
Cannabidiol-Induced Autophagy Ameliorates Tau Protein Clearance
Abstract
Graphical Abstract:
Supplementary Information:
The online version contains supplementary material available at 10.1007/s12640-025-00729-3.
Article type: Research Article
Keywords: CBD, Tau protein, Autophagy, Neuroprotection
Affiliations: https://ror.org/02k5swt12grid.411249.b0000 0001 0514 7202Department of Pharmacology, Escola Paulista de Medicina, Universidade Federal de São Paulo, Rua Três de Maio, 100, São Paulo, SP CEP: 04044-020 Brazil; https://ror.org/02k5swt12grid.411249.b0000 0001 0514 7202Department of Biological Sciences, Universidade Federal de São Paulo, Diadema Campus, Diadema, SP Brazil; https://ror.org/02k5swt12grid.411249.b0000 0001 0514 7202Laboratory of Molecular and Translational Endocrinology, Escola Paulista de Medicina, Universidade Federal de São Paulo, São Paulo, SP Brazil; National Institute for Translational Medicine (INCT-TM, CNPq), Ribeirão Preto, Brazil; https://ror.org/036rp1748grid.11899.380000 0004 1937 0722Department of Neuroscience and Behavior, Universidade de São Paulo, USP, Ribeirão Preto, Brazil; https://ror.org/02jx3x895grid.83440.3b0000 0001 2190 1201Department of Cell and Developmental Biology, University College London, London, UK
License: © Crown 2025 CC BY 4.0 Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
Article links: DOI: 10.1007/s12640-025-00729-3 | PubMed: 39900844 | PMC: PMC11790692
Relevance: Relevant: mentioned in keywords or abstract
Full text: PDF (3.4 MB)
Introduction
Tau is a phosphoprotein predominantly expressed in cells of the central nervous system (CNS) that confers stability to microtubules (Wang and Liu ref. 2008). The excessive phosphorylation, misfolding and aggregation of tau protein cause cellular stress, leading to activation of cell death pathways, and consequently neurodegeneration (Fontaine et al. ref. 2015). Some neurodegenerative diseases can be classified as tauopathies, including those characterized by dementia, such as Alzheimer’s disease (AD), frontotemporal dementia (FTD), Pick’s disease and progressive supranuclear palsy (Goedert and Spillantini ref. 2011). AD is the most common neurodegenerative disease associated with aging and one of the main causes of dementia in the elderly (Castellani et al. ref. 2010). The pathophysiology of AD is characterized by the presence of two distinct biomarkers: (1) amyloid plaques, composed by β-amyloid peptide (βA) that aggregates in extracellular space and (2) neurofibrillary tangles, composed by hyperphosphorylated tau, which leads to cellular stress and cognitive impairment (Fulga et al. ref. 2007). Hyperphosphorylated tau detaches from microtubules disrupting its structure and leading to intracellular accumulation of isolated tau in neurons (Stoothoff and Johnson ref. 2005). In addition, studies suggest that in tauopathies, including AD, there is an impairment of protein degradation pathways, such as autophagy (Nassif and Hetz ref. 2012; Zhang et al. ref. 2017; Festa et al. ref. 2021).
Autophagy is a lysosomal catabolic degradation process based on double-membrane vesicles formation called autophagosomes, and it is essential for survival, development, and maintenance of cellular homeostasis (Yang and Klionsky ref. 2010). The blockage of autophagy can induce tau accumulation and aggregation in neurodegenerative processes, which was observed in in vivo and in vitro models of tauopathies (Liu et al. ref. 2009; Salama et al. ref. 2018; Silva et al. ref. 2019) and in post-mortem brains from AD patients (Piras et al. ref. 2016; Long et al. ref. 2020). Studies have demonstrated that autophagy is one of the main pathways involved in tau degradation and that dysfunctions in the autophagy-lysosome system lead to the formation of insoluble tau oligomers and aggregates, while the induction of autophagy can reduce the amount of these aggregates (Congdon et al. ref. 2012; Krüger et al. ref. 2012; Caballero et al. ref. 2021; Hamano et al. ref. 2021). Several studies have shown that the endocannabinoid system can modulate autophagy (Gugliandolo et al. ref. 2020). This system, which includes cannabinoid receptors type 1 (CB1R) and type 2 (CB2R), endogenous cannabinoids, and the enzymes involved in their synthesis and degradation, play a crucial role in maintaining the balance between protein aggregation and clearance. Growing evidence underscores its capacity to regulate autophagic processes, leading to insights into the mechanisms underlying neurodegenerative diseases (Hiebel et al. ref. 2014; Basavarajappa et al. ref. 2017; Gugliandolo et al. ref. 2020).
The deletion for CB1R in mice promoted the pathological accumulation of proteins, which are not degraded by lysosomal enzymes through autophagic flux, influencing the onset and the course of brain aging (Piyanova et al. ref. 2013). The deletion of CB2R impairs the accumulation of the autophagic protein LC3-II in the spinal cord of an experimental model of autoimmune encephalomyelitis in mice, while the activation of CB2R reverted this effect (Shao et al. ref. 2014). Recently, we demonstrated that cannabidiol (CBD) induced autophagy in neural cells through the activation of CB1, CB2 and TRPV1 receptors by regulating the phosphorylation of ERK1/2 and AKT kinases in a mTORC1 independent manner (Vrechi et al. ref. 2021). Thus, the modulation of autophagy by cannabinoids could be considered as a new strategy and a possible approach to deal with neurodegenerative processes, since it also suppresses the accumulation of proteins, eliminating aggregation-prone proteins and damaged organelles that may be involved in neurodegeneration (Hara et al. ref. 2006; Komatsu et al. ref. 2006).
