
Anandamide Influences Interleukin-1β Synthesis and IL-1 System Gene Expressions in the Ovine Hypothalamus during Endo-Toxin-Induced Inflammation
Abstract
Simple Summary:
Pro-inflammatory cytokines are considered to be one of the most important mediators affecting the function of central nervous system during an immune/inflammatory challenge. It was found that in acting on different hypothalamic nuclei, pro-inflammatory cytokines influence the centrally regulated processes including reproduction. Recently, it has been shown that the endocannabinoid system and endogenous cannabinoids may attenuate the inflammatory response. Therefore, in our study we examined the influence of anandamide, one of the earliest known endocannabinoids, on the synthesis of interleukin (IL)-1β and IL-1 system gene expressions in the hypothalamic structures involved in gonadotropin-releasing hormone (GnRH)-ergic activity, and thus the central control of reproduction, during immune stress induced by endotoxin injection. It was found that anandamide inhibited lipopolysaccharide (LPS)-stimulated synthesis of IL-1β in the hypothalamus, likely affecting posttranscriptional levels of this cytokine synthesis. Anti-inflammatory effect of anandamide at the level of central nervous system might also result from its stimulating action on IL-1 antagonist and IL-1 type II receptor gene expression. This study suggests the potential of endocannabinoids and/or their metabolites in the inhibition of inflammatory process at the level of the central nervous system, as well as their usefulness in the therapy of inflammation-induced neuroendocrine disorders, but further detailed research is required to investigate this issue.
Abstract:
This study evaluated the effect of anandamide (AEA) on interleukin (IL)-1β synthesis and gene expression of IL-1β, its type I (IL-1R1) and II (IL-1R2) receptors, and IL-1 receptor antagonist (IL-1RN) in the hypothalamic structures, involved in the central control of reproduction, during inflammation. Animals were intravenously (i.v.) injected with bacterial endotoxin-lipopolysaccharide (LPS) (400 ng/kg) or saline, and two hours after LPS administration., a third group received i.v. injection of AEA (10 μg/kg). Ewes were euthanized one hour later. AEA injection (p < 0.05) suppressed LPS-induced expression of IL-1β protein in the hypothalamus. The gene expression of IL-1β, IL-1RN, and IL-1R2 in the hypothalamic structures was higher (p < 0.05) in animals treated with both LPS and AEA in comparison to other experimental groups. AEA administration did not influence LPS-stimulated IL-1R1 gene expression. Our study shows that AEA suppressed IL-1β synthesis in the hypothalamus, likely affecting posttranscriptional levels of this cytokine synthesis. However, anti-inflammatory effect of AEA might also result from its stimulating action on IL-1RN and IL-1R2 gene expression. These results indicate the potential of endocannabinoids and/or their metabolites in the inhibition of inflammatory process at the level of central nervous system, and therefore their usefulness in the therapy of inflammation-induced neuroendocrine disorders.
Article type: Research Article
Keywords: anandamide, endocanabinoids, inflammation, endotoxin, LPS, IL-1β, hypothalamus, central nervous system
Affiliations: Department of Genetic Engineering, The Kielanowski Institute of Animal Physiology and Nutrition, Polish Academy of Sciences, 05-110 Jabłonna, Poland; m.tomczyk@ifzz.pl (M.T.); jb.tst@wp.pl (J.B.); Department of Animal Physiology, The Kielanowski Institute of Animal Physiology and Nutrition, Polish Academy of Sciences, 05-110 Jabłonna, Poland; d.tomaszewska@ifzz.pl; Faculty of Health Sciences, Warsaw School of Engineering and Health, 02-366 Warsaw, Poland; anna.herman@onet.pl
License: © 2021 by the authors. CC BY 4.0 Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Article links: DOI: 10.3390/ani11020484 | PubMed: 33673103 | PMC: PMC7918765
Relevance: Moderate: mentioned 3+ times in text
Full text: PDF (1.1 MB)
1. Introduction
During inflammation induced by viral or bacterial infection, a number of immune and non-immune cells produce cytokines and other inflammatory mediators able to affect physiological processes in the organism, including those occurring in the central nervous system (CNS) [ref. 1]. Acting on different hypothalamic nuclei, pro-inflammatory cytokines may influence the centrally regulated processes such as food intake, thermogenesis, rest/activity circadian rhythm, and reproduction [ref. 2,ref. 3]. Both acute and prolonged inflammations decrease the circulating concentration of luteinizing hormone (LH) and therefore affect the reproductive process [ref. 4,ref. 5]. These changes are connected mainly with decreased gonadotropin-releasing hormone (GnRH) secretion in the hypothalamus [ref. 6,ref. 7,ref. 8,ref. 9]. It is worth mentioning that in sheep, which were used as an animal model in the present study, GnRH neurons do not form dense clusters but are spread from the brain septum and the diagonal band of Broca through the preoptic area (POA) to the anterior hypothalamus (AHA) and the medial basal hypothalamus (MBH). Nonetheless, more than a half of the population of GnRH-ergic neurons in the hypothalamus have perikarya located in the POA [ref. 10]. The inflammation caused by numerous physiological pathways impede GnRH/LH secretion, however, centrally acting pro-inflammatory cytokine-interleukin (IL)-1β is thought to be among the most important mediators inhibiting reproductive processes during the immune challenge [ref. 1,ref. 11,ref. 12]. During inflammatory stimuli, IL-1β is produced at the periphery by specific cells, mainly monocytes and macrophages and numerous other cells including lymphocytes T and B, microglia cells, endothelial cells, astrocytes, oligodendrocytes, and neurons [ref. 13,ref. 14]. It was previously shown that peripheral cytokines (e.g., IL-1β) are able to reach the CNS by crossing the fenestrated capillaries in the structures such as the choroid plexus (CP), median eminence, or organum vasculosum of the lamina terminalis [ref. 11]. At the period of inflammation, these cytokines may also reach the brain via the blood–brain barrier by saturated, self-inhibitable transport mechanisms [ref. 15]. Moreover, in recent studies it was shown that during inflammation activated cells of the CP might express and release pro-inflammatory cytokines into the cerebrospinal fluid (CSF) [ref. 16,ref. 17,ref. 18]. However, the local synthesis of centrally acting inflammatory cytokines in the CNS is another important source of such proteins. In numerous studies, it was observed that pro-inflammatory interleukins, including IL-1β, are synthetized directly in the hypothalamus [ref. 4,ref. 14,ref. 19,ref. 20]. The ability of IL-1β to impair secretion of GnRH enables the presence of interleukin 1 receptor (IL-1R1) in the hypothalamic structures involved in GnRH-ergic activity [ref. 14,ref. 21]. Moreover, the expression of IL-1Rs was demonstrated directly on GnRH neurons. This fact proves that IL-1β ligands could take part in the modulation of these cells’ activity [ref. 22].
