
The endocannabinoidome–gut microbiome–brain axis as a novel therapeutic target for autism spectrum disorder
Abstract
Introduction:
Autism spectrum disorder (ASD) is characterized by disruption of the gut–brain axis, which leads to behavioral, psychiatric, metabolic and gastrointestinal symptoms. Effective ASD treatments are limited. Research highlights the roles of the endocannabinoidome (eCBome) and gut microbiome (GM), both crucial for brain and gut function. This review summarizes research on therapeutic targets within the eCBome–GM–brain axis for ASD and related comorbidities.
Discussion:
Evidence suggests that reduced levels of eCBome mediators, like oleoylethanolamide and anandamide, and altered cannabinoid type 1 and type 2 (CB1 and CB2) receptors activity may contribute to ASD symptoms, making them promising targets. Modulating the eCBome–GM–brain axis with inhibitors of fatty acid amide hydrolase (FAAH), transient receptor potential vanilloid 1, and monoacylglycerol lipase (MAGL) may improve repetitive, stereotypical, and sensory behaviors, and alleviate sociability impairments, depression and anxiety. However, inhibition of FAAH and MAGL may also induce ADHD-like behaviors, which can be reversed by CB1 inverse agonists. Targeting metabotropic glutamate receptor 5 to increase levels of the eCBome mediator 2-arachidonoylglycerol (2-AG) may benefit ASD-related behaviors. eCBome mediators such as 2-AG, 1/2-palmitoylglycerol and palmitoylethanolamide may also help manage ASD- and GI-related symptoms, and systemic inflammation. Other potential therapeutic targets that deserve further investigation are eCBome-related receptors G-protein-coupled receptor 55 and peroxisome proliferator-activated receptors-alpha and -gamma, and the cyclooxygenase-2/prostaglandin E2 pathway, which may address hyperactivity and repetitive behaviors. Additionally, mucin-degrading genera like Akkermansia and Ruminococcus may improve ASD-related GI symptoms such as hypersensitivity and inflammation. Selective antibiotics against specific Clostridium strains may improve irritability and aggression. In ASD with ADHD and OCD, treatments may involve modulating the CB1 and CB2 receptor, and bacterial families like Ruminococcaceae and Lachnospiraceae. Lastly, modulating the abundance of anti-inflammatory genera like Prevotella and Anaeroplasma, and taxa associated with gut health such as Roseburia may also offer therapeutic value.
Conclusion:
The eCBome–GM–brain axis is a promising target for ASD treatment, meriting further clinical and preclinical research.
Supplementary Information:
The online version contains supplementary material available at 10.1186/s12929-025-01145-7.
Article type: Review Article
Keywords: Autism spectrum disorder, Endocannabinoidome, Gut microbiome, Gut–brain axis, Mental health, Metabolic health, Gastrointestinal health, Therapeutic targets, Biomarkers
Affiliations: https://ror.org/01pxwe438grid.14709.3b0000 0004 1936 8649Department of Psychiatry, McGill University, Montreal, QC H4H 1R2 Canada; https://ror.org/03a64bh57grid.8158.40000 0004 1757 1969Department of Experimental Medicine, Division of Molecular Biology, Biotechnology and Histology, University of Campania, 80138 Naples, Campania Italy; https://ror.org/02aqtvv10grid.512214.1European Biomedical Research Institute of Salerno (EBRIS), 84125 Salerno, Campania Italy; https://ror.org/04sjchr03grid.23856.3a0000 0004 1936 8390Centre de Recherche de L’Institut Universitaire De Cardiologie Et De Pneumologie de Québec, Département of Médecine, Université Laval, Quebec, QC G1 V 4G5 Canada; https://ror.org/04sjchr03grid.23856.3a0000 0004 1936 8390Canada Excellence Research Chair On the Microbiome-Endocannabinoidome Axis in Metabolic Health (CERC-MEND), Université Laval, Quebec, QC G1 V 0 V6 Canada; https://ror.org/04sjchr03grid.23856.3a0000 0004 1936 8390Institut sur la Nutrition et les Aliments Fonctionnels, and Centre NUTRISS, École de Nutrition, Université Laval, Quebec, QC G1 V 0 V6 Canada; https://ror.org/03wyf0g15grid.473581.c0000 0004 1761 6004Joint International Unit Between the CNR of Italy, Institute of Biomolecular Chemistry, and Université Laval on Chemical and Biomolecular on the Microbiome and its Impact on Metabolic Health and Nutrition (UMI-MicroMeNu), Quebec, QC G1 V 0 V6 Canada
License: © The Author(s) 2025 CC BY 4.0 Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
Article links: DOI: 10.1186/s12929-025-01145-7 | PubMed: 40605060 | PMC: PMC12220735
Relevance: Moderate: mentioned 3+ times in text
Full text: PDF (1.7 MB)
Introduction
Autism Spectrum Disorder (ASD), a neurodevelopmental condition affecting 1 in 36 individuals [ref. 1], presents challenges including atypical social communication and interaction, stereotyped language, atypical interests, repetitive and stereotyped behavior [ref. 2], and heightened risk for psychiatric and gastrointestinal (GI) comorbidities [ref. 3–ref. 6]. Struggling to integrate into society, adults with ASD often face unemployment [ref. 7], loneliness [ref. 8], and financial hardships [ref. 9], further exacerbating their already burdened physical and mental wellbeing. Existing behavioral and pharmacological interventions have fallen short in providing evidence of treatment effectiveness [ref. 10, ref. 11]. Therefore, investigating novel therapeutic targets for ASD is encouraged. Recently, two emerging sources of chemical signals—the endocannabinoidome (eCBome) and the gut microbiome (GM)—have raised hopes for developing new therapies to address brain dysregulations [ref. 12, ref. 13].
The eCBome is an intricate regulatory network that includes the eCB system, with its two major eCBs N-arachidonoyl-ethanolamine (anandamide, AEA) and 2-arachidonoyl-glycerol (2-AG), and cannabinoid receptors (CB) of type 1 and 2. The eCBome also includes endogenous and GM-derived eCB-like lipid compounds, a wide array of biosynthetic and inactivating enzymes, receptors, and ion channels, forming a complex web of over 100 mediators, 20 enzymes, and 20 receptors known [ref. 14]. The eCBome is involved in social behavior, mood, synaptic plasticity, pain, sleep, GM bacterial composition, gut barrier permeability and gut immunity [ref. 15–ref. 21], all of which are altered in ASD and can be targeted therapeutically [ref. 22].
The GM is the community of microorganisms that resides in the digestive tracts, and is involved in various processes, including: (1) digestion, absorption, detoxification and vitamin synthesis, as well as production of short-chain fatty acids (SCFAs) and other metabolites that are essential for maintaining gut and brain health and providing energy; (2) immune system regulation, inflammation, gut and brain immunity and respective barrier permeability; (3) metabolic functions—the GM influences the host energy balance and plays a role in obesity and metabolic syndrome; and (4) the gut–brain axis and related behavioural, affective and cognitive functions. The bidirectional communication between the GM and the brain involves neural, endocrine, and immune pathways, and has been linked to various neuro-psychiatric conditions, including ASD and its comorbidities [ref. 23, ref. 24].
A bidirectional interaction between the eCBome and the GM has emerged recently [ref. 25–ref. 27], and the effect of this interaction on the brain are altered in states such as ASD. For example, altered eCBome in ASD [ref. 22, ref. 28, ref. 29] may reflect changes in GM composition (dysbiosis, a pathological shift of GM composition) [ref. 30–ref. 32] and vice-versa. Dysbiosis contributes to disruptions in gut immunity, gut barrier integrity, and gut, systemic, and neuroinflammation [ref. 33–ref. 39], exacerbating GI, systemic, and brain-related symptoms. Certain bacteria and their metabolites can also directly cross the blood–brain barrier, exerting neurotoxic effects [ref. 40] and contributing to ASD-like behaviors [ref. 41–ref. 44]. Some GM strains have been reported to influence the eCB system and exert anti-inflammatory effects [ref. 45]. Additionally, some eCB system modulators have shown promising results in alleviating or reversing ASD symptoms in animals [ref. 46–ref. 53]. While there are several studies on the role of the eCB system in ASD, little is known about the eCBome. Due to its roles, targeting eCBome receptors in ASD may have therapeutic potential at both the gut and brain level [ref. 54–ref. 58].
The present article examines the eCBome–GM–brain axis as an emerging therapeutic target in ASD.
Animal models to study the gut–brain axis
Animal models have been critical in investigating the gut–brain axis and its role in neurological conditions, including ASD. Germ-free (GF) mice are among the most commonly used models, as they are raised in a sterile environment devoid of any microorganisms. This allows researchers to observe the impact of the absence of a microbiome on brain function and behavior [ref. 59]. These mice can then be colonized with a single or multiple microbial communities to investigate how specific microbes influence the brain, or subjected to a fecal matter transplant (FMT), where feces from one individual or mouse are transplanted into a GF mouse, to examine the role of the gut microbiome specifically.
Conventionalized ex-GF mice, which are initially GF and then colonized with specific microbiota, help researchers explore the impact of particular bacterial communities on brain development and behavior. Monocolonization experiments, where a single bacterial species is introduced into GF mice, help pinpoint the effects of individual microbes on brain function. Also antibiotic treatments can be used to selectively deplete or alter the gut microbiota, allowing studies on how changes in microbial composition affect neurological outcomes [ref. 60].
Additionally, more complex approaches like using cocktails of probiotics, which consist of multiple beneficial bacteria, help understand how a diverse microbiome influences the brain [ref. 60]. In some studies, a pathogen and a protective strain are introduced to study the interaction between harmful and beneficial microbes and their combined effects on brain function. Vagotomy (severing the vagus nerve) [ref. 61] and chemical sympathectomy (disrupting sympathetic nervous system activity through chemical means) [ref. 62, ref. 63] can also be employed as a model to block specific neural pathways and explore how gut-derived signals impact brain activity.
Other models also provide valuable insights into the gut–brain axis. Humanized mouse models, where animals are engineered to express human microbiota or genes [ref. 64], allow for the study of human-specific microbial influences on the brain [ref. 65]. An FMT can reveal how the gut microbiota from different individuals (e.g., ASD) and mouse models (i.e. mice harbouring risk gene mutations for ASD) influences brain function [ref. 66]. Dietary interventions are another important tool—by altering the animals’ diet, researchers can investigate how different nutritional components (e.g., high-fat, fiber-rich, “Cafeteria diet”) affect the gut microbiome and, in turn, brain function and behavior [ref. 67].
Moreover, neuroinflammation models (e.g., LPS injections) [ref. 68] allow for the study of how inflammation in the gut may influence brain function, particularly in the context of ASD. Chronic stress models, such as repeated social stress or maternal separation [ref. 69], help examine the role of stress-induced changes in the microbiome on brain activity and behavior [ref. 70]. Knockout and transgenic mouse models (e.g., Shank3, Fmr1 knockout mice; addressed in “Genetic models of ASD” section) enable the exploration of gene-environment interactions by investigating how specific genetic mutations associated with neurodevelopmental disorders affect gut–brain signaling (see “Genetic models of ASD” section).
Finally, chronic illness models such as those induced by dextran sodium sulfate (DSS) [ref. 71] or trinitrobenzenesulfonic acid (TNBS) [ref. 72], which mimic conditions like IBD, can provide insights into how gut diseases or inflammation influence brain development and behavior. Social interaction models, including social defeat or social preference tests, also help examine how microbiome alterations affect social behaviors [ref. 73, ref. 74]—an area of particular interest in ASD [ref. 75].
These models offer valuable insights into the interactions between the gut microbiome, the brain, and behaviors associated with neurodevelopmental disorders like ASD.
Use of germ-free mice to assess the impact of specific bacterial species on the gut–brain axis
A number of studies have explored the effect of single-strain colonization in GF mice. While many of these studies focus on the impact of specific bacteria on colonization dynamics throughout the gut and gastrointestinal and systemic inflammation, only a few have investigated their direct effects on the gut–brain axis, influencing behavior and mental health. Philip et al. [ref. 76] investigated the effect of E. coli strains (JM83, HA107) and Salmonella enterica serovar Typhimurium in GF mice. Their study demonstrated that neural plasticity changes rapidly after initial gut microbial colonization and that this process involves innate immune signaling to the brain. The migration of dendritic cells plays a key role in this communication. Nishino et al. [ref. 77] observed the colonization effect of strains like Brauia coccoides and Bifidobacterium infantis on behavior, such as reduced anxiety and locomotor activity. This research supports the notion that gut microbiota can modulate neurotransmitter levels and neurodevelopment, which in turn affect behavior. Liu et al. [ref. 78] administered Lactobacillus plantarum PS128 in GF mice. The findings demonstrated that live PS128 is safe for chronic ingestion through the gut brain axis and can serve as an anxiolytic agent to regulate the motor functions and mood of the host via modulation of neurotransmitters such as dopamine and serotonin. Parker et al. [ref. 79] demonstrated that Candida albicans can translocate from the gut to the brain, where it can trigger an inflammatory response. Indeed, in their study, the yeast was often found in close proximity of activated microglial cells. This research supports the idea that disruptions in the gut not only of bacteriome but also of the mycobiome and the intestinal barrier may be important contributors to neurological diseases.
Many other studies have addressed the effect of specific single-strain bacteria on intestinal motility [ref. 80], enteric immune activity and systemic inflammation [ref. 81–ref. 100]. The explored outcomes on immune function could therefore putatively be an indirect index of their potential impact on the gut–brain axis.
ASD symptoms and etiology
Autism Spectrum Disorder is a neurodevelopmental condition whose core symptoms include atypical social communication and interaction, stereotyped language, repetitive and stereotyped behavior, and atypical interests [ref. 101]. Individuals with ASD have up to 78% increased risk of psychiatric comorbidities compared to neurotypical individuals [ref. 102]. These include anxiety [ref. 103], depressive syndrome [ref. 104], obsessive compulsive disorder (OCD) [ref. 105], attention deficit hyperactivity disorder (ADHD) [ref. 106], as well as atypical behaviors such as sociability impairment [ref. 107], sensory behavior [ref. 108], hyperarousal [ref. 109], irritability [ref. 110], tantrum and/or self-injury behavior [ref. 111]. Up to 70% of individuals with ASD also suffer from GI [ref. 112] and metabolism-related disorders [ref. 113] such as gut dysbiosis, digestive enzyme deficits [ref. 114] and metabolic syndrome [ref. 113, ref. 115].
An epidemiological study from 2022 reports significant variations in ASD prevalence across continents. The highest prevalence is observed in Africa (2963/10,000), followed by Oceania (258/10,000) and the Americas (129/10,000), while lower rates are reported in Asia (34/10,000) and Europe (116/10,000) [ref. 116]. These discrepancies may arise from genetic, environmental, and diagnostic differences but also from dietary patterns that shape gut microbiota composition [ref. 117].
In Africa, food insecurity and poor maternal nutrition during pregnancy can impair fetal brain development, increasing ASD risk [ref. 118]. Additionally, prenatal and early-life exposure to toxins, pollutants, and infections may further contribute to ASD pathophysiology [ref. 119, ref. 120]. Diet plays a crucial role in shaping gut microbiota, which in turn influences neurodevelopment. Western diets, rich in fats, sugars, and processed foods, are associated with gut microbiota dysbiosis, increased intestinal permeability, and systemic inflammation—factors implicated in ASD [ref. 121, ref. 122]. In contrast, Mediterranean and traditional Asian diets, characterized by high fiber intake and fermented foods, promote microbial diversity and the production of short-chain fatty acids (SCFAs), which exert metabolic [ref. 123] and neuroprotective effects [ref. 124–ref. 126]. Notably, Western dietary patterns have been linked to cognitive decline, whereas Mediterranean diets have demonstrated cognitive benefits [ref. 127]. Thus, epidemiological evidence indirectly supports the link between GM composition and ASD symptom severity and prevalence.
Animal models employed to investigate ASD symptoms and etiology
ASD is highly heterogeneous not only in its phenotypic presentation but also in its etiology. The disorder is often classified into different sub-populations based on genetic and epigenetic modifications [ref. 128]. Preclinical models of ASD replicate certain phenotypes through genetic, epigenetic, or combined factors, providing controlled environments to study the disorder’s complexities and test hypotheses. These models can be divided into two main categories: genetic models and environmental models [ref. 129].
Genetic models of ASD
The genetic architecture of autism is complex and lacks a single causative gene, but clear genetic bases are identifiable in about 20–25% of cases [ref. 130]. ASD often co-occurs with rare Mendelian neurodevelopmental disorders such as fragile X syndrome (FXS) [ref. 131, ref. 132], Rett syndrome (RTT) [ref. 133, ref. 134], neurofibromatosis type 1 (NF1) [ref. 135], and tuberous sclerosis complex (TSC) [ref. 136] as well as copy number variations like those in Phelan–McDermid syndrome, which involves mutations in SH3 and multiple ankyrin repeat domains 3 (SHANK3) [ref. 137].
Fragile X syndrome is caused by an expanded CGG trinucleotide repeat in the fragile X messenger ribonucleoprotein 1 (FMR1) gene, leading to disrupted synaptic function [ref. 131] and ASD-like behavioral phenotypes such as intellectual disability, hyperarousal, and impulsivity [ref. 132]. Mutations in the methyl-CpG binding protein 2 (MECP2) gene cause Rett syndrome, leading to developmental regression, motor and language impairments, stereotypic movements, seizures, and autonomic dysfunction [ref. 133, ref. 134]. Autosomal dominant mutations in the neurofibromin 1 (NF1) gene cause tumor formation and neuronal dysfunction, leading to cognitive, language, and behavioral deficits, with high rates of ASD [ref. 135]. Mutations in tuberous sclerosis complex 1 (TSC1) and tuberous sclerosis complex 2 (TSC2) genes disrupt neuronal development, resulting in seizures, intellectual disability, and ASD in about 50% of affected individuals [ref. 138]. SHANK3 mutations in Phelan–McDermid syndrome results in severe expressive language delays, hypotonia, and ASD-related behaviors [ref. 137].