Here we investigated the possible neuroprotective role of CBD in a neuroblastoma cellular model of tauopathy, in an inducible Tet-On system that conditionally overexpresses tau. Our study provides evidence that CBD induces autophagy and reduces the accumulation of tau, which could represent a potential new therapeutic strategy for neurodegeneration.
Methods
Drugs
Cannabidiol (CBD) was purchased from BSPG-Pharm (Sandwich, Kent, UK); Arachidonyl-2´-chloroethylamide hydrate (ACEA), 1-(2,3-Dichlorobenzoyl)-5-methoxy-2-methyl-(3-(morpholin-4-yl)ethyl)-1 H-indole hydrochloride (GW-405833), wortmannin, chloroquine, doxycycline hydrochloride hemiethanolate hemihydrate, ammonium chloride and Earle’s Balanced Salt Solution (EBSS) were purchased from Sigma-Aldrich Chemical Co. (St Louis, MO, USA).
Cell Culture
Human neuroblastoma (SH-SY5Y) cell line was maintained in high glucose Dulbeccos’s modified Eagle’s medium, supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin (Thermo Fisher Scientific) and kept in 37 °C in a humidified CO2 atmosphere. The undifferentiated cells used as tauopathy model was previously established and described (Costa et al. ref. 2022). Briefly, cells were transduced using inducible Tet-On conditional expression system, containing the tetracycline-controlled reverse transactivation. Cells were infected with the pLVX-Tight-Puro-EGFP-Tau WT vector, to conditionally express tau. The Tet-On system and the expression of EGFP-Tau(WT) is activated by doxycycline (1 µg/mL) treatment for 72 h.
Cell Viability Assay
To investigate the neuroprotective role of cannabinoid agonists in a tauopathy cellular model, the SH-SY5Y cell line was transduced with the lentivirus containing the vector pVLX-GFP-Tau0N4R Wild type (WT), as previously described (Costa et al. ref. 2022). Cells were plated and the Tet-On system was activated with doxycycline (1 µg/mL) for 72 h. After this period, cells were treated with CBD (100, 250 nM, 1 and 10 µM), ACEA and GW405833 (100, 250 nM, 1 and 2 µM) for 24 h. Cell viability was performed using the 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT, Sigma-Aldrich) reduction assay. To evaluate cell viability, the culture medium was removed and MTT solution (0.5 mg/mL) in serum-free DMEM was added and incubated at 37 °C for 4 h. The medium was then gently removed, and the formazan crystals were solubilized in dimethyl sulfoxide (DMSO, Sigma-Aldrich). Colorimetric determination of MTT reduction was measured at 570 nm with a reference wavelength at 630 nm in a microplate reader (BioTek). Data are presented as relative expression to control group, normalized as 100%.
Western Blotting
To assess whether treatment with CBD could reduce the cytosolic levels of tau, the system was activated with doxycycline (1 µg/mL) for 72 h, followed by treatment with CBD (100, 250 nM, 1 and 10 µM) for 24 h. After, cells expressing EGFP-Tau WT and treated with CBD were collected in lysis buffer (150 mM NaCl, 1% NP-40, 0.5% deoxycholic acid, 0.1% SDS, 50 mM Tris pH 8.0, and 2 mM MgCl2) supplemented with protease and phosphatase inhibitors cocktails. The insoluble fraction was removed by centrifugation at 15,000 ×g, 4 °C for 10 min and the protein concentration in supernatants was determined using the Bradford assay (Bio-Rad). Samples were prepared in NuPAGE™ LDS Sample Buffer (4X) sample buffer (Thermo Fischer Scientific). Total protein lysates (20–30 µg) were loaded and subjected to SDS-PAGE, transferred to PVDF membranes (Millipore) using a Trans-Blot cell system (Bio-Rad) and blocked for 1 h at room temperature with 5% non-fat dry milk in Tris buffered saline solution containing 1% Tween-20 (TBS-T). Membranes were incubated with primary antibodies overnight at 4 °C: anti-LC3B (Cell Signaling Technology Inc., #2775S, 1:2000,), anti-p-AT8-tau (pSer202/pTrh205, Thermo Fisher Scientific, #MN1020, 1:1000), anti-p-AT180-tau (Trh231, Thermo Fisher Scientific, #MN1040, 1:1000), anti-tau-5 (Abcam, #ab80579, 1:1000) and anti-p62 (MBL #PM045, 1:2000), α-tubulin (Sigma-Aldrich, #T8203, 1:5000) or GAPDH (Sigma-Aldrich, #G8795, 1:5000) as housekeep control. The membranes were incubated with appropriate horseradish peroxidase-conjugate secondary antibody (1:5,000) for 1 h at room temperature. Protein signals were visualized using the Western Lightning Plus-ECL chemiluminescence system (Perkin Elmer) and the luminescence was captured using an Uvitec chemidoc-imaging platform (UVITEC Alliance 4.7, Cambridge). The quantification was performed using the Uvitec Alliance software and the protein bands were normalized relative to the internal control expression. Data are presented as relative expression to control group.
mCherry-LC3 Puncta Formation
The EGFP-Tau WT cells were plated on 25 mm glass coverlips and treated with doxycycline for 72 h. Cells were transfected as previously described in Costa et al. (ref. 2022), using mCherry-LC3 (5 µg) and lipofectamine 3000 reagent (10 µL) (Thermo Fisher Scientific) according to manufacturer’s protocol. After 24 h of plasmid expression, cells were treated with CBD (100 nM and 10 µM) or Earle’s Balanced Salt Solution (EBSS) medium for 2 h. Cells were fixed with 4% paraformaldehyde and coverslips were prepared in microscope slides with Fluoromount (Sigma Aldrich). Images were randomly acquired in a Zeiss LSM 780 confocal microscope (Zeiss LSM 780 Axiovert 200 M, Carl Zeiss) at 63× magnification, using 543/615 nm emission filters, and analyzed using the ImageJ software (NIH).