Recently, the majority of scientific research has been focused on the importance of the endocannabinoid system and endogenous cannabinoids. It was found that cannabinoids acting through their corresponding receptors are able to attenuate the inflammatory response [ref. 23,ref. 24]. Anandamide (AEA) is the earliest known endocannabinoid that takes part in the immune system regulation [ref. 25,ref. 26]. It may influence various physiological processes acting through two major cannabinoid receptors: type 1 (CB1) and 2 (CB2). CB2, also known as “spleen type”, is predominantly expressed by the immune cells but also, to a lesser extent, in the brain [ref. 24,ref. 27]. CB1 mainly occurs in the CNS; it has been detected in the cerebral cortex, limbic system structures, cerebellum, pituitary gland, and above all else in the hypothalamus [ref. 28]. Having in mind the fact that on immune cells both CB1 and CB2 receptors have been detected, it could be suggested that cannabinoids play a substantial role in the immune system regulation. It was demonstrated that administration of delta9-tetrahydrocannabinol (THC) into mice generated a noticeable apoptosis in T and dendritic cells. Cannabinoids may downregulate the production of cytokine and chemokine and, as presented in some models, may upregulate T-regulatory cells (Tregs) in order to suppress inflammatory responses [ref. 29]. From this point of view, cannabinoids may be considered as a potent treatment against inflammatory disorders. It is worth mentioning, that in our pilot study on ewes, the lack of effect of single AEA injection on the IL-1β and IL-1RN expression in the hypothalamus was found.
Therefore, the aim of our study was to check whether peripheral administration of AEA will influence the expression of IL-1β protein and mRNA encoding IL-1β, its type I and II receptors, and interleukin 1 receptor antagonist (IL-1RN) in the hypothalamic structures involved in GnRH-ergic activity in ewe during inflammation induced by injection of a lipopolysaccharide (LPS)—bacterial endotoxin present in the outer membrane of Gram-negative bacteria.
2. Materials and Methods
2.1. Animals and Experimental Design
The study was performed on 18 adult 6-year-old Blackhead ewes during the reproductive season (from September to October). The animal acclimation to the experimental conditions was 1 month before the experiment. Ewes body condition score was kept at an estimated level of 3 points on a 5-point scale. The animal housing system was customized for sheep requirements, according to regulations of animal welfare. Each ewe was kept in individual pen in the fold, and in order to prevent separation stress, we gave them visual contact with other animals. The good health and condition of animals was assured by commercial concentrates with hay fed to ewes and not limited access to water, according to the recommendations of the National Research Institute of Animal Production for adult ewes [ref. 30].
The synchronization of estrous cycle stage was made by the Chronogest CR (Merck Animal Health, Boxmeer, the Netherlands) according to the procedures described in our previous studies [ref. 31,ref. 32]. Each ewe had a vaginal sponge containing 20 mg of cronolone (Chronogest CR; Merck Animal Health, Boxmeer, the Netherlands) placed for 14 days. After the sponge removal, animals were intramuscularly injected with 500 IU pregnant mare’s serum gonadotropin (PMSG; Folligon; Merck Animal Health, Boxmeer, the Netherlands). The experimental procedures started 7 days following PMSG injection, in luteal phase of estrus cycle. An immune/inflammatory challenge was induced by the intravenous (i.v.) administration of 400 ng/kg LPS from Escherichia coli 055:B5 (Sigma-Aldrich, St. Louis, MO, USA), dissolved in saline (0.9% w/v NaCl) (Baxter, Deerfield, IL, USA) at a concentration of 10 mg/L.
All experimental procedures carried out on animals were performed according to the guidelines of the Local Ethics Committee of Warsaw University of Life Sciences-SGGW (authorization no. WAW2/70/2017).
2.2. Experimental Procedures
Venous catheters were implanted into the jugular vein on the day prior to the experiment. Ewes were randomly divided into 3 groups: control (n = 6), LPS-treated (n = 6), and treated with LPS and AEA (n = 6) (see Table 1). Two hours after LPS injection, the animals from the third group additionally received i.v. injection of AEA (Sigma-Aldrich, St. Louis, MO, USA) at a dose of 10 µg/kg body weight chosen on the basis of a previous study [ref. 33]. Immediately after the blood collection, ewes were euthanized, and their brains were instantaneously removed from the skulls. Then, the brain structures such as POA, AHA, and MBH were dissected and at once frozen in liquid nitrogen. Collected hypothalamic tissues were stored at −80 °C.
Table 1: Experiment organization chart.
| Group | No. of Animals | Experimental Treatment I | Dose (ng/kg) | Experimental Treatment II | Dose (µg/kg) | |
|---|---|---|---|---|---|---|
| 1 | Control | 6 | NaCl | 0 | NaCl | 0 |
| 2 | LPS treated | 6 | LPS | 400 | NaCl | 0 |
| 3 | LPS + AEA i.v. treated | 6 | LPS | 400 | AEA i.v. | 10 |
| Total number of animals | 18 | |||||
LPS: lipopolysaccharide; AEA: anandamide; i.v.: intravenous.
2.3. IL-1β Concentration Assesment
The concentrations of IL-1β in the hypothalamus were assayed using IL-1β ELISA kits (cat. no. E0041Sh; Bioassay Technology Laboratory, Shanghai, China) designed and validated for sheep. In the case of the ELISA assay, the hypothalamic tissues collected from each ewe were combined into 1 sample and homogenized according to the method described elsewhere [ref. 31]. The assays procedure was performed according to the instructions provided by the manufacturer. Plates incubation and absorbance measurement at wavelength of 450 nm were estimated using the VersaMax reader (Molecular Devices LLC., Sunnyvale, CA, USA). The assay sensitivity was 0.24 pg/mL. IL-1β concentration in the hypothalamus was computed relative to total protein content of the sample assayed by the Bradford method.