Rodent models genetically engineered to mimic genetic forms of ASD offer a means to directly study the genetic contributions to ASD, investigating the behavioral phenotypes observed in ASD. It is important to note that not all ASD models exhibit robust construct validity. For example, models like the BTBR and C58/J mouse naturally display idiopathic autism-like behaviors. In such cases, the model validity is assessed based on face validity. Ultimately, no single animal model can entirely capture the complexity of ASD. However, collectively, these models provide a way to approximate it [ref. 139].
Environmental models of ASD
Despite high heritability, the lack of perfect concordance between monozygotic twins [ref. 130] suggests that environmental factors also play a role in the etiopathogenesis of ASD. Key environmental risk factors during the perinatal period include advanced parental age, maternal diabetes, maternal infections, medication use during pregnancy, and toxin exposure, though their associations with ASD vary [ref. 140, ref. 141]. Therefore, these environmental models of ASD study epigenetic causes of autism mainly focusing on the maternal immune system activation (MIA) and prenatal exposure to medications and toxins [ref. 142]. Exposure to valproic acid (VPA), a widely studied antiepileptic and mood stabilizer, serves as a model of MIA during pregnancy, along with other compounds like thalidomide, misoprostol, and propionic acid [ref. 143]. These environmental models are often combined with genetic modifications to explore gene × environment interactions.
Germ-free mice to investigate the gut–brain axis in ASD
GF mice have been instrumental in dissecting causal relationships between gut microbes and ASD-like behaviors. In some studies, these mice have been found to exhibit social deficits, increased anxiety-like traits, and altered brain development, mirroring core ASD phenotypes [ref. 59]. Mechanistically, the absence of microbiota in GF mice may affect microglial maturation, HPA axis regulation, neurotransmitter systems, and immune activation, all of which contribute to ASD pathophysiology [ref. 144, ref. 145]. When gut microbiota are reintroduced into these mice, researchers can observe the effects of specific bacterial species on ASD-related behaviors. For instance, Bifidobacterium infantis, a species commonly used in microbiota studies, has been shown to alleviate anxiety-like behaviors and stress responses in GF mice, indicating its potential role in modulating behavior [ref. 77, ref. 144, ref. 145].
FMT studies in GF mice also provide insights into how specific bacteria contribute to ASD-related behaviors. FMT from healthy donors to GF mice has been shown to improve social behaviors, whereas FMT from ASD donors can induce ASD-like behaviors. For example, Xiao et al. [ref. 146] demonstrated that microbiota from ASD individuals induced ASD-like behaviors in GF mice, with certain bacterial species likely playing a key role in these effects. These findings suggest that specific bacteria in the gut microbiota can influence ASD-like behaviors, providing a preliminary picture of how individual species contribute to neurodevelopment and behavior in preclinical models.
The influence of gut microbiota on preclinical models of ASD
Gut microbiota plays a critical role in shaping preclinical models used to study ASD, for example by altering intestinal barrier integrity [ref. 147–ref. 151], immune responses, neuroinflammation [ref. 152–ref. 154], and neural signaling [ref. 155]—factors that contribute to ASD-like behaviors. However, despite clear associations between gut dysbiosis and ASD phenotypes [ref. 156], findings across ASD models remain inconsistent [ref. 157]. Differences in genetic background, environmental conditions and sex may contribute to these discrepancies, affecting the reproducibility and interpretation of microbiota-driven effects in ASD research.
One key mechanism through which microbiota influences ASD models (both genetic [ref. 147, ref. 148], environmental [ref. 149] and idiopathic ones [ref. 150, ref. 151]) is the disruption of gut barrier integrity, leading to systemic inflammation and altered brain function. These models display increased intestinal permeability [ref. 99–ref. 103], yet some studies report no significant differences, suggesting that microbiota-driven barrier dysfunction may be strain- or condition-dependent [ref. 157]. Additionally, gut microbiota composition differs across ASD models, with alterations in α-diversity, β-diversity, and specific taxa linked to ASD-like traits (reviewed in [ref. 156]). While some studies report consistent microbial signatures associated with ASD, others find conflicting patterns, even within the same model [ref. 156], emphasizing the complexity of microbiota contributions. Sex differences further modulate these findings, influencing microbiota composition and ASD-related behaviors in genetic [ref. 158, ref. 159], environmental [ref. 160–ref. 163], and idiopathic [ref. 150] models.
Microbiota-derived metabolites, particularly SCFAs, further shape ASD models. Butyrate, which modulates gut integrity and neuroinflammation [ref. 152, ref. 164], is elevated in several environmental [ref. 160], genetic [ref. 159], and idiopathic [ref. 151] models of ASD. However, not all studies report this increase—Rett syndrome models, for instance, show no significant changes in butyrate levels [ref. 165]. Interestingly, sodium butyrate administration improves atypical behaviors and cognition in BTBR [ref. 33, ref. 166] and VPA models [ref. 167], yet its effects on gut microbiota composition in ASD remain unclear, with some studies suggesting modulation of gut microbiota and permeability in environmental model-specific responses [ref. 168–ref. 170]. Conversely, propionate (PA), another SCFA, consistently induces ASD-like behaviors in rodents, highlighting how microbial-produced SCFAs can either mitigate or worsen ASD traits depending on the model.
Microbiota-targeted interventions, including probiotics and antibiotics, further illustrate the variability in microbiota-immune-brain interactions across ASD models. While Bacteroides fragilis restored gut permeability and improved behavioral deficits, including reducing repetitive behaviors and anxiety-like symptoms, in MIA models [ref. 149], it did not fully rescue social deficits, highlighting the model-dependent limitations of microbiota-based interventions. Similarly, Lactobacillus reuteri improved social and repetitive behaviors in multiple (genetic [ref. 157, ref. 158], idiopathic [ref. 157], and environmental [ref. 157, ref. 171]) ASD models. Antibiotic studies also show interesting results—vancomycin treatment in MIA models improved ASD-like behaviors, sociability and anxiety [ref. 172], streptomycin rebalanced gut dysbiosis and memory deficits in another MIA model [ref. 173], while neomycin improved social behavior in a genetic model [ref. 174].
Co-housing studies demonstrated how gut microbiota can influence ASD-like traits in preclinical maternal environmental models [ref. 175] through microbial exchange via coprophagy and grooming [ref. 176]. Early life is a critical period for these interactions, which may shape neurodevelopment. For example, Lammert et al. [ref. 175] showed that offspring of segmented filamentous bacteria (SFB)-negative dams exhibited ASD-like behaviors only when co-housed with SFB-positive dams, indicating the importance of microbiota exchange in model outcomes. Additionally, Buffington et al. [ref. 171] found that co-housing maternal high-fat diet offspring with control mice improved social behaviors and microbiota composition, suggesting a microbiota-driven rescue effect. These findings highlight the potential of co-housing as a method to study the role of microbiota in ASD models, though further research is needed to confirm these effects.
The eCBome–GM–brain axis as a model to investigate therapeutic targets in ASD and comorbidities
Individuals with ASD suffer from both gut- and brain-related comorbidities. Gastrointestinal symptoms in ASD are thought to be caused by central sensory augmentation, altered modulation of GI epithelial permeability including visceral hypersensitivity, and altered enteric nervous system-mediated motility and secretion [ref. 177]. Increased permeability and disruption of gut and blood–brain barrier integrity are observed in individuals with ASD [ref. 178, ref. 179]. This permeability results in a higher antigenic load from the GI tract entering circulation and crossing the BBB, inducing a brain immune response [ref. 180, ref. 181]. Pro-inflammatory mediators may directly communicate with microglia through the neurovascular network, creating a reactive environment that negatively affects the functioning of neuronal circuits responsible for mood regulation [ref. 182].
Immune dysfunction, gut dysbiosis, and the eCBome in ASD are closely intertwined. Gut dysbiosis contributes to immune activation and exacerbates intestinal permeability and inflammation, leading to GI symptoms [ref. 179, ref. 183]. The GI system maintains a close connection with the brain through the eCBome, primarily mediated by CB1 receptors and transient receptor potential cation channels subfamily V member 1 (TRPV1) in myenteric and vagal fibers [ref. 184, ref. 185], peroxisome proliferator-activated receptor alpha (PPAR-α), and oleoyl-lysophosphatidylinositol receptor 1 (GPR119) in enteroendocrine epithelial cells of the small intestine [ref. 186, ref. 187]. These receptors influence myenteric neuron activity, nerve function, and the release of GI and neuroactive neuropeptides. Dysbiosis in ASD affects mental health via the GM production of molecules that influence cognitive or social behaviors, impacting myenteric and vagal nerve activity directly or via the bloodstream, exacerbating ASD and systemic symptom severity [ref. 180, ref. 188, ref. 189].
Notably, neurotransmitters like serotonin or gamma-aminobutyric acid (GABA), and eCB-like mediators such as N-acyl amides that interact with G-protein-coupled receptors, produced from commensal microorganisms, can modulate host eCBome receptors [ref. 190–ref. 193]. Additionally, significant changes in GM abundance and composition can alter the expression of eCBome receptors and enzymes, and the concentrations of eCBome mediators in the gut, influencing the eCBome–GM–brain axis [ref. 27, ref. 194]. For instance, antibiotic-induced dysbiosis alters the levels of eCBs and their mediator congeners, ultimately leading to decreased neurogenesis and ASD-like behaviors, which can be normalized by treatment with probiotics such as Lactobacillus casei [ref. 195]. Specific GM taxa influence the eCB system, inducing the expression of CB1 and CB2 receptors, leading to anti-inflammatory and analgesic effects [ref. 45, ref. 196]. The administration of various strains of Lactobacillus and Bifidobacterium (genera underrepresented in ASD) have shown promise in improving ASD symptoms and gut integrity in both preclinical and clinical studies [ref. 149, ref. 171, ref. 196, ref. 197]. Indeed, several clinical trials administering probiotics containing Lactobacillus and Bifidobacterium strains (NCT04939974, NCT06448767, NCT04942522, NCT03514784, NCT02903030, NCT03369431, NCT02708901, NCT04293783, NCT06126185, NCT06650644, NCT05307744, NCT03982290) are ongoing or were recently completed. Others have administered Lactobacilli and Bifidobacteria in combination with prebiotics (NCT06126185, NCT04944901, NCT05151601, NCT02086110, NCT04639141). Fecal microbiota transplant-based therapies in children and adults with ASD are also under investigation (NCT06419530, NCT03426826, NCT04246398, NCT03408886, NCT04630847, NCT06030752, NCT04948814, NCT06290258, NCT02504554, NCT06503978, NCT04182633, NCT03829878).
The eCBome–GM–brain axis in ASD, anxiety, sociability and behavioral impairment
Gut microbial diversity is significantly disrupted in ASD, with a reduced ratio of Bacteroidetes to Firmicutes and an increase in Proteobacteria [ref. 198, ref. 199]. High levels of Proteobacteria in ASD, including opportunistic LPS-producing pathogens, may lead to systemic inflammation and impair antioxidant mechanisms [ref. 34, ref. 35]. These changes in turn can contribute to altered social behavior and anxio-depressive tendencies [ref. 200], comorbidities often manifested in ASD. Given the involvement of inflammation in ASD pathophysiology, increasing endocannabinoid (eCB) tone has been proposed as a potential therapeutic strategy [ref. 16, ref. 201].
Individuals with ASD exhibit reduced serum levels of the eCBs anandamide (AEA) and oleoylethanolamide (OEA) [ref. 29]. Notably, OEA and AEA increase oxytocin signaling [ref. 202–ref. 204]. Oxytocin administration was reported to improve several ASD symptoms and behaviors, such as enhancing social cognition [ref. 205, ref. 206] and behavior [ref. 207], emotional recognition of facial expressions [ref. 208], and reducing anxiety [ref. 209–ref. 213]. AEA by increasing oxytocin led to social reward and improved social impairment in ASD mouse models [ref. 53, ref. 204]. Therefore, interventions that restore AEA levels—such as CB1 receptor activation or inhibition of fatty acid amide hydrolase (FAAH) (the enzyme responsible for degrading AEA [ref. 214–ref. 216])—may help mitigate these symptoms [ref. 53, ref. 204]. In preclinical models, increasing eCB signaling via CB1 activation has been shown not only to enhance social reward [ref. 53], but also to reduce aggression [ref. 217], and stress-induced impulsivity [ref. 218, ref. 219]. These ASD-related symptoms will be discussed in detail in the next section.
However, the effects of CB1 modulation are highly context-dependent, as CB1 receptors regulate both GABAergic (inhibitory) and glutamatergic (excitatory) neurotransmission [ref. 220]. In some cases, CB1 antagonists (which reduce CB1 activity) have been reported to exert either anxiolytic or anxiogenic effects, depending on which neurotransmitter system they predominantly affect. Specifically, CB1 blockade reduces anxiety when targeting GABAergic pathways but increases anxiety when interfering with glutamatergic ones [ref. 220] (Table 1). These findings suggest that the effect of CB1 modulation in ASD may depend on the specific neurochemical transmission and alterations present in different ASD subtypes.
