
Comparative Analysis of Cannabidiol and Risperidone on Behavioral and Neurochemical Outcomes, and Neurodevelopment Markers in a Zebrafish Model of Embryonic Exposure to Sodium Valproate
Abstract
Autism spectrum disorder (ASD) is a neurodevelopmental condition characterized by social communication deficits, repetitive behaviors, and sensory abnormalities. Sodium valproate (VPA) exposure during embryonic development is a well‐established preclinical model for ASD, leading to increased oxidative stress in the developing brain, including lipid peroxidation, which affects cell proliferation and organization. This study aimed to investigate the potential therapeutic effects of cannabidiol (CBD) and risperidone (RISP) in reversing ASD‐like behaviors and associated neurobiological alterations induced by embryonic VPA exposure in a zebrafish model. Zebrafish embryos were exposed to 125 μM VPA for 2 days post‐fertilization (dpf). At 3–4 dpf, embryos were treated with 0.06 μM CBD or 1 μM RISP. Behavioral assays were conducted to assess hyperlocomotion and aggressive behavior. At 7 dpf, lipid peroxidation levels were measured, and expression of glial fibrillary acidic protein (GFAP) and calcium/calmodulin (CaM) were analyzed to evaluate neurobiological changes. VPA exposure resulted in increased hyperlocomotion and aggression. CBD treatment effectively reversed these behaviors, while RISP showed limited efficacy. Additionally, CBD reduced lipid peroxidation and restored anandamide levels, whereas RISP did not exhibit these effects. CBD also normalized GFAP and CaM expression, indicating restoration of glial function and excitatory/inhibitory balance. CBD demonstrated a better efficacy and safety profile compared to RISP in reversing ASD‐like behaviors and associated neurobiological alterations in the zebrafish model. These findings suggest that CBD may offer a safer and more effective therapeutic alternative for managing ASD‐related symptoms.
Article type: Research Article
Keywords: autism spectrum disorder, cannabidiol, endocannabinoids, lipid peroxidation, risperidone, zebrafish
Affiliations: Department of Pharmacology Ribeirão Preto Medical School, University of São Paulo Ribeirão Preto SP Brazil; Center for Cannabinoid Research, Mental Health Building Ribeirão Preto Medical School, University of São Paulo Ribeirão Preto SP Brazil; Department of Clinical Analyses, Toxicology and Food Science School of Pharmaceutical Sciences of Ribeirão Preto, University of São Paulo São Paulo SP Brazil; National Institute of Science and Technology for Translational Medicine (INCT TM), CNPQ/FAPESP/CAPES Ribeirão Preto SP Brazil; Department of Neuroscience and Behavior Ribeirão Preto Medical School, University of São Paulo São Paulo SP Brazil; Laboratory of Psychiatric Neuroimaging (LIM21) Hospital das Clínicas da Faculdade de Medicina da Universidade de Sao Paulo (HCFMUSP) São Paulo SP Brazil
License: © 2025 The Author(s). Autism Research published by International Society for Autism Research and Wiley Periodicals LLC. CC BY 4.0 This is an open access article under the terms of the http://creativecommons.org/licenses/by/4.0/ License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Article links: DOI: 10.1002/aur.70151 | PubMed: 41293963 | PMC: PMC12729503
Relevance: Relevant: mentioned in keywords or abstract
Full text: PDF (5.4 MB)
Introduction
Autism spectrum disorder (ASD) is a complex neurodevelopmental condition marked by a wide range of clinical manifestations and severity. Symptoms include deficits in social–emotional reciprocity, executive function impairments, communication difficulties, and sensory processing challenges (Lord et al. ref. 2018). Recent data report that ASD affects about 1 in 36 children, being 4.3 times more prevalent in males (Maenner et al. ref. 2023). Additionally, ASD is often accompanied by intellectual disability, aggression, hyperactivity, social anxiety, epilepsy, sleep disturbances, and gastrointestinal symptoms (Lord et al. ref. 2018; Pedrazzi et al. ref. 2022).
Besides genetic factors and abnormal brain development during critical periods (LeBlanc and Gillis ref. 2012; Medina et al. ref. 2023), environmental stressors, infections, and teratogenic drugs, including sodium valproate (VPA), are recognized as risk factors (Karimi et al. ref. 2017) for ASD (Christensen et al. ref. 2013). Prenatal VPA exposure is a well‐established model for studying ASD‐like features in rodents (Schneider and Przewłocki ref. 2005; Wagner et al. ref. 2006), due to its effects on oxidative stress balance, histone acetylation, serotonin turnover, and excitatory/inhibitory balance during neurodevelopment (Fereshetyan et al. ref. 2021; Mabunga et al. ref. 2015; Nicolini and Fahnestock ref. 2018).
Despite major advances in the understanding of factors that might increase vulnerability to ASD, the pharmacological treatment remains suboptimal, focused on the relief of specific symptoms (Aishworiya et al. ref. 2022). In this context, antipsychotic drugs such as risperidone (RISP) and aripiprazole are the only FDA‐approved medications for ASD patients, but they exhibit limited efficacy depending on the severity of the ASD as reported across clinical studies (Broadstock et al. ref. 2007; Murray et al. ref. 2015).
Recently, CBD, the main non‐psychotomimetic phytocannabinoid derived from Cannabis sativa, has shown promise in improving neuropsychiatric symptoms in ASD patients (Hacohen et al. ref. 2022; Pretzsch et al. ref. 2019; Silva Jr et al. ref. 2024) and cognitive function in mouse models of ASD (Pedrazzi et al. ref. 2025). Additional studies also support the efficacy of CBD‐enriched extracts in enhancing sociability in individuals with ASD (Barchel et al. ref. 2019; Bilge and Ekici ref. 2021; Silva Jr et al. ref. 2024). However, the characterization of the mechanism associated with CBD effects remains poorly understood in the context of ASD.
In this study, we developed a rapid and reproducible platform for ASD drug screening by inducing persistent ASD‐like traits through embryonic exposure to VPA in zebrafish. Indeed, different zebrafish VPA‐ASD study models have been developed through different exposure methods, evaluating changes in the molecular, proteomic, cellular, and behavioral patterns (for an extensive review, please read Camussi et al. ref. 2025). Additionally, we explore possible mechanisms associated with the long‐term behavioral, neurochemical, and neurodevelopmental effects of CBD and RISP during the larval stages.
Material and Methods
Animals
Wild type zebrafish (Danio rerio) embryos and larvae were obtained from the facility at the School of Pharmaceutical Science of Ribeirão Preto, University of São Paulo (Ribeirão Preto, Brazil). The adults were kept in a ZebTEC (Tecniplast, Italy) recirculating system using water obtained by a reverse osmosis system at standard conditions (pH 7.5 ± 0.5, temperature 26°C ± 1°C, dissolved oxygen at 95% saturation, and conductivity 750 ± 50 μS/cm, 14 h:10 h light/dark cycle). The experimental protocols were approved by the Ethics Committee on Animal Use at the University of São Paulo (CEUA/FCFRP no. 19.1.243.60.4). Fertilized eggs obtained by placing male and female breeding stock in a 2:1 ratio in a 1 L aquarium, were washed using freshly prepared embryo medium (2 mM CaCl2⋅2H2O, 0.5 mM MgSO4⋅7H2O, 0.75 mM NaHCO3, and 0.07 mM KCl) and observed under a stereomicroscope (Zoom Microscope SMZ 1500, Nikon), and those in the cleavage stage (4–64 cells) were randomly selected for experiments. During the first 120 h post‐fertilization (hpf), the larvae feed from their yolk sac, eliminating the need for additional feeding during the experiments. At 6 days post‐fertilization (dpf), external feeding was offered (microflocculated larval food; Alcon). Before the experimental assays, larvae were fasted overnight.