Measurement of Tau Clearance
Cells were cultured (1 × 104) in black 96-well plates, followed by the activation of Tet-On system with doxycycline (1 µg/mL) for 72 h. After this period, cells were pretreated with wortmannin (250 nM) and chloroquine (25 µM) for 30 min, followed by the treatments with CBD (100, 250 nM, 1 and 10 µM), ACEA and GW-405,833 (100, 250 nM, 1 and 2 µM) for 24 h. Nuclei were stained with Hoechst 33,342 (2 µg/mL) for 15 min. The EGFP-tau fluorescence was analyzed in a high-content screening equipment (IN Cell Analyzer 2200, GE Healthcare). For each condition tested, a total of 56 images were acquired from wells, in a 40x magnification. Images were analyzed by the IN Cell Investigator High-Content analysis software v1.3, and data were expressed as the average fluorescence intensity normalized by the control group (Dox+).
Statistics
Comparisons between groups were performed using one or two-way Analysis of Variance (ANOVA) test with the subsequent Dunnett, Tukey or Sidak post-hoc tests when appropriate. For all assessments, the assumed significance level was p ≤ 0.05 and data are expressed as the mean ± standard error of the mean (S.E.M.).
Results
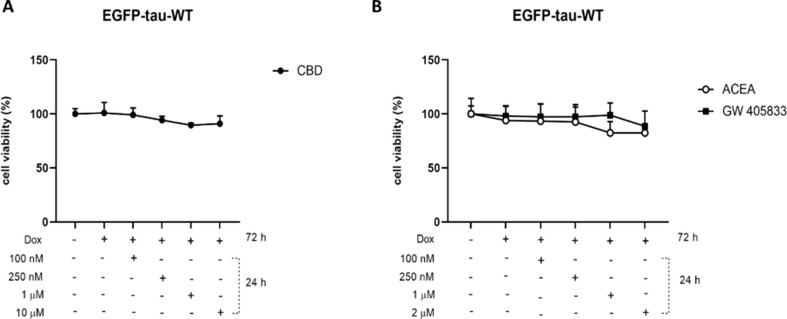
CBD, ACEA and GW-405,833 Treatments do not Affect Cell Viability
To characterize the overexpression of EGFP-Tau WT construct (~ 90 kD), the expression of total tau (Tau-5) was evaluated after 72 h of system activation with doxycycline (1 µg/mL) (Suppl. Figure 1 A-B). Data demonstrated the increase in tau-5 expression in the group treated with doxycycline, when compared to non-treated control group. Immunoblot analysis also showed no variation in endogenous tau when the Tet-On system is activated. After the activation of Tet-On system, the toxicity of cannabidiol and the selective agonists ACEA (CB1) and GW-405,833 (CB2) receptors, were evaluated. As shown in Fig. 1, there were no significant alterations in cell viability in the groups.
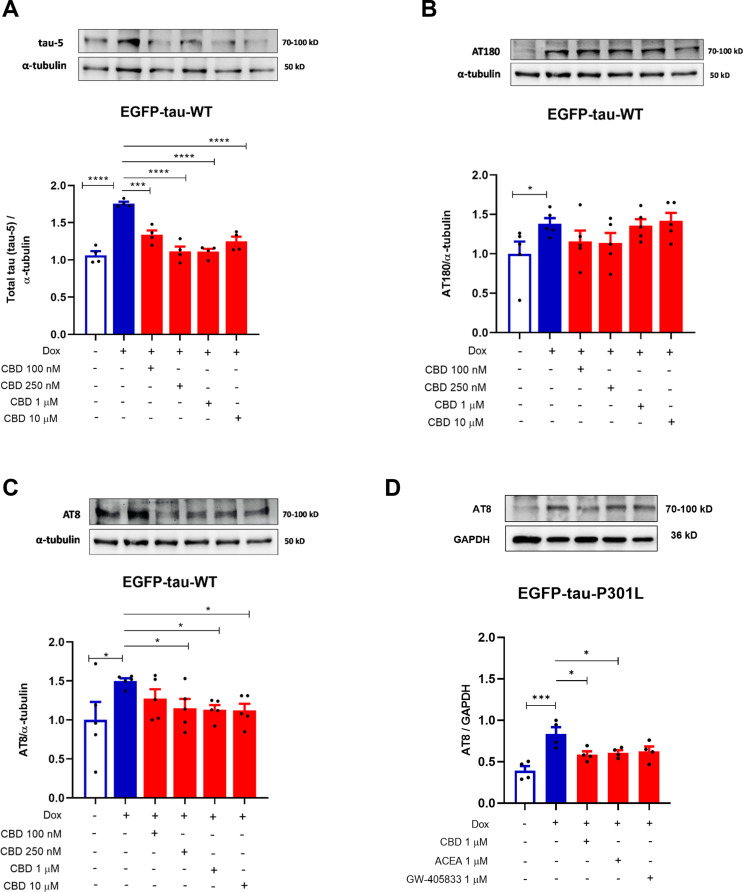
Cannabidiol Induces Tau Clearance
Next, after in Tau expressing SH-SY5Y cells (Dox+) presented a significant increase of tau-5 and its sites of phosphorylation AT8 (pSer202/pThr205) and AT180 (pThr231) (69% (p < 0.001); 50%, p = 0.0206 and 38.7%, p = 0.0490; respectively), when compared to the control (Dox−) (Fig. 2A-C). The results showed that the treatment with CBD significantly decreased the levels the phosphorylation sites at AT8 pSer202/pThr205 (tau AT8) by 35% (p = 0.020), 37% (p = 0.015) and 38% (p = 0.013), at 250 nM, 1 and 10 µM, respectively (Fig. 2A). On the other hand, no significant differences were observed in the AT180 phosphorylation (Fig. 2B). Additionally, CBD was able to decrease total tau (Tau-5) by 42% (p < 0.001), 64%, 64% and 50% (p < 0.0001) at 100, 250 nM, 1 and 10 µM, respectively, when compared to Dox+ (Fig. 2C). These data indicated that activation of Tet-On system induces the accumulation of tau total (Tau-5) and the phosphorylation of AT8 and AT180 proteins. Importantly, all concentrations tested of CBD abrogated the tau-5 and AT8 phosphorylation. Given this, we investigated the effect of CBD and the cannabinoid agonists ACEA and GW405833 on the AT8 phosphorylation site in the EGFP-tau-P301L cell line, which carries a mutated tau protein. Interestingly, CBD and ACEA at 1 µM after 24 h also reduced phosphorylation of AT8 in mutated tau by 30% (p = 0.0251) and 27% (p = 0.0465), respectively (Fig. 2D).