Table 1: The degenerative and therapeutic role of the eCBome in ASD
| eCBome role | Key findings | Model | References |
|---|---|---|---|
| Degenerative | Reduced eCB levels, increased eCBome enzyme degradation, and upregulated cannabinoid receptors (CBRs) contribute to ASD-like behaviors such as hyperactivity, repetitive actions, and social dysfunction | Children with ASD and in the VPA-induced animal model | [ref. 47] |
| Reduced levels of anandamide (AEA) promote anxiety | Male Sprague–Dawley rats | [ref. 214] | |
| CB1 antagonist rimonabant caused depressive-like effects | Male Wistar rats | [ref. 216] | |
| CB2 inhibition increases aggression, preferentially in female mice | CB2 receptor KO mice [286]Sprague–Dawley rats [287] | [ref. 286, ref. 287] | |
| CB1 receptor KO mice exhibited increased context dependent-anxiety-like behavior, sociability impairment and aggression | CB1 receptor KO mice | [ref. 317] | |
| Excessive AEA levels can activate TRPV1 receptors and exacerbate anxiety | TRPV1 receptor KO C57BL/6 J mice | [ref. 197] | |
| Chronic administration of CB1 receptor agonists during adolescence but not during adulthood impairs emotional behaviour and monoaminergic neurotransmission | Sprague–Dawley rats | [ref. 243] | |
| Disruptions in the functional activity of mGlu5Rs may underpin synaptic dysfunction and behavioral deficits in ASD | Shank 3 knockout mice | [ref. 249] | |
| PPARγ downregulation enhances fear, emotional response to acute stress and exacerbates anxiety | PPARγNestinCre KO mice [253]C57BL/6 J mice [255] | [ref. 253, ref. 255] | |
| PPARα downregulation enhances fear, emotional response to acute stress and exacerbates anxiety | PPARα KO mice [254]C57BL/6 J mice [255] | [ref. 254, ref. 255] | |
| Abnormal COX2/PGE2 signaling in COX2 deficient mice leads to autism-related behaviors, with males showing hyperactivity, anxiety, and repetitive behaviors, and females exhibiting social impairments | COX2-deficient mice | [ref. 274, ref. 275] | |
| COX2-deficiency leads to autism-related behaviors, such as hyperactivity and atypical communication | Children with ASD | [ref. 278] | |
| Misuse of misoprostol (a PGE2 analogue) during early pregnancy is linked to a higher incidence of neurodevelopmental disorders, including Moebius syndrome and ASD | Children with ASD | [ref. 279–ref. 281] | |
| Altered AEA degradation, due to decreased FAAH activity in lymphocytes is associated with attention-deficit/hyperactivity disorder (ADHD) | Adolescences with ADHD | [ref. 295] | |
| CB1 receptor blockade increases anxiety when interfering with glutamatergic function | CD1 mice and Wistar rats | [ref. 220] | |
| Striatal CB1 receptors on GABA terminals are insensitive in ADHD and do not respond to their agonists | DAT–CI mice | [ref. 296] | |
| FAAH and MAGL inhibition causes ADHD-like behaviors | SHR rats | [ref. 298] | |
| mGluR5 activation induces long-term depression at presynaptic sites, reducing synaptic transmission and contributing to neuronal dysfunction in the non-activated pathway | Visual cortical slices from mice | [ref. 306] | |
| DAGLα and NAPE-PLD enzymes, CB1, CB2 and oxytocin receptors are downregulated in OCD | DAT heterozygous rats with compulsive behaviors and individuals with OCD | [ref. 309] | |
| Therapeutic | Increasing anandamide (AEA) signaling, through anandamide-deactivating enzyme FAAH inhibition ameliorates social disabilities | Male Wistar rats [16]Unspecified male mice [53]BTBR and fmr1−/− mice [53]VPA-mice [52, 237, 238] | [ref. 16, ref. 52, ref. 53, ref. 237, ref. 238] |
| AEA-mediated signaling at CB1 receptors, driven by oxytocin, controls social reward and improves social impairments | C57Bl6 J male mice [204]BTBR and fmr1−/− mice [53] | [ref. 53, ref. 204] | |
| CB1 receptor blockade reduces anxiety when targeting GABAergic pathways | CD1 mice and Wistar rats | [ref. 220] | |
| Low doses of the CB1 receptor antagonist rimonabant and the neutral antagonist NESS0327 normalize memory deficits and cognitive impairments | Fmr1 KO mice | [ref. 49] | |
| Increasing eCB signaling via CB1 activation reduces aggression, and stress-induced impulsivity | Male C57BL/6 JNarl mice [217]Male CB1 KO [283] | [ref. 217, ref. 283] | |
| CBD rescues cognitive deficits by enhancing short-term memory through GPR55 receptor activation in the hippocampus and modulating anandamide metabolism | Fmr1-Δexon 8 rats | [ref. 318] | |
| Beta-caryophyllene-induced CB2R activity mediates antidepressant, and memory-enhancing effects | Wistar rats | [ref. 232] | |
| CB2 receptor activity leads to the upregulation of PGC-1α and PPAR-γ activity, exerting anxiolytic and anti-inflammatory effects | Wistar rats | [ref. 232] | |
| CB1 receptor agonists exert antidepressant effects | Male Long-Evans rats [240]Male BALB/c mice [241]Male Sprague–Dawley rats [242] | [ref. 240–ref. 242] | |
| FAAH inhibition improves aversion memory and relieves anxiety behavior | Wistar Kyoto rats [236]VPA-mice [52] | [ref. 52, ref. 236] | |
| Activation of the endocannabinoid system via CB1 receptor agonists, FAAH inhibitors, or eCB uptake inhibitors exerts antidepressant-like effects and enhances the efficacy of traditional antidepressants | Male Wistar rats | [ref. 216] | |
| 2AG signaling enhanced by a monoacylglycerol lipase (MAGL) inhibitor reduces aggressiveness and increases victimization | Male CD1 mice | [ref. 284] | |
| CBD decreases social isolation-induced aggressive behaviors potentially by increasing the activation of 5-HT1 A and CB1 receptors | Swiss mice | [ref. 285] | |
| Acute administration of CB2 agonism significantly reduced isolation-induced aggressivity | OF1 mice | [ref. 286] | |
| 2AG signaling enhanced by a MAGL inhibitor reduces OCD-like behavior | Sprague–Dawley rats | [ref. 287] | |
| Increasing AEA signaling, through FAAH inhibition ameliorates communication deficits | VPA-mice | [ref. 52] | |
| Increasing AEA signaling, through FAAH inhibition ameliorates repetitive behavior | VPA-mice | [ref. 238] | |
| Dual FAAH/TRPV1 show more anxiolytic effects than selective FAAH inhibitors alone | TRPV1-KO C57BL/6 J mice | [ref. 197] | |
| Selective inhibitors of MAGL improve repetitive and stereotypical behaviors, hyperactivity, and anxiety-like behavior | VPA mice [47]Fmr1−/− mice on a C57BL/6 J [246]Fmr1 KO mice [247] | [ref. 47, ref. 246, ref. 247] | |
| Increasing 2-AG levels through a selective inhibitor of MAGL improves sociability, social preference, and cognitive functioning | VPA mice | [ref. 47] | |
| Selective inhibitors of MAGL improve sensory behavior | Fmr1 KO mice | [ref. 247] | |
| CB1 receptor antagonim improved memory impairments | Fmr1 KO mice | [ref. 49] | |
| Palmitoylethanolamide (PEA) supplementation improves repetitive and stereotyped behaviors and sociability | BTBR T + tf/J mice [54]VPA mice [250]BTBR T + tf/J mice [54] | [ref. 54, ref. 250] | |
| PEA supplementation enhances language skills, and improves sensory sensitivity and sociability | Children with autism | [ref. 251] | |
| PEA reduces repetitive behavior and increases sociability while mitigating GI inflammatory disorders and intestinal permeability | BTBR T + tf/J mice | [ref. 54] | |
| Amygdala PPARγ activity reverses emotional response to acute stress and anxiety | PPARγNestinCre KO mice | [ref. 253] | |
| GPR55 receptor agonists exert anxiolytic-like effects in the medial orbital cortex of mice with acute stress | Male C57BL mice | [ref. 256] | |
| PPAR-α antagonists significantly attenuate social impairment, repetitive behavior, hyperactivity, anxiety, and low exploratory activity in environmental models of ASD | VPA mice [257]PPA-treated rats [258] | [ref. 257, ref. 258] | |
| Increased 2-AG levels normalize sensory behavior and improve anxiety-like and hyperactivity behaviors | Fmr1 KO mice | [ref. 247] | |
| A CB1 receptor inverse agonist (SLV-319) causes a reduction of ADHD-like behaviors such as hyperactivity and anxiety | SHR rats | [ref. 298] | |
| Endocannabinoids and CB1 receptors (activation) upregulate heterosynaptic long-term depression, contributing to synaptic plasticity. BDNF prevents the suppressive effects of eCBs, maintaining transmitter release at presynaptic sites | Visual cortical slices from mice | [ref. 306] | |
| BDNF triggers the release of eCB, which act on CB1 receptors at presynaptic GABA terminals to regulate inhibitory synaptic transmission | Neocortical slices from mice | [ref. 307] | |
| eCB and CB1 receptors activation facilitate long-term potentiation at pyramidal neurons, contributing to synaptic plasticity. BDNF enhances this process, promoting sensory information processing | Thalamocortical slices from rats | [ref. 308] |
Summary of key findings on the degenerative and therapeutic role of the endocannabinoidome (eCBome) in various behavioral and neuropsychiatric conditions occurring in autism spectrum disorder (ASD). The table outlines the observed eCBome alterations in different animal models and human samples, highlighting behavioral, biochemical, and functional changes. The table also provides information about the findings, model, and references
eCB endocannabinoid, CBR cannabinoid receptor, AEA anandamide, CB1 Cannabinoid receptor 1, CB2 cannabinoid receptor 2, TRPV1 transient receptor potential vanilloid 1, mGlu5R metabotropic glutamate receptor 5, PPARγ peroxisome proliferator-activated receptor gamma, PPARα peroxisome proliferator-activated receptor alpha, COX2 cyclooxygenase 2, PGE2 prostaglandin E2, FAAH fatty acid amide hydrolase, MAGL monoacylglycerol lipase, OCD obsessive–compulsive disorder, 2-AG 2-arachidonoylglycerol, GABA gamma-aminobutyric acid, GPR55 G-protein-coupled receptor 55, BDNF brain-derived neurotrophic factor, SHR spontaneously hypertensive rat, DAT–CI dopamine transporter–cocaine interaction, KO knockout, VPA valproic acid, BTBR brain-targeted receptor, Fmr1 fragile X mental retardation 1, OF1 outbred field mice, SLV-319 selective ligand for variant 319, PEA palmitoylethanolamide
Interestingly, gut microbiota may further contribute to ASD-related disruptions in CB1 signaling. In clinical studies, lower oxytocin serum levels in ASD correlate with increased relative abundance of Clostridia genera [ref. 221]. These bacteria, along with Desulfovibrio (also elevated in ASD) [ref. 198, ref. 222], are propionate-producing bacteria [ref. 223]. Excessive levels of propionate induce dysbiosis, neurotoxicity, and ASD-like behaviors [ref. 224–ref. 226], potentially by altering neurotransmitters such as dopamine, GABA, and serotonin [ref. 227–ref. 230]. Intriguingly, blocking CB1 receptors in the gut increases propionate-producing bacteria, though it also elicited beneficial effects on metabolic syndrome [ref. 231].
Beyond CB1, CB2 receptors have emerged as a promising therapeutic target for treating anxiety, depression, and memory deficits in ASD, along with the associated metabolic comorbidities. Specifically, beta-caryophyllene, an agonist of CB2 and PPAR-γ receptors, improved not only metabolic parameters but also psychiatric and behavioral symptoms by exerting anti-inflammatory effects in the PFC of rats with neuroinflammation and metabolic syndrome [ref. 232].
Anxiety and depression often involve an overexpression of FAAH in the amygdala and hippocampus [ref. 214–ref. 216]. Indeed, in animal models of ASD, FAAH inhibitor improved aversion memory and relieved anxiety behavior [ref. 233] and depression [ref. 216, ref. 234–ref. 236], reversed social impairment [ref. 52, ref. 53, ref. 237, ref. 238], communication deficits, stereotyped behavior [ref. 52], and repetitive and atypical emotion-related behaviors [ref. 238]. However, excessive AEA levels can activate TRPV1 receptors, which may paradoxically exacerbate anxiety. This explains why dual FAAH/TRPV1 blockers have shown more consistent anxiolytic effects than selective FAAH inhibitors alone [ref. 239].
Thus, we speculate that the abundance of propionate-producing bacteria and the downregulation of CB1 receptors in the gut, along with decreased concentrations of OEA and AEA, may contribute to ASD symptoms and pathophysiology. Targeting this axis and modulating it with eCBome enzyme/receptor (such as FAAH/TRPV1) blockers may have therapeutic potential in ASD-related anxiety and depressive tendencies as well as behavioral and sociability impairment (Fig. 1).
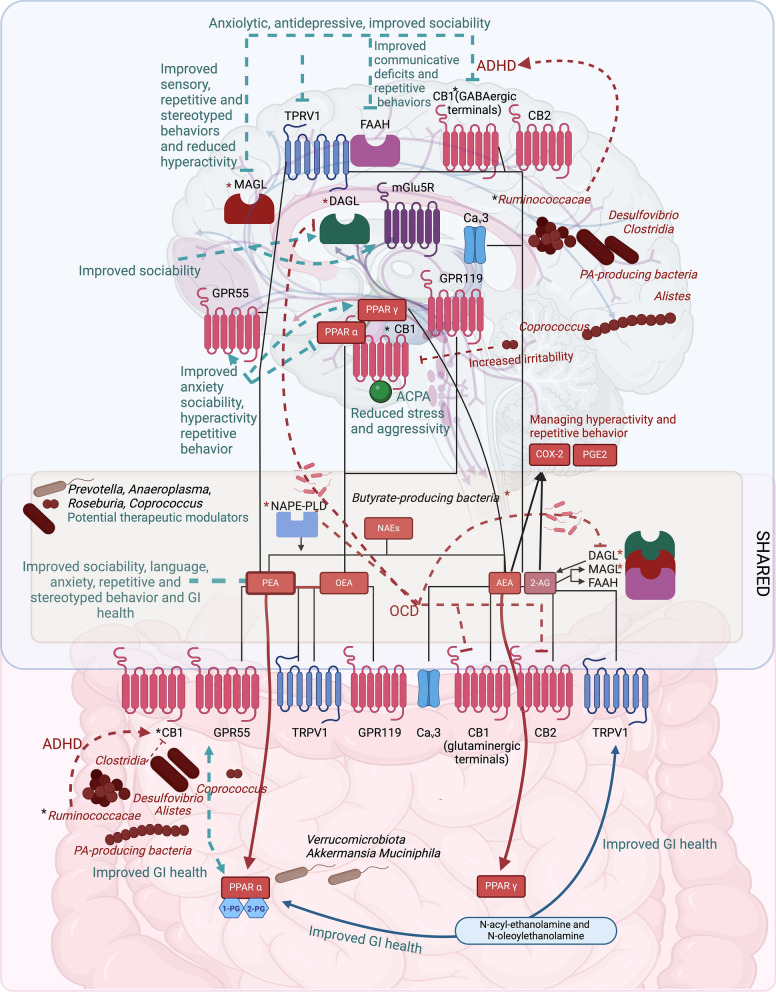
Preclinical studies further suggest that CB1 receptor agonists exert antidepressant effects [ref. 240–ref. 242], though findings in adolescent rats remain controversial [ref. 243]. CB2 receptor agonists such as cannabidiol (CBD) may also hold therapeutic value for ASD (see Supplementary Table 1). Overall, the differential roles of CB1 in ASD can be understood in terms of regional and neurotransmitter-specific mechanisms. CB1 activation may be beneficial for social behavior, aggression, and reward processing, whereas CB1 inhibition could be useful for hyperactivity-related symptoms (e.g., ADHD-like behaviors). Meanwhile, CB2 activation, FAAH inhibition, and dual FAAH/TRPV1 blockade may expand the therapeutic potential of eCB-based interventions in ASD.
In addition to OEA and AEA, palmitoylethanolamide (PEA) and 2-arachidonoylglycerol (2-AG) are also found at reduced levels in serum [ref. 29, ref. 244], while CB2 receptor expression is increased in peripheral blood mononuclear cells in individuals with ASD [ref. 245]. Selective inhibitors of monoacylglycerol lipase (MAGL), an enzyme that breaks down 2-AG, improved repetitive and stereotypical behaviours, hyperactivity, anxiety-like behavior [ref. 246, ref. 247], sociability, social preference, cognitive functioning [ref. 47], and sensory behavior [ref. 247].
In the Fmr1 model of ASD, which recapitulates fragile X syndrome (FXS), a failure of diacylglycerol Lipase α (DAGL-α), the main enzyme responsible for synthesizing 2-AG, was associated with metabotropic glutamate 5 receptor (mGlu5R) deficits [ref. 248]. Additionally, disruptions in the functional activity of mGlu5Rs was observed in the Shank 3 knockout model of ASD [ref. 249]. We speculate that increasing mGlu5R activity may enhance the activity of DAGL, thereby raising 2-AG levels, and exerting beneficial effects on ASD symptoms.
Studies on PEA demonstrate its dual effects on both the gut and brain: its supplementation improved repetitive and stereotyped behaviors and sociability in ASD models [ref. 54, ref. 250], enhanced language skills, and improved sensory sensitivity and sociability in children with autism [ref. 251], while mitigating GI inflammatory disorders and intestinal permeability [ref. 54]. These effects are possibly elicited through activity at its proposed targets, i.e. G protein-coupled receptor 55 (GPR55) [ref. 252], PPAR-α [ref. 54], and indirectly via enhancement of eCB levels or activity [ref. 252]. Two clinical trials administering PEA are ongoing (NCT06187090) and, of which one (NCT05182697) administers PEA in combination with cannabidiol to obtain an “entourage effect”. Brain PPAR-γ and PPAR-α are implicated in emotional stress and anxiety in preclinical models [ref. 253–ref. 255]. GPR55, which is activated by both eCB and non-eCB ligands, plays a role in anxiety regulation [ref. 256], suggesting a broader involvement of the eCBome in anxiety modulation. Interestingly, treatment with a PPAR-α antagonist significantly attenuated social impairment, repetitive behavior, hyperactivity, anxiety, and low exploratory activity in environmental models of ASD [ref. 257, ref. 258]. Thus, we speculate that the therapeutic effects of PEA can be potentiated through the combination of PEA and modulators of GPR55, PPAR-γ, and PPAR-α activity.
Modulation of PPAR-α and TRPV1 also shows therapeutic potential for treating GI disorders such as gut dysbiosis, permeability issues, and systemic inflammation, which are often comorbid in ASD. This modulation can be achieved through the respective ligands of these receptors, such as PEA, N-oleoylethanolamine, as well as the TRPV1 agonist capsaicin [ref. 54, ref. 259–ref. 261]. Interestingly, in FXS, the concentrations of eCBs was not altered; however, the receptors and enzymes involved in the metabolism of eCBs, such as FAAH, MAGL, CB1 on GABAergic terminals, and mGlu5R, were affected [ref. 246, ref. 247].
In conditions like ASD and FXS, increased levels of mucin-degrading bacteria (e.g., Verrucomicrobiota and Akkermansia) can serve as biomarkers for gut barrier integrity, permeability, and inflammation [ref. 33, ref. 36, ref. 37, ref. 39]. Higher levels of these bacteria may impair hippocampal function and cognitive processes related to neuronal development [ref. 262, ref. 263], potentially exacerbating psychiatric symptoms through a disrupted gut–brain axis [ref. 38]. Interestingly, while the species A. muciniphila has been associated with beneficial effects in restoring gut barrier function and reducing intestinal permeability, it also possesses inflammatory properties due to increased exposure of immune cells to microbial antigens when the mucosal layer is compromised [ref. 36]. This double-edged role may contribute to inflammatory conditions, as suggested not only in ASD but also in neurodegenerative diseases like multiple sclerosis and Parkinson’s disease [ref. 36]. Nevertheless, in a mouse model of metabolic syndrome, supplementation with A. muciniphila improved gut permeability and increased the levels of 2-AG and its mono-acyl-glycerol congeners [ref. 264]. In obese and overweight men, supplementation of this bacterium elevated the circulating levels of eCBome mediators, 1-palmitoyl-glycerol (1-PG) and 2-palmitoyl-glycerol (2-PG), which bind to PPAR-α receptors with beneficial effects on lipid metabolism and energy regulation [ref. 265]. Positive modulation of PPAR-α and TRPV1 by eCBome-active GM compounds could thus offer therapeutic benefits in treating ASD and associated GI and metabolic symptoms.
Additionally, fecal microbial transplantation from wild-type mice into Fmr1−/y mice increased A. muciniphila levels, while simultaneously ameliorating cognitive and social deficits [ref. 266]. This raises uncertainty about whether A. muciniphila is strictly beneficial or detrimental in the context of the Fmr1 mutation. At any rate, the mucin-degrading activity of A. muciniphila might not necessarily imply a negative role in neurodegenerative and neuropsychiatric disorders [ref. 36], as such activity might become pathological only after that other more pathogenic bacteria have already compromised the mucosal intestinal barrier, as suggested by the fact that the relative abundance of this species is decreased in obesity [ref. 267].
