
Cannabis consumption is associated with altered steroid metabolism in young men
Abstract
Background:
Cannabis use has been hypothesized to alter endocrine function. We aimed at investigating this hypothesis through extended steroid profiling in young men.
Methods:
Using liquid chromatography – tandem mass spectrometry (LC-MS/MS), 70 endogenous steroids were reliably identified in serum samples from 47 cannabis consumers and 47 controls. Seven major steroids were subject to absolute quantification, while the others were considered as relative concentrations.
Results:
This cross-sectional study shows a global increase in androgen levels among cannabis consumers. Androstenedione (A4), testosterone (T), and dihydrotestosterone (DHT) are among the most significantly increased steroids. In contrast, C11-oxy androgens show no significant change upon cannabis use. This pattern suggests that phytocannabinoids might selectively affect gonadal androgen synthesis without altering adrenal or peripheral pathways, possibly via direct effects on the testes, or disruption of the hypothalamic–pituitary–gonadal (HPG) axis function. Additionally, two progesterone metabolites, 11β-hydroxyprogesterone (11β-OHP4) and 5β-dihydroprogesterone (5β-DHP4), are markedly elevated in cannabis consumers. When the cannabis user group is stratified according to the corresponding usage biomarkers, 11β-OHP4 proves to be a biomarker of general exposure, whereas 5β-DHP4 displays a dose-dependent relationship.
Conclusions:
These findings highlight the value of extended steroid profiling for investigating hormonal variations and evidence a possible link between cannabis consumption and altered male endocrine function.
Article type: Research Article
Keywords: Endocrine system and metabolic diseases, Metabolomics, Steroid hormones
Affiliations: https://ror.org/01swzsf04grid.8591.50000 0001 2175 2154School of Pharmaceutical Sciences, University of Geneva, Geneva, Switzerland; https://ror.org/01swzsf04grid.8591.50000 0001 2175 2154Institute of Pharmaceutical Sciences of Western Switzerland, University of Geneva, Geneva, Switzerland; https://ror.org/03wma5x570000 0004 0373 8123Swiss Centre for Applied Human Toxicology (SCAHT), Basel, Switzerland; Service of Clinical Chemistry and Toxicology, Central Institute of Hospitals, Hospital of Valais, Sion, Switzerland; https://ror.org/01swzsf04grid.8591.50000 0001 2175 2154Department of Internal Medicine, Faculty of Medicine, University of Geneva, Geneva, Switzerland; https://ror.org/01swzsf04grid.8591.50000 0001 2175 2154Department of Genetic Medicine and Development, University of Geneva, Geneva, Switzerland
License: © The Author(s) 2026 CC BY 4.0 Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
Article links: DOI: 10.1038/s43856-026-01469-x | PubMed: 41991730 | PMC: PMC13087242
Relevance: Moderate: mentioned 3+ times in text
Full text: PDF (627 KB)
Introduction
According to the World Health Organization, 2.5% of the world population (i.e., about 150 million people) would consume cannabis (also called marijuana)1. In parallel with increasing rates of recreational use, cannabis is now widely legalized or authorized for medical purposes, making the assessment of its potential adverse effects a growing public health priority. The impact of cannabis on the male reproductive system has been particularly documented but remains controversial2–5. Several studies reported altered semen parameters, such as a lower sperm count, concentration, motility, and viability in semen samples from men exposed to phytocannabinoids2,4,6. These adverse effects are thought to be mediated by the endocannabinoid system (ECS), which comprises lipid-derived neurotransmitters (endocannabinoids) and their receptors (CB1 and CB2), which are expressed throughout the hypothalamic–pituitary–gonadal (HPG) axis5,7. Δ9-tetrahydrocannabinol (THC), the primary psychoactive compound in cannabis, can bind to these two receptors4,5. This interference of THC with the endocannabinoids in the ECS could affect the homeostasis of the HPG axis, which is essential for male reproductive function. Specifically, it could impair the regulation of key reproductive hormones in the hypothalamus and anterior pituitary, including the gonadotropin-releasing hormone (GnRH), the luteinizing hormone (LH), the follicle-stimulating hormone (FSH), and the sex hormones, such as androgens and estrogens4–6,8,9. As CB1 and CB2 receptors are also present in the testes, specifically in Leydig cells, phytocannabinoids may also directly alter testicular steroidogenesis5.
The current state of the art on the hormonal effect of cannabis consumption in males, especially its impact on testosterone (T) levels, has frequently been described as “inconsistent”, “contradictory”, or “conflicting”6,10,11. The only article reporting a significant decrease in circulating T levels in men consuming cannabis dates back to 1974, involved a small cohort (20 consumers compared to 20 controls), and lacked control for key confounding factors12. In contrast, numerous subsequent studies have found no significant differences in blood testosterone concentrations between cannabis users compared to non-users2,10,13–16. However, these studies were either limited by small sample sizes13, or focused on subfertile men in clinical settings, which are not representative of the general population14–16.
More robust cross-sectional studies conducted in recent years in Denmark and the United States have evaluated thousands of young men from the general population and consistently reported higher serum T levels in cannabis users compared to non-users11,17–19. These findings, drawn from large, well-characterized cohorts, suggest a positive association between cannabis use and testosterone, which could be more pronounced either with frequency of use17 or with recency of regular use11. Despite these advances, existing studies are limited by several factors: reliance on self-reported cannabis use without biomarker confirmation, difficulty accounting for all potential confounders, and a narrow focus on testosterone without comprehensive hormonal profiling.
In Switzerland, the mandatory military enrollment for men between 18 and 22 years offers the possibility to recruit participants from the general population and evaluate their reproductive health20. Based on this vast recruitment campaign, sub-cohorts were created to investigate potential associations between male reproductive health outcomes and specific environmental factors. In particular, the gonadotropin axis function was studied in hundreds of participants and associated with their cannabis consumption status, which was confirmed by analyses of phytocannabinoid levels in biological fluids. This work concluded in increased concentrations of endocannabinoids, androgens, estradiol, and sex hormone binding globulin in cannabis smokers, with higher significance in chronic and recent consumers21.