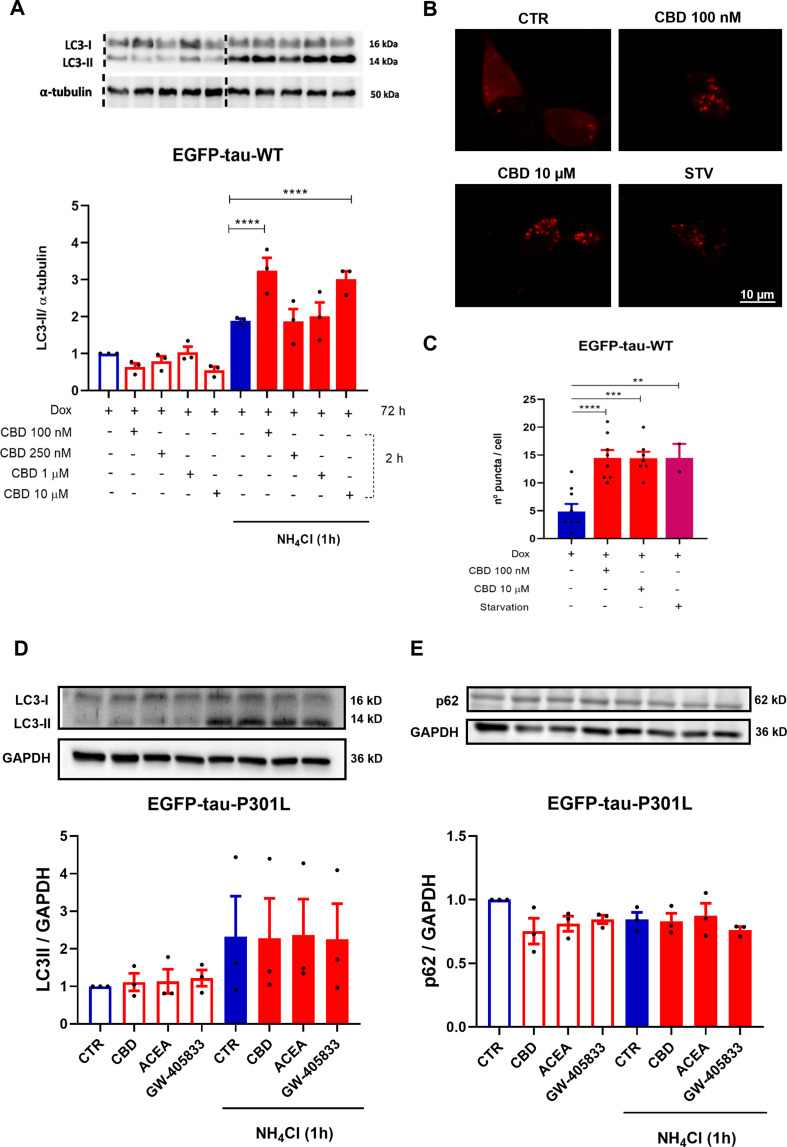
CBD Induces Autophagy in Tau WT Expressing SH-SY5Y Cell Line
Furthermore, investigating the role of autophagy in tau degradation mediated by CBD. For this purpose, LC3-II protein was quantified in the EGFP-Tau WT cell line after treatment with CBD (100, 250 nM, 1 and 10 µM) for 2 h. The cells were also treated in presence and absence of ammonium chloride (NH4Cl, 2 mM), an inhibitor of the final step of autophagic flux, added in the last hour of treatment. As demonstrated in the representative immunoblots, the groups treated with CBD 100 nM and 10 µM had a further increase in LC3-II accumulation, 135.9% and 112.4% (p < 0.0001), when compared to NH4Cl (Fig. 3A). To evaluate the potential of CBD in inducing autophagy, the EGFP-Tau WT cell line overexpressing mCherry-LC3 was treated with CBD at 100 nM and 10 µM for 2 h, as previously demonstrated to activate autophagy. As a positive control, the cells were subjected to nutritional deprivation (starvation – STV) by Earle’s Balanced Salt Solution (EBSS), also for 2 h. The number of autophagosomes was performed in single-cells analysis under a confocal microscope (Fig. 3B). The data showed that there was an increase of mCherry puncta (LC3-II) of 96.1% (p < 0.0001), 95.4% (p = 0.0001) and 96.1% (p = 0.0097), in the CBD (100 nM), CBD (10 µM) and STV treated groups, respectively, when compared to the control (Dox+) (Fig. 3C). At the same concentrations, that CBD increased the levels of LC3-II protein in the immunoblot, it also induced the formation of autophagosome, which were observed in cells transfected with mCherry-LC3. In addition, to investigate the effect of CBD on autophagy in mutated tau, SH-SY5Y cells expressing EGFP-Tau-P301L were used. In contrast to prior findings, CBD and the selective cannabinoid agonists ACEA and GW-405,833 at 1 µM for 2 h did not modulate the levels of LC3-II (p = 0.9844) or p62 (p = 0.9749) (Fig. 3D-E, respectively).
