
The role of endocannabinoid signaling in the cytoskeleton functionality in migrating neurons
Abstract
A fraction of neurons migrating through the developing brain are known to show nuclear envelope rupture and herniation of the chromatin in cytoplasm. We recently reported powerful streams of chromatin rupturing nuclear envelopes together with the plasma membranes in migrating cerebral neurons in mouse embryos. Such chromatin streams represent a novel form of cell pathology, which we named ‘piercing nuclear hernia’ (PNH). Simultaneous piercing of the nuclear and plasma membranes exposes nucleoplasm and cytoplasm to the intercellular space and may result in accidental cell death which, in contrast to the programed cell death mechanisms, are not detectable using biochemical or immunochemical markers for apoptosis, autophagy, or necrotic type of cell death. We also showed that the disfunction of the endocannabinoid system increases the probability of nuclear membrane rupture and chromatin herniation in developing brain. Indeed, about 40% of migrating neurons in cannabinoid type 1 receptor knock-out mouse embryos and wild type embryos exposed to two different agonists of the cannabinoid receptor show nuclear envelope ruptures or/and PNHs. This indicates that deviations from optimal functioning of the endocannabinoid system in under- or over-activity may trigger analogous mechanisms increasing the membrane’s vulnerability and chromatin herniation. The role of increased intranuclear pressure and cytoskeleton malfunction in the mechanism of nuclear envelope rupture is documented and commonly accepted. In accordance, our results provide evidence that optimal endocannabinoid signaling plays a role in cytoskeleton functionality in migrating neurons. In a fraction of neurons, catastrophic rupture of the nuclear and plasma membranes provokes ultrastructural pathology in the mitochondria and other organelles. At the same time, other neurons with PNH show generally normal ultrastructure that may indicate a mechanism of neuronal cell body repair. Further studies of neuronal cell body recovery may identify yet unknown molecular mechanisms and become instrumental for increasing regenerative capacity of neurons during neurodegenerative diseases, after traumatic brain injury and ischemic conditions. On the other hand, the demonstrated novel pathology of PNH in migrating cells and the procedure of its upregulation may be useful for inducing breaks of the plasma membrane and death of metastatic tumor cells.
Article type: Research Article
Keywords: cannabinoid type 1 receptor, nuclear envelope rupture, cell membrane rupture and repair, accidental cell death, brain development pathology, recovery of neurons
Affiliations: Department of Neuroscience, Yale University School of Medicine and Kavli Institute for Neuroscience, New Haven, Connecticut 06510, USA
License: CC BY 4.0 This is an open- access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Article links: DOI: 10.18103/mra.v13i10.7041 | PubMed: 41695078 | PMC: PMC12904361
Relevance: Moderate: mentioned 3+ times in text
Full text: PDF (1.7 MB)
Introduction
Medical cannabis products are approved or tolerated in many countries and, increasingly, self-administered by pregnant individuals seeking relief from pain, nausea, or depression. The use of marijuana and synthetic cannabinoids during pregnancy and lactation poses potential risks for intrauterine and postnatal brain development, particularly as cannabinoids can appear in breast milk. The extent to which marijuana and medicinal cannabinoids affect immature brains is still the subject of debate and extensive biomedical studies [Reviewed in1–4].
The endocannabinoid system is involved in several cognitive and physiological processes, including cardiovascular regulation, fertility, pregnancy, prenatal and postnatal development, activity of the immune system, food consumption and energy metabolism1,5–12. Endocannabinoid signaling via cannabinoid type 1 receptor (CB1R) participates in retrograde synaptic modulation but also involves non-synaptic pathways13–15. In addition to abundant expression in the axonal plasma membrane, CB1R accumulates in the membranes of intracellular vesicles in the cell bodies of adult animals. As demonstrated in rodent and rhesus macaque embryos, CB1R also accumulates in immature interneurons tangentially migrating through the embryonic cerebral marginal zone and projection neurons vertically migrating through the intermediate zone and cortical plate16–20. One demonstrated role of CB1R, which putatively involve the intracellular vesicles, is self-inhibition of cholecystokinin-expressing interneurons and a subpopulation of cortical pyramidal neurons through the cell’s autonomous Ca2+-dependent production of endocannabinoids and K+ channel activation21,22. Two known endocannabinoids – anandamide and 2-arachidonoylglycerol (2-AG) – show divergent behaviors regarding CB1R functionality. Particularly, anandamide, but not 2-AG, inhibits voltage-gated Na+ channels located in the intracellular compartments of the cell bodies rather than in the synaptic boutons, whereas presynaptic endocannabinoid activity is predominantly attributed to 2-AG23. The endocannabinoid system influences synapse target selection by pyramidal neurons and GABAergic interneurons through CB1R internalization from axonal filopodia and chemorepulsion of the growth cones24,25. We previously demonstrated that, in cannabinoid type 1 receptor knock-out (CB1R−/−) mouse embryos, migrating cerebral projection neurons deviated from the vertical orientation, implicating CB1R’s role in cellular migration18. In vivo inhibition of the cannabinoid receptors disrupted migration of immature neurons from the rostral migratory stream in young postnatal mice, as evident by the decreasing length and increasing branching of their leading processes26. Cell reorganization during migration, as well as maintenance of the shape of mature cells, are in high degree determined by the cytoskeleton components such as centrosomes, microtubules, actin, myosin II, dynein and others27–30. However, the molecular mechanisms of the endocannabinoid system’s action in the cytoarchitecture of the developing brain are still enigmatic.
Piercing nuclear hernia is a novel ultrastructural pathology of migrating neurons
Recently, we reported novel ultrastructural pathology in developing neurons related to CB1R knock-out and in utero application of synthetic cannabinoids31. Particularly, we documented numerous local breakups of the NE accompanied by herniation of the nuclear chromatin into the cytoplasm (referred as NE rupture hereafter) in neurons migrating through the mouse embryo cerebrum (Figure 1A). Surprisingly, in a fraction of the cells, streams of herniated chromatin ruptured not only the NE, but also the plasma membrane, exposing cytoplasm and nucleoplasm to the intercellular space (Figure 1B). As it was the first description of simultaneous rupture of the NE and the plasma membrane, we named this cellular pathology ‘piercing nuclear hernia’ (PNH). For detailed characterization of this phenomenon, we performed extensive analyses of electron microscopy imagery of migrating neurons in CB1R−/− embryos, wild type control mouse embryos and embryos after in utero application of CB1R agonists CP-55940 and WIN 55,212–2. Application of two different CB1R agonists produced similar chromatin herniation, the extent of which was dose-dependent. Using three-dimensional electron microscopy analysis, we identified 122 nuclear hernias (including 65 NE ruptures and 57 PNHs). In CB1R−/− embryos and in wild type embryos shortly exposed to high doses of CB1R agonists, about 40% of randomly analyzed migrating projection neurons and interneurons demonstrated NE ruptures or PNHs (Figure 1C). This indicates that deviations from optimal functioning of the endocannabinoid system in either direction – knock-out of CB1R or its temporary overstimulation – increase likelihood of altered cellular nucleus and provoke chromatin herniation in a similar way31.
