
Chemical constituents of industrial hemp roots and their anti-inflammatory activities
Abstract
Objective:
Although the chemical constituents of the aerial parts of Cannabis have been extensively studied, phytochemicals of Cannabis roots are not well characterized. Herein, we investigated the chemical constituents of industrial hemp (Cannabis sativa L.) roots and evaluated the anti-inflammatory activities of phytochemicals isolated from the hemp roots extract.
Methods:
An ethyl acetate extract of hemp roots was subjected to a combination of chromatographic columns to isolate phytochemicals. The chemical structures of the isolates were elucidated based on spectroscopic analyses (by nuclear magnetic resonance and mass spectrometry). The anti-inflammatory effects of phytochemicals from hemp roots were evaluated in an anti-inflammasome assay using human monocyte THP-1 cells.
Results:
Phytochemical investigation of hemp roots extract led to the identification of 32 structurally diverse compounds including six cannabinoids (1–6), three phytosterols (26–28), four triterpenoids (22–25), five lignans (17–21), and 10 hydroxyl contained compounds (7–16), three fatty acids (29–31), and an unsaturated chain hydrocarbon (32). Compounds 14–21, 23, 27, and 32 were identified from the Cannabis species for the first time. Cannabinoids (1–5) reduced the level of cytokine tumor necrosis-alpha (by 38.2, 58.4, 47.7, 52.2, and 56.1%, respectively) and 2 and 5 also decreased the interleukin-1β production (by 42.2 and 92.4%, respectively) in a cell-based inflammasome model. In addition, non-cannabinoids including 11, 13, 20, 25, 29, and 32 also showed selective inhibition of interleukin-1β production (by 23.7, 22.5, 25.6, 78.0, 24.1, 46.6, and 25.4%, respectively) in THP-1 cells.
Conclusion:
The phytochemical constituent of a hemp roots extract was characterized and compounds from hemp roots exerted promising anti-inflammatory effects.
Supplementary Information:
The online version contains supplementary material available at 10.1186/s42238-022-00168-3.
Article type: Case Report
Keywords: Hemp, Cannabinoids, Inflammasome
Affiliations: grid.12981.330000 0001 2360 039XResearch Center for Drug Discovery, School of Pharmaceutical Sciences, Sun Yat-sen University, 510006 Guangzhou, China; grid.20431.340000 0004 0416 2242Bioactive Botanical Research Laboratory, Department of Biomedical and Pharmaceutical Sciences, College of Pharmacy, University of Rhode Island, 02881 Kingston, RI USA
License: © The Author(s) 2023 CC BY 4.0 Open AccessThis article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
Article links: DOI: 10.1186/s42238-022-00168-3 | PubMed: 36642726 | PMC: PMC9841654
Relevance: Core topic: mentioned in title and keywords
Full text: PDF (1.2 MB)
Introduction
The medicinal uses of Cannabis roots for a variety of maladies are supported by empirical practice and emerging scientific evidence. The ancient Chinese herbology Compendium of Materia Medica (“Bencao Gangmu” in Chinese or Great Pharmacopoeia) was one of the earliest books that documented the use of hemp (Cannabis sativa L.) roots as a folk medicine to soothe the pain. The traditional Chinese medicine system also uses hemp roots as a common practice to treat Reqi (“hot air” or heatiness)-related symptoms (e.g., mouth ulcers) by indigenous people from Southwest China regions. Modern scientific studies provide more rigorous evidence of the biological activities of cannabis roots. Several pre-clinical studies reported that cannabis root extracts exert various pharmacological effects including anti-inflammatory, estrogenic, liver protective, and anti-cancer activities (Elhendawy et al., ref. 2019; Kornpointner et al. ref. 2021; Lima, et al., ref. 2021; Ryz et al., ref. 2017). Notably, there are fewer reported studies on cannabis roots as compared to the plant’s aerial parts including the inflorescences and leaves. This is possibly due to the lower density of glandular trichomes in cannabis roots, which leads to a lower level of cannabinoids biosynthesis (Stout et al., ref. 2012). Moreover, the mechanisms of actions of enzymes that contribute to the biosynthesis of phytochemicals in the roots of cannabis are not fully elucidated (Sirikantaramas and Taura ref. 2017). Nevertheless, several studies on hemp roots have reported the isolation and identification of several phytochemicals including cannabinoid-type of compounds, such as cannabidiol (CBD), tetrahydrocannabinol (THC), cannabichromene, cannabigerol, and cannabigerolic acid, as well as non-cannabinoid-type of compounds including flavonoids, triterpenes, polyphenols, and alkaloids (Stout et al., ref. 2012). In addition, a study characterized the