Drugs
Sodium valproate (VPA; Prati‐Donaduzzi, Brazil), RISP (Prati‐Donaduzzi, Brazil) freshly prepared in stock solutions of VPA (12.5 mM), and RISP (0.03 mM) using ultrapure water were used. The stock solution of CBD (99.9% pure CBD, BSPG Pharm, Brentwood, UK) (9.54 mM) was prepared in DMSO. To avoid DMSO‐induced toxicity, all experiments were performed with DMSO diluted at 0.01% (v/v) (ISO 7346‐3, 1996) (Brigante et al. ref. 2018).
Experimental Procedures
Embryonic Exposure to VPA
Fish embryo toxicity (FET) test was conducted (OECD ref. 2025) to evaluate the impacts of the VPA during development. For that, eggs were randomly and individually placed in 24‐well plates, containing 2.0 mL of tank water + each VPA concentration (5, 25, 125, 625, and 1250 μM). Exposure of VPA occurred for 48 h; lethality (egg coagulation, non‐detached tail, absence of somites, and no heartbeat), sublethality (eye development, spontaneous movements, blood circulation, pigmentation, and swim bladder inflation), and teratogenicity (malformed head, otoliths, tail, or heart; scoliosis; yolk sac deformity; and altered growth) were evaluated daily (Nagel ref. 2002) using a stereomicroscope (Stemi 508, Carl Zeiss, Germany) equipped with a camera (AxioCam ICc 5, Zeiss, Germany).
To assess effects on spontaneous embryonic movement, the tail coiling activity test was accomplished as previously reported (de Oliveira et al. ref. 2021). Tests were conducted in triplicate. At 24 hpf, non‐hatched embryos from each group exposed to VPA (5, 25, 125, 625, and 1250 μM) were grouped on an excavated slide and placed under a stereomicroscope (Stemi 508, Zeiss, 0.63). After a 5‐min acclimation period, tail coiling behavior was recorded for 3 min using Zen 2.3 Image software (Zeiss) and videos recorded were processed using DanioScope software (Noldus), which computed the burst activity frequency, defined as the percentage of time the embryo exhibited spontaneous movement (n = 20 per group, three independent replicates).
Behavioral Procedures
Experiment 1: Standardization of VPA Model
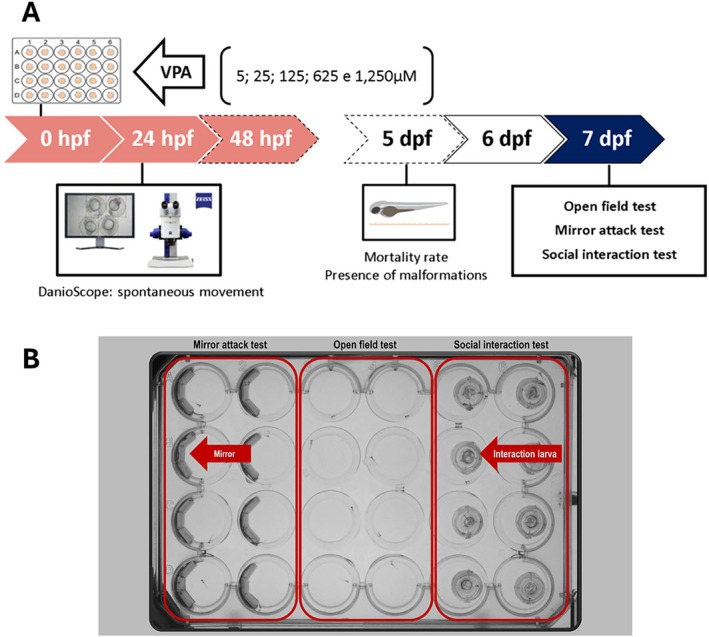
Experiment 1 aimed to determine the optimal working concentration of VPA that reliably induces neurodevelopmental changes in a zebrafish model for ASD without causing teratogenicity or lethality. After the embryonic VPA exposure (Figure 1A, see Section 2.3.1 for details), on the 7th day post‐fertilization (dpf), independent groups of zebrafish larvae were subjected to the following behavioral tests: open field (for motor activity and swimming coordination), the mirror attack test (for assessing aggressive behavior), and the social interaction (SI) test (to measure social behavior) (Figure 1B). At the end of the analysis, the larvae were euthanized under deep anesthesia.
Experiment 2: Characterization of theBehavioral Effects of Different CBD Concentrations in the Zebrafish Model of VPA‐Induced Behavioral Changes Following Embryonic Exposure
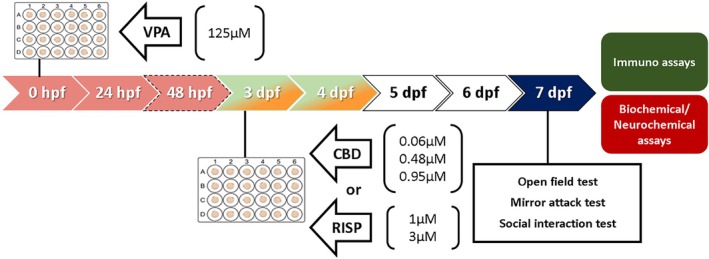
Once the VPA model was set (work concentration of 125 μM), we tested if the treatments could reverse the behavioral effects of VPA embryonic exposure (first 48 hpf). Then, on 3–4 dpf, larvae were exposed to different concentrations of vehicle (negative control), CBD, or RISP (positive control) (Figure 2). After a 2‐day drug washout period, independent groups of larvae were subjected to behavioral tests (for further details, see Figure 2 caption). At the end of the experiments, the larvae were euthanized and prepared for biochemical assays, neurotransmitter measurements, or immunofluorescence protocols (for further details, see Figure 2).
Behavioral Tests
All behavioral tests were conducted in triplicate during the dark cycle (n = 20–24 larvae per group).
Open Field Test (OFT)
Each larva was individually transferred to a new well in a modified 4–24‐well plate (Figure 1B), filled with 2 mL of tank water. The larvae were given 15 min to freely explore the well, consisting of a 5‐min habituation period followed by a 10‐min testing phase. The total distance traveled during the final 10 min of the test was recorded and analyzed automatically using the Zebrabox (View Point Life Sciences, France) (Dwivedi et al. ref. 2019).
Mirror Attack Test
This test was used to assess aggressive behavior. The wells of the modified 4–24‐well plate were filled with 2 mL of tank water and had half of their walls lined with a semicircular mirror (Figure 1B), designed to simulate the presence of another fish in the well. The larvae were allowed to freely explore the well with the first 5 min serving as a habituation period, followed by a 10‐min test. During the test, the number of attacks/bites directed toward the mirror and the total time spent in the mirror area were recorded and analyzed (Pham et al. ref. 2012).