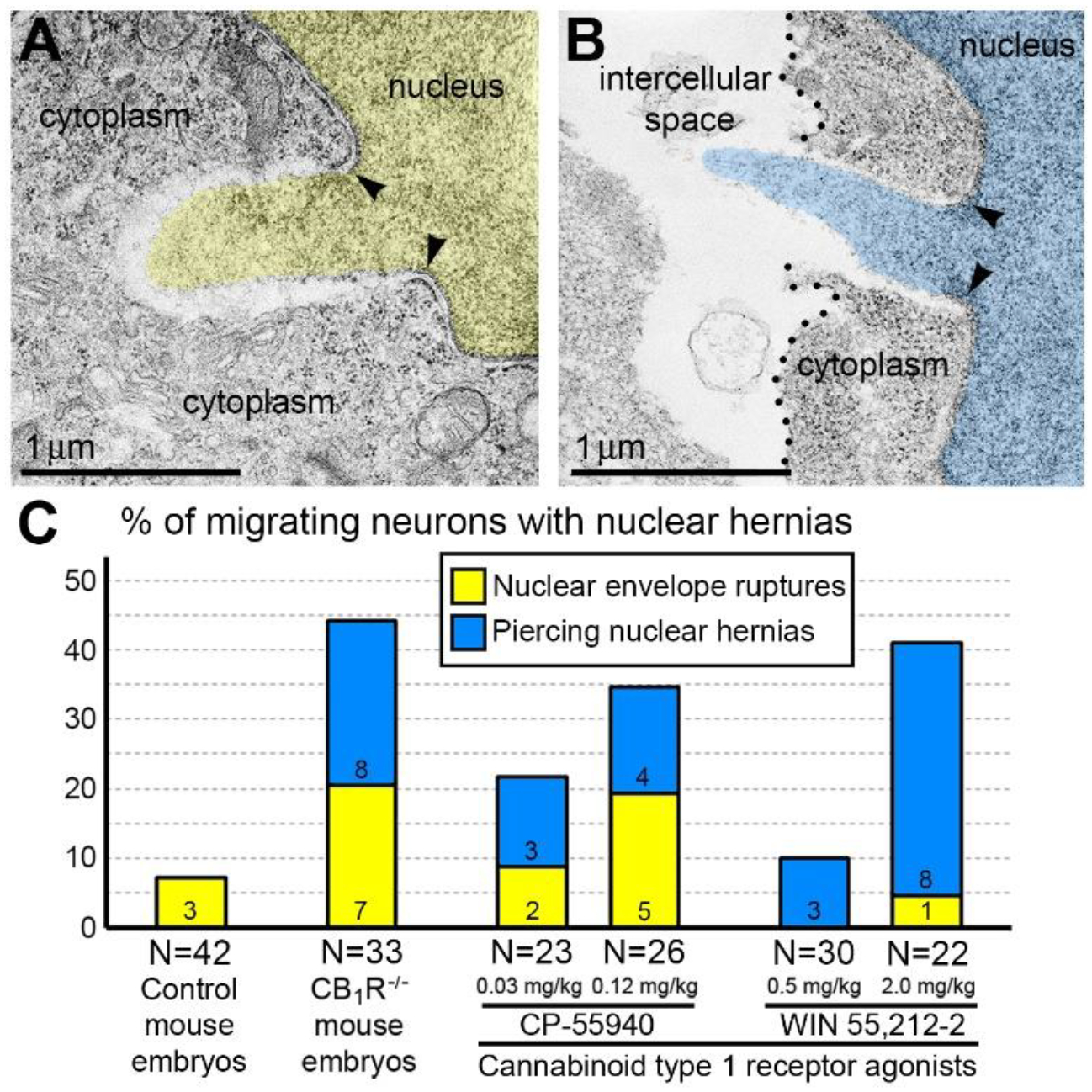
Morphometric analyses showed that volume of the streams of herniated chromatin varied from 0.02 to 1.63 μm3 (Published in31). In some analyzed cells, the PNH stream was narrow and reached the length of several microns (Figure 2). Although the molecular mechanism of PNH remains unclear, the large volume and length of the herniated chromatin streams point to abnormally high intranuclear pressure during nuclear translocation – a condition that predisposes cells for NE rupture32. Our discovery of a correlation between the disorder of the endocannabinoid system and frequency of NE ruptures and PNHs could be useful for better understanding the effects of cannabinoids in the developing brain (Reviewed in1,2). Identification of in vivo experimental conditions that increase frequency and power of chromatin herniation may be instrumental for further investigation of the mechanisms of the membrane’s rupture. Here, we review probable cellular mechanisms and developmental consequences of the detected cell pathology as well as the directions of further studies and prospective medicinal application of the obtained knowledge.
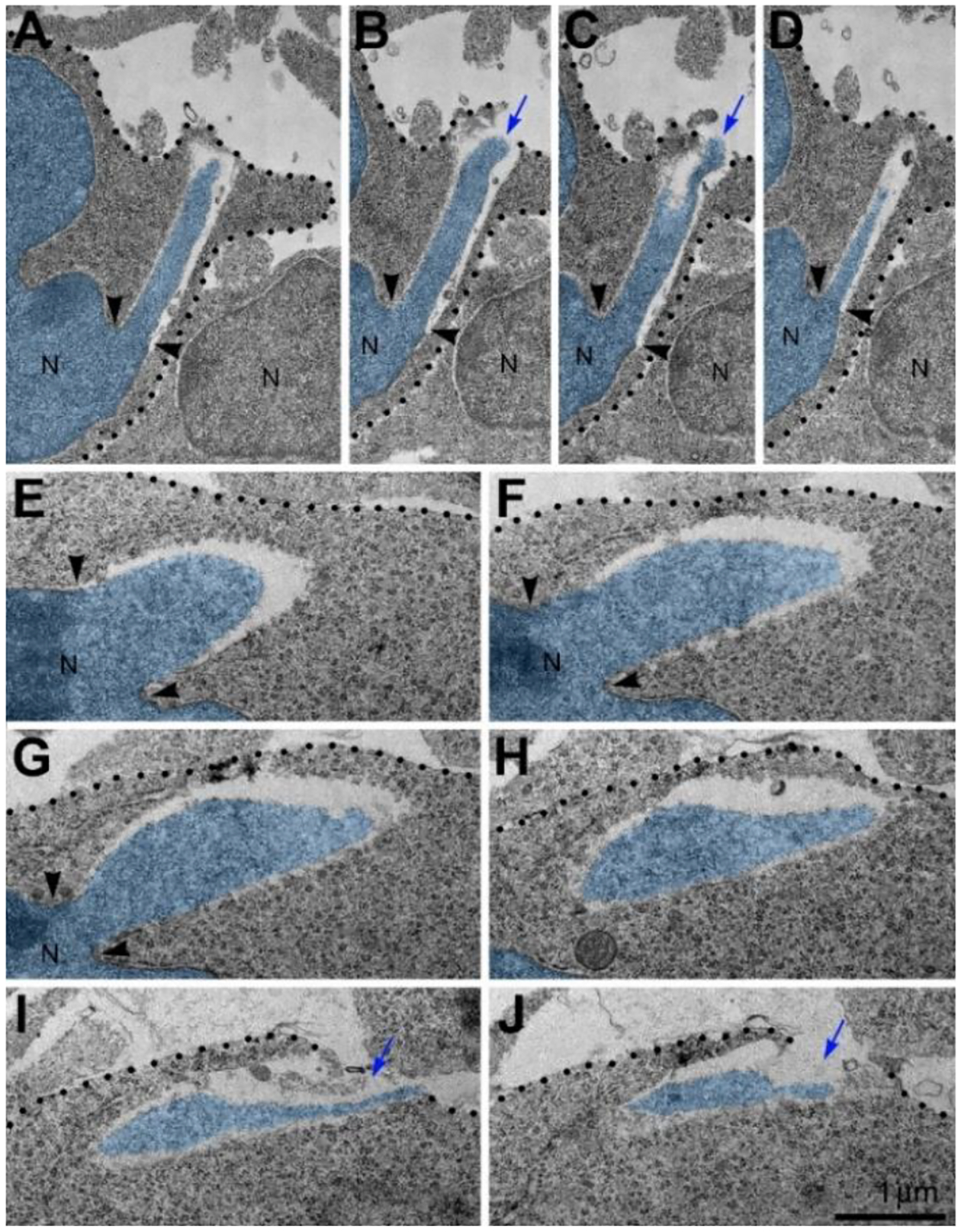
Mechanisms of nuclear envelope rupture and recovery
The nuclear envelope (NE) consists of the inner and outer nuclear membranes and the nuclear lamina – a composite matrix assembled on the inner nuclear membrane. The main function of the NE is to allow molecular exchange between the nucleoplasm and the cytoplasm through specified nuclear pores. Recent works highlighted the dynamic properties of the nuclear membranes and demonstrated that dysregulation of their functions has significant consequences for the cell33–37. In addition to biochemical inputs, nuclei of the cells may be exposed to intrinsic and extrinsic mechanical forces transmitted by the cytoskeleton and nucleoskeleton that trigger dynamic changes in nuclear morphology33,35,36,38. Ruptures of the NE have been observed in several species in vitro and in vivo indicating that it is a consequence of the environmental conditions rather than a species-specific phenomenon [Reviewed in39]. For example, NE rupture was observed in neurons migrating during brain development40,41. NE ruptures were extensively studied in many cancer cell lines during cell migration through tightly constricted areas34,42–44. NE ruptures can be induced in non-tumorigenic cell lines by generating a lamin B1 deficiency or depletion of two major tumor suppressors, p53 and retinoblastoma (Rb1)40,45,46. Cultivated cells can also be provoked to experience NE rupture by external mechanical force38,44,47, human immunodeficiency virus infection48, and activation of apoptosis, inflammation, or autophagy45,49–51. Emerging data suggests that extensive deformation of the cell and its nucleus during constrained migration transmits substantial physical stress to the NE and may result in its rupture, which in turn leads to chromatin herniation, uncontrolled exchange of nucleocytoplasmic content, DNA damage, and cell death33,40,42,44,52,53. Distribution of a diffusible fluorescent marker from a ruptured nucleus through the cytoplasm was observed as early as minutes after NE rupture, indicating the high speed of the reaction42,43,48,54,55. The full impact of nucleocytoplasmic mixing is likely to be extensive yet remains poorly understood. Depletion, or mutation of nuclear lamin proteins as well as conditions that impair the connections between the inner nuclear membrane and the chromatin increase the probability of NE ruptures32,34,35,40,42,43,47,52,56–58. Thus, NE ruptures may be caused by defects in lamina organization, or increased intranuclear pressure transmitted by actin cytoskeleton and/or nucleoskeleton32. Several NE repair mechanisms have been proposed, including attachment of endoplasmic reticulum sheets to the exposed chromatin, spreading of the preserved outer nuclear membrane, plugging with membrane fragments, and resealing by protein complexes [Reviewed in39,59,60]. As identified using fluorescent live cell imaging and other methods, BAF (barrier-to-autointegration factor), ECRT-III (the endosomal sorting complexes required for transport-III) and several nuclear membrane proteins such as emerin, LEM-domain containing protein 2 and lamin A/C are involved in the repair of the nuclear envelope54,61–63. Cancer and immune cells migrating in vitro through constricted microchannels of various sizes reseal their ruptured NE using ESCRT-III and by doing so reduce DNA damage and cell death42,44. An individual NE rupture may persist for minutes to hours before repair, however, unrepaired NE ruptures eventually result in the cell’s death42–44,54,64.
Mechanisms of plasma membrane rupture and recovery
Separation of cellular inner content from the environment is one of the fundamental challenges of life due to the variety of mechanical and biochemical stresses that pose constant risks of compromise to the function and viability of the cell. To counteract this threat, eukaryotic cells have developed efficient repair mechanisms, which are heavily conserved across species and seem to have co-evolved with the emergence of vital properties of the plasma membrane65–68. Membrane repair mechanisms include distinct reactions depending on the size of the rupture [Reviewed in66,68,69]. Namely, tiny membrane injuries (less than a nanometer) may be repaired spontaneously by reciprocal attraction of the hydrophobic domains of lipids. Injuries larger than a few nanometers in diameter, such as those caused by bacterial pore-forming toxins, require the help of an active membrane repair mechanism that