cannabinoids content (ranging between 0.001 and 0.004%) in three chemovars, which was lower than the other chemotypes of compounds such as sterols (0.06–0.09%) and triterpenoids (0.13–0.24%) (Jin et al. ref. 2020). Notably, some unique compounds have been isolated from cannabis roots. For instance, a phenolic amide, namely, N-trans-coumaroyltyramine, was isolated from the roots of a Cannabis sp. grown in Mississippi, USA (Pollastro et al., ref. 2018). In addition, several arylnapthalene-type of compounds including cannabisins B and G were only found in fruits and roots of cannabis (Sakakibara et al., ref. 1995). However, to date, the phytochemical constituents of hemp roots and their biological activities are not been fully investigated. Our group has initiated a program to systematically investigate the biological activities of non-psychedelic cannabinoids (Liu et al., ref. 2021, ref. 2020, ref. 2022; Ma, Li, et al., ref. 2021; Ma, Xu, et al., ref. 2021; Puopolo et al., ref. 2022). During this course, we revealed that the anti-inflammatory activity of CBD is associated with its inhibitory effect on the activation of NLR-family-pyrin-domain-containing 3 (NLR3) inflammasome, which is a vital molecular target for the production of pro-inflammatory cytokine (i.e., interleukin 1 beta; IL-1β) (Liu et al. ref. 2020). However, the phytochemical constituents of hemp roots from the region of Southwest China (i.e., Yunnan Province) and the inhibitory effects of cannabinoids from hemp roots are still not clear. Herein, we report the isolation and identification of phytochemicals of hemp roots as well as the biological evaluation of isolated phytochemicals from hemp roots using a cell-based inflammasome model.
Materials and methods
Chemical reagents
The roots of industrial hemp were provided by Mawang Shenzhen Co. Ltd. (Shenzhen, China) and a voucher sample (No. MG20191230) is deposited in the School of Pharmaceutical Sciences of Sun Yat-Sen University. Lipopolysaccharides (LPS), nigericin, phorbol 12-myristate 13-acetate (PMA), and phosphate-buffered saline were purchased from Sigma-Aldridge Chemical Co (St. Louis, MO, USA).
Extraction and isolation
Dried hemp roots (20 kg) were pulverized and extracted with 95% EtOH (3 × 150 L) for 48 h at room temperature. The solvent was evaporated in vacuo, and the dried ethyl acetate extract (732 g) was suspended in water and partitioned with ethyl acetate 3 times to obtain an ethyl acetate extract (350 g). This extract was chromatographed over a combination of columns (silica gel, Sephadex LH-20, thin-layer chromatography, and semi-preparative HPLC) using various eluting solvents to obtain following purified compounds (see detailed separation and purification in the Supplementary Data): compounds 1 (4 mg, 0.00002%), 2 (31 mg, 0.000155%), 3 (12 mg, 0.00006%), 4 (7 mg, 0.000035%), 5 (9 mg, 0.000045%), 6 (6 mg, 0.00003%), 7 (12 mg, 0.00006%), 8 (90 mg, 0.00045%), 9 (48 mg, 0.00024%), 10 (22 mg, 0.00011%), 11 (125 mg, 0.000625%), 12 (8 mg, 0.00004%), 13 (9 mg, 0.000045%), 14 (11 mg, 0.000055%) 15 (30 mg, 0.00015%), 16 (6 mg, 0.00003%), 17 (43 mg, 0.000215%), 18 (89 mg, 0.000445%), 19 (8 mg, 0.00004%), 20 (67 mg, 0.000335%), 21 (314 mg, 0.00157%), 22 (28 mg, 0.00014%), 23 (17 mg, 0.000085%), 24 (239 mg, 0.001195%), 25 (1.3 g, 0.0065%), 26 (15 mg, 0.000075%), 27 (14 mg, 0.00007%), 28 (18 mg, 0.00009%), 29 (31 mg, 0.000155%), 30 (51 mg, 0.000255%), 31 (25 mg, 0.000125%), and 32 (14 mg, 0.00007%).
Cell culture
Human monocyte THP-1 cells were purchased from the American Type Culture Collection (ATCC, Rockville, MD, USA) and cultured in the Roswell Park Memorial Institute 1640 medium supplemented with 10% fetal bovine serum (Gibco, Life Technologies, Gaithersburg, MD, USA).
Inflammasome inhibition assay
The anti-inflammasome effects of cannabinoids isolated from hemp root were evaluated by measuring the production of lipopolysaccharide (LPS)- and nigericin-induced secretion of pro-inflammatory cytokines including interleukin-1β (IL-1β) and tumor necrosis factor-α (TNF-α) using a previously reported method (Liu et al. ref. 2020). Briefly, THP-1 cells were seeded at a density of 5 × 104 cells per well in a 48-well plate and differentiated with phorbol 12-myristate 13-acetate (PMA) (25 nM) for 48 h. Then, culture medium was removed and replaced with PMA-free medium for 24 h. Next, LPS (100 ng/mL) was added and incubated for 4 h, followed by adding hemp root samples (compounds 1–5; 50 µM) and further incubated for 1 h. Then, nigericin (10 µM) was added and incubated with the cells for 4 h. The cell culture supernatant was collected to measure the levels of IL-1β and TNF-α using the ELISA kits (BioLegend, San Diego, CA, USA).