CBD-Induced Autophagy Reduces Tau Accumulation Via Autophagy
As previously demonstrated, CBD induces autophagy and a tau reduction after 2 and 24 h of treatment. Using selective CBR agonists, EGFP-Tau WT-overexpressing cells were treated at different concentrations CBD, ACEA and GW405833 in association with the autophagic inhibitors: wortmannin (250 nM) and chloroquine (25 µM). Wortmannin is a specific inhibitor of phosphoinositidine 3-kinases (PI3Ks), used as an inhibitor of autophagy since it inhibits the initiation of the autophagosome (HUNG et al., ref. 2009). Chloroquine is an autophagic blocker since it interferes in the fusion of autophagosome and lysosome. Thus, after the activation of Tet-On system, cells were pretreated with the autophagic inhibitors for 30 min, followed by CBD treatment and the selective agonists of CB1 and CB2, ACEA and GW405833 (1 and 2 µM), respectively for 24 h. The quantification of GFP fluorescence demonstrated that CBD (1 and 10 µM) decreased the fluorescence intensity of EGFP-Tau WT by 34.8% (p = 0.014) and 39.9% (p = 0.004), respectively, when compared to the control (Dox+) (Fig. 4A). The association of CBD with wortmannin did not affect the fluorescence intensity when compared to the control group. The groups co-treated with chloroquine increased 45% and 46% (p = 0.005) in fluorescence intensity in the groups treated with 1 µM and 10 µM CBD, respectively. These data demonstrated that chloroquine blocked the autophagic flux, causing the accumulation of tau and, consequently, increased the intensity of GFP fluorescence.
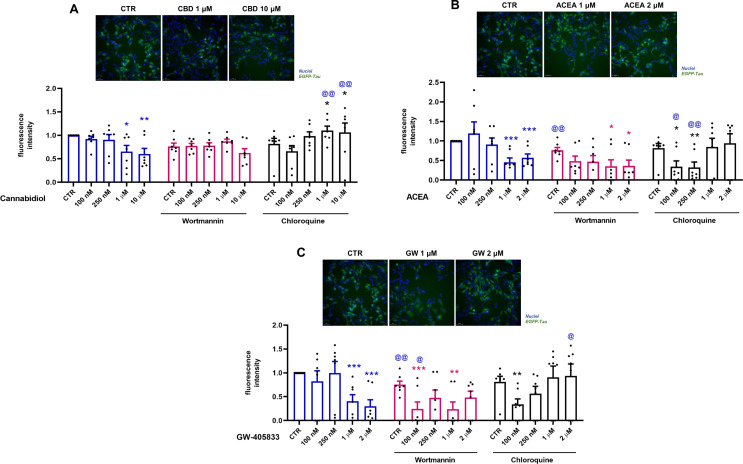
Cells treated with ACEA showed a significant decrease of 54.6% and 42.9% (p = 0.001) in the fluorescence intensity of EGFP-Tau WT at 1 and 2 µM, respectively (Fig. 4B). The association of ACEA and wortmannin caused a decrease of 23.9% (p = 0.004) of EGFP-Tau WT fluorescence when compared to control. Additionally, wortmannin at 1 and 2 µM led to a decrease of 41% (p = 0.027) and 40% (p = 0.025) in fluorescence intensity in relation to the control, respectively.
The co-treatment with chloroquine decreased in 47.7% (p = 0.017) and 49.5% (p = 0.005) compared to the control and 85.2% (p = 0.017) and 58.6% (p = 0.010) at 100 and 250 nM, respectively.
Cells treated with GW-405,833 showed a significant decrease of 59.3% and 69.9% (p = 0.001) in the EGFP-Tau WT fluorescence intensity at 1 and 2 µM, respectively (Fig. 4C). Co-treatment of GW-405,803 with wortmannin led to a further decreased of 51% (p = 0.001) e 51.1% (p = 0.0.007) in fluorescence intensity in relation to the wortmannin control, at 100 nM and 1 µM, respectively. Also, GW-405,803 was able to decrease 24.8% (p = 0.002) and 75.8% (p = 0.027) in relation to their respective groups without inhibitors, control group and group treated with 100 nM, respectively. The co-treatment with chloroquine showed a reduction of 47% (p = 0.010) in relation to chloroquine alone and an increase of 93.03% (p = 0.034) in the group treated with 2 µM in relation to its non-treated group (Fig. 4C). In summary, these data showed that the cannabinoids CBD, ACEA and GW-405,833 decreased the intensity of tau fluorescence. However, when using the autophagic inhibitors, wortimanin and chloroquine, only the CBD (1 and 10 µM) and GW-405,833 (2 µM) + chloroquine groups, reversed the decrease in the intensity of EGFP-Tau WT, demonstrating the possible participation of autophagy in the degradation of tau in these groups.