includes endosomal uptake or exosomal secretion of the bacteria-lined pores. Repair of very large disruptions of the plasma membrane (hundreds to thousands of nanometers in diameter) requires complicated vesicle-vesicle and vesicle-plasma membrane fusion mechanisms. The initial repair steps include plugging the membrane rupture with conglomerates of vesicles or membrane whorls that crudely ‘patch’ the rupture, restricting the leak of cytoplasm and providing time for reconstruction of intact plasma membrane66,68–70. A major trigger for the signaling cascade that precedes membrane resealing is the influx of Ca2+ from the intercellular space through the membrane disruption. Subsequent vesicle deposition and fusion involves complex Ca2+-dependent pathways acting through the cAMP signaling cascade, cytoskeleton reconstruction, SNAP receptors, the phospholipase enzymes, calpain proteases and others. The membrane patch serving a temporary barrier is subsequently remodeled or removed via exocytic or endocytic machinery66,68,69. Alternatively, if the damaged plasma membrane is not repaired within minutes, activation of apoptotic or necrotic pathways mediated through Ca2+ influx often results in cell death69,71. Repair mechanisms of large membrane ruptures are effective for certain cell types, e.g., muscular cells, oocytes, epithelial cells, and invasive cancer cells69,70. While neurons may effectively restore transected axons, recovery of neuronal cell bodies following severe ruptures of plasma membranes has not been demonstrated65.
Probable death or survival of neurons with piercing nuclear hernia
PNH detected in migrating neurons catastrophically breaks isolation of the inner content of the cell from the environment31. Cells exposed to extreme physical, chemical, or mechanical stimuli may immediately lose their structural integrity and die in an uncontrollable manner termed ‘accidental cell death’, which is opposite to mechanisms of programed cell death such as apoptosis, autophagy, or necrosis-type cell death65,72–75. Cases of accidental cell death are virtually undetectable using biochemical or immunochemical methods such as TUNEL staining, caspase immunolabeling, or labeling of autophagy markers. Electron microscopy visualizes organelles regardless of their functional conditions and can detect dead cells or cellular remnants75.
Given the high frequency of cells with NE rupture and PNH in certain experimental conditions (Figure 1C) one might expect to detect many dead cells. Surprisingly, we did not observe a considerable number of cellular remnants or cells with ultrastructural features of necrotic, apoptotic or autophagy type degradation in the embryos containing numerous cells with herniated nuclei. We also did not identify massive swelling of the mitochondrial matrix that was recognized to be irreversible and foreshadowing cell death through a necrotic mechanism76,77. Instead, a fraction of the cells showed reduction of mitochondrial length – evidence of mitochondrial fission. It is known that morpho-functional conditions of mitochondria correlate with general cellular functionality, while predominance of mitochondrial fission over fusion may serve as evidence of disordered cellular energetics or other moderate malfunctions76–82. We often observed dramatic heterogeneity of ultrastructure between certain cells from the same sample. For example, cells with PNH demonstrated short mitochondria while adjacent non-herniated cells showed normally sized mitochondria (Figure 3). At the same time, other cells with PNH showed normal ultrastructure of mitochondria and other organelles31. Different time courses of NE rupture (likely occurring in the span of seconds42,43,48,54,55) and mitochondrial reaction (delay in the span of hours76) may partially explain the divergence between emergence of PNH and reaction of organelles in the embryos temporarily exposed to CB1R agonists. Nevertheless, the absence of a considerable number of dead cells in CB1R−/− embryos, which constantly experience disorder of endocannabinoid signaling, raises a supposition that PNH may be repaired rather than always be fatal for the cell. This observation aligns with the fact that CB1R−/− mice generally do not exhibit obvious behavioral phenotypes83. In accordance, partial recovery of cells after NE rupture has been documented39,59. Surviving neurons were reported after transection of the axons, but not after damage of the cell bodies65. We considered morphological features that might promote recovery of migrating neurons with PNH. Namely, for a fraction of the cells with PNH, the area of the plasma membrane rupture was closely surrounded by adjacent cell bodies or processes, which may contribute to decreasing the flow of cytoplasm and blocking the leak (Figure 4). Although the effectiveness of such blocking is difficult to estimate in direct experiments, reduction of the cytoplasm leak because of ‘patching’ by adjacent cells, together with a possible delay of the organelles’ reaction, may explain dissimilarity of the ultrastructure in several cells with PNH. Thus, although death of cells with ruptured NE and plasma membrane is probable, PNH in mouse embryo brains may be non-fatal for a fraction of the affected cells. This suggests the possibility of recovery for catastrophically damaged neuronal cell bodies, which has not been demonstrated previously. Deeper study of the newly detected role of the endocannabinoid system in rupture and recovery of cell membranes could uncover presently unknown molecular mechanisms that prove to be instrumental for increasing regenerative capacity of neuronal cell bodies65,66,68,70,71,84.