Therefore, Verrucomicrobiota, and specifically A. muciniphila, can modulate eCBome mediator levels. This makes them both potential biomarkers and therapeutic targets for ASD-related symptoms and GI pathophysiology.
In the valproic acid-exposed and BTBR models of ASD, treatment with butyrate improved recognition memory and social behavior [ref. 166, ref. 167]. Butyrate modulates eCB tone in gut epithelial cells by regulating enzymes involved in eCBs synthesis and breakdown, such as N-acyl-phosphatidylethanolamine-specific phospholipase D (NAPE-PLD), DAGL-α, MAGL [ref. 268] and FAAH [ref. 269]. This modulation indicates that excessive butyrate levels may alter eCBs levels in ASD. However, clinical studies have reported conflicting findings regarding the abundance of butyrate-producing bacteria and fecal butyrate levels in ASD populations [ref. 270–ref. 272], mostly concerning Faecalibacteria and Faecalibacterium prausnitzii concentrations. Further research is needed to determine whether modulating butyrate-producing bacteria is beneficial or detrimental for addressing the dysregulated eCBome–GM–brain axis in ASD.
Endogenous cannabinoids are substrates for cyclooxygenase-2 (COX-2) and can be oxygenated by COX-2 to form new classes of prostaglandins [ref. 273]. The signaling of two eCBome mediators, prostaglandin E2 (PGE2) and COX-2, is altered in ASD [ref. 274]. As PGE2 plays a crucial role in brain development, abnormalities in the COX-2/PGE2 pathway have been linked to ASD [ref. 274, ref. 275]. Studies report lower arachidonic acid levels alongside increased COX-2 and PGE2 in the blood plasma of individuals with ASD [ref. 276, ref. 277], correlating these changes with sensory abnormalities [ref. 277]. Polymorphisms in the PTGS2 gene, which encodes COX-2, have also been associated with hyperactivity and atypical communication in ASD [ref. 278]. Misuse of misoprostol (a PGE2 analogue) during early pregnancy is linked to a higher incidence of neurodevelopmental disorders, including Moebius syndrome and ASD [ref. 279–ref. 281]. Moreover, COX-1/2 deficient mice display ASD-like behaviors, such as hyperactivity, repetitive behaviors, and atypical social interaction [ref. 274], alongside disruptions in ASD-related biological pathways [ref. 275]. Therefore, the COX-2/PGE2 pathway also represents a viable potential therapeutic target in ASD.
The eCBome–GM–brain axis in ASD, irritability and aggressivity
The challenges faced by individuals with ASD in articulating their own mental and emotional states, recognizing those of others, and coping with sensory sensitivity, unmet needs, and societal criticism often lead to discrimination and social isolation [ref. 282], as well as increased irritability, meltdowns, tantrums, or self-injurious behaviors [ref. 283].
In preclinical models, social isolation disrupts brain development, particularly in the ventral hippocampus (vHPC), increasing excitatory activity and exacerbating stress-induced impulsive aggression [ref. 217]. Activation of the CB1 receptor by agonists like arachidonylcyclopropylamide reduces aggression in stress tests without affecting overall activity [ref. 217]. CB1 activation also elevates levels of eCBs such as AEA and 2-AG in the vHPC, and by inhibiting c-Fos expression, mitigates biting, offensive behavior, and impulsive aggression in rodents [ref. 217–ref. 219]. Additionally, in preclinical models, enhancing 2-AG signaling through MAGL inhibition reduced aggressiveness while increasing victimization [ref. 284], and cannabidiol decreased social isolation-induced aggressive behaviors likely by activating both CB1 (indirectly) and 5-HT1 A receptors [ref. 285]. Interestingly, also acute administration of the CB2 agonist significantly reduced isolation-induced aggressivity [ref. 286], while CB2 receptor downregulation showed increased aggressivity [ref. 286, ref. 287]. The findings suggest that disruptions in social interaction and isolation contribute to heightened aggression and altered brain activity, particularly in the vHPC, and that targeting the endocannabinoid eCBome—via CB1 and CB2 receptor activation or MAGL inhibition—may mitigate these effects, highlighting potential therapeutic strategies for managing aggression and social deficits in ASD.
Clostridium, a bacterium whose abundance is negatively associated with CB1 and CB2 activity [ref. 288], correlates positively with irritability in ASD symptoms [ref. 289, ref. 290]. At the neuronal level, the gut abundance of Clostridium, and specifically the taxa Coprococcus, shows negative associations with changes in critical analysis and reasoning skills in children with ASD [ref. 115]. Therefore, CB1 activation can potentially suppress impulsive aggression by increasing AEA and 2-AG [ref. 217–ref. 219] and reducing Clostridium abundance [ref. 288]. The eCB system appears to regulate both neuronal activity and GM composition related to irritability and aggression, suggesting it could be a therapeutic target for individuals with ASD experiencing aggressive or self-injurious behaviors.
While current treatments like probiotics for ASD often include Streptococcus strains [ref. 291], these taxa are found to be significantly increased in children with ASD and irritability. Another overabundant bacterium in ASD-related irritability is Alistipes [ref. 289], a SCFA-producing bacterium primarily generating propionate and acetate, which may contribute to neurotoxicity and dysbiosis [ref. 225]. Notably, acetate administration is used to model ASD epigenetically due to its neurotoxic effects. Furthermore, propionic acidemia induces ASD-like behaviors, possibly by altering neurotransmitters such as dopamine [ref. 227, ref. 292, ref. 293]. Inhibition of CB1 receptors has been shown to increase levels of both propionate-producing bacteria and dopamine [ref. 231]. Hence, targeting the negative interaction between CB1 and Alistipes may hold promise as a therapeutic approach for managing ASD-related aggressivity and related comorbid behaviors.
The eCBome–GM–brain axis in ASD, ADHD and OCD
The role of the eCBome as a therapeutic target in ADHD remains unclear. ADHD is associated with dysregulated dopamine signaling [ref. 294], which may affect eCB levels [ref. 295]. Increased AEA levels have been observed in ADHD, possibly due to hyperactive dopamine D2 receptors [ref. 295]. Preclinical models of ADHD display a loss of striatal CB1 receptors on GABAergic terminals and a dysfunctional FAAH polymorphism [ref. 296, ref. 297]. Interestingly, MAGL and FAAH inhibition in rats induced ADHD-like behaviors, which were reversed by a CB1 receptor inverse agonist [ref. 298]. Conversely, MAGL inhibition in models with hyperactivity, such as Fmr1 mice, restored eCB-mediated synaptic plasticity, and reduced hyperactivity, anxiety, and other ASD behaviors [ref. 47, ref. 246, ref. 247]. Additionally, CB1 antagonists improved memory impairments in the Fmr1 model of ASD [ref. 49].
Also, treatment with a PPAR-α antagonist significantly attenuated hyperactivity, anxiety, and other ASD behaviors in models of ASD [ref. 257, ref. 258].
Short chain fatty acids production alterations are implicated in ADHD. Among SCFAs, butyrate modulates the expression of eCBome enzymes such as MAGL, NAPE-PLD and DAGL in a dose-dependent manner [ref. 268], as well as modulating neurotransmitter (e.g., GABA) levels [ref. 166] and CB1 receptors signaling [ref. 299].
In ASD with ADHD, there is increased abundance of Odoribacter, particularly O. splanchnicus, whereas species from the Ruminococcaceae family, known for butyrate production, like F. prausnitzii, are decreased. F. prausnitzii has been negatively associated with ADHD severity and hyperactivity [ref. 300]. However, Ruminococcacae_UGC was positively correlated with the risk of ADHD and with lack of attention [ref. 301, ref. 302], potentially due to its association with microbial species capable of affecting GABA neurotransmitter levels [ref. 303]. We therefore speculate that alterations in SCFA levels might affect the availability of the eCBs AEA and 2-AG, impacting synaptic plasticity and neurotransmitters implicated in ADHD pathophysiology.
Variation of a genus from the Lachnospiraceae family (Coprococcus) showed a trend of being negatively associated with inattention symptoms [ref. 301, ref. 302]. The genera Butyricicoccus, Roseburia, Desulfovibrio, Lachnospiraceae NC2004 group, and Romboutsia were protective factors for ADHD, with the abundance of the genus Butyricicoccus being negatively correlated with the risk of ADHD [ref. 302]. The differential abundance of gut bacteria in ADHD, as reviewed elsewhere [ref. 303], shows distinct patterns from that observed in ASD. This suggests unique microbial signatures associated with either condition. Together, further research is needed to uncover the interplay between SCFA-producing bacteria and the eCBome in ADHD comorbid with ASD.
Obsessive–compulsive disorder involves complex neurobiological mechanisms, including altered brain-derived neurotrophic factor (BDNF) expression and oxytocin signaling [ref. 304, ref. 305], both of which interact with the eCBome [ref. 204, ref. 306–ref. 309]. In a translational study on blood samples from individuals with OCD and OCD rat models, downregulation of enzymes responsible for eCB biosynthesis, such as DAGLα and NAPE-PLD, was correlated with reduced oxytocin receptor expression. Moreover, CB1 and CB2 receptor expression was downregulated, with a significant inverse correlation observed between BDNF and CB2 gene expression [ref. 309]. These findings underscore intricate relationships between the eCBome, BDNF, and oxytocin signaling in OCD and its co-occurrence with ASD.
Indeed, as mentioned above, many eCBome components have beneficial effects in ASD co-occurring with OCD behavior. Treatment with a PPAR-α antagonist and PEA significantly attenuated repetitive [ref. 54, ref. 250, ref. 257, ref. 258] and stereotyped behaviors in ASD models [ref. 54, ref. 250], while COX-1/2 deficient mice display repetitive behaviors alongside ASD-like behaviors [ref. 274, ref. 275].
Differences in GM taxonomic composition, particularly with regard to SCFA-producing bacteria, such as Lachnospiraceae and Ruminococcaceae, have been noted in OCD models and ASD [ref. 310, ref. 311]. Specifically, genera such as Anaerostipes and Odoribacter were reduced in individuals with OCD, while in the Fmr1 mouse model of ASD they were over-abundant [ref. 33, ref. 310]. The genus Oscillospira instead was under-represented in OCD models, mirroring observations in the Fmr1 knockout mouse model [ref. 310, ref. 312].
In a preclinical model of OCD, a lower abundance of Prevotella and Anaeroplasma, both known for their anti-inflammatory properties, was observed [ref. 313], consistent with gut microbiota composition findings in ASD [ref. 314]. Youth suffering from pediatric autoimmune neuropsychiatric disorders associated with streptococcal infections (PANDAS), which exhibit OCD-like symptoms [ref. 315], showed a decrease in the genera Roseburia and Coprococcus, but an increase of Odoribacter and Oscillospira [ref. 316], contrasting with previous studies in OCD individuals [ref. 310]. The genera Roseburia and Coprococcus are known for their protective properties for gut health and are also reduced in ASD and FXS. Kamble and Dandekar reviewed a wide spectrum of bacteria involved in OCD, considering its comorbidity with other psychiatric disorders, although they did not discuss specific links to ASD [ref. 316].
Conclusions
The eCBome–GM–brain axis is emerging as a promising therapeutic target for ASD and its systemic and psychiatric comorbidities. Dysregulated eCB levels, particularly reduced OEA and AEA concentrations and altered CB1 receptor activity, may contribute to ASD symptoms. Modulating this axis, through inhibitors of FAAH, TRPV1, and MAGL holds potential for sociability impairments, depression and anxiety. However, inhibition of FAAH and MAGL may induce ADHD-like behaviors, which could be reversed with a CB1 receptor inverse agonist. Conversely, MAGL inhibition may restore eCB-mediated synaptic plasticity, reduce hyperactivity, anxiety, aggressivity and improve repetitive, stereotypical, and sensory behaviors. Targeting mGlu5R to elevate 2-AG through DAGL activation could also provide therapeutic benefits in ASD, while the COX-2/PGE2 pathway may serve as another target to manage hyperactivity and repetitive behaviors. Supplementation with PEA has shown promise in improving repetitive and stereotyped behaviors, language skills, and GI symptoms, including intestinal permeability and inflammation. GPR55, PPAR-γ, and PPAR-α receptors may hold further therapeutic promise within this axis, particularly through modulation by PEA. Targeting CB1 and CB2 receptor activation may mitigate aggressivity in ASD.
Mucin-degrading bacterial taxa represent potential targets for managing ASD symptoms and GI pathophysiology. Selective antibiotics against specific Clostridium strains may improve irritability and aggression. Changes in SCFAs levels, for example through prebiotics, may influence eCB availability, impacting synaptic plasticity and neurotransmitter systems. Anti-inflammatory bacteria like Prevotella and Anaeroplasma, alongside gut health biomarkers such as Roseburia and Coprococcus, may offer therapeutic value. Conflicting findings regarding butyrate-producing bacteria in ASD populations suggest caution when using these bacteria as targets. Further research is needed to elucidate their role in modulating the eCBome–GM–brain axis in ASD.
In conclusion, the eCBome–GM–brain axis represents a promising, multifaceted therapeutic target for ASD and its comorbidities, warranting further clinical and preclinical research to clarify its therapeutic potential and refine targeted interventions.