SI Test
SI was assessed by placing a larva in a modified 4–24‐well plate filled with 2 mL of tank water. The well was divided into two semicircles, each with a diameter of 6.6 mm, using an adapted transparent slide that allowed each larva to visualize its counterpart in the adjacent semicircle (Figure 1B). The test consisted of a 5‐min habituation period, followed by a 10‐min testing phase. During the test, the time spent in the interaction zone, defined as the area of the well corresponding to a length, was analyzed automatically using the Zebrabox (View Point Life Sciences, France).
Biochemical Assays
At the end of behavioral experiments, larvae were euthanized in liquid nitrogen (hypothermia) and stored at −80°C. Larval samples were subjected to ultrasonic homogenization with 1.5 mL of 0.1 M KPBS (pH 7.4). The homogenate was then equally divided into two aliquots: one designated for lipid peroxidation (LPO) analysis and the other for mitochondrial isolation. To isolate the mitochondria, the samples were centrifuged at 10,000 g for 20 min at 4°C, and catalase activity (CAT), glutathione‐S‐transferase (GST) activity, and protein quantification were assessed. All biochemical assays were conducted using a microplate spectrophotometer (Multiskan‐Sky) along with SkanIt software (version 6.0.2, Thermo Scientific, USA) to ensure accurate measurements and data analysis. The results of the biochemical assays were normalized to the total protein concentration, quantified by Bradford assay (Bradford ref. 1976).
GST Activity
The GST activity was continuously monitored for 30 min using a microplate spectrophotometer set to a wavelength of 340 nm by measuring the reaction between reduced glutathione and 1‐chloro‐2,4‐dinitrobenzene (CDNB). The enzymatic activity was normalized to the total protein content and expressed as nmol/min/mg of protein (Habig et al. ref. 1974).
CAT
The enzymatic CAT was continuously monitored for 2 min by measuring the decrease in absorbance at a wavelength of 240 nm, corresponding to the consumption of hydrogen peroxide (H2O2). Similar to the procedure for GST, the CAT was normalized to the total protein content and expressed in appropriate units (Claiborne ref. 1985).
Lipid Oxidation Quantification
For the LPO assay, the homogenate was treated with 2,6‐di‐tert‐butyl‐4‐methylphenol at a concentration of 4% (v/v), diluted in methanol, to prevent further oxidation. The endogenous LPO products were quantified as reactive substances reacting with thiobarbituric acid (TBARS), expressed as pmol per larva. The analysis was conducted at a wavelength of 535 nm (Bird and Draper ref. 1984; Ohkawa et al. ref. 1979) and normalized by total protein.
Neurochemical Analyses by ELISA
To assess the levels of serotonin (5‐HT), dopamine (DA), anandamide (AEA), and 2‐arachidonoylglycerol (2‐AG) in the larvae, three clusters of 30 zebrafish larvae at 7 dpf were collected for each experimental group (n = 3 replicates per pool of 30 larvae). Quantifications were performed using commercial ELISA: Serotonin/5‐Hydroxytryptamine (ST/5‐HT) ELISA Kit (Cat. No. MBS2513872), Fish Dopamine (DA) ELISA Kit (Cat. No. MBS912178), General Anandamide (AEA) ELISA Kit (Cat. No. MBS2097802), and General 2‐Arachidonoylglycerol ELISA Kit (Cat. No. MBS2031975), following the manufacturer’s instructions (kits from MyBioSource, USA).
Immunofluorescence Assay and Microscopy
Intact larvae from behavioral assays were euthanized by hypothermia and fixed overnight at 4°C in 4% paraformaldehyde (PFA) with gentle agitation (300 rpm). The next day, samples were washed three times for 5 min each in PBS (0.01 M, pH 7.4), then dehydrated through graded methanol (25%, 50%, 75%, 100%, 5 min each) and stored at −20°C in 100% methanol. For the staining procedure, larvae were washed three times (5 min each) in 1% PBS‐Triton and gradually rehydrated in 75%, 50%, and 25% methanol in PBS (5 min each), followed by three additional washes in 1% PBS‐Triton. Depigmentation was performed by incubating larvae for 5 min in 3% hydrogen peroxide and 1% potassium hydroxide in PBS. Antigen retrieval was carried out with Proteinase K (10 μg/mL) for 40 min, followed by three washes in 1% PBS‐Triton. The blocking step was performed for 2 h at room temperature in 5% fetal bovine serum, 4 mg/mL bovine serum albumin, 1% Triton X‐100, and 1% DMSO in PBS. Larvae were then incubated overnight at 4°C with primary antibodies diluted in blocking solution (see Table S1 in Supporting Information), followed by six 30‐min washes in 1% PBS‐Triton. Secondary antibody incubation was performed overnight at 4°C under gentle agitation (see Table S1 in Supporting Information). The next day, larvae were incubated for 1 h in Hoechst (1:10,000, Invitrogen) diluted in 1% PBS‐Triton. After the final washes, larvae were mounted on glass slides using Fluoromount‐G (Invitrogen) and coverslipped. Imaging was performed using multiphoton microscopy (LSM 780 Axio Observer, Carl Zeiss) at 40× magnification (MOSAIC mode), with images acquired in z‐stacks and analyzed with Zen 3.5 software (Carl Zeiss). Quantification was conducted on the total brain area, and results were expressed as relative fluorescence intensity (FI) for each immunostaining (n = 2–6 larvae/group).
Statistical Analyses
When the assumptions of normality and homoscedasticity were met, significant differences between treatments were assessed using one‐way ANOVA or two‐way ANOVA, followed by Duncan’s post hoc test. Data are presented as the mean ± standard error of the mean (SEM). For nonparametric data, significant differences were evaluated using the Kruskal–Wallis test, followed by Dunn’s post hoc test, with results expressed as median and interquartile ranges. Statistical significance was defined as p ≤ 0.05.
Results
Embryonic Exposure to VPA in Zebrafish Induces Long‐Term Behavioral Changes Associated With ASD
In rodents, embryonic VPA exposure induces social deficits, altered exploration, impaired motor coordination, and aggression. To investigate if similar patterns occur in zebrafish, we characterized the effects of VPA during early development by continuous observation from early stages to larvae in the same individuals, a clear experimental advantage over the rodent model. Exposure to different VPA concentrations (5, 25, 125, 625, 1250 μM) for 24 h significantly altered tail coiling behavior (χ 2 = 125.81; p < 0.001, Figure 3B). Specifically, 125 μM (p < 0.05) and 625 μM (p < 0.01) increased spontaneous movement, while 1250 μM of VPA reduced this behavior (p < 0.001). Higher VPA concentrations (625 and 1250 μM) also increased malformations (F 5,12 = 346.12; p < 0.001, Figure 3D) and mortality rates (F 5,12 = 358.53; p < 0.001, Figure 3C), with 100% lethality at the concentration of 1250 μM. The VPA concentrations of 5, 25, and 125 μM did not induce malformation and mortality (Figure 3C,D) throughout the experimental window, and therefore, were selected to continue the experiments.