Gaining further insights into steroid metabolism in young men of reproductive age is essential to assess the potential hormonal imbalances associated with cannabis use. Most of the current state of the art is restricted to serum testosterone, with few exceptions, such as a recent Swiss study that also monitored androstenedione (A4), cortisol (F), and dehydroepiandrosterone sulfate (DHEAS)21.
To address this limitation, the present investigation applies an extended steroid profiling approach to a subset of the Swiss cohorts described previously20,21, comprising 47 cannabis users and 47 matched controls. This extended profiling performed with liquid chromatography (LC) hyphenated to tandem mass spectrometry (MS/MS) covers 171 target steroids from diverse subclasses, including androgens, progestogens, estrogens, corticosteroids, bile acids, oxysterols, and phase II metabolites (glucuronides and sulfates). Among the 70 reliably detected endogenous steroids, alterations in metabolic patterns related to testicular steroidogenesis are associated with cannabis consumption, providing insight into its systemic endocrine effects.
Methods
Sample selection
Participants were recruited nationwide in Switzerland from 2005 to 2017 as previously described20. The ethics committees of the cantons of Vaud (17-01-2005, 01/02), Zürich (EK-StV-Nr. 27-2006), Ticino (Rif.CE 1886), and Geneva (2016-01674) approved the present study. All participants signed a written informed consent. Serum samples were retrieved from the study of Zufferey et al.21. Zufferey et al. quantified cannabis biomarkers (THC and THC-COOH) in the same serum samples that were studied in the present work (see Section S10)21. Two groups were constructed to differentiate 47 participants with confirmed cannabis consumption (i.e., declared consumption and positive concentrations of THC and THC-COOH detected in serum) from 47 participants with no detectable THC and THC-COOH in serum and who did not declare any cannabis use (categorized as controls). All participants were aged 18–23 at the time of sampling. Most samples were taken in late afternoon, and there was no significant difference in sampling time of day between THC-positive and control groups, considerably reducing the influence of diurnal variations as a confounder. BMI, which might also be a confounding factor, was available for each participant. The group of cannabis consumers was further separated into chronic or occasional consumers according to their circulating THC-COOH level21,22. Participants were considered chronic users when THC-COOH was quantified at more than 40 µg/L in serum. All cannabis users were defined according to their declared substance use within the previous 7 days. Samples from these participants reflect very recent cannabis use (<12 h, based on the model of Huestis et al.23).
Chemicals
LC-MS “Optima” grade solvents, i.e., Acetonitrile (ACN), Methanol (MeOH), and Water (H2O), were purchased from Fisher Scientific. Formic Acid (FA) was acquired from Biosolve Chimie at ULC-MS purity (>99%). Ammonium Fluoride (NH4F) (>99.99% purity) was purchased from Sigma-Aldrich (Merck KGaA).
Analytical standards of endogenous steroids and 13C-labeled internal standards were supplied by Sigma-Aldrich (Merck KGaA), Steraloids Inc., and LGC Standards (LGC Ltd).
Sample preparation
Between collection and extraction, all serum samples were stored at −80 °C. The procedure for the preparation of serum samples for multi-targeted steroid analysis was described in detail elsewhere24. Briefly, 750 µL of protein precipitation solution (ACN / MeOH, 9:1 v/v) containing 13C-labeled internal standards was added to 250 µL serum samples. After centrifugation, supernatants were filtered through HLB Prime 30 mg cartridges (96-well format, Waters Corp.). Extracts resulting from this “reversed solid-phase extraction” were evaporated to dryness and reconstituted in 50 µL Water / Methanol (1:1, v/v). The injection volume for each sample at the LC-MS/MS analysis was 5 µL.
LC-MS/MS analysis
The multi-targeted LC-MS/MS method for extended steroid profiling was previously described in detail24. According to this protocol, seven steroid metabolites were subject to absolute quantification through a one-point internal calibration strategy, thanks to the commercial availability of 13C-labeled standards. The other steroids were analyzed semi-quantitatively using MRM peak areas. This corresponds to a semi-targeted assay according to the classification of Beger et al.25.
The separation of steroids was performed with a Biphenyl stationary phase (Restek Raptor Inert Biphenyl, 2.1 × 100 mm, 1.8 µm) and a H2O/MeOH mobile phase gradient (from 40 to 100 % methanol in 18 min). LC flow rate was 0.4 mL/min. A concentration of 0.01 % of FA was added to the mobile phase. NH4F was added post-column to enhance the ionization of steroids.
Mass spectrometry was achieved with a Xevo TQ-XS Triple Quadrupole equipped with a ZSpray ESI source (Waters Corp.). Multiple Reaction Monitoring (MRM) mode was used for data acquisition, utilizing MS/MS transitions that were previously optimized on neat standards. Both negative polarity and positive polarity transitions could be acquired simultaneously (polarity switching). The selected transitions for the cohort acquisition stemmed from a preliminary analysis of a pooled QC sample from this cohort, which suggested the tentative detection of 94 endogenous steroids out of the 171 target compounds, along with the 14 isotope-labeled internal standards (see Supplementary Data 1).
Data processing
MRM chromatograms were acquired in MassLynx (Version 4.2, Waters Corp.) and processed in Skyline (Version 24.1, “molecule” interface, MacCoss Lab Software) with manual peak verification and integration.
Peak areas of endogenous steroids were normalized by peak areas of spiked 13C-labeled internal standards (SILs) in the same sample. The attribution of a given SIL to a given endogenous compound was made based on the following criteria, by decreasing order of importance: mass spectrometry acquisition polarity, steroid class, and retention time difference. A summary of the SIL/analyte pairs is presented in Supplementary Data 2.