Statistical analysis
Data are presented as mean ± standard deviation of values from three replicated experiments. Statistical analysis was performed using the GraphPad Prism9 software (GraphPad, La Jolla, CA, USA) with a one-way analysis of variance with multiple comparisons. The significance was noted as p < 0.05 (*), p < 0.01 (**), p < 0.001 (***), and p < 0.0001 (****).
Results and discussion
Isolation and identification of phytochemicals isolated from hemp roots
Thirty-two phytochemicals (1–32) were isolated from the hemp roots extract, and their chemical structures (Fig. 1) were elucidated on the basis of spectroscopic analysis (by NMR and MS) and comparisons of the literature reported values as follows: cannabinol (1) (Wang et al., ref. 2017); (9R,10 S)-6,6,9-trimethyl-3-pentyl-7,8,9,10-tetrahydro-6 H-benzo[c]chromene-1,9,10-triol (2) (Li et al., ref. 2020); (-)-7R-cannabicoumarononic acid A (3) (Radwan et al., ref. 2009); (9R,10 S)-6,6,9-trimethyl-3-propyl-7,8,9,10-tetrahydro-6 H-benzo[c]chromene-1,9,10-triol (4) (Li et al., ref. 2020); cannabivarin (5) (Wang et al., ref. 2017); cannabidiol (6) (Marchetti et al. ref. 2019); 4-hydroxy-3-methoxybenzaldehyde (7) (Ito et al., ref. 2001); 4-hydroxybenzaldehyde (8) (Bouaicha et al., ref. 1994); 4-hydroxy-3,5-dimethoxybenzaldehyde (9) (Xue et al., ref. 2013); syringic acid (10) (Queirós et al., ref. 2020); vanillic acid (11) (Xu et al. ref. 2022); 2,6-dimethoxyphenol (12) (Goda et al., ref. 1987); (2E)-3-(4-hydroxyphenyl)-2-propenoic acid (13) (Seal, ref. 2013); (3 S)-3-hydroxy-β-ionone (14) (DellaGreca et al., ref. 2004); (-)-dihydrovomifoliol (15) (Bennett et al., ref. 1990); 1 H-indole-3-carboxaldehyde (16) (Wang et al., ref. 2012); 3-(4-hydroxyphenyl)-2-propenoic acid ethyl ester (17) (Pizzolatti et al., ref. 2006); 4-hydroxyphenethyl trans-ferulate (18) (Fujita et al., ref. 1995); 1,2-diguaiacyl-1-ethoxy-3-propanol (19) (Mohammed et al., ref. 2022); (+)-4-ketopinoresinol (20) (Chen et al., ref. 2012); dadahol A (21) (Devanathan and Stalin ref. 2020); (+)-oleanolic acid (22) (He et al., ref. 2021), epipseudotaraxastanediol (23) (Hinge et al., ref. 1966); 3-epifriedelinol (24) (Sousa et al., ref. 2012); 9,12-octadecadienoic acid (9Z,12Z)-friedelin (25) (Zhou & Guo, ref. 2009); β-sitosterol (26) (Kadowaki et al., ref. 2003); 5α,8α-epidioxy-(22E,24R)-ergosta-6,22-dien-3β-ol (27) (Gao et al., ref. 2007); stigmast-4-en-3-one (28) (Smania et al., ref. 2003); 9,12-octadecadienoic acid (9Z,12Z)-methyl ester (29) (Huh et al., ref. 2010); α-linolenic acid (30) (Kishino et al., ref. 2003); linoleic acid (31) (Yoon et al., ref. 2015); and dotriacont-1-ene (32) (Chen et al., ref. 2010). The isolated phytochemicals can be divided into two classes, the cannabinoid-type (1–6) and the non-cannabinoid-type (7–32). Among the cannabinoids, compound 1 (cannabinol) is a derivative of tetrahydrocannabinol, which is the major psychedelic (hallucinogenic substance triggering non-ordinary states of consciousness, also referred to as psychoactive or psychotropic) cannabinoid found in cannabis (Sampson, ref. 2021). In contrast to tetrahydrocannabinol, cannabinol only shows mild psychedelic effects (Rhee et al., ref. 1997). Other cannabinoid-type compounds isolated from the hemp roots were non-psychedelic (Fig. 1).