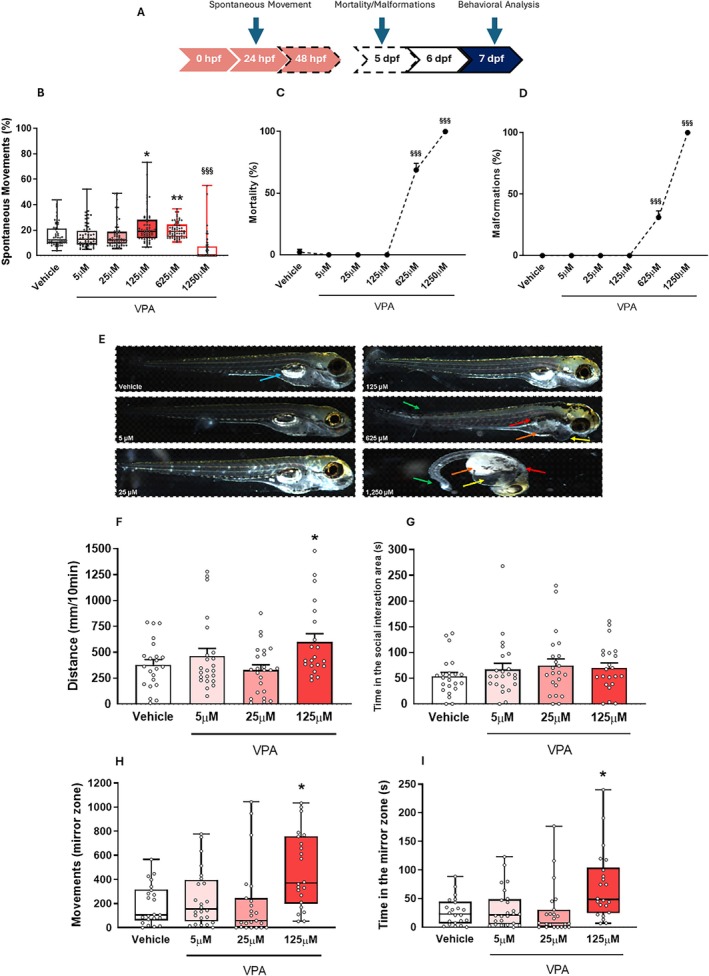
To assess the long‐term effects of non‐teratogenic concentrations of VPA (5, 25, 125 μM), we tested 7 dpf larvae in behavioral paradigms: mirror attack (aggression), SI, and open field (locomotor activity) (Figure 3F–I). In the OFT, 125 μM VPA larvae showed increased locomotor activity compared to controls (F 3,86 = 3.42; p < 0.05, Figure 3F). Larvae exposed to 125 μM VPA also exhibited increased movements (χ 2 = 16.02; p < 0.01, Figure 3H) and time (χ 2 = 15.82; p < 0.01, Figure 3I) in the mirror area, indicating increased aggressive behavior. No significant differences were found in SI (F 3,86 = 0.71; p > 0.05, Figure 3G). Based on these results, 125 μM was selected for subsequent experiments for testing drug efficacy.
CBD is More Potent and Safer Than RISP in Reversing VPA Embryonic Exposure‐Induced ASD‐Like Behaviors in Zebrafish Larvae
RISP is commonly prescribed for managing ASD symptoms. Therefore, as a positive control for our model, we tested whether RISP (1 and 3 μM) could counteract behavioral changes induced by embryonic VPA (125 μM) exposure in zebrafish larvae. Our results suggest that RISP at a concentration of 1 μM effectively reversed ASD‐like behaviors (at 7 dpf) induced by VPA exposure (Figure S1 in Supporting Information).
Next, given CBD’s broad action in neurodevelopmental models, we evaluated CBD at 0.06, 0.48, and 0.95 μM alongside a vehicle (0.01% DMSO) and RISP (1 μM, Figure 4A). In the mirror attack test, both VPA‐exposed groups showed an increased number of movements (F 1,205 = 6.51; p < 0.05) and time spent in the mirror area (F 1,205 = 5.78; p < 0.05) compared to vehicle‐treated groups (Figure 4B,C). Treatment with CBD (0.06 μM) or RISP (1 μM) significantly reduced these parameters compared to untreated VPA groups (since CBD and RISP have different vehicles—DMSO or saline, respectively) (F 5,120 = 2.27; p < 0.05). No significant difference between CBD and RISP was detected (Figure 4B,C).
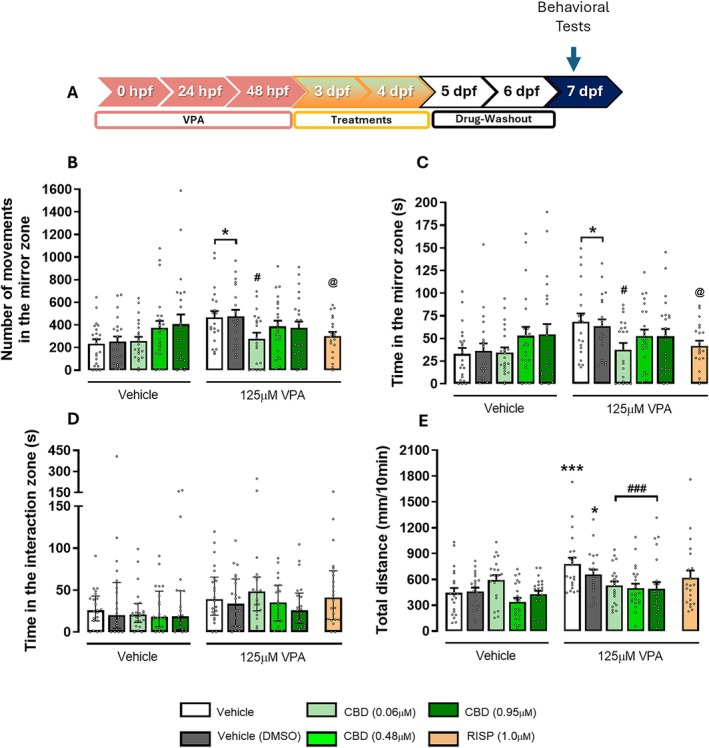
To determine if any CBD concentrations affect locomotor activity in 7 dpf zebrafish larvae exposed to 125 μM VPA during the embryonic period, larvae were tested in the OFT (Figure 4E). Both VPA‐exposed non‐treated groups (saline or 0.01% DMSO) had significantly higher locomotor activity than the vehicle control group (F 4,205 = 3.58; p < 0.01). In contrast, VPA‐exposed larvae treated with all the CBD concentrations showed a significant reduction in total distance compared to the untreated VPA group (p < 0.001, Figure 4E). No effects on the SI test were observed (χ 2 = 12.77, Figure 4D).
During the collection of larval samples in plastic vials filled with PBS following euthanasia, we made an interesting observation: the larvae treated with RISP (independent of the stimulus) tended to settle at the bottom of the vial, which was distinctly different from the behavior of larvae in the vehicle and CBD treatment groups. A close morphological analysis revealed that RISP induced notable changes in the swim bladder development in these larvae (Figure S2 in Supporting Information).
Given that CBD and RISP reversed the behavioral effects induced by VPA at different concentrations (0.06 μM for CBD and 1 μM for RISP), we conclude that, in this model, CBD is more potent than RISP in reversing VPA‐induced ASD‐like behaviors in zebrafish larvae at 7 dpf. Additionally, given the close relationship between the swim bladder and the primitive lung, we propose that CBD exhibits a safer profile than RISP in our zebrafish model.