Supplementary Materials
References
- MJ Maenner, Z Warren, AR Williams, E Amoakohene, AV Bakian, DA Bilder. Prevalence and characteristics of autism spectrum disorder among children aged 8 years—autism and developmental disabilities monitoring network, 11 sites, United States, 2020. MMWR Surveill Summ, 2023. [DOI | PubMed]
- C Lord, M Elsabbagh, G Baird, J Veenstra-Vanderweele. Autism spectrum disorder. The lancet, 2018. [DOI]
- BWJH Penninx, SMM Lange. Metabolic syndrome in psychiatric patients: overview, mechanisms, and implications. Dialogues Clin Neurosci, 2018. [DOI | PubMed]
- EK Nousen, JG Franco, EL Sullivan. Unraveling the mechanisms responsible for the comorbidity between metabolic syndrome and mental health disorders. Neuroendocrinology, 2013. [DOI | PubMed]
- T Barlattani, C D’Amelio, A Cavatassi, D De Luca, R Di Stefano, A Di Berardo. Autism spectrum disorders and psychiatric comorbidities: a narrative review. Journal of Psychopathology, 2023
- MF Casanova, RE Frye, C Gillberg, EL Casanova. Comorbidity and autism spectrum disorder. Front Psych, 2020
- SM Bury, D Hedley, M Uljarević. Restricted, repetitive behaviours and interests in the workplace: barriers, advantages, and an individual difference approach to autism employment. Repetitive and restricted behaviors and interests in autism spectrum disorders: from neurobiology to behavior, 2021
- D Hedley, M Uljarević, M Wilmot, A Richdale, C Dissanayake. Understanding depression and thoughts of self-harm in autism: a potential mechanism involving loneliness. Res Autism Spectr Disord, 2018. [DOI]
- SM Bury, RL Flower, R Zulla, DB Nicholas, D Hedley. Workplace social challenges experienced by employees on the autism spectrum: an international exploratory study examining employee and supervisor perspectives. J Autism Dev Disord, 2021. [DOI | PubMed]
- AA Pallathra, L Cordero, K Wong, ES Brodkin. Psychosocial interventions targeting social functioning in adults on the autism spectrum: a literature review. Curr Psychiatry Rep, 2019. [DOI | PubMed]
- OD Howes, M Rogdaki, JL Findon, RH Wichers, T Charman, BH King. Autism spectrum disorder: consensus guidelines on assessment, treatment and research from the British Association for Psychopharmacology. J Psychopharmacol, 2018. [DOI | PubMed]
- S Pietropaolo, G Marsicano. The role of the endocannabinoid system as a therapeutic target for autism spectrum disorder: lessons from behavioral studies on mouse models. Neurosci Biobehav Rev, 2022. [DOI | PubMed]
- A Mehra, G Arora, G Sahni, M Kaur, H Singh, B Singh. Gut microbiota and autism spectrum disorder: from pathogenesis to potential therapeutic perspectives. J Tradit Complement Med, 2023. [DOI | PubMed]
- V Di Marzo. The endocannabinoidome as a substrate for noneuphoric phytocannabinoid action and gut microbiome dysfunction in neuropsychiatric disorders. Dialogues Clin Neurosci, 2020. [DOI | PubMed]
- S Zou, U Kumar. Cannabinoid receptors and the endocannabinoid system: signaling and function in the central nervous system. Int J Mol Sci, 2018. [DOI | PubMed]
- V Trezza, R Damsteegt, A Manduca, S Petrosino, LW Van Kerkhof, RJ Pasterkamp. Endocannabinoids in amygdala and nucleus accumbens mediate social play reward in adolescent rats. J Neurosci, 2012. [DOI | PubMed]
- N Acharya, S Penukonda, T Shcheglova, AT Hagymasi, S Basu, PK Srivastava. Endocannabinoid system acts as a regulator of immune homeostasis in the gut. Proc Natl Acad Sci USA, 2017. [DOI | PubMed]
- D Siniscalco, S Schultz, AL Brigida, N Antonucci. Inflammation and neuro-immune dysregulations in autism spectrum disorders. Pharmaceuticals, 2018. [DOI | PubMed]
- PD Cani, H Plovier, M Van Hul, L Geurts, NM Delzenne, C Druart. Endocannabinoids—at the crossroads between the gut microbiota and host metabolism. Nat Rev Endocrinol, 2016. [DOI | PubMed]
- SS Habib, K Al-Regaiey, S Bashir, M Iqbal. Role of endocannabinoids on neuroinflammation in autism spectrum disorder prevention. J Clin Diagn Res, 2017. [PubMed]
- M Zou, D Li, L Li, L Wu, C Sun. Role of the endocannabinoid system in neurological disorders. Int J Dev Neurosci, 2019. [DOI | PubMed]
- EA Mayer, D Padua, K Tillisch. Altered brain-gut axis in autism: comorbidity or causative mechanisms?. BioEssays, 2014. [DOI | PubMed]
- I Sekirov, SL Russell, LCM Antunes, BB Finlay. Gut microbiota in health and disease. Physiol Rev, 2010. [DOI | PubMed]
- LH Morais, HL Schreiber, SK Mazmanian. The gut microbiota–brain axis in behaviour and brain disorders. Nat Rev Microbiol, 2021. [DOI | PubMed]
- C Silvestri, V Di Marzo. The gut microbiome–endocannabinoidome axis: a new way of controlling metabolism, inflammation, and behavior. Function, 2023. [DOI | PubMed]
- C Manca, M Shen, B Boubertakh, C Martin, N Flamand, C Silvestri. Alterations of brain endocannabinoidome signaling in germ-free mice. Biochim Biophys Acta Mol Cell Biol Lipids, 2020. [DOI | PubMed]
- C Manca, B Boubertakh, N Leblanc, T Deschênes, S Lacroix, C Martin. Germ-free mice exhibit profound gut microbiota-dependent alterations of intestinal endocannabinoidome signaling. J Lipid Res, 2020. [DOI | PubMed]
- M De Pol, NJ Kolla. Endocannabinoid markers in autism spectrum disorder: a scoping review of human studies. Psychiatry Res, 2021. [DOI | PubMed]
- A Aran, M Eylon, M Harel, L Polianski, A Nemirovski, S Tepper. Lower circulating endocannabinoid levels in children with autism spectrum disorder. Mol Autism, 2019. [DOI | PubMed]
- F Liu, J Li, F Wu, H Zheng, Q Peng, H Zhou. Altered composition and function of intestinal microbiota in autism spectrum disorders: a systematic review. Transl Psychiatry, 2019. [DOI | PubMed]
- JT Morton, D-M Jin, RH Mills, Y Shao, G Rahman, D McDonald. Multi-level analysis of the gut–brain axis shows autism spectrum disorder-associated molecular and microbial profiles. Nat Neurosci, 2023. [DOI | PubMed]
- M Xu, X Xu, J Li, F Li. Association between gut microbiota and autism spectrum disorder: a systematic review and meta-analysis. Front Psychiatry, 2019. [DOI | PubMed]
- F Altimiras, JA Garcia, I Palacios-García, MJ Hurley, R Deacon, B González. Altered gut microbiota in a fragile X syndrome mouse model. Front Neurosci, 2021. [DOI | PubMed]
- N-R Shin, TW Whon, J-W Bae. Proteobacteria: microbial signature of dysbiosis in gut microbiota. Trends Biotechnol, 2015. [DOI | PubMed]
- Y Zhu, PM Carvey, Z Ling. Altered glutathione homeostasis in animals prenatally exposed to lipopolysaccharide. Neurochem Int, 2007. [DOI | PubMed]
- JR Bedarf, F Hildebrand, LP Coelho, S Sunagawa, M Bahram, F Goeser. Functional implications of microbial and viral gut metagenome changes in early stage L-DOPA-naïve Parkinson’s disease patients. Genome Med, 2017. [DOI | PubMed]
- SY Geerlings, I Kostopoulos, WM de Vos, C Belzer. Akkermansia muciniphila in the human gastrointestinal tract: when, where, and how?. Microorganisms, 2018. [DOI | PubMed]
- C Bostrom, SY Yau, N Majaess, M Vetrici, J Gil-Mohapel, BR Christie. Hippocampal dysfunction and cognitive impairment in Fragile-X Syndrome. Neurosci Biobehav Rev, 2016. [DOI | PubMed]
- BP Ganesh, R Klopfleisch, G Loh, M Blaut. Commensal Akkermansia muciniphila exacerbates gut inflammation in Salmonella Typhimurium-infected gnotobiotic mice. PLoS ONE, 2013. [DOI | PubMed]
- JL Dempsey, M Little, JY Cui. Gut microbiome: an intermediary to neurotoxicity. Neurotoxicology, 2019. [DOI | PubMed]
- SR Shultz, NA Aziz, L Yang, M Sun, DF MacFabe, TJ O’Brien. Intracerebroventricular injection of propionic acid, an enteric metabolite implicated in autism, induces social abnormalities that do not differ between seizure-prone (FAST) and seizure-resistant (SLOW) rats. Behav Brain Res, 2015. [DOI | PubMed]
- DF MacFabe, NE Cain, F Boon, K-P Ossenkopp, DP Cain. Effects of the enteric bacterial metabolic product propionic acid on object-directed behavior, social behavior, cognition, and neuroinflammation in adolescent rats: relevance to autism spectrum disorder. Behav Brain Res, 2011. [DOI | PubMed]
- WA Kandeel, NA Meguid, G Bjørklund, EM Eid, M Farid, SK Mohamed. Impact of clostridium bacteria in children with autism spectrum disorder and their anthropometric measurements. J Mol Neurosci, 2020. [DOI | PubMed]
- R Keşli, C Gökçen, U Buluğ, Y Terzi. Investigation of the relation between anaerobic bacteria genus clostridium and late-onset autism etiology in children. J Immunoassay Immunochem, 2014. [DOI | PubMed]
- C Rousseaux, X Thuru, A Gelot, N Barnich, C Neut, L Dubuquoy. Lactobacillus acidophilus modulates intestinal pain and induces opioid and cannabinoid receptors. Nat Med, 2007. [DOI | PubMed]
- FA Moreira, N Kaiser, K Monory, B Lutz. Reduced anxiety-like behaviour induced by genetic and pharmacological inhibition of the endocannabinoid-degrading enzyme fatty acid amide hydrolase (FAAH) is mediated by CB1 receptors. Neuropharmacology, 2008. [DOI | PubMed]
- M Zou, Y Liu, S Xie, L Wang, D Li, L Li. Alterations of the endocannabinoid system and its therapeutic potential in autism spectrum disorder. Open Biol, 2021. [DOI | PubMed]
- A Busquets-Garcia, M Gomis-Gonzalez, T Guegan, C Agustin-Pavon, A Pastor, S Mato. Targeting the endocannabinoid system in the treatment of fragile X syndrome. Nat Med, 2013. [DOI | PubMed]
- M Gomis-Gonzalez, A Busquets-Garcia, C Matute, R Maldonado, S Mato, A Ozaita. Possible therapeutic doses of cannabinoid type 1 receptor antagonist reverses key alterations in fragile X syndrome mouse model. Genes, 2016. [DOI | PubMed]
- A Gururajan, DA Taylor, DT Malone. Cannabidiol and clozapine reverse MK-801-induced deficits in social interaction and hyperactivity in Sprague-Dawley rats. J Psychopharmacol, 2012. [DOI | PubMed]
- JS Kaplan, N Stella, WA Catterall, RE Westenbroek. Cannabidiol attenuates seizures and social deficits in a mouse model of Dravet syndrome. Proc Natl Acad Sci USA, 2017. [DOI | PubMed]
- M Servadio, F Melancia, A Manduca, A di Masi, S Schiavi, V Cartocci. Targeting anandamide metabolism rescues core and associated autistic-like symptoms in rats prenatally exposed to valproic acid. Transl Psychiatry, 2016. [DOI | PubMed]
- D Wei, D Dinh, D Lee, D Li, A Anguren, G Moreno-Sanz. Enhancement of anandamide-mediated endocannabinoid signaling corrects autism-related social impairment. Cannabis Cannabinoid Res, 2016. [DOI | PubMed]
- C Cristiano, C Pirozzi, L Coretti, G Cavaliere, A Lama, R Russo. Palmitoylethanolamide counteracts autistic-like behaviours in BTBR T+tf/J mice: contribution of central and peripheral mechanisms. Brain Behav Immun, 2018. [DOI | PubMed]
- SW Saliba, H Jauch, B Gargouri, A Keil, T Hurrle, N Volz. Anti-neuroinflammatory effects of GPR55 antagonists in LPS-activated primary microglial cells. J Neuroinflammation, 2018. [DOI | PubMed]
- N Malek, K Popiolek-Barczyk, J Mika, B Przewlocka, K Starowicz. Anandamide, acting via CB2 receptors, alleviates LPS-induced neuroinflammation in rat primary microglial cultures. Neural Plast, 2015. [PubMed]
- A Stančić, K Jandl, C Hasenöhrl, F Reichmann, G Marsche, R Schuligoi. The GPR55 antagonist CID16020046 protects against intestinal inflammation. Neurogastroenterol Motil, 2015. [DOI | PubMed]
- H Sumida, E Lu, H Chen, Q Yang, K Mackie, JG Cyster. GPR55 regulates intraepithelial lymphocyte migration dynamics and susceptibility to intestinal damage. Sci Immunol, 2017. [DOI]
- P Luczynski, K-A McVey Neufeld, CS Oriach, G Clarke, TG Dinan, JF Cryan. Growing up in a bubble: using germ-free animals to assess the influence of the gut microbiota on brain and behavior. Int J Neuropsychopharmacol, 2016. [DOI | PubMed]
- M Jans, L Vereecke. A guide to germ-free and gnotobiotic mouse technology to study health and disease. FEBS J, 2024. [DOI | PubMed]
- Y Liu, P Forsythe. Vagotomy and insights into the microbiota–gut–brain axis. Neurosci Res, 2021. [DOI | PubMed]
- PA Muller, M Schneeberger, F Matheis, P Wang, Z Kerner, A Ilanges. Microbiota modulate sympathetic neurons via a gut–brain circuit. Nature, 2020. [DOI | PubMed]
- I Mishra, KB Pullum, KN Eads, AR Strunjas, NT Ashley. Peripheral sympathectomy alters neuroinflammatory and microglial responses to sleep fragmentation in female mice. Neuroscience, 2022. [DOI | PubMed]
- JC Park, S-H Im. Of men in mice: the development and application of a humanized gnotobiotic mouse model for microbiome therapeutics. Exp Mol Med, 2020. [DOI | PubMed]
- J Lu, L Lu, Y Yu, J Cluette-Brown, CR Martin, EC Claud. Effects of intestinal microbiota on brain development in humanized gnotobiotic mice. Sci Rep, 2018. [DOI | PubMed]
- E Avolio, I Olivito, E Rosina, L Romano, T Angelone, A De Bartolo. Modifications of behavior and inflammation in mice following transplant with fecal microbiota from children with autism. Neuroscience, 2022. [DOI | PubMed]
- S Mörkl, J Wagner-Skacel, T Lahousen, S Lackner, SJ Holasek, SA Bengesser. The role of nutrition and the gut–brain axis in psychiatry: a review of the literature. Neuropsychobiology, 2020. [DOI]
- A Skrzypczak-Wiercioch, K Sałat. Lipopolysaccharide-induced model of neuroinflammation: mechanisms of action, research application and future directions for its use. Molecules, 2022. [DOI | PubMed]
- M Rincel, M Darnaudéry. Maternal separation in rodents: a journey from gut to brain and nutritional perspectives. Proceedings of the Nutrition Society, 2020. [DOI | PubMed]
- F Dunphy-Doherty, SM O’Mahony, VL Peterson, O O’Sullivan, F Crispie, PD Cotter. Post-weaning social isolation of rats leads to long-term disruption of the gut microbiota-immune-brain axis. Brain Behav Immun, 2018. [DOI | PubMed]
- C Yang, D Merlin. Unveiling colitis: a journey through the dextran sodium sulfate-induced model. Inflamm Bowel Dis, 2024. [DOI | PubMed]
- I Silva, R Pinto, V Mateus. Preclinical study in vivo for new pharmacological approaches in inflammatory bowel disease: a systematic review of chronic model of TNBS-induced colitis. J Clin Med, 2019. [DOI | PubMed]
- S Higashida, H Nagai, K Nakayama, R Shinohara, M Taniguchi, M Nagai. Repeated social defeat stress impairs attentional set shifting irrespective of social avoidance and increases female preference associated with heightened anxiety. Sci Rep, 2018. [DOI | PubMed]
- SD Iñiguez, FJ Flores-Ramirez, LM Riggs, JB Alipio, I Garcia-Carachure, MA Hernandez. Vicarious social defeat stress induces depression-related outcomes in female mice. Biol Psychiat, 2018. [DOI | PubMed]
- B Rein, K Ma, Z Yan. A standardized social preference protocol for measuring social deficits in mouse models of autism. Nat Protoc, 2020. [DOI | PubMed]
- V Philip, N Kraimi, H Zhang, J Lu, G Palma, C Shimbori. Innate immune system signaling and intestinal dendritic cells migration to the brain underlie behavioral changes after microbial colonization in adult mice. Brain Behav Immun, 2025. [DOI | PubMed]
- R Nishino, K Mikami, H Takahashi, S Tomonaga, M Furuse, T Hiramoto. Commensal microbiota modulate murine behaviors in a strictly contamination-free environment confirmed by culture-based methods. Neurogastroenterol Motil, 2013. [DOI | PubMed]
- WH Liu, HL Chuang, YT Huang, CC Wu, GT Chou, S Wang. Alteration of behavior and monoamine levels attributable to Lactobacillus plantarum PS128 in germ-free mice. Behav Brain Res, 2016. [DOI | PubMed]
- A Parker, SA James, C Purse, A Brion, A Goldson, A Telatin. Absence of bacteria permits fungal gut-to-brain translocation and invasion in germfree mice but ageing alone does not drive pathobiont expansion in conventionally raised mice. Front Aging Neurosci, 2022. [DOI | PubMed]
- E Husebye, PM Hellström, F Sundler, J Chen, T Midtvedt. Influence of microbial species on small intestinal myoelectric activity and transit in germ-free rats. Am J Physiol Gastrointest Liver Physiol, 2001. [DOI | PubMed]