2.4. Relative Gene Expression Determination
We followed the previously published methods of Herman et al. [ref. 31]. For the total RNA isolation from the collected structures such as POA, AHA, and MBH, we used the NucleoSpin RNA kit (MACHEREY-NAGEL GmbH and Co, Düren, Germany). The concentration and purity of isolated RNA were quantified spectrophotometrically by measuring the optical density at 230, 260, and 280 nm in a NanoDrop 1000 instrument (Thermo Fisher Scientific Inc., Waltham, MA, USA). The verification of RNA integrity was carried out by electrophoresis using 1% agarose gel stained with ethidium bromide (Sigma-Aldrich, St. Louis, MO, USA). The synthesis of complementary DNA (cDNA) was performed using 1000 ng of total RNA and components of Maxima First Strand cDNA Synthesis Kit for RT-qPCR (Thermo Fisher Scientific Inc., Waltham, MA, USA).
Real-time RT-PCR analysis was carried out with the use of the HOT FIREPol EvaGreen qPCR Mix Plus (Solis BioDyne, Tartu, Estonia) and HPLC-grade oligonucleotide primers (Genomed, Warszawa, Poland) in line with the method described elsewhere [ref. 34]. Specific primers to determine the expression of the genes of interest and housekeeping genes were chosen on the basis of our previous experience (see Table 2). One reaction mixture of total volume amounting 15 μL contained 3 μL of PCR Master Mix, 10.05 μL of RNase-free water, 0.45 μL of primers (0.225 μL each primer), and 1.5 μL of the cDNA template. The PCR reactions were carried out using Rotor-Gene 6000 instrument (Qiagen, Dusseldorf, Germany) with the following protocol: 95 °C for 15 min and 30–35 cycles of 95 °C for 10 s for denaturation, 59 °C for 20 s for annealing, and 72 °C for 10 s for extension. A final melting curve analysis and agarose gel electrophoresis of PCR products were performed to verify the specificity of the amplification reaction. The relative gene expression was calculated using the comparative quantification option [ref. 35] of the Rotor Gene 6000 software 1.7. (Qiagen, Dusseldorf, Germany) with reference to the mean expression of 3 housekeeping genes: glyceraldehyde-3-phosphate dehydrogenase (GAPDH), β-actin (ACTB), and histone deacetylase1 (HDAC1).
Table 2: The list of all genes analyzed by real-time PCR.
| GenBank Acc. No. | Gene | Amplicon Size (bp) | Forward/Reverse | Sequence 5′→3′ | Reference |
|---|---|---|---|---|---|
| NM_001034034 | GAPDH(glyceraldehyde-3-phosphate dehydrogenase) | 134 | forward | AGAAGGCTGGGGCTCACT | [ref. 2] |
| reverse | GGCATTGCTGACAATCTTGA | ||||
| U39357 | ACTB(beta actin) | 168 | forward | CTTCCTTCCTGGGCATGG | [ref. 2] |
| reverse | GGGCAGTGATCTCTTTCTGC | ||||
| BC108088.1 | HDAC1(histone deacetylase1) | 115 | forward | CTGGGGACCTACGGGATATT | [ref. 2] |
| reverse | GACATGACCGGCTTGAAAAT | ||||
| X54796.1 | IL-1B(interleukin 1 beta) | 137 | forward | CAGCCGTGCAGTCAGTAAAA | [ref. 2] |
| reverse | GAAGCTCATGCAGAACACCA | ||||
| NM_001206735.1 | IL-1R1(interleukin 1 receptor, type I) | 124 | forward | GGGAAGGGTCCACCTGTAAC | [ref. 2] |
| reverse | ACAATGCTTTCCCCAACGTA | ||||
| NM_001046210.1 | IL-1R2(interleukin 1 receptor, type II) | 161 | forward | CGCCAGGCATACTCAGAAA | [ref. 36] |
| reverse | GAGAACGTGGCAGCTTCTTT | ||||
| NM_001308595.1 | IL-1RN(interleukin 1 receptor antagonist) | 145 | forward | AGGATCTGGGATGTCAACCA | [ref. 36] |
| reverse | CATGGATCCCCAGGAACATA |
2.5. Statistical Analysis
Statistical analysis was performed using the STATISTICA 10 software (Stat Soft. Inc., Tulsa, OK, USA). The results of gene and protein expression were analyzed using one-way analysis of variance (ANOVA) to identify the influence of anandamide on protein and gene expression during immune challenge and were followed by a post hoc Fisher’s least significance test. The results are presented as the mean ± standard error of the mean (S.E.M.); statistical significance was set at p < 0.05.
3. Results
3.1. Effect of AEA Injection on LPS-Induced Synthesis of IL-1β in the Hypothalamus of Endotoxin-Treated Ewes
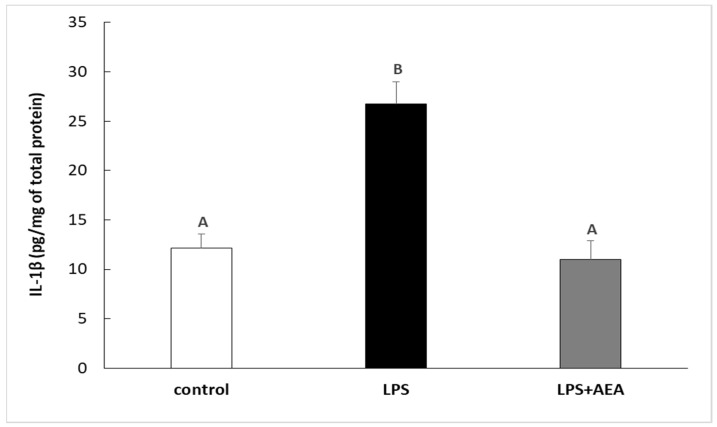
Injection of LPS increased (p < 0.05) the concentration of IL-1β protein in the hypothalamic tissue in comparison with the control group. Contrarily, the peripheral administration of AEA diminished this stimulatory effect of LPS on the expression of IL-1β protein in the ovine hypothalamus. The concentration of IL-1β in the group treated simultaneously with LPS and AEA was lower (p < 0.05) in comparison to the group treated only with LPS and did not differ from the IL-1β expression determined in control individuals (Figure 1).