Steroid features were excluded from the dataset if they were absent from at least 50% of the samples from this study. A peak was considered missing in a sample if its peak area was lower than the mean peak area measured in the procedural blanks. Missing values were replaced by one-third of this mean “blank” peak area before multivariate analysis. Compounds were also excluded if the coefficient of variation (CV) of their normalized peak area in 10 pooled QCs exceeded 30%. A summary of quality control parameters for all the steroid compounds is given in Supplementary Data 3.
The determination of absolute concentrations of seven steroids for which SILs were commercially available was performed using a previously described one-point calibration strategy and an automated in-house workflow implemented in Python 3.9.24,26.
Statistics
The design of this clinical study involved two groups of 47 cannabis users and 47 matched controls. Among the cannabis users, 14 were classified as “chronic” while 33 were classified as “occasional” for the corresponding comparisons.
Univariate analyses were conducted with Prism (Version 10.3.1, GraphPad). T-tests were performed with or without Welch correction, depending on the homogeneity of variance in the two groups. Two-tailed p-values were calculated. Welch correction was applied when the variance in the two groups was significantly different (F-test, p < 0.05).
Multivariate analyses, including Principal Component Analysis (PCA), Orthogonal Partial Least Square—Discriminant Analysis (OPLS-DA), and Partial Least Squares regression (PLS), were performed after unit variance scaling using the software SIMCA (Version 17.0.2, Sartorius AG).
Results & Discussion
The concentration of bioactive androgens is significantly higher in cannabis users
Using the previously proposed one-point internal calibration approach24, seven major compounds of the steroid biosynthesis pathway in humans were accurately quantified using their corresponding 13C-labeled standard: androstenedione (A4), testosterone (T), 17α-hydroxyprogesterone (17α-OHP4), progesterone (P4), 11-deoxycortisol (S), cortisol (F), and cortisone (E) (Table 1). The absolute concentrations measured were consistent with established reference ranges for men in their early twenties27, and aligned well with previous findings in a similar Swiss cohort21.
Table 1: Absolute concentrations (in nmol/L) of seven major steroids in serum samples from cannabis users and non-users. P-values were obtained from two-tailed t-tests without multiple comparison adjustment
| THC-positive(n = 47) | Controls(n = 47) | Difference between groups (THC + – THC-) | ||||||
|---|---|---|---|---|---|---|---|---|
| Min | Max | Mean | Min | Max | Mean | Difference | p-value | |
| A4 | 1.64 | 7.19 | 3.70 | 1.12 | 5.96 | 2.96 | +0.75 | 0.008 (**) |
| T | 7.2 | 32.2 | 19.3 | 6.6 | 28.9 | 15.7 | +3.5 | 0.002 (**) |
| 17α-OHP4 | 0.46 | 3.93 | 1.96 | 0.42 | 3.77 | 1.62 | +0.34 | 0.03 (*) |
| P4 | 0.169 | 0.594 | 0.297 | 0.143 | 0.513 | 0.272 | +0.025 | 0.19 (ns) |
| S | 0.09 | 3.45 | 1.07 | 0.16 | 3.06 | 1.01 | +0.06 | 0.70 (ns) |
| F | 56 | 366 | 235 | 60 | 413 | 241 | −6 | 0.72 (ns) |
| E | 14.7 | 58.4 | 33.0 | 11.4 | 51.1 | 33.5 | −0.5 | 0.79 (ns) |
Notably, serum levels of A4 and T, the two quantified androgens, were significantly higher in THC-positive individuals (p = 0.008 and p = 0.002, respectively). A modest but statistically significant increase was also observed for 17α-OHP4 (p = 0.03). In contrast, no significant differences were observed for P4, S, F, and E.
This finding supports the conclusion drawn by Gundersen et al. in a large cross-sectional cohort from Denmark, which evidenced a 7% higher T concentration in marijuana users after adjustment for confounders17. The same observation was reported in another large cross-sectional study led in the U.S., where T was measured with higher concentrations in THC-positive participants, regardless of the frequency and recency of use19.
Although displaying a smaller androgenic activity than T, A4 is the major precursor of T and is thus essential in masculine sex hormone metabolism28. Its significantly higher concentration in cannabis consumers further supports previous findings of Zufferey et al.21.
Absolute quantitative data on these seven major steroids were complemented by extended steroid profiling. Out of the 171 target steroids (Supplementary Data 1), 77 were consistently identified in serum, with 70 meeting stringent quality control criteria (see Supplementary Fig. S1, Supplementary Data 3). These encompassed a broad range of steroid subclasses: 17 androgens, 15 progestogens, 3 estrogens, 20 corticosteroids, 3 oxysterols, 9 glucuronides, 5 sulfates, and 5 bile acids (see Supplementary Fig. S2). This depth of profiling exceeds that of most previous targeted or untargeted steroidomic studies in human serum29–32, and closely mirrors the steroidome found in certified human blood reference materials24. When no absolute concentration was determined, data analysis of the extended steroid profile was achieved using analyte peak areas normalized by peak areas of 13C-labeled internal standards (ISTDs) as representative of concentration.
Within the extended steroid profile, 5α-dihydrotestosterone (DHT) was of particular interest given the previous observation on T and A4. DHT is the most potent androgen in humans, even more than T, as it features a twice higher ability to bind to the androgen receptor, and a five-fold lower dissociation rate33. The present study demonstrates that serum DHT is significantly higher in THC-positive men (p = 0.029). This confirms the interest of measuring DHT and other androgens in addition to T to support hypotheses regarding steroidogenesis and androgen activity in men. As summarized in Fig. 1, the levels of all three bioactive androgens of gonadal origin were significantly higher among cannabis users.