- YP Lee, CC Chiu, TJ Lin, SW Hung, WC Huang, CF Chiu. The germ-free mice monocolonization with Bacteroides fragilis improves azoxymethane/dextran sulfate sodium induced colitis-associated colorectal cancer. Immunopharmacol Immunotoxicol, 2019. [DOI | PubMed]
- CC Chiu, YH Ching, YC Wang, JY Liu, YP Li, YT Huang. Monocolonization of germ-free mice with Bacteroides fragilis protects against dextran sulfate sodium-induced acute colitis. Biomed Res Int, 2014. [DOI | PubMed]
- SC Kim, SL Tonkonogy, CA Albright, J Tsang, EJ Balish, J Braun. Variable phenotypes of enterocolitis in interleukin 10-deficient mice monoassociated with two different commensal bacteria. Gastroenterology, 2005. [DOI | PubMed]
- N Ibnou-Zekri, S Blum, EJ Schiffrin, T von der Weid. Divergent patterns of colonization and immune response elicited from two intestinal Lactobacillus strains that display similar properties in vitro. Infect Immun, 2003. [DOI | PubMed]
- M Schwarzer, D Srutkova, I Schabussova, T Hudcovic, J Akgün, U Wiedermann. Neonatal colonization of germ-free mice with Bifidobacterium longum prevents allergic sensitization to major birch pollen allergen Bet v 1. Vaccine, 2013. [DOI | PubMed]
- M Manohar, DO Baumann, NA Bos, JJ Cebra. Gut colonization of mice with actA-negative mutant of Listeria monocytogenes can stimulate a humoral mucosal immune response. Infect Immun, 2001. [DOI | PubMed]
- MV Herías, C Hessle, E Telemo, T Midtvedt, LA Hanson, AE Wold. Immunomodulatory effects of Lactobacillus plantarum colonizing the intestine of gnotobiotic rats. Clin Exp Immunol, 1999. [DOI | PubMed]
- M Waidmann, O Bechtold, JS Frick, HA Lehr, S Schubert, U Dobrindt. Bacteroides vulgatus protects against Escherichia coli-induced colitis in gnotobiotic interleukin-2-deficient mice. Gastroenterology, 2003. [DOI | PubMed]
- J Xia, S Jiang, L Lv, W Wu, Q Wang, Q Xu. Modulation of the immune response and metabolism in germ-free rats colonized by the probiotic Lactobacillus salivarius LI01. Appl Microbiol Biotechnol, 2021. [DOI | PubMed]
- M Schwarzer, A Repa, C Daniel, I Schabussova, T Hrncir, B Pot. Neonatal colonization of mice with Lactobacillus plantarum producing the aeroallergen Bet v 1 biases towards Th1 and T-regulatory responses upon systemic sensitization. Allergy, 2011. [DOI | PubMed]
- PA Ruiz, M Hoffmann, S Szcesny, M Blaut, D Haller. Innate mechanisms for Bifidobacterium lactis to activate transient pro-inflammatory host responses in intestinal epithelial cells after the colonization of germ-free rats. Immunology, 2005. [DOI | PubMed]
- MT Henke, EM Brown, CD Cassilly, H Vlamakis, RJ Xavier, J Clardy. Capsular polysaccharide correlates with immune response to the human gut microbe Ruminococcus gnavus. Proc Natl Acad Sci USA, 2021. [DOI | PubMed]
- EB Troy, DL Kasper. Beneficial effects of Bacteroides fragilis polysaccharides on the immune system. Front Biosci, 2010. [DOI]
- SK Mazmanian, CH Liu, AO Tzianabos, DL Kasper. An immunomodulatory molecule of symbiotic bacteria directs maturation of the host immune system. Cell, 2005. [DOI | PubMed]
- SS Seregin, N Golovchenko, B Schaf, J Chen, NA Pudlo, J Mitchell. NLRP6 protects Il10−/− mice from colitis by limiting colonization of Akkermansia muciniphila. Cell Rep, 2017. [DOI | PubMed]
- T Hudcovic, P Petr Hermanova, H Kozakova, O Benada, O Kofronova, M Schwarzer. Priority order of neonatal colonization by a probiotic or pathogenic Escherichia coli strain dictates the host response to experimental colitis. Front Microbiol, 2024. [DOI | PubMed]
- JL Pope, Y Yang, RC Newsome, W Sun, X Sun, M Ukhanova. Microbial colonization coordinates the pathogenesis of a Klebsiella pneumoniae infant isolate. Sci Rep, 2019. [DOI | PubMed]
- C Yang, I Mogno, EJ Contijoch, JN Borgerding, V Aggarwala, Z Li. Fecal IgA levels are determined by strain-level differences in Bacteroides ovatus and are modifiable by gut microbiota manipulation. Cell Host Microbe, 2020. [DOI | PubMed]
- JL Sonnenburg, CT Chen, JI Gordon. Genomic and metabolic studies of the impact of probiotics on a model gut symbiont and host. PLoS Biol, 2006. [DOI | PubMed]
- JB Lynch, EL Gonzalez, K Choy, KF Faull, T Jewell, A Arellano. Gut microbiota Turicibacter strains differentially modify bile acids and host lipids. Nat Commun, 2023. [DOI | PubMed]
- T Hirota, BH King. Autism spectrum disorder: a review. JAMA, 2023. [DOI | PubMed]
- NE Rosen, C Lord, FR Volkmar. The diagnosis of autism: from Kanner to DSM-III to DSM-5 and beyond. J Autism Dev Disord, 2021. [DOI | PubMed]
- V Nimmo-Smith, H Heuvelman, C Dalman, M Lundberg, S Idring, P Carpenter. Anxiety disorders in adults with autism spectrum disorder: a population-based study. J Autism Dev Disord, 2020. [DOI | PubMed]
- MJ Hollocks, JW Lerh, I Magiati, R Meiser-Stedman, TS Brugha. Anxiety and depression in adults with autism spectrum disorder: a systematic review and meta-analysis. Psychol Med, 2019. [DOI | PubMed]
- C Aymerich, M Pacho, A Catalan, N Yousaf, V Pérez-Rodríguez, MJ Hollocks. Prevalence and correlates of the concurrence of autism spectrum disorder and obsessive compulsive disorder in children and adolescents: a systematic review and meta-analysis. Brain Sci, 2024. [DOI | PubMed]
- Y Rong, C-J Yang, Y Jin, Y Wang. Prevalence of attention-deficit/hyperactivity disorder in individuals with autism spectrum disorder: a meta-analysis. Res Autism Spectr Disord, 2021. [DOI]
- G Vivanti, T Hamner, NR Lee. Neurodevelopmental disorders affecting sociability: recent research advances and future directions in autism spectrum disorder and Williams syndrome. Curr Neurol Neurosci Rep, 2018. [DOI | PubMed]
- L Balasco, G Provenzano, Y Bozzi. Sensory abnormalities in autism spectrum disorders: a focus on the tactile domain, from genetic mouse models to the clinic. Front Psych, 2020. [DOI]
- L Yi, Q Wang, C Song, ZR Han. Hypo-or hyperarousal? The mechanisms underlying social information processing in autism. Child Dev Perspect, 2022. [DOI]
- T Hirota, M Deserno, E McElroy. The network structure of irritability and aggression in individuals with autism spectrum disorder. J Autism Dev Disord, 2020. [DOI | PubMed]
- FA Satopoh. An overview of aggressive behaviors in children with autism spectrum disorder. Sci Psychiatr, 2024
- LY Hung, KG Margolis. Autism spectrum disorders and the gastrointestinal tract: insights into mechanisms and clinical relevance. Nat Rev Gastroenterol Hepatol, 2024. [DOI | PubMed]
- Z Dan, X Mao, Q Liu, M Guo, Y Zhuang, Z Liu. Altered gut microbial profile is associated with abnormal metabolism activity of Autism Spectrum Disorder. Gut Microbes, 2020. [DOI | PubMed]
- L Hetemi. Food intolerance and autism. Arch Med, 2020
- H Zhao, X Gao, L Xi, Y Shi, L Peng, C Wang. Mo1667 fecal microbiota transplantation for children with autism spectrum disorder. Gastrointest Endosc, 2019. [DOI]
- J Wang, B Ma, J Wang, Z Zhang, O Chen. Global prevalence of autism spectrum disorder and its gastrointestinal symptoms: a systematic review and meta-analysis. Front Psychiatry, 2022. [DOI | PubMed]
- NH Nogay, M Nahikian-Nelms. Can we reduce autism-related gastrointestinal and behavior problems by gut microbiota based dietary modulation? A review. Nutr Neurosci, 2021. [DOI | PubMed]
- M Kittana, A Ahmadani, KE Williams, A Attlee. Nutritional status and feeding behavior of children with autism spectrum disorder in the Middle East and North Africa Region: a systematic review. Nutrients, 2023. [DOI | PubMed]
- G Bjørklund, AV Skalny, MM Rahman, M Dadar, HA Yassa, J Aaseth. Toxic metal (loid)-based pollutants and their possible role in autism spectrum disorder. Environ Res, 2018. [DOI | PubMed]
- OS Von Ehrenstein, H Aralis, M Cockburn, B Ritz. In utero exposure to toxic air pollutants and risk of childhood autism. Epidemiology, 2014. [DOI | PubMed]
- IJ Malesza, M Malesza, J Walkowiak, N Mussin, D Walkowiak, R Aringazina. High-fat, western-style diet, systemic inflammation, and gut microbiota: a narrative review. Cells, 2021. [DOI | PubMed]
- M Kittana, A Ahmadani, F Al Marzooq, A Attlee. Dietary fat effect on the gut microbiome, and its role in the modulation of gastrointestinal disorders in children with autism spectrum disorder. Nutrients, 2021. [DOI | PubMed]
- KV Gantenbein, C Kanaka-Gantenbein. Mediterranean diet as an antioxidant: the impact on metabolic health and overall wellbeing. Nutrients, 2021. [DOI | PubMed]
- S Charisis, M Yannakoulia, N Scarmeas. Diets to promote healthy brain ageing. Nat Rev Neurol, 2025. [DOI | PubMed]
- E Schneider, R Balasubramanian, A Ferri, PD Cotter, G Clarke, JF Cryan. Fibre & fermented foods: differential effects on the microbiota–gut–brain axis. Proc Nutr Soc, 2024. [DOI | PubMed]
- FE Eroğlu, N Sanlier. Effect of fermented foods on some neurological diseases, microbiota, behaviors: mini review. Crit Rev Food Sci Nutr, 2023. [DOI | PubMed]
- MI Muñoz-García, MA Martínez-González, C Razquin, M Fernández-Matarrubia, F Guillén-Grima, E Toledo. Exploratory dietary patterns and cognitive function in the “Seguimiento Universidad de Navarra” (SUN) prospective cohort. Eur J Clin Nutr, 2022. [DOI | PubMed]
- SS Jeste, DH Geschwind. Disentangling the heterogeneity of autism spectrum disorder through genetic findings. Nat Rev Neurol, 2014. [DOI | PubMed]
- K Yenkoyan, A Grigoryan, K Fereshetyan, D Yepremyan. Advances in understanding the pathophysiology of autism spectrum disorders. Behav Brain Res, 2017. [DOI | PubMed]
- S Banerjee, M Riordan, MA Bhat. Genetic aspects of autism spectrum disorders: insights from animal models. Front Cell Neurosci, 2014. [DOI | PubMed]
- DB Budimirovic, A Schlageter, S Filipovic-Sadic, DD Protic, E Bram, EM Mahone. A genotype-phenotype study of high-resolution FMR1 nucleic acid and protein analyses in fragile X patients with neurobehavioral assessments. Brain Sci, 2020. [DOI | PubMed]
- M Mila, M Alvarez-Mora, I Madrigal, L Rodriguez-Revenga. Fragile X syndrome: an overview and update of the FMR1 gene. Clin Genet, 2018. [DOI | PubMed]
- M Palmieri, D Pozzer, N Landsberger. Advanced genetic therapies for the treatment of Rett syndrome: state of the art and future perspectives. Front Neurosci, 2023. [DOI | PubMed]
- U Petriti, DC Dudman, E Scosyrev, S Lopez-Leon. Global prevalence of Rett syndrome: systematic review and meta-analysis. Syst Rev, 2023. [DOI | PubMed]
- J Choi, S An, SY Lim. Current concepts of neurofibromatosis type 1: pathophysiology and treatment. Arch Craniofac Surg, 2022. [DOI | PubMed]
- JH Miles. Autism spectrum disorders—a genetics review. Genet Med, 2011. [DOI | PubMed]
- JN Crawley. Translational animal models of autism and neurodevelopmental disorders. Dialogues Clin Neurosci, 2012. [DOI | PubMed]
- RA Mitchell, M Mitchell, K Williams. The autism spectrum disorder phenotype in children with tuberous sclerosis complex: a systematic review and meta-analysis. Dev Med Child Neurol, 2022. [DOI | PubMed]
- S Assimopoulos, A Beauchamp, JP Lerch. Preclinical models of autism spectrum disorder. Neurodevelopmental pediatrics: genetic and environmental influences, 2023
- A Modabbernia, E Velthorst, A Reichenberg. Environmental risk factors for autism: an evidence-based review of systematic reviews and meta-analyses. Mol Autism, 2017. [DOI | PubMed]
- K Iwata, H Matsuzaki, N Takei, T Manabe, N Mori. Animal models of autism: an epigenetic and environmental viewpoint. J Cent Nerv Syst Dis, 2010. [DOI | PubMed]
- PH Patterson. Maternal infection and immune involvement in autism. Trends Mol Med, 2011. [DOI | PubMed]
- Z Ergaz, L Weinstein-Fudim, A Ornoy. Genetic and non-genetic animal models for autism spectrum disorders (ASD). Reprod Toxicol, 2016. [DOI | PubMed]
- L Desbonnet, G Clarke, F Shanahan, TG Dinan, J Cryan. Microbiota is essential for social development in the mouse. Mol Psychiatry, 2014. [DOI | PubMed]
- M Crumeyrolle-Arias, M Jaglin, A Bruneau, S Vancassel, A Cardona, V Daugé. Absence of the gut microbiota enhances anxiety-like behavior and neuroendocrine response to acute stress in rats. Psychoneuroendocrinology, 2014. [DOI | PubMed]
- L Xiao, J Yan, T Yang, J Zhu, T Li, H Wei. Fecal microbiome transplantation from children with autism spectrum disorder modulates tryptophan and serotonergic synapse metabolism and induces altered behaviors in germ-free mice. mSystems, 2021. [DOI | PubMed]
- S Hosie, M Ellis, M Swaminathan, F Ramalhosa, GO Seger, GK Balasuriya. Gastrointestinal dysfunction in patients and mice expressing the autism-associated R451C mutation in neuroligin-3. Autism Res, 2019. [DOI | PubMed]
- AK Sauer, J Bockmann, K Steinestel, TM Boeckers, AM Grabrucker. Altered intestinal morphology and microbiota composition in the autism spectrum disorders associated SHANK3 mouse model. Int J Mol Sci, 2019. [DOI | PubMed]
- EY Hsiao, SW McBride, S Hsien, G Sharon, ER Hyde, T McCue. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell, 2013. [DOI | PubMed]
- L Coretti, C Cristiano, E Florio, G Scala, A Lama, S Keller. Sex-related alterations of gut microbiota composition in the BTBR mouse model of autism spectrum disorder. Sci Rep, 2017. [DOI | PubMed]
- AV Golubeva, SA Joyce, G Moloney, A Burokas, E Sherwin, S Arboleya. Microbiota-related changes in bile acid & tryptophan metabolism are associated with gastrointestinal dysfunction in a mouse model of autism. EBioMedicine, 2017. [DOI | PubMed]
- B Dalile, L Van Oudenhove, B Vervliet, K Verbeke. The role of short-chain fatty acids in microbiota–gut–brain communication. Nat Rev Gastroenterol Hepatol, 2019. [DOI | PubMed]
- Y Dong, C Cui. The role of short-chain fatty acids in central nervous system diseases. Mol Cell Biochem, 2022. [DOI | PubMed]
- KJ O’Riordan, MK Collins, GM Moloney, EG Knox, MR Aburto, C Fülling. Short chain fatty acids: microbial metabolites for gut–brain axis signalling. Mol Cell Endocrinol, 2022. [DOI | PubMed]
- YP Silva, A Bernardi, RL Frozza. The role of short-chain fatty acids from gut microbiota in gut–brain communication. Front Endocrinol, 2020. [DOI]
- AM Tartaglione, V Pazienza, G Calamandrei, L Ricceri. A snapshot of gut microbiota data from murine models of autism spectrum disorder: still a blurred picture. Neurosci Biobehav Rev, 2023. [DOI | PubMed]
- M Sgritta, SW Dooling, SA Buffington, EN Momin, MB Francis, RA Britton. Mechanisms underlying microbial-mediated changes in social behavior in mouse models of autism spectrum disorder. Neuron, 2019. [DOI | PubMed]
- L Tabouy, D Getselter, O Ziv, M Karpuj, T Tabouy, I Lukic. Dysbiosis of microbiome and probiotic treatment in a genetic model of autism spectrum disorders. Brain Behav Immun, 2018. [DOI | PubMed]
- K Neier, TE Grant, RL Palmer, D Chappell, SM Hakam, KM Yasui. Sex disparate gut microbiome and metabolome perturbations precede disease progression in a mouse model of Rett syndrome. Commun Biol, 2021. [DOI | PubMed]
- CG De Theije, H Wopereis, M Ramadan, T van Eijndthoven, J Lambert, J Knol. Altered gut microbiota and activity in a murine model of autism spectrum disorders. Brain Behav Immun, 2014. [DOI | PubMed]
- F Liu, K Horton-Sparks, V Hull, RW Li, V Martínez-Cerdeño. The valproic acid rat model of autism presents with gut bacterial dysbiosis similar to that in human autism. Mol Autism, 2018. [DOI | PubMed]
- AM Tartaglione, A Villani, MA Ajmone-Cat, L Minghetti, L Ricceri, V Pazienza. Maternal immune activation induces autism-like changes in behavior, neuroinflammatory profile and gut microbiota in mouse offspring of both sexes. Transl Psychiatry, 2022. [DOI | PubMed]
- S Liu, E Li, Z Sun, D Fu, G Duan, M Jiang. Altered gut microbiota and short chain fatty acids in Chinese children with autism spectrum disorder. Sci Rep, 2019. [DOI | PubMed]
- RM Stilling, M Van De Wouw, G Clarke, C Stanton, TG Dinan, JF Cryan. The neuropharmacology of butyrate: the bread and butter of the microbiota–gut–brain axis?. Neurochem Int, 2016. [DOI | PubMed]
