Clinical implication and immunological landscape analyses of ANLN in pan‐cancer: A new target for cancer research
Abstract
Background:
Anillin is a F‐actin binding protein (ANLN) mainly involved in the process of cytokinesis and known to be dysregulated in diverse cancers. However, the role of ANLN in pan‐cancer prognosis and tumor immunity remains unclear.
Methods:
Gene expression profiles of 31 solid tumors were downloaded from The Cancer Genome Atlas (TCGA) database. ANLN mRNA and protein expression were quantified using quantitative real‐time PCR (qRT‐PCR) and immunohistochemistry (IHC). Protein expression of ANLN was further confirmed in Human Protein Atlas (HPA) database. Cox regression and Kaplan–Meier analysis were utilized to assess the prognostic value of ANLN in pan‐cancer. The correlation between ANLN and different immune gene markers and infiltration cells was analyzed via ESTIMATE and CIBERSORT. A BLCA immunotherapy cohort: IMvigor (210) was used to confirm the role of ANLN in immune response.
Results:
ANLN upregulation was detected in 21 types of cancers and was associated with poor overall survival (OS), disease‐free interval (DFI), and progression‐free interval (PFI) in most cancers except in THYM (Thymoma). Additionally, correlation analysis revealed a significantly positive association between ANLN expression and tumor mutation burden (TMB), microsatellite instability (MSI), immune cells infiltration. and immune checkpoint genes in various cancers. The BLCA immunotherapy cohort confirmed that patients with higher ANLN level had better immune responses and longer OS.
Conclusion:
ANLN may serve as a prognostic biomarker for pan‐cancer. ANLN upregulation is associated with higher TMB, MSI, and immune cell infiltration in multiple types of tumors, shedding new light for cancer treatment.
Article type: Research Article
Keywords: ANLN, immune infiltration, pan‐cancer, prognosis, TMB
Affiliations: Department of Radiation Oncology Shanghai Tenth People’s Hospital of Tongji University Shanghai China; Department of Urology The Second Affiliated Hospital of Nanjing Medical University Nanjing China; Department of Radiotherapy & Oncology The Affiliated Suzhou Hospital of Nanjing Medical University Suzhou China; Department of Pathology The Second Affiliated Hospital of Nanjing Medical University Nanjing China; Department of Oncology The Affiliated Hospital of Jiangnan University Wuxi China; Department of Pathology The Affiliated Suzhou Hospital of Nanjing Medical University Suzhou China; Department of Radiation Oncology Dushu Lake Hospital Affiliated to Soochow University (Medical Center of Soochow University) Suzhou China; State Key Laboratory of Radiation Medicine and Protection Soochow University Suzhou China; Department of Oncology First Affiliated Hospital of Nanjing Medical University Nanjing China
License: © 2022 The Authors. Cancer Medicine published by John Wiley & Sons Ltd. CC BY 4.0 This is an open access article under the terms of the http://creativecommons.org/licenses/by/4.0/ License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Article links: DOI: 10.1002/cam4.5177 | PubMed: 36030492 | PMC: PMC9972146
Relevance: Moderate: mentioned 3+ times in text
Full text: PDF (15.9 MB)
INTRODUCTION
Anillin (ANLN) is a F‐actin binding protein originally isolated from Drosophila embryo.ref. cam45177-bib-0001 It is highly conserved and mainly involved in the cortical cytoskeletal dynamics during cytokinesis and cellularization.ref. cam45177-bib-0002 Dysregulations of ANLN is observed in a wide range of malignant tumors and its overexpression is closely linked to unfavorable outcomes in colorectal cancer,ref. cam45177-bib-0003 breast cancer,ref. cam45177-bib-0004 and liver cancer.ref. cam45177-bib-0005 However, the role of ANLN in pan‐cancer remains still unclear.
The tumor microenvironment (TME) consists of not only cancer cells, but also tumor‐associated normal cells, including infiltrating stromal and immune cells.ref. cam45177-bib-0006 Increasing evidence suggests that TME has multiple effects on tumorigenesis, development, and prognosis.ref. cam45177-bib-0007, ref. cam45177-bib-0008, ref. cam45177-bib-0009, ref. cam45177-bib-0010, ref. cam45177-bib-0011 However, the possible mechanisms of stromal and immune components remain elusive. Immunotherapy has proved to be a powerful clinical strategy for cancer treatment.ref. cam45177-bib-0012 Monoclonal antibodies targeting cytotoxic T lymphocytes‐associated antigen 4 (CTLA4), programed death‐1 (PD‐1) receptor, and its ligand (PD‐L1) have achieved clinical success in some cancers.ref. cam45177-bib-0013, ref. cam45177-bib-0014, ref. cam45177-bib-0015 However, only a subset of patients could benefit from current immunotherapies.ref. cam45177-bib-0016 Thus, the discovery of effective immunotherapy targets is of great clinical significance.