3.2. Effect of AEA Injection on IL-1 Gene Expression System in the Hypothalamus of Endotoxin-Treated Ewes
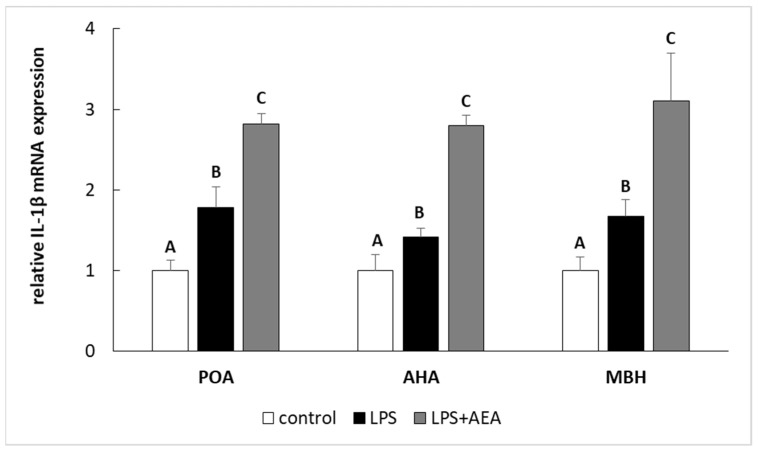
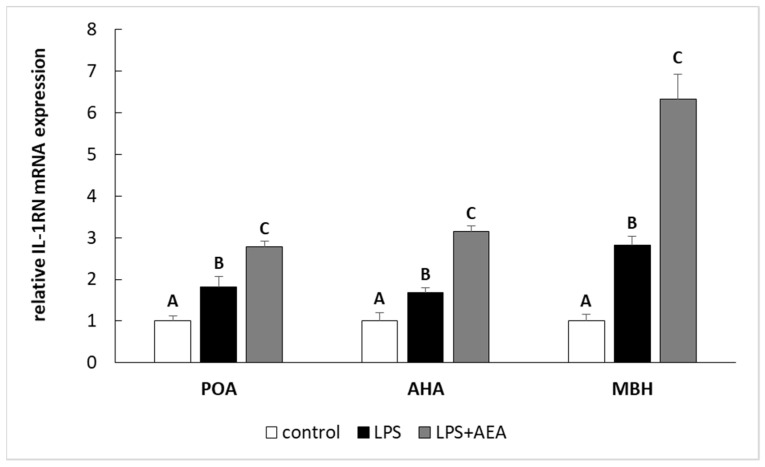
Inflammation induced by LPS treatment increased (p < 0.05) the relative level of IL-1β and IL-1RN gene expression in all studied structures such as the POA, AHA, and MBH. Furthermore, in the animals simultaneously treated with LPS and AEA, the gene expression of both IL-1β (Figure 2) and IL-1RN (Figure 3) in all analyzed hypothalamic structures was higher (p < 0.05) compared to control and LPS-treated groups.
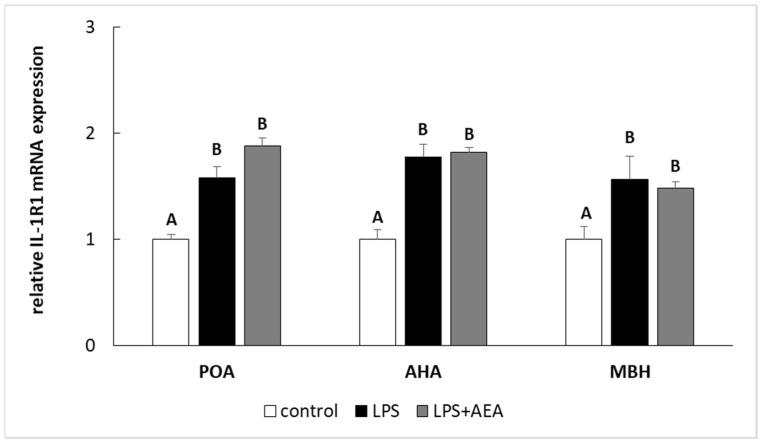
Endotoxin treatment elevated (p < 0.05) the level of IL-1R1 gene expression in all analyzed hypothalamic structures. On the other hand, the gene expression of IL-1R1 in these hypothalamic structures in ewe simultaneously treated with LPS and AEA did not differ in comparison to this gene expression in the endotoxin-treated group; however, it was higher (p < 0.05) when compared to the control animals (Figure 4).
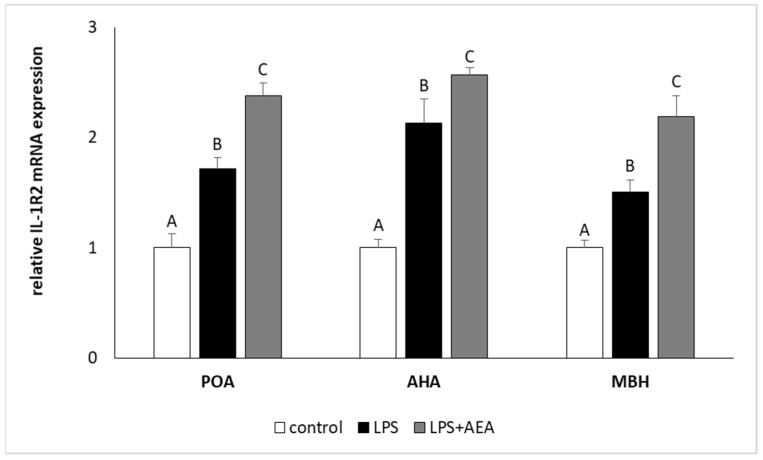
Injection of LPS elevated (p < 0.05) the level of IL-1R2 gene expression in all studied hypothalamic structures. However, in contrast to IL-1R1 gene expression, the simultaneous administration of LPS and AEA increased (p < 0.05) IL-1R2 gene expression in the POA, AHA, and MBH in comparison to both control and LPS-treated groups (Figure 5).