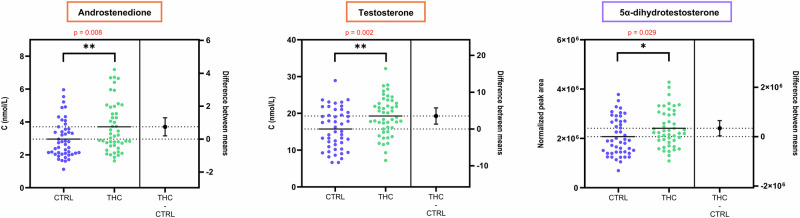
Adrenal androgen synthesis is not affected by cannabis consumption
Further insights into the complex male steroidome in the context of cannabis usage were obtained using multivariate analysis. An Orthogonal Partial Least Squares—Discriminant Analysis (OPLS-DA) model was used to discriminate THC-positive (n = 47) and THC-negative (n = 47) participants based on their circulating steroid profile (M1, see Section S2 of the Supplement). A 7-fold cross-validation of the optimal model with one predictive and one orthogonal component resulted in the following metrics: R²Xp = 0.055, R²Y = 0.49, Q² = 0.19. Although the low value of Q² indicates a low predictive ability, all Q² values obtained under the null hypothesis in a permutation test with 100 permutations were markedly lower than the observed Q², meaning that the OPLS-DA model can be considered statistically significant (see Supplementary Fig. S6)34.
Consistent with absolute quantification, the OPLS-DA model confirmed higher levels of T, A4, and DHT in THC-positive individuals (see Fig. 2). Variable Importance in Projection (VIP) scores further highlighted these androgens as key contributors to the group separation, with T and DHT ranking second and fourth among all variables (see Supplementary Fig. S5).
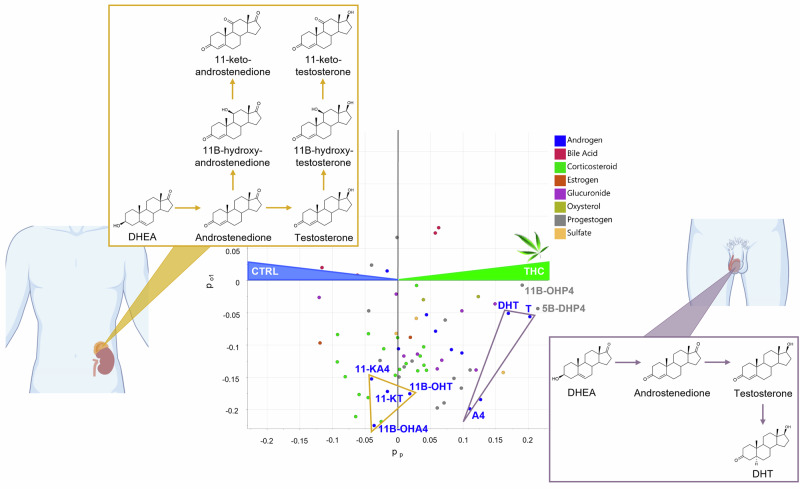
However, not all androgens contributed equally to the discrimination between cannabis users and non-users. Dehydroepiandrosterone (DHEA), a precursor of androgen synthesis, did not contribute to group separation in the model. As, in contrast, 17α-OHP4 is significantly increased in cannabis users (see Table 1), this suggests that the increase of gonadal androgens (A4, T, and DHT) may be mediated by the Δ4 pathway (involving 17α-OHP4 as precursor of A4) instead of the Δ5 pathway (production of A4 via DHEA).
All C11-oxy androgens (11β-hydroxytestosterone (11β-OHT), 11β-hydroxyandrostenedione (11β-OHA4), 11-ketotestosterone (11-KT), and 11-ketoandrostenedione (11-KA4)) also displayed similar levels in both groups according to M1 loadings (see Fig. 2). Notably, 11β-hydroxydihydrotestosterone (11β-OHDHT) was not among the target compounds, and 11-ketodihydrotestosterone (11-KDHT) was targeted but not detected. These findings were confirmed by univariate analyses, which showed no significant differences in C11-oxy androgen levels between users and non-users (p > 0.5, see Supplementary Fig. S7).
C11-oxy androgens are synthesized almost exclusively in the adrenal cortex via the enzymatic activity of the 11β-monooxygenase CYP11B1 and the 11β-dehydrogenase HSD11B2, which act on the precursor androgen substrates35–38. As such, their circulating concentrations predominantly reflect adrenal androgenic output39, in contrast to A4, T, and DHT levels for which testicular synthesis largely prevails in men33.
It can thus be concluded that adrenal and peripheral androgen biosynthesis in men is not affected by cannabis usage. It is particularly relevant to pinpoint that the highly potent 11-KT, which has similar bioactivity to DHT36, is not higher in cannabis users, and therefore does not affect androgenic activity compared to non-users.
Two metabolites of progesterone are strongly related to cannabis consumption
In the OPLS-DA model M1, the highest and third-highest VIP values were attributed to two progestogens, namely 5β-dihydroprogesterone (5β-DHP4) and 11β-hydroxyprogesterone (11β-OHP4), which were more concentrated in serum from cannabis consumers (see Fig. 2 and Supplementary Fig. S5). Multivariate trends were confirmed by t-tests that revealed a strong relationship between these two steroid compounds and cannabis usage. The high statistical significance of this finding was characterized by a p-value of 1.9 ∙ 10−5 for 5β-DHP4 and 5.7 ∙ 10−5 for 11β-OHP4 (see Fig. 3A).