Discussion
Several studies have reported a possible crosstalk between the tau hyperphosphorylation and autophagy dysfunctions (Nassif and Hetz ref. 2012; Silva et al. ref. 2019; Di Meco et al. ref. 2020). Tau hyperphosphorylations were found in Atg7 knockout mouse brains, which was reverted after the restoration of autophagy (Inoue et al. ref. 2012). Besides the assembly and the stabilization of microtubules, tau protein is crucial for the maintenance of retrograde traffic and maturation of autophagosomes, as well as their fusion with lysosomes (Dickey et al. ref. 2007). In fact, tau hyperphosphorylation can lead to instability of the microtubule cytoskeleton, which can inhibit autophagosome traffic and favor the accumulation of immature autophagosomes in axons (Nassif and Hetz ref. 2012; Piras et al. ref. 2016).
Increasing evidence indicate that the activation of CB1 and CB2 receptors, by natural or synthetic agonists, mediates neuroprotective effects in in vitro and in vivo models of AD, reducing the deleterious effects of βA peptide or tau hyperphosphorylation (Esposito et al. ref. 2007; Martin-Moreno et al. ref. 2011; Janefjord et al. ref. 2014; Ahmed et al. ref. 2015). Furthermore, Esposito et al. (ref. 2006) showed that CBD (from 100 nM to 10 µM) inhibited tau hyperphosphorylation in PC12 cell line stimulated by βA peptide (Esposito et al. ref. 2006).
Here the high levels of AT8 levels in EGFP-Tau WT cells and in the mutated tau cell line EGFP-Tau-P301L were attenuated by the treatment with CBD (250 nM, 1 or 10 µM) for 24 h. These results showed that the Tet-On system activation induced tau AT8 on pSer202/pThr205 sites, and CBD treatment decreased these phosphorylations, indicating a potential neuroprotective effect. Interestingly, CBD (100, 250 nM, 1 and 10 µM, 24 h) was able to decrease total tau, strongly suggesting a total tau clearance. This evidence was reported in other studies, in mice overexpressing human tau (PK−/−TauVLW), a frontotemporal complex model, Sativex® induced a marked reduction in neurofibrillary tangles (Casarejos et al. ref. 2013).
However, it was not possible to observe a decrease in AT180 phosphorylation after treatment with CBD (100, 250 nM, 1 and 10 µM, 24 h). The pThr231 (Tau AT180) was observed only in some neurofibrillary tangles, most of which were found in amyloid plaques in the cortex of patients with AD and FTD (Spillantini et al. ref. 1996). However, a study using a transgenic model in zebrafish, expressing human tau P301L, has demonstrated that AT180 is present in tau hyperphosphorylation, but there was not related to neurofibrillary tangles formation (Cosacak et al. ref. 2017).
Several studies have suggested that cannabinoid compounds (Δ9-THC and CBD) potentiated autophagy reducing the deposition of tau proteins. Piras et al. (ref. 2016) showed that hyperphosphorylated tau was co-localized on surfaces labeled for LC3-II and p62 proteins in the brain of patients with progressive supranuclear palsy and corticobasal degeneration (Piras et al. ref. 2016). Our results demonstrated autophagy activation in cells overexpressing EGFP-Tau WT cells after treatment with CBD (100 nM and 10 µM, 2 h). This result was confirmed in EGFP-Tau WT cells transfected with the mCherry-LC3, indicating an increase in the number of puncta, indicative of autophagosome formation.
Next, to evaluate the participation of autophagy on tau expression, the EGFP-Tau WT cells were treated with the cannabinoid compounds CBD, ACEA and GW-405,833 with the pre-treatment with classic autophagic inhibitors wortmannin or chloroquine. The use of ACEA (CB1 agonist) and GW-405,833 (CB2 agonist) stems from their ability to provide targeted insights into the specific pathways mediated by cannabinoid receptor activation. Unlike CBD, which interacts with multiple biological targets, these selective agonists enable a focused evaluation of the roles of CB1 and CB2 in autophagy and tau clearance, complementing the broader effects observed with CBD. The results showed that the cannabinoid compounds: CBD (1 and 10 µM), ACEA and GW-405,833, at 1 and 2 µM, were able to significantly reduce the intensity of fluorescence in the EGFP-Tau WT cells. Libro et al. (ref. 2017) also observed that CBD led to the downregulation of genes linked to AD, including genes that encode kinases responsible for phosphorylation of tau (Libro et al. ref. 2017). Furthermore, other studies observed that ACEA (CB1 agonist) and WIN55,212-2 (CB1/CB2 agonist) decreased tau phosphorylation in mouse models of AD (Esposito et al. ref. 2006; Aso et al. ref. 2012). We also investigated the effects of CBD on autophagy modulation in mutated tau-P301L. CBD, ACEA, and GW-405,833 were found to block autophagy flux in this cell line, as reported by Costa et al. (ref. 2022). Then, in order to evaluate the role of autophagy in tau clearance mediated by cannabinoid compounds, EGFP-Tau WT cells were treated with wortmannin (500 nM, 30 min) prior of treatment with different concentrations of CBD, ACEA and GW-405,833. Wortmannin, a non-specific inhibitor of PI3K-related enzymes (Feldman and Shokat ref. 2010), consequently inhibiting the autophagosome formation (Hung et al. ref. 2009). Interestingly, the wortmannin-only group had a significant decrease in EGFP-Tau WT fluorescence intensity. Thus, the cannabinoid compounds ACEA and GW-405,833 decreased the intensity of tau fluorescence, but in an mTOR-independent manner, since wortmannin inhibits PI3K enzymes related to this pathway. These data corroborate previous data published by our group, that shows that CBD activates autophagy in a mTOR-independent manner (Vrechi et al. ref. 2021). However, how wortmannin increased the intensity of tau (hyperphosphorylation) in EGFP-Tau WT cells have not been elucidated. In addition, the EGFP-Tau WT cells were also treated with chloroquine. Of note, groups treated with chloroquine and CBD (1 and 10 µM) increased GFP intensity. These results suggested CBD was able to decrease EGFP-Tau WT via autophagy, since chloroquine potentially inhibit the lysosomal degradation, the last step of autophagic process. One study revealed that chloroquine (50 µM) induced the accumulation of autophagic vacuoles and blocked autophagic outflow in retinoic acid-differentiated SH-SY5Y cells (Pivtoraiko et al. ref. 2010). Taken together, the collecting data provides evidence that CBD was able to reduce total tau via AT8 pSer202/pTrh205 dephosphorylation and autophagy activation. Moreover, the cannabinoid compounds CBD, ACEA and GW-504,833 were able to decrease the fluorescence intensity of the EGFP-Tau WT, demonstrating the possible protein degradation in this model. Furthermore, in the presence of chloroquine, an autophagic inhibitor, the treatment with CBD (1 and 10 µM) and GW-405,833 (2 µM) reversed tau clearance, indicating the participation of autophagy in tau degradation in these groups.