4. Discussion
We have shown that LPS-induced inflammation stimulated the synthesis of IL-1β in the hypothalamus both at mRNA and protein levels. These results were not surprising and confirmed the conclusions achieved in other studies that showed that systemic inflammation increases the expression of IL-1β in the hypothalamus of many species including rat [ref. 37], guinea pig [ref. 38], and sheep [ref. 2,ref. 31,ref. 39,ref. 40]. It is worth mentioning that at least a partial increase in the expression of hypothalamic IL-1β could be induced by stress provoked by LPS injection because it was proved that stress condition also stimulates the synthesis of IL-1β in the hypothalamus [ref. 19]. It was also shown that LPS-induced inflammation increased IL-1RN gene expression in the hypothalamus. IL-1RN is a natural anti-inflammatory factor. One of its major features is high affinity binding to the IL-1R1 with no conformational changes. As a result, IL-1RN blocks the IL-1b/IL-1R1 binding. Thus, IL-1β is unable to interact with corresponding receptor, and the intracellular signal that promotes further immune stimulation does not arise [ref. 41]. Moreover, IL-1RN is a terminal anti-inflammatory factor that can not only competitively antagonize the biological effects of IL-1, but also inhibit the effects of other inflammatory factors [ref. 42]. Furthermore, it was reported that the synthesis of IL-1RN rises during various pathological states characterized by local or systemic inflammation [ref. 43]. Therefore, the increased IL-1RN synthesis in the hypothalamus during peripheral inflammation suggests that three hours after LPS, not only do inflammatory processes occur, but also anti-inflammatory mechanisms are activated in order to protect the central tissues against unrestrained development of the inflammatory response.
Endotoxin-induced inflammation also increased the gene expression of IL-1 receptor type 1 and 2 in all analyzed hypothalamic structures. This observation is consistent with the results of previous studies in which peripheral inflammation is thought to stimulate the gene expression of IL-1R1 and IL-1R2 in the brain [ref. 31,ref. 39,ref. 40,ref. 44]. IL-1R1 plays an important role in initiating inflammatory response. It is a signal-transmitting receptor, triggered by both IL-1α and IL-1β ligands. The IL-1R1, with its intracellular domain, is responsible for initializing the inflammatory signaling processes in target cells [ref. 45]. The activation of IL-1R1 through its agonist is required to elicit intracellular signal and further development of immune response [ref. 46]. On the other hand, IL-1R2 is a membrane-bound protein expressed on the surface of various cells such as endometrial epithelial cells, basophils, neutrophils, monocytes, and activated T and B cells but it was also found in the brain tissue. IL-1R2 inhibits pro-inflammatory IL-1 effect by acting as a decoy receptor and by preventing IL-1/IL-1R1 binding [ref. 43].
The suppressive action of AEA on the synthesis of IL-1β, reported in our study, is generally consistent with other studies that showed that endocannabinoids and cannabinoids exhibit anti-inflammatory properties. It was found that AEA treatment reduced the serum level of pro-inflammatory IL-1β in rats exposed to water immersion and restrain stress [ref. 47]. Anti-inflammatory effects of cannabinoids were also observed in the brain tissue in neurodegenerative diseases occurring with accompanied neuroinflammation. A study performed on neurodegenerative disease murine models indicated that cannabidiol decreased the level of pro-inflammatory cytokines, including IL-1β, in the central nervous system [ref. 48]. In other viral study on mice administration of R(+)WIN55,212, CB1 agonist significantly reduced the expression of mRNA encoding pro-inflammatory cytokines such as IL-1β, IL-6, and tumor necrosis factor α [ref. 49]. Moreover, in vitro studies on rat microglial cells showed that cannabinoids inhibited LPS-stimulated gene expression of pro-inflammatory cytokines including IL-1β [ref. 50]. Although many studies suggest that there is a link between cannabinoids and TLR signaling pathways that leads to the synthesis of pro-inflammatory cytokines including IL-1β [ref. 51], the definition of direct molecular mechanism still awaits further study. On the other hand, the effect of cannabinoids on the IL-1β synthesis may be more elusive. It has been reported that AEA stimulated secretion of IL-1β from RAW264.7 cells [ref. 52]. Moreover, the administration of the psychoactive component of cannabis, delta 9-tetrahydrocannabinol (THC), activated cerebellar microglia and increased the expression of neuroinflammatory markers, including IL-1β [ref. 53]. The same pro-inflammatory effect of THC was reported in studies on murine resident peritoneal macrophage cultures, showing that cannabinoid treatment increased, in a dose-dependent manner, the secretion of IL-1β [ref. 54]. The different effect of AEA on the synthesis of IL-1 at the level of mRNA and protein expression demonstrated in our research indicates that endocannabinoids may influence cytokine expression through different cellular pathways. The findings that changes in IL-1β mRNA expression did not correspond with this cytokine protein synthesis suggests that inhibitory effect of AEA is targeted on a post-transcriptional level of IL-1β synthesis. It is known that after transcription, the inactive IL-1β precursor accumulates in the cytosol until it is processed by the activation of nucleotide-binding domain and leucine-rich repeat pyrin-containing protein-3 (NLRP3) and caspase-1 into an active cytokine [ref. 55,ref. 56]. Therefore, inhibition of NLRP3 inflammasome results in the reduction of the secretion of mature IL-1β. It was previously found that AEA and its COX-2 metabolite-prostaglandin E2 ethanolamide may exert an inhibitory effect on the NLRP3 inflammasome formation and activation [ref. 57]. Moreover, it seems that the inhibitory effect of anandamide on NLRP3 inflammasome activation could be common feature of cannabinoids because recent studies on LPS-nigericin-stimulated THP-1 monocytes also showed that cannabidiol suppressed NLRP3 inflammasome activation [ref. 58]. However, it is also possible that AEA influences the expression of mature IL-1β in the hypothalamic tissue by stimulating processes leading to degradation of this cytokine. It was found that activated matrix metalloproteinase (MMP)-1, MMP-2, MMP-3, and MMP-9 are responsible for degradation of IL-1β [ref. 59]. In turn, it was reported that AEA, acting via CB1 and transient receptor potential vanilloid-1, induced production of MMP-2 [ref. 60].