Hence, our study comprehensively explored the prognostic significance of ANLN in 31 tumors based on The Cancer Genome Atlas (TCGA) data. Subsequently, we explored the potential correlations between ANLN and tumor immune characteristics.
METHODS
Data collection and preprocessing
Gene expression profiles of 31 types of solid tumors including raw counts and Fragments Per Kilobase of transcript per Million (FPKM)‐normalized RNA‐seq data were downloaded from TCGA database (https://portal.gdc.cancer.gov/) on 20 April, 2021 (ACC: adrenocortical carcinoma, BLCA: bladder urothelial carcinoma, BRCA: breast invasive carcinoma, CESC: cervical squamous cell carcinoma, CHOL: cholangiocarcinoma, COAD: colon adenocarcinoma, DLBC: lymphoid neoplasm diffuse large B‐cell lymphoma, ESCA: esophageal carcinoma, GBM: glioblastoma multiforme, LGG: brain lower grade glioma, HNSC: head and neck squamous cell carcinoma, KICH: kidney chromophobe, KIRC: kidney renal clear cell carcinoma, KIRP: kidney renal papillary cell carcinoma, LIHC: liver hepatocellular carcinoma, LUAD: lung adenocarcinoma, LUSC: lung squamous cell carcinoma, MESO: mesothelioma, OV: ovarian serous cystadenocarcinoma, PAAD: pancreatic adenocarcinoma, PCPG: pheochromocytoma and paraganglioma, PRAD: prostate adenocarcinoma, READ: rectum adenocarcinoma, SARC: sarcoma, SKCM: skin cutaneous melanoma, STAD: stomach adenocarcinoma, TGCT: testicular germ cell tumors, THCA: thyroid carcinoma, THYM: thymoma, UCEC: uterine corpus endometrial carcinoma, UCS: uterine carcinosarcoma, UVM: uveal melanoma). Acute myeloid leukemia (LAML) and large B‐cell lymphoma (DLBC) were excluded because they were hematological tumors. The primary data are presented in Table 1. Protein‐encoding genes were aligned to the human reference genome assembly GRCh38. Transcriptome files of 18,321 related genes were normalized with the assistance from R packages “limma” on R platform v3.6.3. and converged into a matrix for further analysis. The difference in ANLN expression was compared between normal tissues and tumor tissues using the Wilcoxon test with a threshold of p < 0.05. ANLN protein expression data in 16 types of cancers were retrieved via the Human Protein Atlas (HPA) database (https://www.proteinatlas.org/).
TABLE 1: Pan‐cancer data acquired from TCGA
| Cancer type | Full name | Tumor samples | Normal samples |
|---|---|---|---|
| ACC | Adrenocortical carcinoma | 79 | 0 |
| BLCA | Bladder urothelial carcinoma | 414 | 19 |
| BRCA | Breast invasive carcinoma | 1109 | 120 |
| CESC | Cervical squamous cell carcinoma and endocervical adenocarcinoma | 306 | 3 |
| CHOL | Cholangiocarcinoma | 36 | 9 |
| COAD | Colon adenocarcinoma | 480 | 41 |
| ESCA | Esophageal carcinoma | 162 | 11 |
| GBM | Glioblastoma multiforme | 169 | 5 |
| HNSC | Head and neck squamous cell carcinoma | 502 | 44 |
| KICH | Kidney chromophobe | 65 | 24 |
| KIRC | Kidney renal clear cell carcinoma | 539 | 72 |
| KIRP | Kidney renal papillary cell carcinoma | 289 | 32 |
| LGG | Brain lower grade glioma | 529 | 0 |
| LIHC | Liver hepatocellular carcinoma | 374 | 50 |
| LUAD | Lung adenocarcinoma | 535 | 59 |
| LUSC | Lung squamous cell carcinoma | 502 | 49 |
| MESO | Mesothelioma | 86 | 0 |
| OV | Ovarian serous cystadenocarcinoma | 379 | 0 |
| PAAD | Pancreatic adenocarcinoma | 178 | 4 |
| PCPG | Pheochromocytoma and paraganglioma | 183 | 3 |
| PRAD | Prostate adenocarcinoma | 499 | 52 |
| READ | Rectum adenocarcinoma | 167 | 10 |
| SARC | Sarcoma | 263 | 2 |
| SKCM | Skin cutaneous melanoma | 471 | 1 |
| STAD | Stomach adenocarcinoma | 375 | 32 |
| TGCT | Testicular germ cell tumors | 156 | 0 |
| THCA | Thyroid carcinoma | 510 | 58 |
| THYM | Thymoma | 119 | 2 |
| UCEC | Uterine corpus endometrial carcinoma | 552 | 35 |
| UCS | Uterine carcinosarcoma | 56 | 0 |
| UVM | Uveal melanoma | 80 | 0 |
Prognosis analysis