Apart from cannabinoid-type compounds (1–6), several non-cannabinoid compounds were identified from hemp roots. These phytochemicals included three phytosterols (26–28), four triterpenoids (22–25), and five lignans (17–21). Several compounds were isolated and identified from Cannabis sp. for the first time as follows. Compounds 20, 21, 23, and 27 were reported from the Cannabis species for the first time. Additionally, 10 hydroxyl-contained compounds (7–16) were isolated from the hemp roots. Among these, compounds 14–19 were identified from Cannabis for the first time. Moreover, three fatty acids (29–31) and an unsaturated hydrocarbon (32) were identified in Cannabis for the first time.
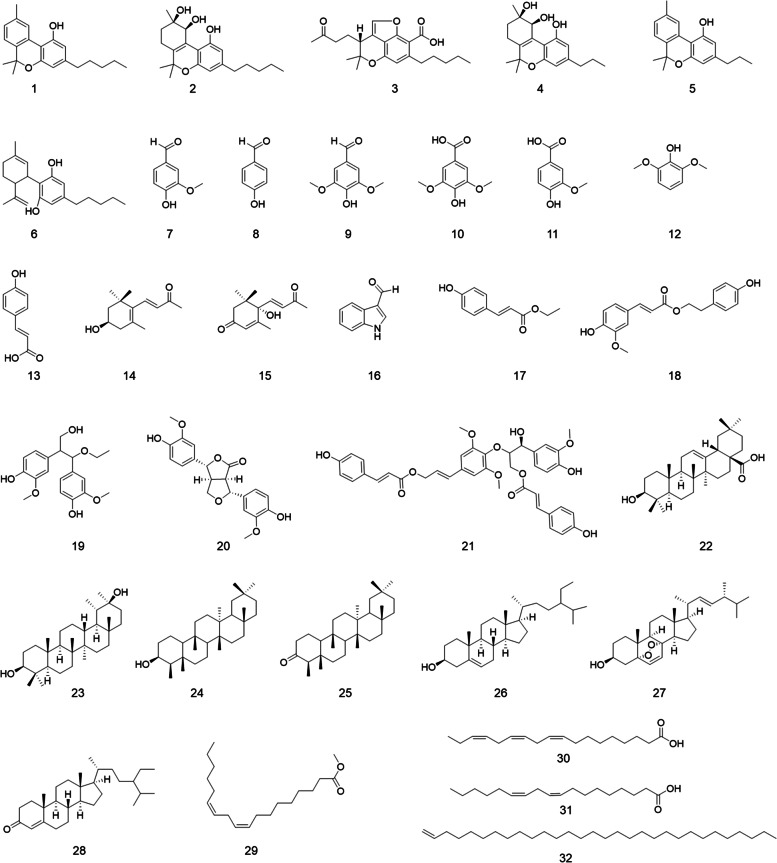
Anti-inflammatory effects of phytochemicals isolated from hemp roots
Our group has previously reported that CBD, the major non-psychedelic cannabinoid in cannabis, confers the anti-inflammatory effect via a specific mechanism, namely, inhibition of inflammasome activation (Liu et al. ref. 2020). However, it is not clear whether other cannabinoids from the hemp roots are inflammasome inhibitors. Thus, an inflammasome inhibition assay was performed to evaluate the anti-inflammatory effects of cannabinoids isolated from hemp roots. The production of IL-1β and TNF-α in inflammasome inducers (LPS and nigericin) stimulated THP-1 cells were significantly increased as compared to the control group (6.1 vs. 1211.6 and 13.0 vs. 102.6 pg/mL, respectively), suggesting that inflammasome in THP-1 cells was activated by the stimulation with LPS and nigericin. Compounds 2 and 5 (50 µM) counteracted LPS-nigericin-induced production of IL-1β by 42.2% and 92.4%, respectively (Fig. 2A). Cannabinoids (1–5) reduced the level of TNF-α by 38.2%, 58.4%, 47.7%, 52.2%, and 56.1%, respectively (Fig. 2B). To confirm whether the anti-inflammatory of these cannabinoids were selective inhibitors of the inflammasome, we used a small molecule with known a selective inflammasome inhibitor effect (Wu et al., ref. 2020), namely, MCC950, as a positive control. MCC950 (10 µM) showed a specific inhibitory effect against the activation of the inflammasome by reducing the level of IL-1β (by 95.8%) without decreasing the level of TNF-α. Additionally, a small molecule with inhibitory effects on the production of TNF-α (DaSilva et al., ref. 2019; Xu et al., ref. 2018), namely, urolithin A, was included as a positive control (as a TNF-α inhibitor) to validate the anti-inflammatory assay. Urolithin A reduced the level of TNF-α by 51.7% as compared to the model group. With these validated bioassays, we were able to evaluate the selective anti-inflammasome activity of the isolates. Although our previously reported study showed that CBD is a specific inflammasome inhibitor (Liu et al. ref. 2020), cannabinoids isolated from hemp roots extract exerted general anti-inflammatory effects rather than specific inhibition of inflammasome activation. The inhibitory effects of cannabinoids from hemp roots on the secretion of pro-inflammatory cytokine (TNF-α) may be accounted for the overall anti-inflammatory activities of Cannabis roots extracts in previously reported studies (Lima, et al., ref. 2021; Ryz et al., ref. 2017). However, several factors that may influence the anti-inflammatory effects of tested cannabinoids should be examined. For instance, the cytotoxicity of these cannabinoids in THP-1 cells may be accounted for the reduction of the pro-inflammatory cytokines. Thus, we assessed the viability of THP-1 cells treated with cannabinoids. Compounds 1–4 showed no significant cytotoxic effect (viability > 95%) at the concentration used for the anti-inflammasome assay (50 µM) whilst cannabivarin (5) reduced the cell viability by 54.7%, which possibly contributed to its suppressive effect on the production of IL-1β and TNF-α (see cell viability data in the Supplementary Data Figure S1). Therefore, further mechanistic investigation on the anti-inflammatory effects of cannabinoids from hemp roots should be conducted at non-cytotoxic concentrations. Nevertheless, this is the first study on the specific anti-inflammasome activity of cannabinoids isolated from hemp roots.