CBD Reduces LPO in Zebrafish: A Potential Mechanism Underlying Its Neuroprotective and Behavioral Effects
Although observational studies suggest benefits of CBD on ASD symptoms, its molecular mechanisms remain unclear. To explore this, we analyzed the antioxidant system and neurochemical profiles in zebrafish brains using the lowest effective CBD concentration (0.06 μM) and compared its effects with RISP (Figure 5A). Analysis of CAT (Figure 5B) and GST activity (Figure 5C) showed no significant changes after VPA exposure or RISP/CBD treatments. Conversely, LPO levels, assessed via TBARS, were significantly reduced by CBD treatment, indicating a protective antioxidant effect, regardless of VPA exposure (F 2,30 = 5.33; p < 0.05, Figure 5D).
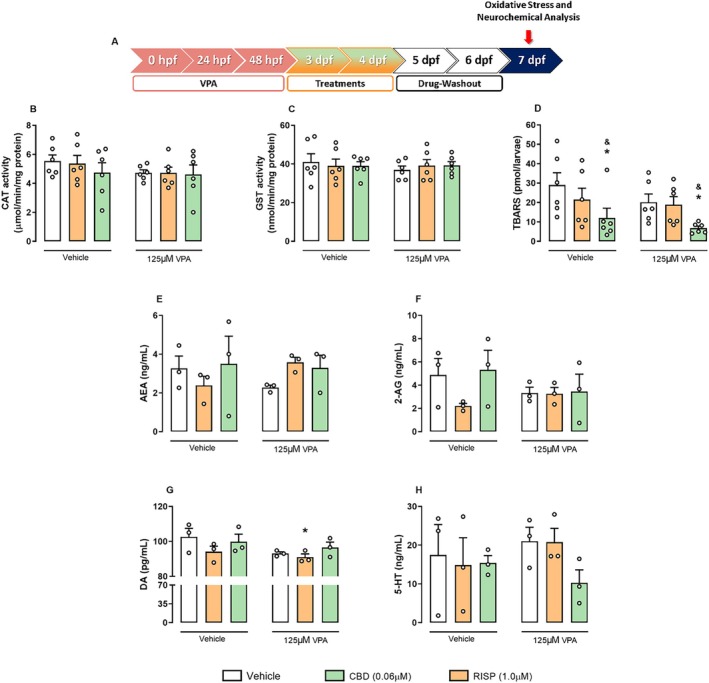
We also evaluated the levels of the neurotransmitters, DA and 5‐HT, and the endocannabinoids, 2‐AG and AEA. Although VPA treatment showed a graphical trend in reducing AEA (F 2,12 = 2.8; p = 0.10, Figure 5E) and 2‐AG (F 2,12 = 1.01; p > 0.05, Figure 5F) levels, they remained unchanged across groups. DA levels (F 1,12 = 3.09; p > 0.05, Figure 5G) were significantly reduced in VPA‐exposed larvae treated with RISP compared to controls (p < 0.05). However, 5‐HT levels (F 2,12 = 0.66; p > 0.05, Figure 5H) showed no significant changes after VPA exposure or RISP/CBD treatment.
The Embryonic Exposure to VPA Induces Complex Changes in the Neurodevelopment Markers in Zebrafish Larvae: CBD’S and RISP’S Effects
Neurodevelopment relies on fine‐tuned cellular regulation, and any disruptive processes may contribute to ASD (Satterstrom et al. ref. 2020). Here, we evaluated key neurodevelopmental markers at 7 dpf, including proliferation (pHH3, PCNA) and apoptosis (Caspase‐3) markers, GFAP expression (astrocyte, radial glial, and possibly neural stem cells), and the calcium signaling proteins, CaM and PV (Figure 6A).
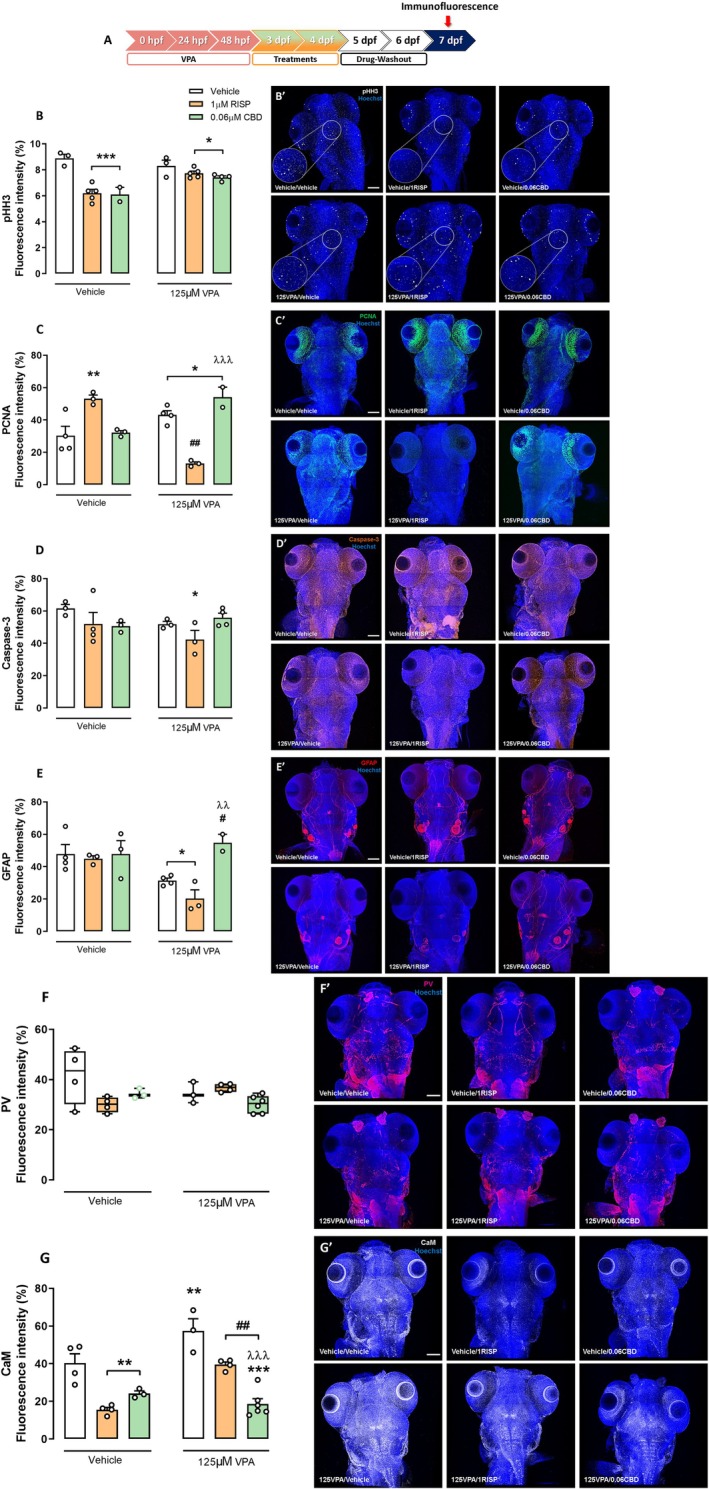
For pHH3, both VPA‐exposed and control larvae treated with 1 μM RISP or 0.06 μM CBD showed a significant reduction in FI compared to controls (F 2,16 = 21.29; p < 0.001; Figure 6B). VPA exposure tended to increase PCNA expression (Figure 6C), with 1 μM RISP increasing PCNA in non‐VPA larvae, but decreasing it in VPA‐exposed ones (F 2,13 = 35.60; p < 0.001). CBD increased PCNA in VPA‐exposed larvae (p < 0.05). RISP reduced Caspase‐3 expression in VPA‐exposed larvae (p < 0.05; Figure 6D). GFAP analysis revealed reduced expression in VPA‐exposed larvae treated with vehicle or RISP compared to controls (F 1,13 = 6.95; p < 0.05). CBD restored GFAP levels compared to VPA‐non‐treated (p < 0.05) and VPA‐RISP‐treated groups (p < 0.01) (Figure 6F).