- A Gallucci, K Patterson, A Weit, W Van Der Pol, L Dubois, A Percy. Microbial community changes in a female rat model of Rett syndrome. Prog Neuropsychopharmacol Biol Psychiatry, 2021. [DOI | PubMed]
- N Kratsman, D Getselter, E Elliott. Sodium butyrate attenuates social behavior deficits and modifies the transcription of inhibitory/excitatory genes in the frontal cortex of an autism model. Neuropharmacology, 2016. [DOI | PubMed]
- K Takuma, Y Hara, S Kataoka, T Kawanai, Y Maeda, R Watanabe. Chronic treatment with valproic acid or sodium butyrate attenuates novel object recognition deficits and hippocampal dendritic spine loss in a mouse model of autism. Pharmacol Biochem Behav, 2014. [DOI | PubMed]
- D Zhou, Q Pan, F-Z Xin, R-N Zhang, C-X He, G-Y Chen. Sodium butyrate attenuates high-fat diet-induced steatohepatitis in mice by improving gut microbiota and gastrointestinal barrier. World J Gastroenterol, 2017. [DOI | PubMed]
- G Chen, X Ran, B Li, Y Li, D He, B Huang. Sodium butyrate inhibits inflammation and maintains epithelium barrier integrity in a TNBS-induced inflammatory bowel disease mice model. EBioMedicine, 2018. [DOI | PubMed]
- W Yong, Y Zhao, X Jiang, P Li. Sodium butyrate alleviates pre-eclampsia in pregnant rats by improving the gut microbiota and short-chain fatty acid metabolites production. J Appl Microbiol, 2022. [DOI | PubMed]
- SA Buffington, GV Di Prisco, TA Auchtung, NJ Ajami, JF Petrosino, M Costa-Mattioli. Microbial reconstitution reverses maternal diet-induced social and synaptic deficits in offspring. Cell, 2016. [DOI | PubMed]
- S Kim, H Kim, YS Yim, S Ha, K Atarashi, TG Tan. Maternal gut bacteria promote neurodevelopmental abnormalities in mouse offspring. Nature, 2017. [DOI | PubMed]
- JM Saunders, JL Moreno, D Ibi, M Sikaroodi, DJ Kang, R Muñoz-Moreno. Gut microbiota manipulation during the prepubertal period shapes behavioral abnormalities in a mouse neurodevelopmental disorder model. Sci Rep, 2020. [DOI | PubMed]
- DE Septyaningtrias, C-W Lin, R Ouchida, N Nakai, W Suda, M Hattori. Altered microbiota composition reflects enhanced communication in 15q11-13 CNV mice. Neurosci Res, 2020. [DOI | PubMed]
- CR Lammert, EL Frost, AC Bolte, MJ Paysour, ME Shaw, CE Bellinger. Cutting edge: critical roles for microbiota-mediated regulation of the immune system in a prenatal immune activation model of autism. J Immunol, 2018. [DOI | PubMed]
- AC Ericsson, CL Franklin. Manipulating the gut microbiota: methods and challenges. ILAR J, 2015. [DOI | PubMed]
- J Wasilewska, M Klukowski. Gastrointestinal symptoms and autism spectrum disorder: links and risks—a possible new overlap syndrome. Pediatr Health Med Therap, 2015. [DOI]
- M Fiorentino, A Sapone, S Senger, SS Camhi, SM Kadzielski, TM Buie. Blood–brain barrier and intestinal epithelial barrier alterations in autism spectrum disorders. Mol Autism, 2016. [DOI | PubMed]
- L de Magistris, V Familiari, A Pascotto, A Sapone, A Frolli, P Iardino. Alterations of the intestinal barrier in patients with autism spectrum disorders and in their first-degree relatives. J Pediatr Gastroenterol Nutr, 2010. [DOI | PubMed]
- Elevated plasma cytokines in autism spectrum disorders provide evidence of immune dysfunction and are associated with impaired behavioral outcome. Brain Behav Immun, 2011. [DOI | PubMed]
- F Li, H Ke, S Wang, W Mao, C Fu, X Chen. Leaky gut plays a critical role in the pathophysiology of autism in mice by activating the lipopolysaccharide-mediated toll-like receptor 4-myeloid differentiation factor 88-nuclear factor kappa B signaling pathway. Neurosci Bull, 2023. [DOI | PubMed]
- JM Parrott, T Oster, HY Lee. Altered inflammatory response in FMRP-deficient microglia. iScience, 2021. [DOI | PubMed]
- F Laudisi, C Stolfi, G Bevivino, C Maresca, E Franzè, E Troncone. GATA6 deficiency leads to epithelial barrier dysfunction and enhances susceptibility to gut inflammation. J Crohns Colitis, 2021. [DOI]
- SM Hermes, MC Andresen, SA Aicher. Localization of TRPV1 and P2X3 in unmyelinated and myelinated vagal afferents in the rat. J Chem Neuroanat, 2016. [DOI | PubMed]
- K Matsumoto, T Hosoya, K Tashima, T Namiki, T Murayama, S Horie. Distribution of transient receptor potential vanilloid 1 channel-expressing nerve fibers in mouse rectal and colonic enteric nervous system: relationship to peptidergic and nitrergic neurons. Neuroscience, 2011. [DOI | PubMed]
- LM Lauffer, R Iakoubov, PL Brubaker. GPR119 is essential for oleoylethanolamide-induced glucagon-like peptide-1 secretion from the intestinal enteroendocrine L-cell. Diabetes, 2009. [DOI | PubMed]
- R Yang, P Wang, Z Chen, W Hu, Y Gong, W Zhang. WY-14643, a selective agonist of peroxisome proliferator-activated receptor-α, ameliorates lipopolysaccharide-induced depressive-like behaviors by preventing neuroinflammation and oxido-nitrosative stress in mice. Pharmacol Biochem Behav, 2017. [DOI | PubMed]
- H Wei, KK Chadman, DP McCloskey, AM Sheikh, M Malik, WT Brown. Brain IL-6 elevation causes neuronal circuitry imbalances and mediates autism-like behaviors. Biochim Biophys Acta, 2012. [DOI | PubMed]
- G Sharon, NJ Cruz, D-W Kang, MJ Gandal, B Wang, Y-M Kim. Human gut microbiota from autism spectrum disorder promote behavioral symptoms in mice. Cell, 2019. [DOI | PubMed]
- LJ Cohen, D Esterhazy, S-H Kim, C Lemetre, RR Aguilar, EA Gordon. Commensal bacteria make GPCR ligands that mimic human signalling molecules. Nature, 2017. [DOI | PubMed]
- M Yano Jessica, K Yu, P Donaldson Gregory, G Shastri Gauri, P Ann, L Ma. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell, 2015. [DOI | PubMed]
- AR Best, WG Regehr. Serotonin evokes endocannabinoid release and retrogradely suppresses excitatory synapses. J Neurosci, 2008. [DOI | PubMed]
- A Glinert, V Zlidennyy, S Turjeman, E Sharon, R Schweitzer, S Khatib. What’s GABA got to do with it? A potential link between the microbiome, schizophrenia, and the endo-cannabinoid system. Psychiatry Res, 2024. [DOI | PubMed]
- F Guida, S Boccella, C Belardo, M Iannotta, F Piscitelli, F De Filippis. Altered gut microbiota and endocannabinoid system tone in vitamin D deficiency-mediated chronic pain. Brain Behav Immun, 2020. [DOI | PubMed]
- F Guida, F Turco, M Iannotta, D De Gregorio, I Palumbo, G Sarnelli. Antibiotic-induced microbiota perturbation causes gut endocannabinoidome changes, hippocampal neuroglial reorganization and depression in mice. Brain Behav Immun, 2018. [DOI | PubMed]
- G Gioacchini, G Rossi, O Carnevali. Host–probiotic interaction: new insight into the role of the endocannabinoid system by in vivo and ex vivo approaches. Sci Rep, 2017. [DOI | PubMed]
- E Grossi, S Melli, D Dunca, V Terruzzi. Unexpected improvement in core autism spectrum disorder symptoms after long-term treatment with probiotics. SAGE Open Med Case Rep, 2016. [DOI | PubMed]
- M De Angelis, M Piccolo, L Vannini, S Siragusa, A De Giacomo, DI Serrazzanetti. Fecal microbiota and metabolome of children with autism and pervasive developmental disorder not otherwise specified. PLoS ONE, 2013. [DOI | PubMed]
- F Strati, D Cavalieri, D Albanese, C De Felice, C Donati, J Hayek. New evidences on the altered gut microbiota in autism spectrum disorders. Microbiome, 2017. [DOI | PubMed]
- G Patki, N Solanki, F Atrooz, F Allam, S Salim. Depression, anxiety-like behavior and memory impairment are associated with increased oxidative stress and inflammation in a rat model of social stress. Brain Res, 2013. [DOI | PubMed]
- V Micale, V Di Marzo, A Sulcova, CT Wotjak, F Drago. Endocannabinoid system and mood disorders: priming a target for new therapies. Pharmacol Ther, 2013. [DOI | PubMed]
- G Provensi, R Coccurello, H Umehara, L Munari, G Giacovazzo, N Galeotti. Satiety factor oleoylethanolamide recruits the brain histaminergic system to inhibit food intake. Proc Natl Acad Sci USA, 2014. [DOI | PubMed]
- A Romano, T Cassano, B Tempesta, S Cianci, P Dipasquale, R Coccurello. The satiety signal oleoylethanolamide stimulates oxytocin neurosecretion from rat hypothalamic neurons. Peptides, 2013. [DOI | PubMed]
- D Wei, D Lee, CD Cox, CA Karsten, O Peñagarikano, DH Geschwind. Endocannabinoid signaling mediates oxytocin-driven social reward. Proc Natl Acad Sci USA, 2015. [DOI | PubMed]
- MC Davis, J Lee, WP Horan, AD Clarke, MR McGee, MF Green. Effects of single dose intranasal oxytocin on social cognition in schizophrenia. Schizophr Res, 2013. [DOI | PubMed]
- J Woolley, B Chuang, O Lam, W Lai, A O’Donovan, K Rankin. Oxytocin administration enhances controlled social cognition in patients with schizophrenia. Psychoneuroendocrinology, 2014. [DOI | PubMed]
- Y Geng, W Zhao, F Zhou, X Ma, S Yao, R Hurlemann. Oxytocin enhancement of emotional empathy: generalization across cultures and effects on amygdala activity. Front Neurosci, 2018. [DOI | PubMed]
- S Shahrestani, AH Kemp, AJ Guastella. The impact of a single administration of intranasal oxytocin on the recognition of basic emotions in humans: a meta-analysis. Neuropsychopharmacology, 2013. [DOI | PubMed]
- RH Ring, JE Malberg, L Potestio, J Ping, S Boikess, B Luo. Anxiolytic-like activity of oxytocin in male mice: behavioral and autonomic evidence, therapeutic implications. Psychopharmacology, 2006. [DOI | PubMed]
- MM McCarthy, CH McDonald, PJ Brooks, D Goldman. An anxiolytic action of oxytocin is enhanced by estrogen in the mouse. Physiol Behav, 1996. [DOI | PubMed]
- K Uvnäs-Moberg, S Ahlenius, V Hillegaart, P Alster. High doses of oxytocin cause sedation and low doses cause an anxiolytic-like effect in male rats. Pharmacol Biochem Behav, 1994. [DOI | PubMed]
- M Yoshida, Y Takayanagi, K Inoue, T Kimura, LJ Young, T Onaka. Evidence that oxytocin exerts anxiolytic effects via oxytocin receptor expressed in serotonergic neurons in mice. J Neurosci, 2009. [DOI | PubMed]
- S Sabihi, NE Durosko, SM Dong, B Leuner. Oxytocin in the prelimbic medial prefrontal cortex reduces anxiety-like behavior in female and male rats. Psychoneuroendocrinology, 2014. [DOI | PubMed]
- JM Gray, HA Vecchiarelli, M Morena, TT Lee, DJ Hermanson, AB Kim. Corticotropin-releasing hormone drives anandamide hydrolysis in the amygdala to promote anxiety. J Neurosci, 2015. [DOI | PubMed]
- I Dincheva, AT Drysdale, CA Hartley, DC Johnson, D Jing, EC King. FAAH genetic variation enhances fronto-amygdala function in mouse and human. Nat Commun, 2015. [DOI | PubMed]
- P Adamczyk, A Golda, A McCreary, M Filip, E Przegaliriski. Activation of endocannabinoid transmission induces antidepressant-like effects in rats. Acta Physiol Pol, 2008
- C-H Chang, Y-C Liu, C-Y Sun, C-L Su, P-W Gean. Regulation of stress-provoked aggressive behavior using endocannabinoids. Neurobiol Stress, 2021. [DOI | PubMed]
- M Rodriguez-Arias, F Navarrete, M Daza-Losada, D Navarro, MA Aguilar, P Berbel. CB1 cannabinoid receptor-mediated aggressive behavior. Neuropharmacology, 2013. [DOI | PubMed]
- M Migliaro, AE Ruiz-Contreras, A Herrera-Solís, M Méndez-Díaz, OE Prospéro-García. Endocannabinoid system and aggression across animal species. Neurosci Biobehav Rev, 2023. [DOI | PubMed]
- J Haller, F Matyas, K Soproni, B Varga, B Barsy, B Nemeth. Correlated species differences in the effects of cannabinoid ligands on anxiety and on GABAergic and glutamatergic synaptic transmission. Eur J Neurosci, 2007. [DOI | PubMed]
- M Huang, K Liu, Z Wei, Z Feng, J Chen, J Yang. Serum oxytocin level correlates with gut microbiome dysbiosis in children with autism spectrum disorder. Front Neurosci, 2021. [DOI | PubMed]
- SM Finegold, SE Dowd, V Gontcharova, C Liu, KE Henley, RD Wolcott. Pyrosequencing study of fecal microflora of autistic and control children. Anaerobe, 2010. [DOI | PubMed]
- M De Angelis, R Francavilla, M Piccolo, A De Giacomo, M Gobbetti. Autism spectrum disorders and intestinal microbiota. Gut Microbes, 2015. [DOI | PubMed]
- DF MacFabe. Short-chain fatty acid fermentation products of the gut microbiome: implications in autism spectrum disorders. Microb Ecol Health Dis, 2012
- HS Al-Salem, RS Bhat, L Al-Ayadhi, A El-Ansary. Therapeutic potency of bee pollen against biochemical autistic features induced through acute and sub-acute neurotoxicity of orally administered propionic acid. BMC Complement Altern Med, 2016. [DOI | PubMed]
- P Witters, E Debbold, K Crivelly, KV Kerckhove, K Corthouts, B Debbold. Autism in patients with propionic acidemia. Mol Genet Metab, 2016. [DOI | PubMed]
- R Mitsui, S Ono, S Karaki, A Kuwahara. Neural and non-neural mediation of propionate-induced contractile responses in the rat distal colon. Neurogastroenterol Motil, 2005. [DOI | PubMed]
- C Hao, Z Gao, X Liu, Z Rong, J Jia, K Kang. Intravenous administration of sodium propionate induces antidepressant or prodepressant effect in a dose dependent manner. Sci Rep, 2020. [DOI | PubMed]
- Y-F Hou, C Shan, S-Y Zhuang, Q-Q Zhuang, A Ghosh, K-C Zhu. Gut microbiota-derived propionate mediates the neuroprotective effect of osteocalcin in a mouse model of Parkinson’s disease. Microbiome, 2021. [DOI | PubMed]
- C Morland, A-S Frøland, MN Pettersen, J Storm-Mathisen, V Gundersen, F Rise. Propionate enters GABAergic neurons, inhibits GABA transaminase, causes GABA accumulation and lethargy in a model of propionic acidemia. Biochemical Journal, 2018. [DOI | PubMed]
- P Mehrpouya-Bahrami, KN Chitrala, MS Ganewatta, C Tang, EA Murphy, RT Enos. Blockade of CB1 cannabinoid receptor alters gut microbiota and attenuates inflammation and diet-induced obesity. Sci Rep, 2017. [DOI | PubMed]
- DA Youssef, HM El-Fayoumi, MF Mahmoud. Beta-caryophyllene alleviates diet-induced neurobehavioral changes in rats: the role of CB2 and PPAR-γ receptors. Biomed Pharmacother, 2019. [DOI | PubMed]
- M Qin, Z Zeidler, K Moulton, L Krych, Z Xia, CB Smith. Endocannabinoid-mediated improvement on a test of aversive memory in a mouse model of fragile X syndrome. Behav Brain Res, 2015. [DOI | PubMed]
- SN Umathe, SS Manna, NS Jain. Involvement of endocannabinoids in antidepressant and anti-compulsive effect of fluoxetine in mice. Behav Brain Res, 2011. [DOI | PubMed]
- G Gobbi, F Bambico, R Mangieri, M Bortolato, P Campolongo, M Solinas. Antidepressant-like activity and modulation of brain monoaminergic transmission by blockade of anandamide hydrolysis. Proc Natl Acad Sci USA, 2005. [DOI | PubMed]
- KY Vinod, S Xie, D Psychoyos, BL Hungund, TB Cooper, SM Tejani-Butt. Dysfunction in fatty acid amide hydrolase is associated with depressive-like behavior in Wistar Kyoto rats. PLoS ONE, 2012. [DOI | PubMed]
- DM Kerr, A Gilmartin, M Roche. Pharmacological inhibition of fatty acid amide hydrolase attenuates social behavioural deficits in male rats prenatally exposed to valproic acid. Pharmacol Res, 2016. [DOI | PubMed]
- HF Wu, TY Lu, MC Chu, PS Chen, CW Lee, HC Lin. Targeting the inhibition of fatty acid amide hydrolase ameliorate the endocannabinoid-mediated synaptic dysfunction in a valproic acid-induced rat model of Autism. Neuropharmacology, 2020. [DOI | PubMed]
- V Micale, L Cristino, A Tamburella, S Petrosino, GM Leggio, F Drago. Anxiolytic effects in mice of a dual blocker of fatty acid amide hydrolase and transient receptor potential vanilloid type-1 channels. Neuropsychopharmacology, 2009. [DOI | PubMed]
- MN Hill, BB Gorzalka. Pharmacological enhancement of cannabinoid CB1 receptor activity elicits an antidepressant-like response in the rat forced swim test. Eur Neuropsychopharmacol, 2005. [DOI | PubMed]
- M Rutkowska, O Jachimczuk. Antidepressant-like properties of ACEA (arachidonyl-2-chloroethylamide), the selective agonist of CB1 receptors. Acta Pol Pharm, 2004. [PubMed]
- FR Bambico, N Katz, G Debonnel, G Gobbi. Cannabinoids elicit antidepressant-like behavior and activate serotonergic neurons through the medial prefrontal cortex. J Neurosci, 2007. [DOI | PubMed]