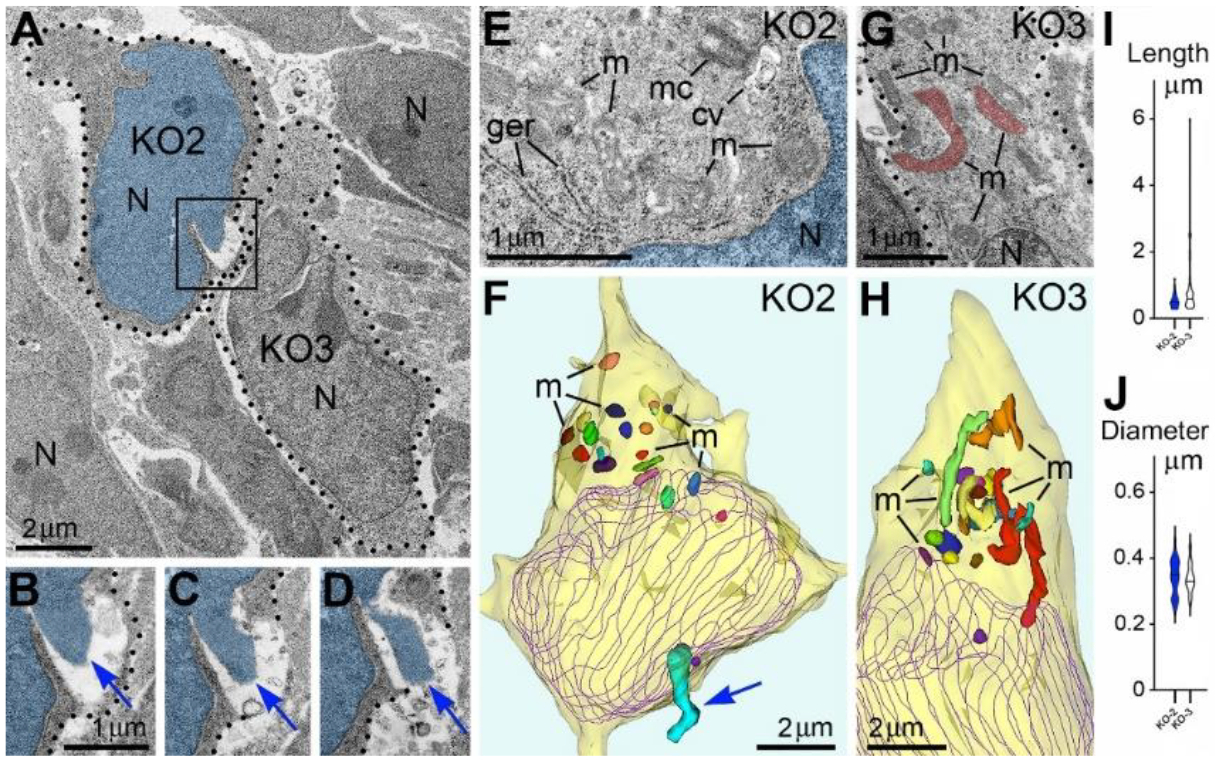
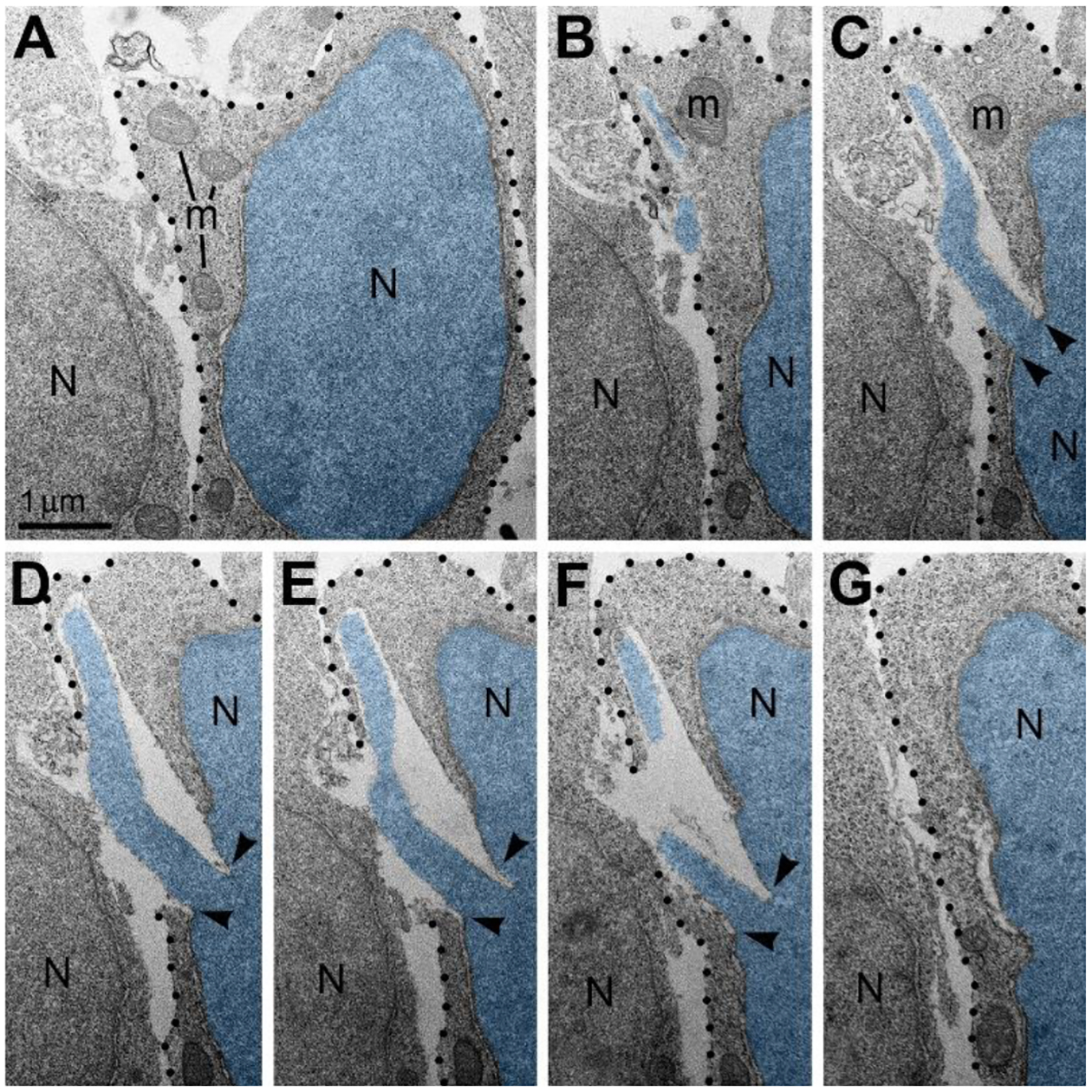
Nuclear envelope ruptures and piercing nuclear hernias suggest role of the endocannabinoid system on cytoskeleton functionality in cell bodies
High percentages of cells with nuclear hernias in the embryo cerebrum of CB1R−/− mice and wild type mice exposed to CB1R agonists CP-55940 and WIN 55,212–2 (Figure 1C) indicate involvement of the endocannabinoid system in the mechanics of NE rupture31. Judging from the large volume and the length of herniated chromatin streams, the NE ruptures and PNHs in the nuclei translocating through tightly packed tissue such as the developing mammalian cerebrum are likely consequences of increased intranuclear pressure and suboptimal function of the cytoskeleton32,52. In accord, chronic treatment of adult rats with WIN 55,212–2 increased expression of neurofilaments Nf-160 and Nf-200, and microtubule-associated protein-2 (MAP-2). Meanwhile, CB1R−/− mice demonstrated a lower expression of the neurofilaments and MAP-2, along with reduced dendritic arborization in the hippocampus85–87. Such deviated expression of cytoskeletal components after stimulation or knock-out of CB1R establishes evidence of cannabinoid signaling participating in neuronal cytoskeleton consolidation. Direct link of endocannabinoid system, and particularly CB1R, with the function of the cytoskeleton was demonstrated in the context of axonal pathfinding. This mechanism involves CB1R internalization from filopodia and chemorepulsion of the axonal growth cones by activating RhoA GTPase (small guanosine triphosphatase) and Rho-associated kinase (ROCK)24. RhoA GTPases are known for their role as molecular switches transducing stimuli to the actin cytoskeleton88,89. CB1R-activation rapidly and reversibly contracts the neuronal actomyosin cytoskeleton through coupling to G12/G13 proteins that produce Rho- and ROCK-mediated non-muscle Myosin II (NM II) activation in the axons of cultured neurons90. Early neurula stage chicken embryos and early tailbud stage frog embryos exposed to CB1R agonist ACEA (arachidonyl-2′-chloroethylamide) or CB1R inverse agonist AM251 demonstrated disordered migration of neural crest cells. The effects of deviated CB1R signaling in over- and under-activity were mimicked by exposing the embryos to NM II inhibitor Blebbistatin, which indicated that CB1R regulated migration of neural cells in chicken and frog embryos through cytoskeleton functionality91. Thus, the unknown role of CB1R in the developing brain of many different species may include maintaining optimal function of the actomyosin cytoskeleton, which is crucial for translocation of neuronal cell bodies.