The calcium signaling/binding proteins (CAM and PV) analyses revealed that VPA embryonic exposure increased CaM expression at 7 dpf, which was reversed by RISP and CBD treatment after drug washout (F 2,18 = 10.17; p < 0.01; Figure 6G). Both drugs reduced CaM in controls. No significant changes were detected for PV expression (χ 2 = 10.93; p > 0.05; Figure 6F). These findings suggest that CBD corrects the excitatory imbalance and astrocyte dysfunction caused by VPA exposure.
Discussion
Our study, in line with previous work (for an extensive review, please read Camussi et al. ref. 2025), demonstrates that embryonic exposure of zebrafish to 125 μM VPA produces long‐term behavioral phenotypes consistent with ASD, validating a fast and reproducible model for rapid drug screening in ASD research. Treatment with the lowest evaluated concentration of CBD (0.06 μM) effectively reversed these behavioral changes, proving to be more potent and safer than RISP (1 μM), a standard treatment for ASD. Importantly, we also begin to uncover a possible mechanistic basis for CBD’s effects, linked to reductions in LPO and changes in markers of neurodevelopment.
The VPA exposure during early zebrafish development replicates several core and associated behavioral and neurochemical features of ASD, consistent with findings in rodent models (Kim et al. ref. 2011; Schneider and Przewłocki ref. 2005). Zebrafish embryos exposed to VPA showed dose‐dependent changes in spontaneous tail coiling, increased malformation rates (e.g., non‐inflated swim bladders and edema), and heightened mortality at higher concentrations. These results align with previous studies using VPA to model ASD‐like symptoms, such as hyperactivity and aggression (Chen et al. ref. 2020; Christensen et al. ref. 2013; Meshalkina et al. ref. 2018). An advantage of the zebrafish model compared to rodents is the ability to observe the embryonic effects of VPA in real time, thanks to external development and egg transparency. Moreover, zebrafish embryos hatch within hours, whereas rodent gestation requires approximately 21 days. Embryos exposed to 125 μM VPA from 4 to 48 hpf exhibited persistent hyperactivity and aggression even after exposure cessation, suggesting long‐lasting neurodevelopmental consequences. Notably, this exposure did not affect survival, malformations, or hatching time, but led to increased spontaneous movement, motor activity, and aggressive‐like behavior at 7 dpf (Dwivedi et al. ref. 2019; Zimmermann et al. ref. 2015). These findings support the well‐documented teratogenic and neurodevelopmental impacts of VPA in humans and rodents (Choi et al. ref. 2016; Ornoy et al. ref. 2023) and further validate the zebrafish as a sensitive and translational model for studying early developmental disturbances relevant to ASD.
Our model also reproduces the long‐term behavioral effects of VPA. Larvae exposed to 125 μM VPA showed increased aggressive behavior, as evidenced by enhanced mirror attack responses, and increased locomotor activity in the OFT—two phenotypes previously associated with hyperactivity, impulsivity, and social dysregulation (Mabunga et al. ref. 2015). Interestingly, no significant changes were observed in the SI test at 7 dpf, suggesting that deficits in social preference may emerge later in development or that our assay conditions lacked the sensitivity to detect subtle impairments at this early stage in the zebrafish larva. The absence of VPA’s effects on SI may be associated with the immature state of social behavior regulation at this developmental point in zebrafish larvae (Buske and Gerlai ref. 2012). However, previous studies suggested that the mirror attack test also has a good face of validity while studying social behaviors (Desjardins and Fernald ref. 2010; Moretz et al. ref. 2007). Together, these results indicate that early exposure to 125 μM VPA reliably induces ASD‐relevant phenotypes without compromising overall survival or development, supporting its use as an optimal concentration for further pharmacological testing.
Although a few studies have reported the protective effects of CBD in rodent models of ASD, and on certain neuropsychiatric symptoms in humans, to the best of our knowledge, CBD has not been tested in ASD models that include positive control groups, such as treatment with antipsychotic drugs (Pedrazzi et al. ref. 2025; Silva Jr et al. ref. 2024). When comparing therapeutic interventions, we found that CBD at a low concentration (0.06 μM) was more effective than RISP (1 μM) in reversing VPA‐induced aggressive behavior and hyperlocomotion. Importantly, larvae treated with RISP displayed swim bladder abnormalities, suggesting potential developmental side effects—a finding consistent with prior reports of RISP’s adverse impacts on growth and organogenesis (Fukushima et al. ref. 2023). Given that individuals with ASD often have communication difficulties or swallowing dysfunctions, they may be at a higher risk of aspiration pneumonia—a complication previously linked to RISP treatment (Wang et al. ref. 2020; Dzahini et al. ref. 2018). In contrast, CBD treatment was not associated with any apparent developmental toxicity, underscoring its safer profile in comparison to RISP.
Mechanistically, our biochemical assays revealed that neither VPA exposure nor treatments with RISP or CBD significantly altered some parameters of oxidative stress (CAT or GST) activities, indicating that these classical antioxidant defenses were not profoundly engaged in our model. However, CBD uniquely reduced the TBARS levels, suggesting a protective effect against LPO. Zebrafish larvae treated with CBD exhibited significantly lower TBARS levels than untreated and RISP‐treated groups. LPO has been implicated in the pathophysiology of ASD (Chauhan and Chauhan ref. 2006; Rose et al. ref. 2012), and its reduction by CBD provides a plausible mechanistic link to its behavioral benefits (Banji et al. ref. 2011). CBD’s antioxidant properties have been characterized across various neuroprotective contexts (Atalay et al. ref. 2019; Campos et al. ref. 2016; Hampson et al. ref. 1998). At the neurochemical level, CBD did not change DA, 5‐HT, or endocannabinoid levels. Preliminary evidence suggests that CBD modulates endocannabinoid tone, including AEA levels (Campos et al. ref. 2013), which could contribute to restoring the neurochemical balance disrupted by early VPA exposure.