Conclusion
CBD induces autophagy promoting tau clearance in an in vitro model of tauopathy. Moreover, CBD, ACEA and GW-405,833 decreased tau expression, which was reversed by chloroquine indicating that autophagy participates in tau clearance. Our results support the relevance of cannabinoid compounds in the autophagic process involved in the degradation of accumulated tau, which has been associated with several neuropathies. Therefore, autophagy is a potential therapeutic target of cannabinoids in neurodegenerative diseases.
Supplementary Materials
References
- E Aso, E Palomer, S Juvés. CB1 agonist ACEA protects neurons and reduces the cognitive impairment of AβPP/PS1 mice. J Alzheimer’s Dis, 2012. [DOI | PubMed]
- BS Basavarajappa, M Shivakumar, V Joshi, S Subbanna. Endocannabinoid system in neurodegenerative disorders. J Neurochem, 2017. [DOI | PubMed]
- B Caballero, M Bourdenx, E Luengo. Acetylated tau inhibits chaperone-mediated autophagy and promotes tau pathology propagation in mice. Nat Commun, 2021. [DOI | PubMed]
- MJ Casarejos, J Perucho, A Gomez. Natural cannabinoids improve dopamine neurotransmission and tau and amyloid pathology in a mouse model of tauopathy. J Alzheimer’s Dis, 2013. [DOI | PubMed]
- RJ Castellani, RK Rolston, MA Smith. Alzheimer disease. Dis Mon, 2010. [DOI | PubMed]
- EE Congdon, JW Wu, N Myeku. Methylthioninium chloride (methylene blue) induces autophagy and attenuates tauopathy in vitro and in vivo. Autophagy, 2012. [DOI | PubMed]
- MI Cosacak, P Bhattarai, L Bocova. Human TAUP301L overexpression results in TAU hyperphosphorylation without neurofibrillary tangles in adult zebrafish brain. Sci Rep, 2017. [DOI | PubMed]
- AJ Costa, RB Oliveira, P Wachilewski. Membrane estrogen receptor ERα activation improves tau clearance via autophagy induction in a tauopathy cell model. Brain Res, 2022. [DOI | PubMed]
- A Di Meco, ME Curtis, E Lauretti, D Praticò. Autophagy dysfunction in Alzheimer’s Disease: mechanistic insights and New Therapeutic opportunities. Biol Psychiatry, 2020. [DOI | PubMed]
- CA Dickey, A Kamal, K Lundgren. The high-affinity HSP90-CHIP complex recognizes and selectively degrades phosphorylated tau client proteins. J Clin Invest, 2007. [DOI | PubMed]
- G Esposito, D De Filippis, R Carnuccio. The marijuana component cannabidiol inhibits β-amyloid-induced tau protein hyperphosphorylation through Wnt/β-catenin pathway rescue in PC12 cells. J Mol Med, 2006. [DOI | PubMed]
- G Esposito, C Scuderi, C Savani. Cannabidiol in vivo blunts b-amyloid induced neuroinflammation by suppressing IL-1b and iNOS expression. Br J Pharmacol, 2007. [DOI | PubMed]
- ME Feldman, KM Shokat. New inhibitors of the PI3K-Akt-mTOR pathway: insights into mTOR Signaling from a New Generation of Tor kinase domain inhibitors (TORKinibs). Phosphoinositide 3-kinase Heal Dis, 2010. [DOI]
- BP Festa, AD Barbosa, M Rob, DC Rubinsztein. The pleiotropic roles of autophagy in Alzheimer’s disease: from pathophysiology to therapy. Curr Opin Pharmacol, 2021. [DOI | PubMed]
- SN Fontaine, JJ Sabbagh, J Baker. Cellular factors modulating the mechanism of tau protein aggregation. Cell Mol Life Sci, 2015. [DOI | PubMed]
- TA Fulga, I Elson-Schwab, V Khurana. Abnormal bundling and accumulation of F-actin mediates tau-induced neuronal degeneration in vivo. Nat Cell Biol, 2007. [DOI | PubMed]
- M Goedert, MG Spillantini. Pathogenesis of the tauopathies. J Mol Neurosci, 2011. [DOI | PubMed]
- A Gugliandolo, F Pollastro, P Bramanti, E Mazzon. Cannabidiol exerts protective effects in an in vitro model of Parkinson’s disease activating AKT/mTOR pathway. Fitoterapia, 2020. [DOI | PubMed]
- T Hamano, S Enomoto, N Shirafuji. Autophagy and tau protein. Int J Mol Sci, 2021. [DOI]
- T Hara, K Nakamura, M Matsui. Suppression of basal autophagy in neural cells causes neurodegenerative disease in mice. Nature, 2006. [DOI | PubMed]