Emergences of nuclear envelope ruptures and piercing nuclear hernias suggest role of cannabinoid type 1 receptor in neuron migration
Early expression of CB1R indicates its probable involvement in prenatal and postnatal development of the mammalian brain17,19. Numerous studies indicate that prenatal inhibition or overstimulation of the endocannabinoid system may provoke long-term consequences in the offspring, for example, altered breathing, disturbed suckling, and memory deficit linked to disorder of glutamate-ergic neurons2,12,92–95, but many aspects of the underlying mechanisms remain enigmatic. Our data indicates that disorder of the endocannabinoid system may provoke NE ruptures and PNHs in migrating neurons31. Although we did not document dead or severely damaged cells, our study does not assume a complete recovery and absence of a long-term effect on the cells experiencing NE ruptures or PNHs. More likely, temporary damage of NE and plasma membrane provokes a delay or disorientation of the cell migration. A body of published evidence supports the involvement of cannabinoid signaling in migration of immature neurons. Particularly, inhibition of cannabinoid receptors or DAGL (diacylglycerol lipase, the enzyme synthetizing the endocannabinoid 2-AG) decreases migration of cultivated neural stem cells and neuroblasts from the rostral migratory stream explant culture. Consequently, activation of cannabinoid receptors or preventing breakdown of 2-AG increases migration26. Early post-natal CB1R−/− mice display higher numbers of projection neurons in deeper cortical layers and lower numbers in the superficial layers identifying altered neuronal migration96. Similar mis-location of neurons was detected in fatty-acid amide hydrolase (enzyme degrading endocannabinoid anandamide) knock-out mice (FAAH−/−) indicating that supposed overstimulation of CB1R through increased amount of anandamide may also disorder cellular migration96. Acute knockdown of CB1R by in utero electroporation of siRNA in mouse embryos delayed colonization of cortical plate by projection neurons; instead, many cells were stacked in the intermediate and subventricular zones97. Thus, delay in radial migration of neurons in the absence or altered functionality of CB1R may be the reason for previously identified moderate disorder of cerebral cytoarchitecture and brain dysfunction in CB1R−/− mice, or cognitive deviations in animals and humans exposed to cannabis during development1,2,12,83,92–95.
Our quantitative immunohistochemical analyses show the probability (rather than unequivocal proof) of a link between chromatin herniation and the amount of CB1R in the cell body31. This leaves the point unsolved, whether cannabinoid signaling disorder upregulates NE ruptures and PNHs directly in the CB1R-expressing cells, or the action is systematic, for example, through vasodilation and hypotensive effects in the entire fetus, placenta or maternal organism1,2,9,10. Accordingly, mouse embryos from pregnant dams exposed to cannabinoid agonist CP-55,940 combined with alcohol vapors demonstrated reduced internal carotid artery blood flow and significant subsequent perinatal mortality; the survived young adult offspring showed induced deficit in neurobehavioral motor outcomes10,98. Our finding opens an opportunity for further investigation if temporary disorder of the endocannabinoid system, for example, in cases of recreational cannabis use by pregnant women, may provoke accidental death or malfunction in a fraction of migrating neurons.
Prospective study of cannabinoid system and piercing nuclear hernia in aspects of medicinal application
Therapeutic benefits of cannabis as an analgesic and antiemetic agent are well-known and applied for centuries3,99. Cannabinoid-based anti-epileptic and anti-spasticity medications such as Epidiolex and Nabiximols (Sativex) have demonstrated efficacy in randomized controlled trials100,101. Inhibition of MAGL (monoacylglycerol lipase – enzyme degrading endocannabinoid 2-AG) has been proposed as a potential therapeutic approach for treatment of diverse neurological and neurodegenerative diseases, such as multiple sclerosis, Alzheimer’s disease, Parkinson’s disease, amyotrophic lateral sclerosis, and traumatic brain injury102–108. Our finding of involvement of the endocannabinoid system in the membrane’s rupture and probably in the recovery of the disordered cells31 may help to reveal potential neuroprotective action of cannabinoids.
Two cannabinoid drugs (dronabinol and nabilone) are approved by the U.S. Food and Drug Administration for the prevention or treatment of cancer-related side effects, such as nausea and vomiting. Numerous in vitro and in vivo experiments on cancer models show that cannabinoids can kill cancer cells, making cannabis-based medicine a promising option to effectively modulate growth of certain tumor types. The observed effects of the cannabinoids’ application include inhibiting tumor growth and metastasis, reducing cell viability by promoting apoptosis, and inhibiting angiogenesis; however, the specific molecular mechanisms at play were not identified in detail. Accordingly, suspected anticancer benefits of cannabinoids have yet to be confirmed in clinical trials109–118. Our findings suggest a novel mechanism of cannabinoid action on the nuclei of migrating cells and, after additional study, may be instrumental for inducing breaks of the plasma membrane in metastatic tumor cells. Although NE rupture resulting in genomic instability might promote cancer progression, unstable NE also represents a particular weakness of metastatic cancer cells. Accordingly, NE ruptures detected in many cancer cell lines during cell migration through tightly constricted areas often result in cell death42–45. Conditions that have potential to increase the probability of NE rupture and death of metastatic cells are promising as cancer treatments34,42. We suggest performing extensive investigation of conditions of PNH generation for the purpose of targeting metastatic cancer cells. The conditions increasing frequency and power of NE ruptures may include manipulations of the endocannabinoid system and probably other molecular mechanisms. This opens an opportunity for a new research direction that may result in the development of therapeutic applications of cannabinoids for targeting cancer cells.
Conclusions
Neurons migrating through the developing cerebral cortex show nuclear envelope ruptures and herniations of the chromatin in cytoplasm. Powerful streams of herniated chromatin may also pierce plasma membranes and expose nucleoplasm and cytoplasm to the intercellular space representing a novel form of ultrastructural cell pathology. Knock-out or over stimulation of the cannabinoid type 1 receptor increases probability and power of the nuclear envelope rupture and chromatin herniation. As previously demonstrated, nuclear envelope ruptures result from disfunction of the cytoskeleton. Thus, deviation of endocannabinoid signaling from optimal functionality in either direction likely provokes disfunction of the cytoskeleton in migrating neurons. Catastrophic damage of the main cellular membranes normally results in accidental cell death. In perspective, this newly discovered pathology of migrating cells and technique of its upregulation may be applied for inducing breaks of the plasma membrane and death of metastatic tumor cells. On the other hand, observed evidence of recovery of neuronal cell bodies, after further study, may become instrumental for increasing regenerative capacity of damaged neurons.