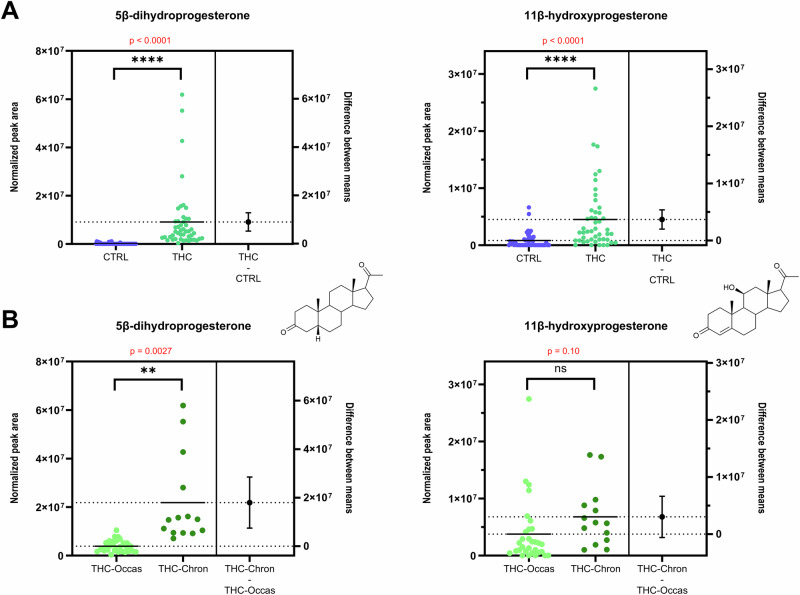
5β-DHP4 and 11β-OHP4 are downstream metabolites of progesterone. 5β-DHP4 is formed through the 5β-reductase pathway. Despite its clear association with cannabis use, the functional role of this metabolite in male reproductive biology remains unknown, warranting further biochemical investigation. However, its presence among testicular steroids was recently suggested in a murine gonadal cell model (MA-10), although not delineated from its 5α-counterpart, 5α-dihydroprogesterone40.
11β-OHP4 is synthesized via the steroid 11β-monooxygenase CYP11B1 and the aldosterone synthase CYP11B241. Interestingly, other CYP11B-derived metabolites, such as 11β-hydroxyandrostenedione (11β-OHA4), did not follow the same trend, suggesting that the increase in 11β-OHP4 is metabolite-specific and not due to generalized upregulation of CYP11B enzymes. In previous reports, 11β-OHP4 was described as a potential precursor of the backdoor pathway and the biosynthesis of C11-oxy steroids41. In the present study, downstream metabolites of 11β-OHP4 in this backdoor pathway, such as 11-ketoprogesterone (11-KP4) and 11-ketodihydrotestosterone (11-KDHT), were targeted, but they could not be detected. This precludes further conclusions on the metabolic fate of 11β-OHP4 in the context of cannabis use. However, similarly to 5β-DHP4, this molecule was evidenced in a Leydig cell mouse model, suggesting that it may be directly produced in the testes40, even though the expression of CYP11B1 in human testes remains uncertain39,42.
Phytocannabinoid levels reveal dose-dependent associations with steroidome changes
A deeper investigation of the relations between androgen levels and THC exposure was performed using Partial Least Squares (PLS) multivariate regression for the THC-positive group, thanks to additional data on serum concentrations of THC and its primary metabolite THC-COOH21. THC and THC-COOH concentrations were considered as independent variables, and the 70 variables of the extended steroid profile dataset were used as X-independent variables (THC-model M2 in Supplementary Section S4; THC-COOH model M3 in Supplementary Section S5). Both models displayed similar relationships between cannabis markers and steroid compounds. In particular, the three main gonadal androgens (A4, DHT, and T) systematically increased when THC and THC-COOH levels were higher, as their coefficients in M2 and M3 were all positive (see Fig. S10 and Fig. S15 in the Supplement). On the other hand, the C11-oxy derivatives were either not significantly related to THC and THC-COOH levels (VIP < 1, see Supplementary Figs. S11 and S16) or were negatively correlated with THC and THC-COOH when their contribution to models M2 and M3 was significant (see Supplementary Figs. S10 and S15).
These results highlight that within cannabis users, increased exposure to phytocannabinoids, characterized by higher levels of THC and THC-COOH, is associated with higher levels of testicular androgens, but not with higher levels of C11-oxy androgens (Supplementary Fig. S12). This reinforces the hypothesis of a specific alteration of gonadal—and not adrenal—steroidogenesis upon cannabis exposure.
Cannabis consumers were further stratified into “chronic” (n = 33) or “occasional” (n = 14) users, based on their blood THC-COOH concentration, a validated biomarker of exposure, using thresholds established by Fabritius et al.21,22.
In contrast to A4, no significant difference in serum T levels was found between chronic and occasional cannabis consumers (p = 0.39, Supplementary Data 4, Supplementary Fig. S17). This is consistent with findings of Fantus et al., who reported a non-linear, “inverse U” relationship between frequency of cannabis use and T levels19, as well as Thistle et al., who stated that the recency of use, rather than frequency, had a greater influence on circulating testosterone concentrations11. In the present study, all “THC-positive” participants declared cannabis use in the last 7 days before sampling, and showed detectable levels of THC in blood. They were thus all recent users and could not be classified more finely according to this criterion. Similarly, no extrapolation to long-term effects on past users can be drawn from the present dataset.
An OPLS-DA model was generated to differentiate chronic and occasional users based on their circulating steroid profile (Model M4, Supplementary Section S7). Interestingly, 5β-DHP4 emerged again as the primary contributor of the discrimination between occasional and chronic consumers (see Supplementary Fig. S19). On the other hand, 11β-OHP4 was not a major contributor to the predictive component of this model (VIP < 1, see Supplementary Fig. S20). Univariate analyses confirmed this trend (see Fig. 3B). The corresponding t-tests showed non-significant differences of 11β-OHP4 levels between chronic and occasional users (p = 0.10), supporting its role as a general exposure marker, which does not vary significantly depending on the acuteness of cannabis consumption.
In contrast, 5β-DHP4 levels were significantly higher in chronic users (p = 0.0027, see Fig. 3B). Therefore, 5β-DHP4 may serve as a dual biomarker, reflecting both cannabis exposure (Fig. 3A) and the intensity or chronicity of use (Fig. 3B). This is further supported by its positive correlation with circulating THC and THC-COOH levels (see Supplementary Sections S4 and S5).