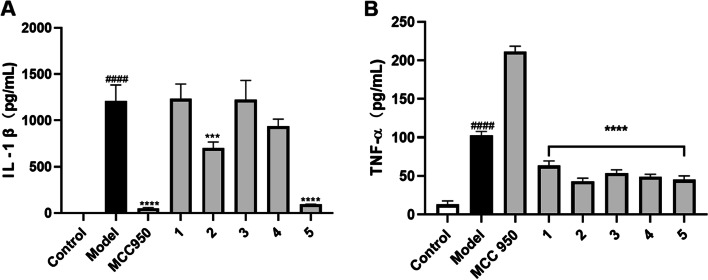
As none of the cannabinoids showed a selective inhibitory effect on inflammasome, we expanded the test samples for the anti-inflammation assay and further evaluated the suppressive effects of other isolates (7–32) on the production of IL-1β (Figure S2). At a concentration of 50 µM, compounds 11, 13, 19, 20, 25, 29, and 32 reduced the production of IL-1β by 23.7, 22.5, 25.6, 78.0, 24.1, 46.6, and 25.4%, respectively (Fig. 3A). These active compounds belong to different chemotypes including simple phenolics (11 and 13), lignans (19 and 20), triterpenoid (25), and unsaturated fatty acids (29 and 32). These active compounds were further assayed in the TNF-α inhibition assay, which showed that, apart from 19, they were inactive in reducing TNF-α production (Fig. 3B). Thus, compounds 11, 13, 20, 25, 29, and 32 showed an inhibitory effect on the activation of the inflammasome and they may contribute to the overall anti-inflammatory activity of cannabis (Lima, et al., ref. 2021; Ryz et al., ref. 2017). The selective inhibitory effects of compounds 11, 13, 20, 25, 29, and 32 on TNF-α production were in agreement with previously reported studies showing that these types of compounds were inflammasome inhibitors (Guo et al., ref. 2017; L’homme et al., ref. 2013; Ma, Huang, et al., ref. 2021; Villar-Lorenzo et al., ref. 2016). Further mechanistic studies are warranted to elucidate whether other pathways (e.g., via the antagonistic effect against the P2 × 7 receptor (Liu et al. ref. 2020)) were involved in the anti-inflammasome effects of these compounds from Cannabis roots.
In summary, a phytochemical investigation of a hemp roots extract led to the identification of 32 compounds including six cannabinoids (1–6) and 26 non-cannabinoid compounds. Isolates 14–21, 23, 27, and 32 were reported from Cannabis for the first time. Cannabinoids from the hemp roots extract exerted anti-inflammatory effects by reducing pro-inflammatory cytokines in an inflammasome model with THP-1 cells. Cannabinoids 2 and 5 reduced LPS-nigericin-induced production of cytokines IL-1β (by 42.2% and 92.4%, respectively) and TNF-α (by 58.4% and 56.1%, respectively). In addition, non-cannabinoids type compounds including 11, 13, 19,20, 25, 29, and 32 also showed promising anti-inflammatory effects. Findings from the current study expanded the understanding of the phytochemical constituents of cannabis roots and their anti-inflammatory effects, which may provide useful information for the utilization of cannabis roots as bioactive natural products.