References
- RJ Rodrigues, JM Marques, A Köfalvi. Cannabis, Endocannabinoids and Brain Development: From Embryogenesis to Adolescence.. Cells., 2024. [DOI]
- MK Mulligan, KM Hamre. Influence of prenatal cannabinoid exposure on early development and beyond.. Adv Drug Alcohol Res., 2023. [DOI | PubMed]
- M Maccarrone, V Di Marzo, J Gertsch. Goods and Bads of the Endocannabinoid System as a Therapeutic Target: Lessons Learned after 30 Years.. Pharmacol Rev., 2023. [DOI | PubMed]
- DC D’Souza, M DiForti, S Ganesh. Consensus paper of the WFSBP task force on cannabis, cannabinoids and psychosis.. World J Biol Psychiatry., 2022. [DOI | PubMed]
- M Koch, L Varela, JG Kim. Hypothalamic POMC neurons promote cannabinoid-induced feeding.. Nature., 2015. [DOI | PubMed]
- V Di Marzo, SK Goparaju, L Wang. Leptin-regulated endocannabinoids are involved in maintaining food intake.. Nature., 2001. [DOI | PubMed]
- M Maccarrone, I Bab, T Bíró. Endocannabinoid signaling at the periphery: 50 years after THC.. Trends in pharmacological sciences., 2015. [DOI | PubMed]
- O Sagredo, J Palazuelos, A Gutierrez-Rodriguez, V Satta, I Galve-Roperh, J Martinez-Orgado. Cannabinoid signalling in the immature brain: Encephalopathies and neurodevelopmental disorders.. Biochemical pharmacology., 2018. [DOI | PubMed]
- B Bányai, Z Vass, S Kiss. Role of CB1 Cannabinoid Receptors in Vascular Responses and Vascular Remodeling of the Aorta in Female Mice.. International journal of molecular sciences., 2023. [DOI]
- SK Rouzer, A Sreeram, RC Miranda. Reduced fetal cerebral blood flow predicts perinatal mortality in a mouse model of prenatal alcohol and cannabinoid exposure.. BMC Pregnancy Childbirth., 2024. [DOI | PubMed]
- YM Morozov, M Koch, P Rakic, TL Horvath. Cannabinoid type 1 receptor-containing axons innervate NPY/AgRP neurons in the mouse arcuate nucleus.. Mol Metab., 2017. [DOI | PubMed]
- M Maccarrone, M Guzmán, K Mackie, P Doherty, T Harkany. Programming of neural cells by (endo)cannabinoids: from physiological rules to emerging therapies.. Nature reviews Neuroscience., 2014. [DOI | PubMed]
- I Katona, TF Freund. Multiple functions of endocannabinoid signaling in the brain.. Annual review of neuroscience., 2012. [DOI]
- D Piomelli. The molecular logic of endocannabinoid signalling.. Nature reviews Neuroscience., 2003. [DOI | PubMed]
- RI Wilson, RA Nicoll. Endocannabinoid signaling in the brain.. Science., 2002. [DOI | PubMed]
- I Katona, EA Rancz, L Acsady. Distribution of CB1 cannabinoid receptors in the amygdala and their role in the control of GABAergic transmission.. J Neurosci., 2001. [PubMed]
- YM Morozov, TF Freund. Post-natal development of type 1 cannabinoid receptor immunoreactivity in the rat hippocampus.. Eur J Neurosci., 2003. [PubMed]
- YM Morozov, K Mackie, P Rakic. Cannabinoid Type 1 Receptor is Undetectable in Rodent and Primate Cerebral Neural Stem Cells but Participates in Radial Neuronal Migration.. International journal of molecular sciences., 2020. [DOI]
- YM Morozov, M Torii, P Rakic. Origin, early commitment, migratory routes, and destination of cannabinoid type 1 receptor-containing interneurons.. Cerebral cortex (New York, NY: 1991)., 2009. [DOI]
- C Bernard, M Milh, YM Morozov, Y Ben-Ari, TF Freund, H Gozlan. Altering cannabinoid signaling during development disrupts neuronal activity.. Proc Natl Acad Sci U S A., 2005. [DOI | PubMed]
- A Bacci, JR Huguenard, DA Prince. Long-lasting self-inhibition of neocortical interneurons mediated by endocannabinoids.. Nature., 2004. [DOI | PubMed]
- S Marinelli, S Pacioni, A Cannich, G Marsicano, A Bacci. Self-modulation of neocortical pyramidal neurons by endocannabinoids.. Nature neuroscience., 2009. [DOI | PubMed]
- LJ Steiger, T Tsintsadze, GB Mattheisen, SM Smith. Somatic and terminal CB1 receptors are differentially coupled to voltage-gated sodium channels in neocortical neurons.. Cell reports., 2023. [DOI | PubMed]
- P Berghuis, AM Rajnicek, YM Morozov. Hardwiring the brain: endocannabinoids shape neuronal connectivity.. Science., 2007. [DOI | PubMed]
- E Keimpema, K Barabas, YM Morozov. Differential subcellular recruitment of monoacylglycerol lipase generates spatial specificity of 2-arachidonoyl glycerol signaling during axonal pathfinding.. J Neurosci., 2010. [DOI | PubMed]
- MJ Oudin, S Gajendra, G Williams, C Hobbs, G Lalli, P Doherty. Endocannabinoids regulate the migration of subventricular zone-derived neuroblasts in the postnatal brain.. J Neurosci., 2011. [DOI | PubMed]
- JW Tsai, KH Bremner, RB Vallee. Dual subcellular roles for LIS1 and dynein in radial neuronal migration in live brain tissue.. Nature neuroscience., 2007. [DOI | PubMed]
- DJ Solecki, L Model, J Gaetz, TM Kapoor, ME Hatten. Par6alpha signaling controls glial-guided neuronal migration.. Nature neuroscience., 2004. [DOI | PubMed]
- P Rakic, E Knyihar-Csillik, B Csillik. Polarity of microtubule assemblies during neuronal cell migration.. Proc Natl Acad Sci U S A., 1996. [DOI | PubMed]
- A Bellion, JP Baudoin, C Alvarez, M Bornens, C Métin. Nucleokinesis in tangentially migrating neurons comprises two alternating phases: forward migration of the Golgi/centrosome associated with centrosome splitting and myosin contraction at the rear.. J Neurosci., 2005. [DOI | PubMed]
- YM Morozov, P Rakic. Abnormalities of the endocannabinoid system produce piercing nuclear hernias in migrating cerebral neurons.. iScience., 2025. [DOI | PubMed]
- EM Hatch, MW Hetzer. Nuclear envelope rupture is induced by actin-based nucleus confinement.. The Journal of cell biology., 2016. [DOI | PubMed]
- Y Kalukula, AD Stephens, J Lammerding, S Gabriele. Mechanics and functional consequences of nuclear deformations.. Nat Rev Mol Cell Biol., 2022. [DOI | PubMed]
- Y Jung-Garcia, O Maiques, J Monger. LAP1 supports nuclear adaptability during constrained melanoma cell migration and invasion.. Nature cell biology., 2023. [DOI | PubMed]
- GR Kidiyoor, Q Li, G Bastianello. ATR is essential for preservation of cell mechanics and nuclear integrity during interstitial migration.. Nature Communications. 2020/09/24, 2020. [DOI]
- G Uzer, G Bas, B Sen. Sun-mediated mechanical LINC between nucleus and cytoskeleton regulates βcatenin nuclear access.. Journal of Biomechanics. 2018/06/06/, 2018. [DOI | PubMed]
- M Shokrollahi, M Stanic, A Hundal. DNA double-strand break-capturing nuclear envelope tubules drive DNA repair.. Nat Struct Mol Biol., 2024. [DOI]
- BE Danielsson, B George Abraham, E Mäntylä. Nuclear lamina strain states revealed by intermolecular force biosensor.. Nature Communications. 2023/06/30, 2023. [DOI]
- J Maciejowski, EM Hatch. Nuclear Membrane Rupture and Its Consequences.. Annual review of cell and developmental biology., 2020. [DOI]