Our experiment also showed that animals co-treated with LPS and AEA are characterized by higher gene expression of IL-1RN and IL-1R2 but not IL-1R1 in all hypothalamic structures involved in GnRH-ergic activity in comparison to control and LPS-treated animals. The findings that in the group treated with LPS and AEA, the gene expression of IL-1RN was significantly raised compared to LPS-treated and control groups suggests that the anti-inflammatory effect of AEA in the hypothalamus may result from the stimulation of IL-1RN synthesis, which in turn competes with IL-1β for binding to its receptor. This mechanism of anti-inflammatory action of cannabinoids was stated in previous in vitro studies that showed potent anti-inflammatory effect of cannabinoids in murine glial and neuronal cell cultures. It was shown that culture of murine glial and neuronal cell incubated with cannabinoid agonists during inflammatory stimuli resulted in increased concentration of IL-1RN in the analyzed supernatants. Moreover, the authors postulated that induction of endogenous IL-1RN is essential for the neuro-protective effects of CBs [ref. 61]. The fact that AEA treatment did not influence IL-1R1 gene expression in the hypothalamus suggests that potential anti-inflammatory effect of AEA does not result from the reduced sensitivity of brain tissue to IL-1 action. However, the obtained results suggest the existence of another, novel mechanism of anti-inflammatory action of endocannabinoids in the central tissue on the basis of the increased synthesis of IL-1R2. As was mentioned above, acting as a decoy receptor IL-1R2 prevents IL-1/IL-1R1 binding, reducing the action of IL-1β. Additionally, IL-1R2 exists in both a membrane-bound and soluble form (sIL-1R2) with biological properties alike to both a decoy receptor and a binding protein [ref. 62]. Therefore, some amount of IL-1R2 from the hypothalamus may be released into the CSF, which in turn may reduce the amount of free, centrally acting IL-1β.
5. Conclusions
Summarizing, our study showed that AEA interfered with the synthesis of interleukin-1β and IL-1 system gene expressions in the hypothalamic structures involved in GnRH-ergic activity during an immune/inflammatory challenge. It was found that AEA inhibited the LPS- stimulated synthesis of central IL-1β in the hypothalamus, likely affecting posttranscriptional levels of this cytokine synthesis. However, anti-inflammatory effect of anandamide at the level of central nervous system might also result from its stimulating action on IL-1RN and IL-1R2 gene expression. However, further detailed research is required to investigate this issue. In sum, our study suggests the potential of endocannabinoids and/or their metabolites in the inhibition of inflammatory process at the level of central nervous system, and therefore their usefulness in the therapy of inflammatory-induced neuroendocrine disorders.
References
- D. Tomaszewska-Zaremba, A. Herman, K. Haziak. How does bacterial endotoxin influence gonadoliberin/gonadotropins secretion and action?. J. Anim. Feed Sci., 2016. [DOI]
- A.P. Herman, A. Krawczyńska, J. Bochenek, H. Antushevich, A. Herman, D. Tomaszewska-Zaremba. Peripheral injection of SB203580 inhibits the inflammatory-dependent synthesis of proinflammatory cytokines in the hypothalamus. BioMed Res. Int., 2014. [DOI | PubMed]
- R.H. McCusker, K.W. Kelley. Immune–neural connections: How the immune system’s response to infectious agents influences behavior. J. Exp. Biol., 2013. [DOI | PubMed]
- A.P. Herman, K. Kopycińska, A. Krawczyńska, K. Romanowicz, D. Tomaszewska-Zaremba. The effect of repeated endo-toxin injections on gonadotropin secretion in ewes. J. Anim. Feed Sci., 2014. [DOI]
- J. Daniel, M. Abrams, L. DeSouza, C. Wagner, B. Whitlock, J. Sartin. Endotoxin inhibition of luteinizing hormone in sheep. Domest. Anim. Endocrinol., 2003. [DOI | PubMed]
- A.P. Herman, J. Skipor, A. Krawczyńska, J. Bochenek, K. Wojtulewicz, H. Antushevich, A. Herman, K. Paczesna, K. Romanowicz, D. Tomaszewska-Zaremba. Peripheral inhibitor of AChE, neostigmine, prevents the inflammatory dependent suppression of GnRH/LH secretion during the follicular phase of the estrous cycle. BioMed Res. Int., 2017. [DOI | PubMed]
- A.P. Herman, J. Krawczyńska, J. Bochenek, K. Haziak, K. Romanowicz, T. Misztal, H. Antushevich, A. Herman, D. To-maszewska-Zaremba. The effect of rivastigmine on the LPS-induced suppression of GnRH/LH secretion during the follicu-lar phase of the estrous cycle in ewes. Anim. Reprod. Sci., 2013. [DOI | PubMed]
- F.J. Karsch, D.F. Battaglia, K.M. Breen, N. Debus, T.G. Harris. Mechanisms for ovarian cycle disruption by immune/inflammatory stress. Stress, 2002. [DOI | PubMed]
- D.F. Battaglia, J.M. Bowen, H.B. Krasa, L.A. Thrun, C. Viguié, F.J. Karsch. Endotoxin inhibits the reproductive neuroen-docrine axis while stimulating adrenal steroids: A simultaneous view from hypophyseal portal and peripheral blood. Endocrinology, 1997. [DOI | PubMed]
- M. Caldani, M. Batailler, M.P. Dubois. LHRH-immunoreactive structures in the sheep brain. Histochemistry, 1988. [DOI | PubMed]
- A.P. Herman, T. Misztal, K. Romanowicz, D. Tomaszewska-Zaremba. Central Injection of Exogenous IL-1β in the Control Activities of Hypothalamic-Pituitary-Gonadal Axis in Anestrous Ewes. Reprod. Domest. Anim., 2011. [DOI | PubMed]