Neurobiologically, GFAP staining initially suggested reduced astrocyte activity in VPA‐exposed animals. CBD treatment restored GFAP expression levels, suggesting a protective effect on glial populations or neurogenic niches disrupted by VPA, as it is important to emphasize that GFAP is also expressed in radial glial cells and neural stem cells during development and in postnatal brains (Alvarez‐Buylla et al. ref. 2001), complicating straightforward interpretation. CBD can also facilitate postnatal neurogenesis (Campos et al. ref. 2013). Additionally, we found that VPA increased the expression of the proliferating cell nuclear antigen (PCNA). RISP treatment significantly reduced this VPA‐induced PCNA upregulation, whereas CBD did not exert a significant effect. These findings may reflect a differential impact of treatments on neurogenesis or gliogenesis processes following early brain injury. Increased PCNA could indicate compensatory neurogenesis or reactive gliosis triggered by VPA (Sharma et al. ref. 2018). The fact that CBD did not reduce PCNA levels suggests that, unlike RISP, CBD may allow ongoing repair or adaptation processes without suppressing endogenous proliferative responses. This could confer an advantage in preserving neural plasticity mechanisms essential for functional recovery in ASD models. Regarding calcium signaling and molecules that influence synapse formation and brain development (Bryan ref. 2010), CaM expression was found elevated in VPA‐exposed animals, but reduced by both RISP and CBD, with CBD producing a more significant normalization. This result could reflect a partial reversal of the disrupted excitatory/inhibitory imbalance, which is considered a hallmark of ASD neuropathology (Nelson and Valakh ref. 2015).
Overall, these findings suggest that CBD not only reverses key behavioral impairments associated with embryonic VPA exposure, but also does so with greater potency and a better safety profile than RISP. Given the translational value of the zebrafish model—offering rapid, high‐throughput behavioral and biochemical screening—our work provides a promising platform for further investigating CBD’s therapeutic potential in ASD and related neurodevelopmental disorders. Future studies should explore the long‐term impacts of CBD treatment beyond early larval stages and elucidate the broader molecular pathways through which CBD confers neuroprotection.
However, as in every preclinical study, our results have limitations that should be acknowledged. First, our behavioral assessments were restricted to early larval stages (7 dpf); thus, we could not evaluate the persistence of VPA‐induced deficits or the durability of CBD’s therapeutic effects in “adolescent/adult” zebrafish. Second, while we identified a reduction in LPO as a potential mechanism underlying CBD’s benefits, this study did not explore other neurobiological pathways, such as inflammatory cascades. Finally, although zebrafish models offer valuable translational insights, the clinical implications of our findings require further investigation in mammalian models and, ultimately, in human studies.
Funding
This work was supported by Fundação de Amparo à Pesquisa do Estado de São Paulo (15/05551‐0, 17/24304‐0, 20/05416‐4, 14/50891‐1) and INCT 2014: Translational Medicine (465458/2014‐9).
Conflicts of Interest
F.S.G., J.A.C., and J.E.C.H. are the coinventors of the patent “Fluorinated CBD compounds, compositions, and uses thereof, Pub. No.: WO/2014/108899, International Application No.: PCT/IL2014/050023,” Def. US number Reg. 62193296; July 29, 2015; INPI on August 19, 2015 (BR1120150164927; Mechoulam R, Zuardi AW, Kapczinski F, Hallak JEC, Guimarães FS, Crippa JAS, Breuer A). Universidade de São Paulo (USP) has licensed this patent to Phytecs Pharm (USP Resolution No. 15.1.130002.1.1). A.C.C., F.S.G., J.A.C., and J.E.C.H. have an agreement with Prati‐Donaduzzi to “develop a pharmaceutical product containing synthetic CBD and prove its safety and therapeutic efficacy in the treatment of epilepsy, schizophrenia, Parkinson’s disease, and anxiety disorders.” A.C.C., F.S.G., J.A.C., and J.E.C.H. are the coinventors of the patent “Cannabinoid‐containing oral pharmaceutical composition, method for preparing and using the same,” INPI on September 16, 2016 (BR 112018005423‐2). J.A.C. was a consultant and/or received speaker fees and/or sits on the advisory board and/or receives research funding and/or receives speaker fees from Janssen‐Cilag, Torrent Pharm, Ease Labs Pharm, Prati‐Donaduzzi, Mantecorp, ArtMed, PurMed Global, BioBrains, and BSPG Pharm over the past 5 years. J.A.C. is a member of the International Advisory Board of the Australian Centre for Cannabinoid Clinical and Research Excellence (ACRE)—National Health and Medical Research Council (NHMRC). The remaining authors declare no conflicts of interest.
Supplementary Materials
References
- Updates on the Management of Autism Spectrum Disorder: Current Policies and Practices in Singapore.”. Neurotherapeutics, 2022. [DOI]
- Identification of Neural Stem Cells in the Adult Vertebrate Brain.”. Brain Research Bulletin, 2001. [DOI]
- Antioxidative and Anti‐Inflammatory Properties of Cannabidiol.”. Antioxidants, 2019. [DOI | PubMed]
- Investigation on the Role of Cannabis sativa Extract on Oxidative Stress and Mitochondrial Enzymes in Neonatal Rat Brain.”. Chinese Journal of Integrative Medicine, 2011. [DOI]
- Oral Cannabidiol Use in Children With Autism Spectrum Disorder to Treat Related Symptoms and Co‐Morbidities.”. Frontiers in Pharmacology, 2019. [DOI | PubMed]
- CBD‐Enriched Cannabis for Autism Spectrum Disorder: An Experience of a Single Center in Turkey and Reviews of the Literature.”. Journal of Cannabis Research, 2021. [DOI | PubMed]
- Comparative Studies on Different Methods of Malonaldehyde Determination.”. Methods in Enzymology, 1984. [DOI | PubMed]
- A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein‐Dye Binding.”. Analytical Biochemistry, 1976. [DOI | PubMed]
- Cannabidiol Did Not Induce Teratogenicity or Neurotoxicity in Exposed Zebrafish Embryos.”. Chemico‐Biological Interactions, 2018. [DOI | PubMed]
- Systematic Review of the Effectiveness of Pharmacological Treatments for Adolescents and Adults With Autism Spectrum Disorder.”. Autism, 2007. [DOI | PubMed]
- Calmodulin and Brain Development.”. Science Signaling, 2010. [DOI]
- Maturation of Shoaling Behavior is Accompanied by Changes in the Dopaminergic and Serotoninergic Systems in Zebrafish.”. Developmental Psychobiology, 2012. [DOI | PubMed]
- Cannabidiol Increases Survival and Promotes Rescue of Cognitive Function in a Murine Model of Cerebral Malaria.”. Neuroscience, 2016. [DOI]
- Multiple Mechanisms Involved in the Large‐Spectrum Therapeutic Potential of Cannabidiol in Psychiatric Disorders.”. Philosophical Transactions of the Royal Society, B: Biological Sciences, 2013. [DOI]
- Valproate‐Induced Model of Autism in Adult Zebrafish: A Systematic Review.”. Cells, 2025. [DOI | PubMed]
- Oxidative Stress in Autism.”. Pathophysiology, 2006. [DOI | PubMed]
- Valproic Acid Exposure During Pregnancy and the Risk of Autism Spectrum Disorders: A Systematic Review and Meta‐Analysis.”. Brain Research Bulletin, 2020. [DOI | PubMed]
- The transgenerational inheritance of autism‐like phenotypes in mice exposed to valproic acid during pregnancy.”. Sci Rep, 2016. [DOI | PubMed]
- Prenatal Valproate Exposure and Risk of Autism Spectrum Disorders and Childhood Autism.”. JAMA, 2013. [DOI | PubMed]
- Claiborne, A. 1985. “Catalase Activity.” In CRC Handbook of Methods for Oxygen Radical Research, edited by R. A. Greenwald , 283–284. CRC Press.