Steroidomics provides further insight into the uncertain mechanisms of male hormonal response to phytocannabinoid intake
While previous studies have explored associations between cannabis consumption, semen quality parameters, and serum hormone levels16,17,21, this is the first to apply comprehensive steroid profiling to healthy young men in relation to their cannabis consumption. A major advantage of our cohort is its narrow age range (18–23 years), which limits age-related hormonal variability seen in other studies13,16. Other known confounding factors, such as time of day at sampling, Body Mass Index (BMI), or tobacco smoking17,43–46, were also carefully investigated and found non-significant in this work (see Section S8 of the Supplement, including Figs. S22 and S23). However, the influence of other confounders originating from the participants’ lifestyle, including diet, alcohol consumption, sleep patterns, stress, etc., cannot be ruled out, as some of these parameters could not be assessed. Furthermore, the hereby-presented results reflect short-term effects on recent cannabis users for a restricted group of young Swiss men. At this stage, they should not be generalized to women, other age groups, or more diverse populations. The cross-sectional nature of this study and the wide array of factors influencing steroid metabolism result in an inherent risk of false positives. Every possible precaution was taken to minimize this risk, and follow-up work will be conducted to confirm the outcome of the present study.
The main finding of this study is the consistent increase of T in cannabis consumers, a trend now corroborated by multiple large-scale cross-sectional studies from both Europe and North America, pointing to higher levels of androgens in cannabis users17,19,21. This growing body of evidence challenges earlier reports suggesting testosterone suppression12,47.
Despite these consistent findings, the underlying mechanisms remain unclear. By combining extended steroid profiling, cannabis biomarker quantification, and broader hormonal analysis, this study enables the exploration of potential pathways responsible for this increase in T.
Fantus et al. proposed several hypotheses to explain elevated T in cannabis users: modulation of LH levels, direct effects on testicular receptors, central suppression, hypothalamic modulation, or interactions at multiple levels. Another possibility is reverse causality, that men with inherently higher testosterone could be more prone to cannabis use due to increased risk-taking behavior19.
It may indeed be argued that cannabis usage is more likely in men with naturally higher levels of T, as this physiological characteristic could lead to increased risk-taking21,48,49. However, the specific relationship between baseline T levels and cigarette smoking or drug abuse was previously described as “significant, but modest”48, and the relationship between T and risk-taking in general might be overstated due to underestimation of social status as a confounding factor50.
A more compelling hypothesis is that phytocannabinoids directly disrupt the homeostasis of the HPG axis due to the presence of cannabinoid receptors along this axis. However, in the present study, no relation could be established between LH and/or FSH levels and sex steroid levels (see Supplementary Section S9). Moreover, LH and FSH concentrations did not significantly differ between THC-positive and THC-negative participants (see Supplementary Fig. S24). The complexity of feedback mechanisms, whereby gonadal steroids modulate pituitary hormone secretion, along with the concurrent elevation of LH and gonadal androgens in users, makes it difficult to attribute increased testosterone specifically to LH modulation38. An additional limitation of this analysis is the pulsatile secretion of LH and FSH, which reduces the comparability of single-time-point measurements. Likewise, hypothalamic modulation via GnRH cannot be directly evaluated, as GnRH is secreted in pulses and does not circulate in the bloodstream. With no measurable intermediates beyond LH and FSH, evidence for a direct hypothalamic effect of phytocannabinoids remains inconclusive. Thus, while a phytocannabinoid-induced disruption of the HPG axis is a plausible explanation, the precise mechanisms remain unclear. Furthermore, the increase of T may also be seen as a homeostatic response to compensate for a decreased sensitivity of the androgen receptor in the presence of phytocannabinoids.
The present work demonstrates that cannabis-related alterations of androgen metabolism are confined to the gonadal part of the HPG axis. This is evidenced by elevated concentrations of the two other major gonadal androgens, A4 and DHT, while adrenal-derived androgens, such as C11-oxy androgens, remain unaffected. This finding rather supports a possible direct effect of phytocannabinoids on the testicular sex hormone synthesis, mediated by CB1 receptors located in Leydig cells.
Another possible source of steroid metabolism alteration that cannot be ruled out in this study is the effect of phytocannabinoids on liver metabolism. It is known that CB1 and CB2 receptors are expressed in the liver4,51. Among the steroids that show increased concentrations upon cannabis exposure according to our multivariate models, several are hydroxylated or reduced metabolites of A4, T, and P4 (see Fig. 2 and Supplementary Figs. S10 and S15), which may indicate increased activity of hepatic cytochrome P450 (CYP) enzymes. However, the literature on the effects of cannabis use on CYP enzymes is conflicting. While both possibilities were reported, there is slightly more in vitro evidence to suggest that THC and CBD inhibit CYP enzymes than induce them, and there is no conclusive evidence from in vivo studies52. Therefore, liver metabolism should be considered as a potential factor in cannabis-related alterations of steroid metabolism, but its role in this study remains uncertain and relatively minor compared to testicular steroidogenesis.
Two compounds from the extended steroid profile, 5β-DHP4 and 11β-OHP4, have been highlighted in this work as potential biomarkers of phytocannabinoid intake in men. While 5β-DHP4 might serve both as a biomarker of exposure and a biomarker of intensity, 11β-OHP4 would better characterize a generic exposure to phytocannabinoids with less significant dose-dependency. It is especially interesting to observe alterations of progesterone metabolism in cannabis users, as progesterone plays a key role in reproductive processes such as LH receptor expression, intracellular signaling in sperm, chemotaxis, and acrosome reaction53–56.
Although this study sheds light on the hormonal effects of cannabis use, its implications for male reproductive health remain uncertain. Data on semen quality in cannabis users from the general population are limited, and findings so far are inconclusive. Recent studies either report no significant differences in semen parameters between users and non-users21, or point to a reduced sperm concentration and total sperm count17. Given the consistent observation of elevated gonadal androgen levels in serum from cannabis users, it is now essential to further investigate how this hormonal profile relates to semen quality in the context of cannabis exposure. There is also a need for further development of relevant in vitro models for the toxicological evaluation of endocrine perturbations in the HPG axis (e.g., human Leydig cells), while current OECD Guidelines for the Testing of Chemicals rely on the adreno-carcinoma H295R cell line to assay steroidogenesis57.