Supplementary Materials
References
- RD Bennett, SM Norman, VP Maier. Intermediate steps in the biosynthesis of abscisic acid from farnesyl pyrophosphate in Cercospora rosicola. Phytochemistry, 1990. [DOI]
- N Bouaicha, P Amade, D Puel, C Roussakis. Zarzissine, a new cytotoxic guanidine alkaloid from the Mediterranean sponge Anchinoe paupertas. J Nat Prod, 1994. [DOI | PubMed]
- JJ Chen, WJ Lin, PC Shieh, IS Chen, CF Peng, PJ Sung. A New Long-Chain Alkene and Antituberculosis Constituents from the Leaves of Pourthiaea lucida. Chem Biodivers, 2010. [DOI | PubMed]
- HH Chen, YT Chen, YW Huang, HJ Tsai, CC Kuo. 4-Ketopinoresinol, a novel naturally occurring ARE activator, induces the Nrf2/HO-1 axis and protects against oxidative stress-induced cell injury via activation of PI3K/AKT signaling. Free Radic Biol Med, 2012. [DOI | PubMed]
- NA DaSilva, PP Nahar, H Ma, A Eid, Z Wei, S Meschwitz, NH Zawia, AL Slitt, NP Seeram. Pomegranate ellagitannin-gut microbial-derived metabolites, urolithins, inhibit neuroinflammation in vitro. Nutr Neurosci, 2019. [DOI | PubMed]
- M DellaGreca, C di Marino, A Zarrelli, B D’Abrosca. Isolation and phytotoxicity of apocarotenoids from Chenopodium album. J Nat Prod, 2004. [DOI | PubMed]
- K Devanathan, AN Stalin. Artocarpus heterophyllus Lam (Moraceae), 2020
- MA Elhendawy, AS Wanas, MM Radwan, NA Azzaz, ES Toson, MA Elsohly. Chemical and biological studies of cannabis sativa roots. Med Cannabis Cannabinoids, 2019. [DOI | PubMed]
- T Fujita, Y Kadoya, H Aota, M Nakayama. A new phenylpropanoid glucoside and other constituents of Oenanthe javanica. Bioscience Biotechnol Biochem, 1995. [DOI]
- JM Gao, M Wang, LP Liu, GH Wei, AL Zhang, C Draghici, Y Konishi. Ergosterol peroxides as phospholipase A2 inhibitors from the fungus Lactarius hatsudake. Phytomedicine, 2007. [DOI | PubMed]
- Y Goda, M Shibuya, U Sankawa. Inhibitors of prostaglandin biosynthesis from Mucuna birdwoodiana. Chem Pharm Bull, 1987. [DOI]
- M Guo, F An, H Yu, X Wei, M Hong, Y Lu. Comparative effects of schisandrin A, B, and C on Propionibacterium acnes-induced, NLRP3 inflammasome activation-mediated IL-1β secretion and pyroptosis. Biomed Pharmacother, 2017. [DOI | PubMed]
- H He, H Li, T Akanji, S Niu, Z Luo, D Li, NP Seeram, P Wu, H Ma. Synthesis and biological evaluations of oleanolic acid indole derivatives as hyaluronidase inhibitors with enhanced skin permeability. J Enzyme Inhib Med Chem, 2021. [DOI | PubMed]
- VK Hinge, SK Paknikar, KG Das, AK Bose, SC Bhattacharyya. Terpenoids-LXXXVI. Structure of epi-ψ-taraxastanonol and epi-ψ-taraxastanediol. Tetrahedron, 1966. [DOI]
- S Huh, YS Kim, E Jung, J Lim, KS Jung, MO Kim, J Lee, D Park. Melanogenesis inhibitory effect of fatty acid alkyl esters isolated from Oxalis triangularis. Biol Pharm Bull, 2010. [DOI | PubMed]
- J Ito, FR Chang, HK Wang, YK Park, M Ikegaki, N Kilgore, KH Lee. Anti-AIDS agents. 48.(1) Anti-HIV activity of moronic acid derivatives and the new melliferone-related triterpenoid isolated from Brazilian propolis. J Nat Prod, 2001. [DOI | PubMed]
- D Jin, K Dai, J Chen. Secondary metabolites profiled in cannabis inflorescences, leaves, stem barks, and roots for medicinal purposes. Sci Rep, 2020. [PubMed]
- E Kadowaki, Y Yoshida, N Baba, S Nakajima. Feeding stimulative activity of steroidal and secoiridoid glucosides and their hydrolysed derivatives toward the olive weevil (Dyscerus perforatus). Z Fur Naturforschung – Sect C J Biosci, 2003. [DOI]
- S Kishino, J Ogawa, A Ando, S Shimizu. Conjugated α-linolenic acid production from α-linolenic acid by Lactobacillus plantarum AKU 1009a. Eur J Lipid Sci Technol, 2003. [DOI]
- Kornpointner C, Sainz Martinez A, Marinovic S, Haselmair-Gosch C, Jamnik P, Schröder K, Löfke C, Halbwirth H. (2021). Chemical composition and antioxidant potential of Cannabis sativa L. roots.Industr Crops Prod.165.10.1016/j.indcrop.2021.113422.