- FR Bambico, N-T Nguyen, N Katz, G Gobbi. Chronic exposure to cannabinoids during adolescence but not during adulthood impairs emotional behaviour and monoaminergic neurotransmission. Neurobiol Dis, 2010. [DOI | PubMed]
- DS Karhson, KM Krasinska, JA Dallaire, RA Libove, JM Phillips, AS Chien. Plasma anandamide concentrations are lower in children with autism spectrum disorder. Mol Autism, 2018. [DOI | PubMed]
- D Siniscalco, A Sapone, C Giordano, A Cirillo, L de Magistris, F Rossi. Cannabinoid receptor type 2, but not type 1, is up-regulated in peripheral blood mononuclear cells of children affected by autistic disorders. J Autism Dev Disord, 2013. [DOI | PubMed]
- K-M Jung, M Sepers, CM Henstridge, O Lassalle, D Neuhofer, H Martin. Uncoupling of the endocannabinoid signalling complex in a mouse model of fragile X syndrome. Nat Commun, 2012. [DOI | PubMed]
- PS Pirbhoy, CR Jonak, R Syed, DA Argueta, PA Perez, MB Wiley. Increased 2-arachidonoyl-sn-glycerol levels normalize cortical responses to sound and improve behaviors in Fmr1 KO mice. J Neurodev Disord, 2021. [DOI | PubMed]
- M Maccarrone, S Rossi, M Bari, V De Chiara, C Rapino, A Musella. Abnormal mGlu 5 receptor/endocannabinoid coupling in mice lacking FMRP and BC1 RNA. Neuropsychopharmacology, 2010. [DOI | PubMed]
- K Lee, Y Vyas, CC Garner, JM Montgomery. Autism-associated Shank3 mutations alter mGluR expression and mGluR-dependent but not NMDA receptor-dependent long-term depression. Synapse, 2019. [DOI | PubMed]
- B Bertolino, R Crupi, D Impellizzeri, G Bruschetta, M Cordaro, R Siracusa. Beneficial effects of co-ultramicronized palmitoylethanolamide/luteolin in a mouse model of autism and in a case report of autism. CNS Neurosci Ther, 2017. [DOI | PubMed]
- N Antonucci, A Cirillo, D Siniscalco. Beneficial effects of palmitoylethanolamide on expressive language, cognition, and behaviors in autism: a report of two cases. Case Rep Psychiatry, 2015. [PubMed]
- F Borrelli, B Romano, S Petrosino, E Pagano, R Capasso, D Coppola. Palmitoylethanolamide, a naturally occurring lipid, is an orally effective intestinal anti-inflammatory agent. Br J Pharmacol, 2015. [DOI | PubMed]
- E Domi, S Uhrig, L Soverchia, R Spanagel, AC Hansson, E Barbier. Genetic deletion of neuronal PPARγ enhances the emotional response to acute stress and exacerbates anxiety: an effect reversed by rescue of amygdala PPARγ function. J Neurosci, 2016. [DOI | PubMed]
- S Chikahisa, D Chida, T Shiuchi, S Harada, N Shimizu, A Otsuka. Enhancement of fear learning in PPARα knockout mice. Behav Brain Res, 2019. [DOI | PubMed]
- B Hempel, M Crissman, S Pari, B Klein, G-H Bi, H Alton. PPARα and PPARγ are expressed in midbrain dopamine neurons and modulate dopamine- and cannabinoid-mediated behavior in mice. Mol Psychiatry, 2023. [DOI | PubMed]
- Q-X Shi, L-K Yang, W-L Shi, L Wang, S-M Zhou, S-Y Guan. The novel cannabinoid receptor GPR55 mediates anxiolytic-like effects in the medial orbital cortex of mice with acute stress. Mol Brain, 2017. [DOI | PubMed]
- R Mirza, B Sharma. Benefits of fenofibrate in prenatal valproic acid-induced autism spectrum disorder related phenotype in rats. Brain Res Bull, 2019. [DOI | PubMed]
- R Mirza, B Sharma. Selective modulator of peroxisome proliferator-activated receptor-α protects propionic acid induced autism-like phenotypes in rats. Life Sci, 2018. [DOI | PubMed]
- MA Karwad, T Macpherson, B Wang, E Theophilidou, S Sarmad, DA Barrett. Oleoylethanolamine and palmitoylethanolamine modulate intestinal permeability in vitro via TRPV1 and PPARα. FASEB J, 2017. [DOI | PubMed]
- M Di Paola, E Bonechi, G Provensi, A Costa, G Clarke, C Ballerini. Oleoylethanolamide treatment affects gut microbiota composition and the expression of intestinal cytokines in Peyer’s patches of mice. Sci Rep, 2018. [DOI | PubMed]
- Y Wang, C Tang, Y Tang, H Yin, X Liu. Capsaicin has an anti-obesity effect through alterations in gut microbiota populations and short-chain fatty acid concentrations. Food Nutr Res, 2020. [DOI | PubMed]
- M Derrien, C Belzer, WM de Vos. Akkermansia muciniphila and its role in regulating host functions. Microb Pathog, 2017. [DOI | PubMed]
- Y Yang, Z Zhong, B Wang, X Xia, W Yao, L Huang. Early-life high-fat diet-induced obesity programs hippocampal development and cognitive functions via regulation of gut commensal Akkermansia muciniphila. Neuropsychopharmacology, 2019. [DOI | PubMed]
- A Everard, C Belzer, L Geurts, JP Ouwerkerk, C Druart, LB Bindels. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc Natl Acad Sci USA, 2013. [DOI | PubMed]
- C Depommier, RM Vitale, FA Iannotti, C Silvestri, N Flamand, C Druart. Beneficial effects of Akkermansia muciniphila are not associated with major changes in the circulating endocannabinoidome but linked to higher mono-palmitoyl-glycerol levels as new PPARα agonists. Cells, 2021. [DOI | PubMed]
- N Goo, HJ Bae, K Park, J Kim, Y Jeong, M Cai. The effect of fecal microbiota transplantation on autistic-like behaviors in Fmr1 KO mice. Life Sci, 2020. [DOI | PubMed]
- Y Xu, N Wang, H-Y Tan, S Li, C Zhang, Y Feng. Function of Akkermansia muciniphila in obesity: interactions with lipid metabolism, immune response and gut systems. Front Microbiol, 2020. [DOI | PubMed]
- IY Hwang, HR Kim, R De Sotto, MW Chang. Engineered probiotics modulate the endocannabinoid system. Biotechnol Notes, 2021. [DOI]
- J Cheng, S Venkatesh, K Ke, MJ Barratt, JI Gordon. A human gut Faecalibacterium prausnitzii fatty acid amide hydrolase. Science, 2024. [DOI | PubMed]
- L Coretti, L Paparo, MP Riccio, F Amato, M Cuomo, A Natale. Gut microbiota features in young children with autism spectrum disorders. Front Microbiol, 2018. [DOI | PubMed]
- X Ding, Y Xu, X Zhang, L Zhang, G Duan, C Song. Gut microbiota changes in patients with autism spectrum disorders. J Psychiatr Res, 2020. [DOI | PubMed]
- L Wang, CT Christophersen, MJ Sorich, JP Gerber, MT Angley, MA Conlon. Elevated fecal short chain fatty acid and ammonia concentrations in children with autism spectrum disorder. Dig Dis Sci, 2012. [DOI | PubMed]
- H Yang, C Chen. Cyclooxygenase-2 in synaptic signaling. Curr Pharm Des, 2008. [DOI | PubMed]
- CT Wong, I Bestard-Lorigados, DA Crawford. Autism-related behaviors in the cyclooxygenase-2-deficient mouse model. Genes Brain Behav, 2019. [DOI | PubMed]
- R Rai-Bhogal, E Ahmad, H Li, DA Crawford. Microarray analysis of gene expression in the cyclooxygenase knockout mice—a connection to autism spectrum disorder. Eur J Neurosci, 2018. [DOI | PubMed]
- SA Brigandi, H Shao, SY Qian, Y Shen, B-L Wu, JX Kang. Autistic children exhibit decreased levels of essential fatty acids in red blood cells. Int J Mol Sci, 2015. [DOI | PubMed]
- A El-Ansary, WM Hassan, H Qasem, UN Das. Identification of biomarkers of impaired sensory profiles among autistic patients. PLoS ONE, 2016. [DOI | PubMed]
- HJ Yoo, IH Cho, M Park, E Cho, SC Cho, BN Kim. Association between PTGS2 polymorphism and autism spectrum disorders in Korean trios. Neurosci Res, 2008. [DOI | PubMed]
- JM Bandim, LO Ventura, MT Miller, HC Almeida, AE Costa. Autism and Möbius sequence: an exploratory study of children in northeastern Brazil. Arq Neuropsiquiatr, 2003. [DOI | PubMed]
- C Vauzelle, D Beghin, M-P Cournot, E Elefant. Birth defects after exposure to misoprostol in the first trimester of pregnancy: prospective follow-up study. Reprod Toxicol, 2013. [DOI | PubMed]
- S Vendramini-Pittoli, ML Guion-Almeida, A Richieri-Costa, JM Santos, NM Kokitsu-Nakata. Clinical findings in children with congenital anomalies and misoprostol intrauterine exposure: a study of 38 cases. J Pediatr Genet, 2013. [PubMed]
- SC Jones, CS Gordon, M Akram, N Murphy, F Sharkie. Inclusion, exclusion and isolation of autistic people: community attitudes and autistic people’s experiences. J Autism Dev Disord, 2022. [DOI | PubMed]
- V Khullar, HP Singh, M Bala. Meltdown/tantrum detection system for individuals with autism spectrum disorder. Appl Artif Intell, 2021. [DOI]
- M Aliczki, ZK Varga, Z Balogh, J Haller. Involvement of 2-arachidonoylglycerol signaling in social challenge responding of male CD1 mice. Psychopharmacology, 2015. [DOI | PubMed]
- A Hartmann, SF Lisboa, AB Sonego, D Coutinho, FV Gomes, FS Guimarães. Cannabidiol attenuates aggressive behavior induced by social isolation in mice: involvement of 5-HT1A and CB1 receptors. Prog Neuropsychopharmacol Biol Psychiatry, 2019. [DOI | PubMed]
- M Rodríguez-Arias, F Navarrete, MC Blanco-Gandia, MC Arenas, MA Aguilar, A Bartoll-Andrés. Role of CB2 receptors in social and aggressive behavior in male mice. Psychopharmacology, 2015. [DOI | PubMed]
- J Fontenot, EC Loetz, M Ishiki, ST Bland. Monoacylglycerol lipase inhibition alters social behavior in male and female rats after post-weaning social isolation. Behav Brain Res, 2018. [DOI | PubMed]
- M Aguilera, P Vergara, V Martínez. Stress and antibiotics alter luminal and wall-adhered microbiota and enhance the local expression of visceral sensory-related systems in mice. Neurogastroenterol Motil, 2013. [DOI | PubMed]
- R Inoue, Y Sakaue, Y Kawada, R Tamaki, Z Yasukawa, M Ozeki. Dietary supplementation with partially hydrolyzed guar gum helps improve constipation and gut dysbiosis symptoms and behavioral irritability in children with autism spectrum disorder. J Clin Biochem Nutr, 2019. [DOI | PubMed]
- DR Rose, H Yang, G Serena, C Sturgeon, B Ma, M Careaga. Differential immune responses and microbiota profiles in children with autism spectrum disorders and co-morbid gastrointestinal symptoms. Brain Behav Immun, 2018. [DOI | PubMed]
- E Santocchi, L Guiducci, F Fulceri, L Billeci, E Buzzigoli, F Apicella. Gut to brain interaction in autism spectrum disorders: a randomized controlled trial on the role of probiotics on clinical, biochemical and neurophysiological parameters. BMC Psychiatry, 2016. [DOI | PubMed]
- JF Cryan, TG Dinan. Mind-altering microorganisms: the impact of the gut microbiota on brain and behaviour. Nat Rev Neurosci, 2012. [DOI | PubMed]
- JF Cryan, KJ O’Riordan, K Sandhu, V Peterson, TG Dinan. The gut microbiome in neurological disorders. Lancet Neurol, 2020. [DOI | PubMed]
- NR Mota, G Poelmans, M Klein, B Torrico, N Fernàndez-Castillo, B Cormand. Cross-disorder genetic analyses implicate dopaminergic signaling as a biological link between attention-deficit/hyperactivity disorder and obesity measures. Neuropsychopharmacology, 2020. [DOI | PubMed]
- D Centonze, M Bari, B Di Michele, S Rossi, V Gasperi, A Pasini. Altered anandamide degradation in attention-deficit/hyperactivity disorder. Neurology, 2009. [DOI | PubMed]
- M Castelli, M Federici, S Rossi, V De Chiara, F Napolitano, V Studer. Loss of striatal cannabinoid CB1 receptor function in attention-deficit / hyperactivity disorder mice with point-mutation of the dopamine transporter. Eur J Neurosci, 2011. [DOI | PubMed]
- A Ahmadalipour, L Mehdizadeh Fanid, N Zeinalzadeh, M Alizadeh, H Vaezi, Z Hassanpour Aydinlou. The first evidence of an association between a polymorphism in the endocannabinoid-degrading enzyme FAAH (FAAH rs2295633) with attention deficit hyperactivity disorder. Genomics, 2020. [DOI | PubMed]
- Y Ito, M Tomizawa, K Suzuki, Y Shirakawa, H Ono, K Adachi. Organophosphate agent induces ADHD-like behaviors via inhibition of brain endocannabinoid-hydrolyzing enzyme(s) in adolescent male rats. J Agric Food Chem, 2020. [DOI | PubMed]
- T Okumura, T Nozu, M Ishioh, S Igarashi, S Kumei, M Ohhira. Centrally administered butyrate improves gut barrier function, visceral sensation and septic lethality in rats. J Pharmacol Sci, 2021. [DOI | PubMed]
- D Gkougka, K Mitropoulos, G Tzanakaki, E Panagouli, T Psaltopoulou, L Thomaidis. Gut microbiome and attention deficit/hyperactivity disorder: a systematic review. Pediatr Res, 2022. [DOI | PubMed]
- J Szopinska-Tokov, S Dam, J Naaijen, P Konstanti, N Rommelse, C Belzer. Investigating the gut microbiota composition of individuals with attention-deficit/hyperactivity disorder and association with symptoms. Microorganisms, 2020. [DOI | PubMed]
- L Wang, Z Xie, G Li, G Li, J Liang. Two-sample Mendelian randomization analysis investigates causal associations between gut microbiota and attention deficit hyperactivity disorder. Front Microbiol, 2023. [DOI | PubMed]
- A Checa-Ros, A Jeréz-Calero, A Molina-Carballo, C Campoy, A Muñoz-Hoyos. Current evidence on the role of the gut microbiome in ADHD pathophysiology and therapeutic implications. Nutrients, 2021. [DOI | PubMed]
- C D’Addario, F Bellia, B Benatti, B Grancini, M Vismara, M Pucci. Exploring the role of BDNF DNA methylation and hydroxymethylation in patients with obsessive compulsive disorder. J Psychiatr Res, 2019. [DOI | PubMed]
- C D’Addario, M Pucci, F Bellia, A Girella, A Sabatucci, F Fanti. Regulation of oxytocin receptor gene expression in obsessive–compulsive disorder: a possible role for the microbiota-host epigenetic axis. Clin Epigenetics, 2022. [DOI | PubMed]
- Y Huang, H Yasuda, A Sarihi, T Tsumoto. Roles of endocannabinoids in heterosynaptic long-term depression of excitatory synaptic transmission in visual cortex of young mice. J Neurosci, 2008. [DOI | PubMed]
- F Lemtiri-Chlieh, ES Levine. BDNF evokes release of endogenous cannabinoids at layer 2/3 inhibitory synapses in the neocortex. J Neurophysiol, 2010. [DOI | PubMed]
- LE Maglio, JA Noriega-Prieto, MJ Maraver, D Fernández de Sevilla. Endocannabinoid-dependent long-term potentiation of synaptic transmission at rat barrel cortex. Cereb Cortex, 2018. [DOI | PubMed]
- F Bellia, A Girella, E Annunzi, B Benatti, M Vismara, A Priori. Selective alterations of endocannabinoid system genes expression in obsessive compulsive disorder. Transl Psychiatry, 2024. [DOI | PubMed]
- TD Jung, PS Jung, L Raveendran, Y Farbod, A Dvorkin-Gheva, B Sakic. Changes in gut microbiota during development of compulsive checking and locomotor sensitization induced by chronic treatment with the dopamine agonist quinpirole. Behav Pharmacol, 2018. [DOI | PubMed]
- X Cao, K Liu, J Liu, Y-W Liu, L Xu, H Wang. Dysbiotic gut microbiota and dysregulation of cytokine profile in children and teens with autism spectrum disorder. Front Neurosci, 2021. [DOI | PubMed]
- J Turna, K Grosman Kaplan, R Anglin, B Patterson, N Soreni, P Bercik. The gut microbiome and inflammation in obsessive-compulsive disorder patients compared to age- and sex-matched controls: a pilot study. Acta Psychiatr Scand, 2020. [DOI | PubMed]
- IM Scheepers, JF Cryan, TFS Bastiaanssen, K Rea, G Clarke, HB Jaspan. Natural compulsive-like behaviour in the deer mouse (Peromyscus maniculatus bairdii) is associated with altered gut microbiota composition. Eur J Neurosci, 2020. [DOI | PubMed]
- A Fattorusso, L Di Genova, GB Dell’Isola, E Mencaroni, S Esposito. Autism spectrum disorders and the gut microbiota. Nutrients, 2019. [DOI | PubMed]
- SE Swedo, J Seidlitz, M Kovacevic, ME Latimer, R Hommer, L Lougee. Clinical presentation of pediatric autoimmune neuropsychiatric disorders associated with streptococcal infections in research and community settings. J Child Adolesc Psychopharmacol, 2015. [DOI | PubMed]
- SR Kamble, MP Dandekar. Implication of microbiota gut–brain axis in the manifestation of obsessive-compulsive disorder: preclinical and clinical evidence. Eur J Pharmacol, 2023. [DOI | PubMed]
- J Haller, B Varga, C Ledent, I Barna, TF Freund. Context-dependent effects of CB1 cannabinoid gene disruption on anxiety-like and social behaviour in mice. Eur J Neurosci, 2004. [DOI | PubMed]
- A Manduca, V Buzzelli, A Rava, A Feo, E Carbone, S Schiavi. Cannabidiol and positive effects on object recognition memory in an in vivo model of fragile X syndrome: Obligatory role of hippocampal GPR55 receptors. Pharmacol Res, 2024. [DOI | PubMed]