Supplementary Materials
- Transparent Peer Review file (PDF)
- Supplementary Information (PDF)
- Description of Additional Supplementary files (DOCX)
- Supplementary Data 1 (XLSX)
- Supplementary Data 2 (XLSX)
- Supplementary Data 3 (XLSX)
- Supplementary Data 4 (XLSX)
- Supplementary Data 5 (XLSX)
- Supplementary Data 6 (XLSX)
- Supplementary Data 7 (XLSX)
References
- 1.World Health Organization. Cannabis. Alcohol, Drugs and Addictive Behaviourshttps://www.who.int/teams/mental-health-and-substance-use/alcohol-drugs-and-addictive-behaviours/drugs-psychoactive/cannabis (2025).
- KS Payne, DJ Mazur, JM Hotaling, AW Pastuszak. Cannabis and male fertility: a systematic review. J. Urol., 2019. [DOI | PubMed]
- 3.Srinivasan, M. et al. The effect of marijuana on the incidence and evolution of male infertility: a systematic review. Cureus10.7759/cureus.20119 (2021).
- BM Fonseca, I Rebelo. Cannabis and cannabinoids in reproduction and fertility: where we stand. Reprod. Sci., 2022. [DOI | PubMed]
- J Lim, E Squire, K-M Jung. Phytocannabinoids, the endocannabinoid system and male reproduction. World J. Mens. Health, 2023. [DOI | PubMed]
- SS Du Plessis, A Agarwal, A Syriac. Marijuana, phytocannabinoids, the endocannabinoid system, and male fertility. J. Assist. Reprod. Genet., 2015. [DOI | PubMed]
- S González. Identification of endocannabinoids and cannabinoid CB1 receptor mRNA in the pituitary gland. Neuroendocrinology, 1999. [DOI | PubMed]
- P Hsiao, RI Clavijo. Adverse effects of cannabis on male reproduction. Eur. Urol. Focus, 2018. [DOI | PubMed]
- 9.Dwyer, A. A. & Quinton, R. Anatomy and Physiology of the Hypothalamic-Pituitary-Gonadal (HPG) Axis. in Advanced Practice in Endocrinology Nursing-Part VII: Male Endocrinology and Reproduction 839–852 (Springer, 2019).
- F Belladelli. The association between cannabis use and testicular function in men: a systematic review and meta-analysis. Andrology, 2021. [DOI | PubMed]
- JE Thistle. Marijuana use and serum testosterone concentrations among U.S. males. Andrology, 2017. [DOI | PubMed]
- RC Kolodny, WH Masters, RM Kolodner, G Toro. Depression of plasma testosterone levels after chronic intensive marihuana use. N. Engl. J. Med., 1974. [DOI | PubMed]
- JK Lisano. Performance and health-related characteristics of physically active males using marijuana. J. Strength Cond. Res., 2019. [DOI | PubMed]
- FL Nassan. Marijuana smoking and markers of testicular function among men from a fertility centre. Hum. Reprod., 2019. [DOI | PubMed]
- TA Teixeira. Marijuana is associated with a hormonal imbalance among several habits related to male infertility: a retrospective study. Front. Reprod. Health, 2022. [DOI | PubMed]
- N Khan. The effects of tobacco and cannabis use on semen and endocrine parameters in infertile males. Hum. Fertil., 2023. [DOI]
- TD Gundersen. Association between use of marijuana and male reproductive hormones and semen quality: a study among 1,215 healthy young men. Am. J. Epidemiol., 2015. [DOI | PubMed]
- ML Eisenberg. Invited commentary: the association between marijuana use and male reproductive health. Am. J. Epidemiol., 2015. [DOI | PubMed]
- RJ Fantus, SD Lokeshwar, TP Kohn, R Ramasamy. The effect of tetrahydrocannabinol on testosterone among men in the United States: results from the National Health and Nutrition Examination Survey. World J. Urol., 2020. [DOI | PubMed]
- R Rahban. Semen quality of young men in Switzerland: a nationwide cross-sectional population-based study. Andrology, 2019. [DOI | PubMed]
- F Zufferey. Gonadotropin axis and semen quality in young Swiss men after cannabis consumption: effect of chronicity and modulation by cannabidiol. Andrology, 2024. [DOI | PubMed]
- M Fabritius, M Augsburger, H Chtioui, B Favrat, C Giroud. Fitness to drive and cannabis: validation of two blood THCCOOH thresholds to distinguish occasional users from heavy smokers. Forensic Sci. Int., 2014. [DOI | PubMed]
- MA Huestis, JE Henningfield, EJ Cone. Blood cannabinoids. II. Models for the prediction of time of marijuana exposure from plasma concentrations of Δ9-tetrahydrocannabinol (THC) and 11-nor-9-carboxy-Δ9-tetrahydrocannabinol (THCCOOH). J. Anal. Toxicol., 1992. [DOI | PubMed]
- M Galmiche. Extended steroid profiling in human serum and plasma with simultaneous quantitative determination using one-point internal calibration. J. Sep. Sci., 2025. [DOI | PubMed]
- RD Beger. Analysis types and quantification methods applied in UHPLC-MS metabolomics research: a tutorial. Metabolomics, 2024. [DOI | PubMed]
- G Visconti. Multitargeted internal calibration for the quantification of chronic kidney disease-related endogenous metabolites using liquid chromatography–mass spectrometry. Anal. Chem., 2023. [DOI | PubMed]
- H Frederiksen. Sex- and age-specific reference intervals of 16 steroid metabolites quantified simultaneously by LC-MS/MS in sera from 2458 healthy subjects aged 0 to 77 years. Clin. Chim. Acta, 2024. [DOI | PubMed]