- NY Chen, Y Yang, TA Weston. An absence of lamin B1 in migrating neurons causes nuclear membrane ruptures and cell death.. Proc Natl Acad Sci U S A., 2019. [DOI | PubMed]
- Y Kamikawa, A Saito, K Imaizumi. Impact of Nuclear Envelope Stress on Physiological and Pathological Processes in Central Nervous System.. Neurochemical research., 2022. [DOI | PubMed]
- CM Denais, RM Gilbert, P Isermann. Nuclear envelope rupture and repair during cancer cell migration.. Science., 2016. [DOI | PubMed]
- JD Vargas, EM Hatch, DJ Anderson, MW Hetzer. Transient nuclear envelope rupturing during interphase in human cancer cells.. Nucleus., 2012. [DOI | PubMed]
- M Raab, M Gentili, H de Belly. ESCRT III repairs nuclear envelope ruptures during cell migration to limit DNA damage and cell death.. Science., 2016. [DOI | PubMed]
- Z Dou, C Xu, G Donahue. Autophagy mediates degradation of nuclear lamina.. Nature., 2015. [DOI | PubMed]
- Z Yang, J Maciejowski, T de Lange. Nuclear Envelope Rupture Is Enhanced by Loss of p53 or Rb.. Mol Cancer Res., 2017. [DOI | PubMed]
- Q Zhang, AC Tamashunas, A Agrawal. Local, transient tensile stress on the nuclear membrane causes membrane rupture.. Molecular biology of the cell., 2019. [DOI | PubMed]
- CM de Noronha, MP Sherman, HW Lin. Dynamic disruptions in nuclear envelope architecture and integrity induced by HIV-1 Vpr.. Science., 2001. [DOI | PubMed]
- Z Dou, K Ghosh, MG Vizioli. Cytoplasmic chromatin triggers inflammation in senescence and cancer.. Nature., 2017. [DOI | PubMed]
- L Lindenboim, D Grozki, AR Amsalem-Zafran. Apoptotic stress induces Bax-dependent, caspase-independent redistribution of LINC complex nesprins.. Cell Death Discov., 2020. [DOI | PubMed]
- L Lindenboim, T Sasson, HJ Worman, C Borner, R Stein. Cellular stress induces Bax-regulated nuclear bubble budding and rupture followed by nuclear protein release.. Nucleus., 2014. [DOI | PubMed]
- M Pho, Y Berrada, A Gunda. Actin contraction controls nuclear blebbing and rupture independent of actin confinement.. Molecular biology of the cell., 2024. [DOI]
- CG Chu, N Lang, E Walsh. Lamin B loss in nuclear blebs is rupture dependent while increased DNA damage is rupture independent.. bioRxiv., 2025. [DOI]
- CT Halfmann, KL Scott, RM Sears, KJ Roux. Mechanisms by which barrier-to-autointegration factor regulates dynamics of nucleocytoplasmic leakage and membrane repair following nuclear envelope rupture.. bioRxiv., 2023. [DOI]
- WH De Vos, F Houben, M Kamps. Repetitive disruptions of the nuclear envelope invoke temporary loss of cellular compartmentalization in laminopathies.. Hum Mol Genet., 2011. [DOI | PubMed]
- A Karoutas, W Szymanski, T Rausch. The NSL complex maintains nuclear architecture stability via lamin A/C acetylation.. Nature cell biology., 2019. [DOI | PubMed]
- SM Schreiner, PK Koo, Y Zhao, SG Mochrie, MC King. The tethering of chromatin to the nuclear envelope supports nuclear mechanics.. Nat Commun., 2015. [DOI | PubMed]
- AD Stephens, EJ Banigan, JF Marko. Chromatin’s physical properties shape the nucleus and its functions.. Current opinion in cell biology., 2019. [DOI | PubMed]
- CT Halfmann, KJ Roux. Barrier-to-autointegration factor: a first responder for repair of nuclear ruptures.. Cell cycle (Georgetown, Tex)., 2021. [DOI | PubMed]
- MC King, CP Lusk, NR Ader. Sense, plug, and seal: proteins as both rapid responders and constitutive barriers supporting organelle compartmentalization.. Molecular biology of the cell., 2025. [DOI]
- AM Young, AL Gunn, EM Hatch. BAF facilitates interphase nuclear membrane repair through recruitment of nuclear transmembrane proteins.. Molecular biology of the cell., 2020. [DOI | PubMed]
- CT Halfmann, RM Sears, A Katiyar. Repair of nuclear ruptures requires barrier-to-autointegration factor.. The Journal of cell biology., 2019. [DOI | PubMed]
- DJ Thaller, D Tong, CJ Marklew. Direct binding of ESCRT protein Chm7 to phosphatidic acid-rich membranes at nuclear envelope herniations.. The Journal of cell biology., 2021. [DOI]
- Y Kamikawa, Z Wu, N Nakazawa, T Ito, A Saito, K Imaizumi. Impact of cell cycle on repair of ruptured nuclear envelope and sensitivity to nuclear envelope stress in glioblastoma.. Cell Death Discov., 2023. [DOI | PubMed]
- C Dias, J Nylandsted. Neural membrane repair at the core of regeneration.. Neural Regen Res., 2024. [DOI | PubMed]
- ST Cooper, PL McNeil. Membrane Repair: Mechanisms and Pathophysiology.. Physiol Rev., 2015. [DOI | PubMed]
- JB Dacks, MC Field. Evolutionary origins and specialisation of membrane transport.. Current opinion in cell biology., 2018. [DOI | PubMed]
- BK Hendricks, R Shi. Mechanisms of neuronal membrane sealing following mechanical trauma.. Neuroscience bulletin., 2014. [DOI | PubMed]
- ML Mencel, GD Bittner. Repair of traumatic lesions to the plasmalemma of neurons and other cells: Commonalities, conflicts, and controversies.. Front Physiol., 2023. [DOI | PubMed]
- C Dias, J Nylandsted. Plasma membrane integrity in health and disease: significance and therapeutic potential.. Cell Discov., 2021. [DOI | PubMed]
- S Xu, TJ Yang, S Xu, YN Gong. Plasma membrane repair empowers the necrotic survivors as innate immune modulators.. Semin Cell Dev Biol., 2024. [DOI | PubMed]
- J Yuan, D Ofengeim. A guide to cell death pathways.. Nature Reviews Molecular Cell Biology., 2023. [DOI]
- L Galluzzi, JM Bravo-San Pedro, I Vitale. Essential versus accessory aspects of cell death: recommendations of the NCCD 2015.. Cell Death Differ., 2015. [DOI | PubMed]
- G Kroemer, L Galluzzi, P Vandenabeele. Classification of cell death: recommendations of the Nomenclature Committee on Cell Death 2009.. Cell Death Differ., 2009. [DOI | PubMed]
- F Adhami, G Liao, YM Morozov. Cerebral ischemia-hypoxia induces intravascular coagulation and autophagy.. Am J Pathol., 2006. [DOI | PubMed]
- YM Morozov, P Rakic. Disorder of Golgi Apparatus Precedes Anoxia-Induced Pathology of Mitochondria.. International journal of molecular sciences., 2023. [DOI]
- YM Morozov, YY Sun, CY Kuan, P Rakic. Alteration of SLP2-like immunolabeling in mitochondria signifies early cellular damage in developing and adult mouse brain.. Eur J Neurosci., 2016. [DOI | PubMed]
- T Navaratnarajah, R Anand, AS Reichert, F Distelmaier. The relevance of mitochondrial morphology for human disease.. Int J Biochem Cell Biol., 2021. [DOI | PubMed]
- JR Friedman, J Nunnari. Mitochondrial form and function.. Nature., 2014. [DOI | PubMed]
- RJ Youle, AM van der Bliek. Mitochondrial fission, fusion, and stress.. Science., 2012. [DOI | PubMed]
- YM Morozov, D Datta, CD Paspalas, AFT Arnsten. Ultrastructural evidence for impaired mitochondrial fission in the aged rhesus monkey dorsolateral prefrontal cortex.. Neurobiol Aging., 2017. [DOI | PubMed]