- C Hiebel, T Kromm, M Stark, C Behl. Cannabinoid receptor 1 modulates the autophagic flux independent of mTOR- and BECLIN1-complex. J Neurochem, 2014. [DOI | PubMed]
- SY Hung, WP Huang, HC Liou, WM Fu. Autophagy protects neuron from Aβ-induced cytotoxicity. Autophagy, 2009. [DOI | PubMed]
- K Inoue, J Rispoli, H Kaphzan. Macroautophagy deficiency mediates age-dependent neurodegeneration through a phospho-tau pathway. Mol Neurodegener, 2012. [DOI | PubMed]
- E Janefjord, JLV Mååg, BS Harvey, SD Smid. Cannabinoid effects on β amyloid fibril and aggregate formation, neuronal and microglial-activated neurotoxicity in vitro. Cell Mol Neurobiol, 2014. [DOI | PubMed]
- M Komatsu, S Waguri, T Chiba. Loss of autophagy in the central nervous system causes neurodegeneration in mice. Nature, 2006. [DOI | PubMed]
- U Krüger, Y Wang, S Kumar, EM Mandelkow. Autophagic degradation of tau in primary neurons and its enhancement by trehalose. Neurobiol Aging, 2012. [DOI | PubMed]
- R Libro, F Diomede, D Scionti. Cannabidiol modulates the expression of alzheimer’s disease-related genes in mesenchymal stem cells. Int J Mol Sci, 2017. [DOI]
- YH Liu, W Wei, J Yin. Proteasome inhibition increases tau accumulation independent of phosphorylation. Neurobiol Aging, 2009. [DOI | PubMed]
- Z Long, J Chen, Y Zhao. Dynamic changes of autophagic flux induced by Abeta in the brain of postmortem Alzheimer’s disease patients, animal models and cell models. Aging, 2020. [DOI | PubMed]
- AM Martin-Moreno, D Reigada, BG Ramirez. Cannabidiol and other Cannabinoids reduce Microglial Activation in Vitro and in vivo: relevance to Alzheimer’s Disease. Mol Pharmacol, 2011. [DOI | PubMed]
- M Nassif, C Hetz. Autophagy impairment: a crossroad between neurodegeneration and tauopathies. BMC Biol, 2012. [DOI | PubMed]
- A Piras, L Collin, F Grüninger. Autophagic and lysosomal defects in human tauopathies: analysis of post-mortem brain from patients with familial Alzheimer disease, corticobasal degeneration and progressive supranuclear palsy. Acta Neuropathol Commun, 2016. [DOI | PubMed]
- VN Pivtoraiko, AJ Harrington, BJ Mader. Low-dose bafilomycin attenuates neuronal cell death associated with autophagy-lysosome pathway dysfunction. J Neurochem, 2010. [DOI | PubMed]
- A Piyanova, O Albayram, CA Rossi. Loss of CB1 receptors leads to decreased cathepsin D levels and accelerated lipofuscin accumulation in the hippocampus. Mech Ageing Dev, 2013. [DOI | PubMed]
- M Salama, M Elhussiny, A Magdy. Dual mTORC1/mTORC2 blocker as a possible therapy for tauopathy in cellular model. Metab Brain Dis, 2018. [DOI | PubMed]
- BZ Shao, W Wei, P Ke. Activating cannabinoid receptor 2 alleviates pathogenesis of experimental autoimmune encephalomyelitis via activation of autophagy and inhibiting NLRP3 inflammasome. CNS Neurosci Ther, 2014. [DOI | PubMed]
- JM Silva, S Rodrigues, B Sampaio-Marques. Dysregulation of autophagy and stress granule-related proteins in stress-driven tau pathology. Cell Death Differ, 2019. [DOI | PubMed]
- MG Spillantini, M Goedert, RA Crowther. Comparison of the neurofibrillary pathology in Alzheimer’s disease and familial presenile dementia with tangles. Acta Neuropathol, 1996. [PubMed]
- WH Stoothoff, GVW Johnson. Tau phosphorylation: physiological and pathological consequences. Biochim Biophys Acta – Mol Basis Dis, 2005. [DOI]
- AIA Van Der Ahmed, MGM Olde Rikkert. Cannabinoids in late-onset Alzheimer’s Disease. Clin Pharmacol Ther, 2015. [DOI | PubMed]
- TAM Vrechi, AHFF Leão, IBM Morais. Cannabidiol induces autophagy via ERK1/2 activation in neural cells. Sci Rep, 2021. [DOI | PubMed]
- JZ Wang, F Liu. Microtubule-associated protein tau in development, degeneration and protection of neurons. Prog Neurobiol, 2008. [DOI | PubMed]
- Z Yang, DJ Klionsky. Eaten alive: a history of macroautophagy. Nat Cell Biol, 2010. [DOI | PubMed]
- Y Zhang, X Chen, Y Zhao. The role of ubiquitin proteasomal system and autophagy-lysosome pathway in Alzheimer’s disease. Rev Neurosci, 2017. [DOI | PubMed]