- A Malaviya, J Gomes. Androstenedione production by biotransformation of phytosterols. Bioresour. Technol., 2008. [DOI | PubMed]
- H Elmongy, M Masquelier, M Ericsson. Development and validation of a UHPLC-HRMS method for the simultaneous determination of the endogenous anabolic androgenic steroids in human serum. J. Chromatogr. A, 2020. [DOI | PubMed]
- O Salamin. Development and validation of a UHPLC–MS/MS method for extended serum steroid profiling in female populations. Bioanalysis, 2020. [DOI | PubMed]
- T Andrieu, T Du Toit, B Vogt, MD Mueller, M Groessl. Parallel targeted and non-targeted quantitative analysis of steroids in human serum and peritoneal fluid by liquid chromatography high-resolution mass spectrometry. Anal. Bioanal. Chem., 2022. [DOI | PubMed]
- L Schiffer. Multi-steroid profiling by UHPLC-MS/MS with post-column infusion of ammonium fluoride. J. Chromatogr. B, 2022. [DOI]
- PM Marchetti, JH Barth. Clinical biochemistry of dihydrotestosterone. Ann. Clin. Biochem. Int. J. Lab. Med., 2013. [DOI]
- Y Xu, R Goodacre. Mind your Ps and Qs – Caveats in metabolomics data analysis. TrAC Trends Anal. Chem., 2025. [DOI]
- L Bloem, K-H Storbeck, L Schloms, A Swart. 11β-hydroxyandrostenedione returns to the steroid arena: biosynthesis, metabolism and function. Molecules, 2013. [DOI | PubMed]
- T Du Toit, MJJ Finken, HM Hamer, AC Heijboer, AC Swart. C11-oxy C19 and C11-oxy C21 steroids in neonates: UPC2-MS/MS quantification of plasma 11β-hydroxyandrostenedione, 11-ketotestosterone and 11-ketoprogesterone. Steroids, 2018. [DOI | PubMed]
- SM Glass. Characterization of human adrenal cytochrome P450 11B2 products of progesterone and androstenedione oxidation. J. Steroid Biochem. Mol. Biol., 2021. [DOI | PubMed]
- 38.Du Toit, T., Naamneh Elzenaty, R. & Flück, C. E. Steroid hormone synthesis. in Reference Module in Biomedical Sciences (Elsevier, 2025).
- S Charoensri. Human gonads do not contribute to the circulating pool of 11-oxygenated androgens. J. Clin. Endocrinol. Metab., 2025. [DOI | PubMed]
- T Du Toit, M Groessl, E Pignatti, AC Swart, CE Flück. Characterization of steroid metabolic pathways in established human and mouse cell models. Int. J. Mol. Sci., 2025. [DOI | PubMed]
- D Van Rooyen, R Gent, L Barnard, AC Swart. The in vitro metabolism of 11β-hydroxyprogesterone and 11-ketoprogesterone to 11-ketodihydrotestosterone in the backdoor pathway. J. Steroid Biochem. Mol. Biol., 2018. [DOI | PubMed]
- Y Imamichi. 11-ketotestosterone is a major androgen produced in human gonads. J. Clin. Endocrinol. Metab., 2016. [DOI | PubMed]
- K Collomp. Altered diurnal pattern of steroid hormones in relation to various behaviors, external factors and pathologies: A review. Physiol. Behav., 2016. [DOI | PubMed]
- LG Deltourbe. Steroid hormone levels vary with sex, aging, lifestyle, and genetics. Sci. Adv., 2025. [DOI | PubMed]
- MS Shiels. Association of cigarette smoking, alcohol consumption, and physical activity with sex steroid hormone levels in US men. Cancer Causes Control, 2009. [PubMed]
- M Mezzullo. Impact of age, body weight and metabolic risk factors on steroid reference intervals in men. Eur. J. Endocrinol., 2020. [DOI | PubMed]
- CM Fronczak, ED Kim, AB Barqawi. The insults of illicit drug use on male fertility. J. Androl., 2012. [DOI | PubMed]
- A Booth, DR Johnson, DA Granger. Testosterone and Men’s health. J. Behav. Med., 1999. [DOI | PubMed]
- JS Peper, PCMP Koolschijn, EA Crone. Development of risk taking: contributions from adolescent testosterone and the orbito-frontal cortex. J. Cogn. Neurosci., 2013. [DOI | PubMed]
- SR Fisk, BJ Miller, J Overton. Why social status matters for understanding the interrelationships between testosterone, economic risk-taking, and gender. Sociol. Compass, 2017. [DOI]
- I Bazwinsky-Wutschke, A Zipprich, F Dehghani. Endocannabinoid system in hepatic glucose metabolism, fatty liver disease, and cirrhosis. Int. J. Mol. Sci., 2019. [DOI | PubMed]
- SA Smith. Effects of cannabidiol and Δ9-tetrahydrocannabinol on cytochrome P450 enzymes: a systematic review. Drug Metab. Rev., 2024. [DOI | PubMed]
- M Oettel, A Mukhopadhyay. Progesterone: the forgotten hormone in men?. Aging Male, 2004. [DOI | PubMed]
- S Publicover, C Barratt. Progesterone’s gateway into sperm. Nature, 2011. [DOI | PubMed]
- S Matsuyama, T DeFalco. Steroid hormone signaling: multifaceted support of testicular function. Front. Cell Dev. Biol., 2024. [DOI | PubMed]
- 56.Wehrli, L. et al. The major phytocannabinoids, delta-9-tetrahydrocannabinol (THC) and cannabidiol (CBD), affect the function of CatSper calcium channels in human sperm. Hum. Reprod. 40, 796-807 (2025).
- 57.OECD. Test Guideline No. 456 – H295R Steroidogenesis Assay. https://www.oecd.org/content/dam/oecd/en/publications/reports/2023/07/test-no-456-h295r-steroidogenesis-assay_g1g14f4a/9789264122642-en.pdf (2023).