- AM Bertholet, T Delerue, AM Millet. Mitochondrial fusion/fission dynamics in neurodegeneration and neuronal plasticity.. Neurobiology of disease., 2016. [DOI | PubMed]
- A Zimmer, AM Zimmer, AG Hohmann, M Herkenham, TI Bonner. Increased mortality, hypoactivity, and hypoalgesia in cannabinoid CB1 receptor knockout mice.. Proc Natl Acad Sci U S A., 1999. [DOI | PubMed]
- HR Bulgart, I Goncalves, N Weisleder. Leveraging Plasma Membrane Repair Therapeutics for Treating Neurodegenerative Diseases.. Cells., 2023. [DOI]
- P Tagliaferro, A Javier Ramos, ES Onaivi, SG Evrard, J Lujilde, A Brusco. Neuronal cytoskeleton and synaptic densities are altered after a chronic treatment with the cannabinoid receptor agonist WIN 55,212–2.. Brain Res., 2006. [DOI | PubMed]
- SJ Jackson, G Pryce, LT Diemel, ML Cuzner, D Baker. Cannabinoid-receptor 1 null mice are susceptible to neurofilament damage and caspase 3 activation.. Neuroscience. 2005/01/01/, 2005. [DOI | PubMed]
- D Soriano, M Vacotto, A Brusco, L Caltana. Neuronal and synaptic morphological alterations in the hippocampus of cannabinoid receptor type 1 knockout mice.. Journal of neuroscience research., 2020. [DOI | PubMed]
- XB Yuan, M Jin, X Xu. Signalling and crosstalk of Rho GTPases in mediating axon guidance.. Nature cell biology., 2003. [DOI | PubMed]
- A Hall. Rho family GTPases.. Biochemical Society transactions., 2012. [DOI | PubMed]
- AB Roland, A Ricobaraza, D Carrel. Cannabinoid-induced actomyosin contractility shapes neuronal morphology and growth.. eLife., 2014. [DOI | PubMed]
- A Mahomed, D Girn, A Pattani. Cannabinoid receptor type 1 regulates sequential stages of migration and morphogenesis of neural crest cells and derivatives in chicken and frog embryos.. J Morphol., 2023. [DOI | PubMed]
- G Mereu, M Fà, L Ferraro. Prenatal exposure to a cannabinoid agonist produces memory deficits linked to dysfunction in hippocampal long-term potentiation and glutamate release.. Proc Natl Acad Sci U S A., 2003. [DOI | PubMed]
- T Antonelli, MC Tomasini, M Tattoli. Prenatal exposure to the CB1 receptor agonist WIN 55,212–2 causes learning disruption associated with impaired cortical NMDA receptor function and emotional reactivity changes in rat offspring.. Cerebral cortex (New York, NY : 1991)., 2005. [DOI]
- KC Tree, M Scotto di Perretolo, J Peyronnet, F Cayetanot. In utero cannabinoid exposure alters breathing and the response to hypoxia in newborn mice.. Eur J Neurosci., 2014. [DOI | PubMed]
- E Fride. The endocannabinoid-CB(1) receptor system in pre- and postnatal life.. Eur J Pharmacol., 2004. [DOI | PubMed]
- J Mulder, T Aguado, E Keimpema. Endocannabinoid signaling controls pyramidal cell specification and long-range axon patterning.. Proc Natl Acad Sci U S A., 2008. [DOI | PubMed]
- J Diaz-Alonso, A de Salas-Quiroga, J Paraiso-Luna. Loss of Cannabinoid CB1 Receptors Induces Cortical Migration Malformations and Increases Seizure Susceptibility.. Cerebral cortex (New York, NY : 1991)., 2017. [DOI]
- SK Rouzer, M Domen, A George, A Bowring, RC Miranda. Early life outcomes of prenatal exposure to alcohol and synthetic cannabinoids in mice.. Drug Alcohol Depend Rep., 2025. [DOI | PubMed]
- A Sideris, J Lauzadis, M Kaczocha. The Basic Science of Cannabinoids.. Anesth Analg., 2024. [DOI | PubMed]
- R Sacco, GC Riccitelli, G Disanto. Effectiveness, Safety and Patients’ Satisfaction of Nabiximols (Sativex(®)) on Multiple Sclerosis Spasticity and Related Symptoms in a Swiss Multicenter Study.. J Clin Med., 2024. [DOI]
- RA Gray, BJ Whalley. The proposed mechanisms of action of CBD in epilepsy.. Epileptic Disord., 2020. [DOI | PubMed]
- A Gil-Ordóñez, M Martín-Fontecha, S Ortega-Gutiérrez, ML López-Rodríguez. Monoacylglycerol lipase (MAGL) as a promising therapeutic target.. Biochemical pharmacology., 2018. [DOI | PubMed]
- RW de Oliveira, CL Oliveira, FS Guimaraes, AC Campos. Cannabinoid signalling in embryonic and adult neurogenesis: possible implications for psychiatric and neurological disorders.. Acta neuropsychiatrica., 2018. [DOI]
- C Chen. Inhibiting degradation of 2-arachidonoylglycerol as a therapeutic strategy for neurodegenerative diseases.. Pharmacol Ther., 2023. [DOI | PubMed]
- A Zanfirescu, A Ungurianu, DP Mihai, D Radulescu, GM Nitulescu. Targeting Monoacylglycerol Lipase in Pursuit of Therapies for Neurological and Neurodegenerative Diseases.. Molecules., 2021. [DOI]
- N Pasquarelli, C Porazik, H Bayer. Contrasting effects of selective MAGL and FAAH inhibition on dopamine depletion and GDNF expression in a chronic MPTP mouse model of Parkinson’s disease.. Neurochemistry International. 2017/11/01/, 2017. [DOI | PubMed]
- LD Schurman, AH Lichtman. Endocannabinoids: A Promising Impact for Traumatic Brain Injury. Review.. Frontiers in Pharmacology. 2017-February-17, 2017. [DOI]
- D Panikashvili, C Simeonidou, S Ben-Shabat. An endogenous cannabinoid (2-AG) is neuroprotective after brain injury.. Nature., 2001. [DOI | PubMed]
- M Guzmán, MJ Duarte, C Blázquez. A pilot clinical study of Δ9-tetrahydrocannabinol in patients with recurrent glioblastoma multiforme.. British Journal of Cancer. 2006/07/01, 2006. [DOI | PubMed]
- M Zaiachuk, N Pryimak, O Kovalchuk, I Kovalchuk. Cannabinoids, Medical Cannabis, and Colorectal Cancer Immunotherapy. Review.. Frontiers in Medicine. 2021-September-24, 2021. [DOI]
- PP Bathula, MB Maciver. Cannabinoids in Treating Chemotherapy-Induced Nausea and Vomiting, Cancer-Associated Pain, and Tumor Growth.. International journal of molecular sciences., 2024
- C Pagano, G Navarra, L Coppola, G Avilia, M Bifulco, C Laezza. Cannabinoids: Therapeutic Use in Clinical Practice.. International journal of molecular sciences., 2022. [PubMed]
- NM Pennant, CV Hinton. The evolution of cannabinoid receptors in cancer.. WIREs Mech Dis., 2023. [DOI | PubMed]
- G Bononi, M Di Stefano, G Poli. Reversible Monoacylglycerol Lipase Inhibitors: Discovery of a New Class of Benzylpiperidine Derivatives.. J Med Chem., 2022. [DOI | PubMed]
- G Nahler. Treatment of malignant diseases with phytocannabinoids: promising observations in animal models and patients.. Exploration of Medicine., 2023. [DOI]
- J Guggisberg, M Schumacher, G Gilmore, DM Zylla. Cannabis as an Anticancer Agent: A Review of Clinical Data and Assessment of Case Reports.. Cannabis Cannabinoid Res., 2022. [DOI | PubMed]
- F Afrin, M Chi, AL Eamens. Can Hemp Help? Low-THC Cannabis and Non-THC Cannabinoids for the Treatment of Cancer.. Cancers (Basel)., 2020. [DOI]
- CJ Fowler. Delta(9) -tetrahydrocannabinol and cannabidiol as potential curative agents for cancer: A critical examination of the preclinical literature.. Clin Pharmacol Ther., 2015. [DOI | PubMed]