- L L’homme, N Esser, L Riva, A Scheen, N Paquot, J Piette, S Legrand-Poels. Unsaturated fatty acids prevent activation of NLRP3 inflammasome in human monocytes/macrophages. J Lipid Res, 2013. [DOI | PubMed]
- J Li, G Wang, Y Qin, X Zhang, HF Wang, HW Liu, LJ Zhu, XS Yao. Neuroprotective constituents from the aerial parts of Cannabis sativa L. subsp. sativa. RSC Adv, 2020. [DOI | PubMed]
- KSB Lima, MEG da Cruz Silva, TC de Lima Araújo, CP da Fonseca Silva, BL Santos, LA de Araújo Ribeiro, PMN Menezes, MG Silva, ÉM Lavor, FS Silva, XP Nunes. Cannabis roots: Pharmacological and toxicological studies in mice. J Ethnopharmacol, 2021. [DOI | PubMed]
- C Liu, A Cai, H Li, N Deng, BP Cho, NP Seeram, H Ma. Characterization of molecular interactions between cannabidiol and human plasma proteins (serum albumin and γ-globulin) by surface plasmon resonance, microcalorimetry, and molecular docking. J Pharm Biomed Anal, 2022. [DOI | PubMed]
- Liu C, Li H, Xu F, Jiang X, Ma H, Seeram N. (2021). Cannabidiol protects human skin keratinocytes from hydrogen-peroxide-induced oxidative stress via modulation of the caspase-1–IL-1β axis. J Nat Prod, 84(5), 1563–1572. 10.1021/acs.jnatprod.1c00083.
- Liu C, Ma H, Slitt L, A., & Seeram P, N. (2020). Inhibitory effect of cannabidiol on the activation of NLRP3 inflammasome is associated with its modulation of the P2X7 receptor in human monocytes. J Nat Prod, 0(0). 10.1021/acs.jnatprod.0c00138.
- H Ma, H Li, C Liu, NP Seeram. Evaluation of cannabidiol’s inhibitory effect on alpha-glucosidase and its stability in simulated gastric and intestinal fluids. J Cannabis Res, 2021. [DOI | PubMed]
- H Ma, F Xu, C Liu, NP Seeram. A network pharmacology approach to identify potential molecular targets for cannabidiol’s anti-inflammatory activity. Cannabis Cannabinoid Res, 2021. [DOI | PubMed]
- Z Ma, Z Huang, L Zhang, X Li, B Xu, Y Xiao, X Shi, H Zhang, T Liao, P Wang. Vanillic acid reduces pain-related behavior in knee osteoarthritis rats through the inhibition of NLRP3 inflammasome-related synovitis. Front Pharmacol, 2021. [DOI]
- Marchetti L, Brighenti V, Rossi MC, Sperlea J, Pellati F, Bertelli D (2019). Use of 13C-qNMR spectroscopy for the analysis of non-psychoactive cannabinoids in fibre-type Cannabis sativa L. (Hemp). Molecules, 24(6). 10.3390/MOLECULES24061138.
- F Mohammed, P Sibley, D Guillaume, N Abdulwali. Chemical composition and mineralogical residence of maple syrup: a comprehensive review. Food Chem, 2022. [DOI | PubMed]
- MG Pizzolatti, LG Verdi, IMC Brighente, TDJC Neiva, J Schripsema, RB Filho. Anticoagulant effect and constituents of Baccharis illinita. Nat Prod Commun, 2006. [DOI]
- F Pollastro, A Minassi, LG Fresu. Cannabis phenolics and their bioactivities. Curr Med Chem, 2018. [DOI | PubMed]
- T Puopolo, C Liu, H Ma, NP Seeram. Inhibitory effects of cannabinoids on acetylcholinesterase and butyrylcholinesterase enzyme activities. Med Cannabis Cannabinoids, 2022. [DOI | PubMed]
- CSGP Queirós, S Cardoso, J Ferreira, I Miranda, MJv Lourenço, H Pereira. Characterization of Hakea sericea fruits regarding chemical composition and extract properties. Waste Biomass Valoriz, 2020. [DOI]
- MM Radwan, MA ElSohly, D Slade, SA Ahmed, IA Khan, SA Ross. Biologically active cannabinoids from high-potency Cannabis sativa. J Nat Prod, 2009. [DOI | PubMed]
- MH Rhee, Z Vogel, J Barg, M Bayewitch, R Levy, L Hanuš, A Breuer, R Mechoulam. Cannabinol derivatives: binding to cannabinoid receptors and inhibition of adenylylcyclase. J Med Chem, 1997. [DOI | PubMed]
- NR Ryz, DJ Remillard, EB Russo. Cannabis roots: a traditional therapy with future potential for treating inflammation and pain. Cannabis Cannabinoid Res, 2017. [DOI | PubMed]
- I Sakakibara, Y Ikeya, K Hayashi, M Okada, M Maruno. Three acyclic bis-phenylpropane lignanamides from fruits of Cannabis sativa. Phytochemistry, 1995. [DOI | PubMed]
- PB Sampson. Phytocannabinoid pharmacology: medicinal properties of Cannabis sativa constituents aside from the “Big Two”. J Nat Prod, 2021. [DOI | PubMed]
- T Seal. Antioxidant and Antiinflammatory Activities of Different Solvent Extracts and Isolated Compounds of Ipomoea pes-caprae (L) Sweet of Sunderban Mangrove Eco-complex. Asian J Chem, 2013. [DOI]
- Sirikantaramas S, Taura F. (2017). Cannabinoids: biosynthesis and biotechnological applications. Cannabis Sativa L – Bot Biotechnol. 183–206. 10.1007/978-3-319-54564-6_8/FIGURES/10.