- S.S. Kang, S.R. Kim, S. Leonhardt, H. Jarry, W. Wuttke, K. Kim. Effect of interleukin-1beta on gonadotropin-releasing hormone (GnRH) and GnRH receptor gene expression in castrated male rats. J. Neuroendocrinol., 2000. [DOI | PubMed]
- N.J. Rothwell, G.N. Luheshi. Interleukin 1 in the brain: Biology, pathology and therapeutic target. Trends Neurosci., 2000. [DOI | PubMed]
- A.P. Herman, D. Tomaszewska-Zaremba. Effect of endotoxin on the expression of GnRH and GnRHR genes in the hypothalamus and anterior pituitary gland of anestrous ewes. Anim. Reprod. Sci., 2010. [DOI | PubMed]
- W.A. Banks, A.J. Kastin, R.D. Broadwell. Passage of Cytokines across the Blood-Brain Barrier. Neuroimmunomodulation, 1995. [DOI | PubMed]
- M. Kowalewska, A. Herman, A. Szczepkowska, J. Skipor. The effect of melatonin from slow-release implants on basic and TLR-4-mediated gene expression of inflammatory cytokines and their receptors in the choroid plexus in ewes. Res. Vet. Sci., 2017. [DOI | PubMed]
- M. Kowalewska, A. Szczepkowska, A. Herman, M. Pellicer-Rubio, M. Jałyński, J. Skipor. Melatonin from slow-release implants did not influence the gene expression of the lipopolysaccharide receptor complex in the choroid plexus of seasonally anoestrous adult ewes subjected or not to a systemic inflammatory stimulus. Small Rumin. Res., 2017. [DOI]
- J. Skipor, M. Kowalewska, A. Szczepkowska, A. Majewska, T. Misztal, M. Jalynski, A.P. Herman, K. Zabek. Plasma and cerebrospinal fluid interleukin-1β during lipopolysaccharide-induced systemic inflammation in ewes implanted or not with slow-release melatonin. J. Anim. Sci. Biotechnol., 2017. [DOI | PubMed]
- D.F. Barnard, K.M. Gabella, A.C. Kulp, A.D. Parker, P.B. Dugan, J.D. Johnson. Sex differences in the regulation of brain IL-1β in response to chronic stress. Psychoneuroendocrinology, 2019. [DOI | PubMed]
- T.G. Li, R. Pu, L. Shui, S.M. Bai, J. Lu, Y.S. Chen, Y. Tu. Effect of moxibustion on inflammatory cytokine levels in hippo-campus and hypothalamus in rats with fatigue. Zhen Ci Yan Jiu, 2019. [PubMed]
- D. Tomaszewska-Zaremba, A.P. Herman, T. Misztal. Does central IL-1b affect GnRH secretion in the hypothalamus of an-oestrous ewes via different regulatory pathways?. J. Anim. Feed Sci., 2013. [DOI]
- P. Igaz, R. Salvi, J.-P. Rey, M. Glauser, F.P. Pralong, R.C. Gaillard. Effects of Cytokines on Gonadotropin-Releasing Hormone (GnRH) Gene Expression in Primary Hypothalamic Neurons and in GnRH Neurons Immortalized Conditionally. Endocrinology, 2006. [DOI | PubMed]
- R. Tanasescu, C.S. Constantinescu. Cannabinoids and the immune system: An overview. Immunobiology, 2010. [DOI | PubMed]
- M. Grill, C. Högenauer, A. Blesl, J. Haybaeck, N. Golob-Schwarzl, N. Ferreirós, D. Thomas, R. Gurke, M. Trötzmüller, H.C. Köfeler. Members of the endocannabinoid system are distinctly regulated in inflammatory bowel disease and colorectal cancer. Sci. Rep., 2019. [DOI | PubMed]
- S.L. Cruz, E. Sánchez-Miranda, J.I. Castillo-Arellano, R.D. Cervantes-Villagrana, A. Ibarra-Sánchez, C. González-Espinosa. Anandamide inhibits FcεRI-dependent degranulation and cytokine synthesis in mast cells through CB2 and GPR55 receptor activation. Possible involvement of CB2-GPR55 heteromers. Int. Immunopharmacol., 2018. [DOI | PubMed]
- D. Centonze, A. Finazzi-Agro, G. Bernardi, M. Maccarrone. The endocannabinoid system in targeting inflammatory neuro-degenerative diseases. Trends Pharmacol. Sci., 2007. [DOI | PubMed]
- K.J. Spiller, G.H. Bi, Y. He, E. Galaj, E.L. Gardner, Z.X. Xi. Cannabinoid CB1 and CB2 receptor mechanisms underlie can-nabis reward and aversion in rats. Br. J. Pharmacol., 2019. [DOI | PubMed]
- M. Maccarrone, T. Wenger. Effects of Cannabinoids on Hypothalamic and Reproductive Function. Handbook of Experimental Pharmacology, 2005
- P. Nagarkatti, R. Pandey, S.A. Rieder, V.L. Hegde, M. Nagarkatti. Cannabinoids as novel anti-inflammatory drugs. Future Med. Chem., 2009. [DOI | PubMed]
- R.R. Noroś. Nutrient Requirements for Cattle and Sheep in the Traditional System, 1993
- A.P. Herman, D. Tomaszewska-Zaremba, M. Kowalewska, A. Szczepkowska, M. Oleszkiewicz, A. Krawczyńska, M. Wójcik, H. Antushevich, J. Skipor. Neostigmine attenuates proinflammatory cytokine expression in preoptic area but not choroid plexus during lipopolysaccharide-induced systemic inflammation. Med. Inflamm., 2018. [DOI | PubMed]
- A. Krawczyńska, H. Antushevich, J. Bochenek, K. Wojtulewicz, B. Pawlina, A.P. Herman, D.A. Zięba. Photoperiodic conditions as a factor modulating leptin influence on pro-inflammatory cytokines and their receptors gene expression in ewe’s aorta. J. Anim. Feed Sci., 2019. [DOI]
- Z. Justinova, M. Solinas, G. Tanda, G.H. Redhi, S.R. Goldberg. The endogenous cannabinoid anandamide and its synthetic analog R(+)-methanandamide are intravenously self-administered by squirrel monkeys. J. Neurosci., 2005. [DOI | PubMed]
- K. Wojtulewicz, D. Tomaszewska-Zaremba, A. Krawczyńska, M. Tomczyk, A.P. Herman. The effect of inflammation on the synthesis of luteinizing hormone and gonadotropin-releasing hormone receptor expression in the pars tuberalis of ewe during different photoperiodic conditions. Can. J. Anim. Sci., 2018. [DOI]
- R. Rasmussen. Quantification on the LightCycler. Rapid Cycle Real-Time PCR, 2001
- A.P. Herman, J. Bochenek, K. Król, A. Krawczyńska, H. Antushevich, B. Pawlina, A. Herman, K. Romanowicz, D. To-maszewska-Zaremba. Central interleukin-1β suppresses the nocturnal secretion of melatonin. Med. Inflamm., 2016. [DOI | PubMed]