- Tail Coiling Assay in Zebrafish (Danio rerio) Embryos: Stage of Development, Promising Positive Control Candidates, and Selection of an Appropriate Organic Solvent for Screening of Developmental Neurotoxicity (DNT).”. Water, 2021. [DOI]
- What Do Fish Make of Mirror Images?”. Biology Letters, 2010. [DOI | PubMed]
- Larval Zebrafish Model for Studying the Effects of Valproic Acid on Neurodevelopment: An Approach Towards Modeling Autism.”. Journal of Pharmacological and Toxicological Methods, 2019. [DOI | PubMed]
- Antipsychotic Drug Use and Pneumonia: Systematic Review and Meta‐Analysis.”. Journal of Psychopharmacology, 2018. [DOI | PubMed]
- Assessment of Behavioral, Morphological and Electrophysiological Changes in Prenatal and Postnatal Valproate Induced Rat Models of Autism Spectrum Disorder.”. Scientific Reports, 2021. [DOI | PubMed]
- Assessment of Risperidone Toxicity in Zebrafish (Danio Rerio) Embryos.”. Comparative Medicine, 2023. [DOI | PubMed]
- Glutathione S‐Transferases: The First Enzymatic Step in Mercapturic Acid Formation.”. Journal of Biological Chemistry, 1974. [PubMed]
- Children and Adolescents With ASD Treated With CBD‐Rich Cannabis Exhibit Significant Improvements Particularly in Social Symptoms: An Open Label Study.”. Translational Psychiatry, 2022. [DOI | PubMed]
- Cannabidiol and (‐)Delta9‐Tetrahydrocannabinol Are Neuroprotective Antioxidants.”. Proceedings of the National Academy of Sciences of the United States of America, 1998. [DOI | PubMed]
- Environmental Factors Influencing the Risk of Autism.”. Journal of Research in Medical Sciences: The Official Journal of Isfahan University of Medical Sciences, 2017. [DOI | PubMed]
- The Critical Period of Valproate Exposure to Induce Autistic Symptoms in Sprague‐Dawley Rats.”. Toxicology Letters, 2011. [DOI | PubMed]
- Behavioral Interventions for Children With Autism Spectrum Disorders.”. Pediatric Clinics of North America, 2012. [DOI | PubMed]
- Autism Spectrum Disorder.”. Lancet, 2018. [DOI | PubMed]
- Exploring the Validity of Valproic Acid Animal Model of Autism.”. Experimental Neurobiology, 2015. [DOI | PubMed]
- Prevalence and Characteristics of Autism Spectrum Disorder Among Children Aged 8 Years – Autism and Developmental Disabilities Monitoring Network, 11 Sites, United States, 2020.”. MMWR Surveillance Summaries, 2023. [DOI]
- Critical Periods and Autism Spectrum Disorders, a Role for Sleep.”. Neurobiology of Sleep and Circadian Rhythms, 2023. [DOI | PubMed]
- Zebrafish Models of Autism Spectrum Disorder.”. Experimental Neurology, 2018. [DOI | PubMed]
- Behavioral Syndromes and the Evolution of Correlated Behavior in Zebrafish.”. Behavioral Ecology, 2007. [DOI]
- Outcomes of Cognitive Behaviour Therapy for Obsessive‐Compulsive Disorder in Young People With and Without Autism Spectrum Disorders: A Case Controlled Study.”. Psychiatry Research, 2015. [DOI | PubMed]
- DarT: The Embryo Test With the Zebrafish Danio Rerio – A General Model in Ecotoxicology and Toxicology.”. ALTEX, 2002. [PubMed]
- Excitatory/Inhibitory Balance and Circuit Homeostasis in Autism Spectrum Disorders.”. Neuron, 2015. [DOI | PubMed]
- The Valproic Acid‐Induced Rodent Model of Autism.”. Experimental Neurology, 2018. [DOI | PubMed]
- OECD . 2025. Test No. 236: Fish Embryo Acute Toxicity (FET) Test, OECD Guidelines for the Testing of Chemicals Section 2. OECD Publishing. 10.1787/9789264203709-en.
- Assay for Lipid Peroxides in Animal Tissues by Thiobarbituric Acid Reaction.”. Analytical Biochemistry, 1979. [DOI | PubMed]
- Valproic Acid in Pregnancy Revisited: Neurobehavioral, Biochemical and Molecular Changes Affecting the Embryo and Fetus in Humans and in Animals: A Narrative Review.”. Int J Mol Sci, 2023. [DOI | PubMed]
- Cannabidiol for the Treatment of Autism Spectrum Disorder: Hope or Hype?”. Psychopharmacology, 2022. [DOI | PubMed]
- Acute Cannabidiol Treatment Reverses Behavioral Impairments Induced by Embryonic Valproic Acid Exposure in Male Mice.”. Pharmacology, Biochemistry, and Behavior, 2025. [DOI | PubMed]
- Pham, M. , J. Raymond , J. Hester , et al. 2012. “Assessing Social Behavior Phenotypes in Adult Zebrafish: Shoaling, Social Preference, and Mirror Biting Tests.” In Zebrafish Protocols for Neurobehavioral Research, edited by A. V. Kalueff and A. M. Stewart , 231–246. Humana Press.
- Effects of Cannabidiol on Brain Excitation and Inhibition Systems: A Randomised Placebo‐Controlled Single Dose Trial During Magnetic Resonance Spectroscopy in Adults With and Without Autism Spectrum Disorder.”. Neuropsychopharmacology, 2019. [DOI | PubMed]
- Evidence of Oxidative Damage and Inflammation Associated With Low Glutathione Redox Status in the Autism Brain.”. Translational Psychiatry, 2012. [DOI | PubMed]
- Large‐Scale Exome Sequencing Study Implicates Both Developmental and Functional Changes in the Neurobiology of Autism.”. Cell, 2020. [DOI | PubMed]
- Behavioral Alterations in Rats Prenatally Exposed to Valproic Acid: Animal Model of Autism.”. Neuropsychopharmacology, 2005. [DOI | PubMed]
- Autism Spectrum Disorder: Classification, Diagnosis and Therapy.”. Pharmacology & Therapeutics, 2018. [DOI | PubMed]
- Evaluation of the Efficacy and Safety of Cannabidiol‐Rich Cannabis Extract in Children With Autism Spectrum Disorder: Randomized, Double‐Blind, and Placebo‐Controlled Clinical Trial.”. Trends in Psychiatry and Psychotherapy, 2024. [DOI | PubMed]
- A New Neurobehavioral Model of Autism in Mice: Pre‐ and Postnatal Exposure to Sodium Valproate.”. Journal of Autism and Developmental Disorders, 2006. [DOI | PubMed]
- Use of Antipsychotics and the Risk of Acute Respiratory Failure Among Adults: A Disease Risk Score‐Matched Nested Case‐Control Study.”. British Journal of Clinical Pharmacology, 2020. [DOI | PubMed]
- Embryological Exposure to Valproic Acid Induces Social Interaction Deficits in Zebrafish (Danio Rerio): A Developmental Behavior Analysis.”. Neurotoxicology and Teratology, 2015. [DOI | PubMed]