- EFA Smania, D Monache, F Smania, A Yunes, RS Cuneo. Antifungal activity of sterols and triterpenes isolated from Ganoderma annulare. Fitoterapia, 2003. [DOI | PubMed]
- GF Sousa, FL Ferreira, LP Duarte, GDF Silva, MCTB Messias, SA Vieira Filho. Structural determination of 3β,11β-dihydroxyfriedelane from Maytenus robusta (Celastraceae) by 1D and 2D NMR. J Chem Res, 2012. [DOI]
- JM Stout, Z Boubakir, SJ Ambrose, RW Purves, JE Page. The hexanoyl-CoA precursor for cannabinoid biosynthesis is formed by an acyl-activating enzyme in Cannabis sativa trichomes. Plant J, 2012. [DOI | PubMed]
- A Villar-Lorenzo, AE Ardiles, AI Arroba, E Hernández-Jiménez, V Pardo, E López-Collazo, IA Jiménez, IL Bazzocchi, Á González-Rodríguez, ÁM Valverde. Friedelane-type triterpenoids as selective anti-inflammatory agents by regulation of differential signaling pathways in LPS-stimulated macrophages. Toxicol Appl Pharmcol, 2016. [DOI]
- RP Wang, HW Lin, LZ Li, PY Gao, Y Xu, SJ Song. Monoindole alkaloids from a marine sponge Mycale fibrexilis. Biochem Syst Ecol, 2012. [DOI]
- G Wang, L Zhu, Y Zhao, S Gao, D Sun, J Yuan, Y Huang, X Zhang, X Yao. A natural product from Cannabis sativa subsp. sativa inhibits homeodomain-interacting protein kinase 2 (HIPK2), attenuating MPP +-induced apoptosis in human neuroblastoma SH-SY5Y cells. Bioorg Chem, 2017. [DOI | PubMed]
- D Wu, Y Chen, Y Sun, Q Gao, H Li, Z Yang, Y Wang, X Jiang, B Yu. Target of MCC950 in Inhibition of NLRP3 inflammasome activation: a literature review. Inflammation, 2020. [DOI | PubMed]
- J Xu, C Yuan, G Wang, J Luo, H Ma, L Xu, Y Mu, Y Li, NP Seeram, X Huang, L Li. Urolithins Attenuate LPS-Induced Neuroinflammation in BV2Microglia via MAPK, Akt, and NF-κB Signaling Pathways. J Agric Food Chem, 2018. [DOI | PubMed]
- Xu Z, Wang C, Yan H, Zhao Z, You L, Ho CT. (2022). Influence of phenolic acids/aldehydes on color intensification of cyanidin-3-O-glucoside, the main anthocyanin in sugarcane (Saccharum officinarum L.). Food Chem. 373(Pt A) 10.1016/J.FOODCHEM.2021.131396.
- Y Xue, G Yao, Z Hu, Z Luo, Y Wang, Y Zhang. Chemical constituents from the leaves of Premna microphylla Turcz. Researchgate Net, 2013. [DOI]
- JY Yoon, JH Kim, KS Baek, GS Kim, SE Lee, DY Lee, JH Choi, SY Kim, HB Park, GH Sung, KR Lee, JY Cho, HJ Noh. A direct protein kinase B-targeted anti-inflammatory activity of cordycepin from artificially cultured fruit body of Cordyceps militaris. Pharmacogn Mag, 2015. [DOI | PubMed]
- W Zhou, S Guo. Components of the sclerotia of Polyporus umbellatus. Chem Nat Compd, 2009. [DOI]