- F. Wahab, N.N. Santos-Junior, R.P.D.A. Rodrigues, L.H.A. Costa, C.H.R. Catalão, M.J.A. Rocha. Interleukin-1 Receptor Antagonist Decreases Hypothalamic Oxidative Stress During Experimental Sepsis. Mol. Neurobiol., 2016. [DOI | PubMed]
- L. Janský, S. Vybíral, D. Pospíšilová, J. Roth, J. Dornand, E. Zeisberger, J. Kamínková. Production of systemic and hypo-thalamic cytokines during the early phase of endotoxin fever. Neuroendocrinology, 1995. [DOI | PubMed]
- A.P. Herman, A. Krawczyńska, J. Bochenek, K. Haziak, H. Antushevitch, A. Herman, D. Tomaszewska-Zaremba. Inhibi-tion of acetylcholinesterase activity by rivastigmine decreases lipopolysaccharide-induced IL-1β expression in the hypothala-mus of ewes. Domest. Anim. Endocrinol., 2013. [DOI | PubMed]
- A. Herman, T. Misztal, D.T. Zaremba. Expression of Interleukin (IL)-1β and IL-1 Receptors Genes in the Hypothalamus of Anoestrous Ewes after Lipopolysaccharide Treatment. Reprod. Domest. Anim., 2010. [DOI | PubMed]
- E. Pinteaux, N.J. Rothwell, H. Boutin. Neuroprotective actions of endogenous interleukin-1 receptor antagonist (IL-1ra) are mediated by glia. Glia, 2006. [DOI | PubMed]
- C. Jin, W.-L. Fu, D.-D. Zhang, W.-W. Xing, W.-R. Xia, Z. Wei, M.-J. Zou, X.-M. Zhu, D.-G. Xu. The protective role of IL-1Ra on intestinal ischemia reperfusion injury by anti-oxidative stress via Nrf2/HO-1 pathway in rat. Biomed. J., 2019. [DOI | PubMed]
- J. Palomo, D. Dietrich, P. Martin, G. Palmer, C. Gabay. The interleukin (IL)-1 cytokine family–Balance between agonists and antagonists in inflammatory diseases. Cytokine, 2015. [DOI | PubMed]
- R. André, D. Lerouet, I. Kimber, E. Pinteaux, N.J. Rothwell. Regulation of expression of the novel IL-1 receptor family members in the mouse brain. J. Neurochem., 2005. [DOI | PubMed]
- A. Bebes, F. Kovács-Sólyom, J. Prihoda, R. Kui, L. Kemény, R. Gyulai. Interleukin-1 Receptors Are Differentially Expressed in Normal and Psoriatic T Cells. Mediat. Inflamm., 2014. [DOI]
- D. Boraschi, A. Tagliabue. The interleukin-1 receptor family. Semin. Immunol., 2013. [DOI | PubMed]
- Z. Warzecha, A. Dembinski, P. Ceranowicz, M. Dembinski, J. Cieszkowski, P. Kownacki, P.C. Konturek. Role of sensory nerves in gastroprotective effect of anandamide in rats. J. Physiol. Pharmacol., 2011. [PubMed]
- M. Mecha, A. Feliu, P.M. Inigo, L. Mestre, F. Carrillo-Salinas, C. Guaza. Cannabidiol provides long-lasting protection against the deleterious effects of inflammation in a viral model of multiple sclerosis: A role for A2A receptors. Neurobiol. Dis., 2013. [DOI | PubMed]
- J.L. Croxford, S.D. Miller. Immunoregulation of a viral model of multiple sclerosis using the synthetic cannabinoid R(+)WIN55,212. J. Clin. Investig., 2003. [DOI | PubMed]
- R.A. Puffenbarger, A.C. Boothe, G.A. Cabral. Cannabinoids inhibit LPS-inducible cytokine mRNA expression in rat micro-glial cells. Glia, 2000. [DOI | PubMed]
- K.L. McCoy. Interaction between Cannabinoid System and Toll-Like Receptors Controls Inflammation. Mediat. Inflamm., 2016. [DOI]
- T. Jourdan, G. Godlewski, R. Cinar, A. Bertola, G. Szanda, J. Liu, J. Tarn, T. Han, B. Mukhopadhyay, M.C. Skarulis. Activation of the Nlrp3 inflammasome in infiltrating macrophages by endocan-nabinoids mediates beta cell loss in type 2 diabetes. Nat. Med., 2013. [DOI | PubMed]
- L. Cutando, A. Busquets-Garcia, E. Puighermanal, M. Gomis-González, J.M. Delgado-García, A. Gruart, R. Maldonado, A. Ozaita. Microglial activation underlies cerebellar deficits produced by repeated cannabis exposure. J. Clin. Investig., 2013. [DOI | PubMed]
- W. Zhu, C. Newton, Y. Daaka, H. Friedman, T.W. Klein. Delta 9-Tetrahydrocannabinol enhances the secretion of interleukin 1 from endotoxin-stimulated macrophages. J. Pharmacol. Exp. Ther., 1994. [PubMed]
- C.A. Dinarello. Overview of the IL-1 family in innate inflammation and acquired immunity. Immunol. Rev., 2018. [DOI | PubMed]
- H. Antushevich. Interplays between inflammasomes and viruses, bacteria (pathogenic and probiotic), yeasts and parasites. Immunol. Lett., 2020. [DOI | PubMed]
- G. Li, M. Xia, J.M. Abais, K. Boini, P.-L. Li, J.K. Ritter. Protective Action of Anandamide and Its COX-2 Metabolite against l-Homocysteine-Induced NLRP3 Inflammasome Activation and Injury in Podocytes. J. Pharmacol. Exp. Ther., 2016. [DOI | PubMed]
- C. Liu, H. Ma, A.L. Slitt, N.P. Seeram. Inhibitory Effect of Cannabidiol on the Activation of NLRP3 Inflammasome Is Associated with Its Modulation of the P2X7 Receptor in Human Monocytes. J. Nat. Prod., 2020. [DOI | PubMed]
- A. Ito, A. Mukaiyama, Y. Itoh, H. Nagase, I.B. Thøgersen, J.J. Enghild, Y. Sasaguri, Y. Mori. Degradation of Interleukin 1β by Matrix Metalloproteinases. J. Biol. Chem., 1996. [DOI | PubMed]
- K. Miyashita, T. Oyama, T. Sakuta, M. Tokuda, M. Torii. Anandamide Induces Matrix Metalloproteinase-2 Production through Cannabinoid-1 Receptor and Transient Receptor Potential Vanilloid-1 in Human Dental Pulp Cells in Culture. J. Endod., 2012. [DOI | PubMed]
- F. Molina-Holgado, E. Pinteaux, J.D. Moore, E. Molina-Holgado, C. Guaza, R.M. Gibson, N.J. Rothwell. Endogenous Interleukin-1 Receptor Antagonist Mediates Anti-Inflammatory and Neuroprotective Actions of Cannabinoids in Neurons and Glia. J. Neurosci., 2003. [DOI | PubMed]
- V.A. Peters, J.J. Joesting, G.G. Freund. IL-1 receptor 2 (IL-1R2) and its role in immune regulation. Brain Behav. Immun., 2013. [DOI | PubMed]