The prognostic values of ANLN, including Overall Survival (OS) and Disease‐Free Interval (DFI) and Progression‐Free Interval (PFI), was assessed via univariate Cox regression model. Samples were clustered into high or low expression cohorts based on the median value of ANLN. Kaplan–Meier (K‐M) curves and the forest plots were utilized to compare the survival time differences. All analyses were carried out using the “survival” and “survminer” package. After that, the associations between ANLN and clinical parameters, including age, sex, and the TNM stage, were assessed by the chi‐square test or Fisher’s exact test. All tests were two‐tailed and a p‐value of 0.05 or less was considered statistically significant.
Correlation between ANLN expression and TMB and MSI
Tumor mutational burden (TMB) is a measure of the amount of somatic coding mutations per DNA megabase.ref. cam45177-bib-0017 Microsatellite instability (MSI) is a molecular tumor phenotype of a deficient mismatch repair system.ref. cam45177-bib-0018 Analysis regarding the correlation between ANLN expression and TMB/MSI was conducted by Pearson correlation analysis and shown in the radar chart using the “fmsb” package.
Correlation between ANLN expression and tumor immunity
ESTIMATE (Estimation of Stromal and Immune cells in Malignant Tumor tissues using Expression data) is a method for estimating the presence of stromal/immune cells in tumor tissues and tumor purity with high accuracy.ref. cam45177-bib-0019 This method was applied to explore the association between the immune and stromal cell ratio with ANLN expression.ref. cam45177-bib-0019
We further explored the correlation between ANLN expression and 22 types of infiltrating immune cells using the “CIBERSORT” algorithm. The association between ANLN expression and common immune checkpoint genes was investigated using the “Reshape2” package and shown in heatmaps by the R “RColorBrewer” package. The Spearman method was utlized to assess the correlation coefficients.
Collection of an immunotherapy‐based cohort
After systematically searching for immunotherapy gene expression profiles, an immunotherapeutic cohort: advanced urothelial cancer with intervention of atezolizumab, an anti‐PD‐L1 antibody (IMvigor210 cohort) with complete clinical information was recruited in our study.ref. cam45177-bib-0020 The complete information of the cohort is available on http://research‐pub.Gene.com/imvigor210corebiologies. The raw count data were normalized by the DEseq2 R package and transformed into the TPM value for further study.
Gene set enrichment analysis
Next, our study explored the potential molecular functions and biological pathways regulated by ANLN. The Gene Set Enrichment Analysis (GSEA) software (http://www.broadinstitute.org/gsea) was used to investigate the enrichment of GO and KEGG pathways between high ANLN expression and low expression groups according to the median value of ANLN. All analyses were conducted with R package “org.Hs.eg.db,” “clusterProfiler,” “enrichplot,” “DOSE/limma.” Gene sets with |NES| > 1, NOM p < 0.05, and FDR q < 0.25 were considered as enrichment significant.
Experimental validation
Fresh tissue samples of 7 LUAD patients were obtained from the Affiliated Suzhou Hospital of Nanjing Medical University. The mRNA and protein expression of ANLN were quantified using quantitative real‐time PCR (qRT‐PCR) and immunohistochemistry (IHC) as previously described.ref. cam45177-bib-0010 Briefly, qRT‐PCR was conducted on a LightCycler 480 (Roche 480, Germany) real‐time thermal cycler, using SYBR Green dye. Each sample was set with three multiple wells. Assays detected with 5 Ct’s less than the negative control (No Template Control, NTC), and with Ct < 35 were included for further data analysis. The melting analysis was added finally to evaluate the specificity of PCR products. Expression levels of ANLN were calculated using 2−ΔΔCt method and normalized to that of GAPDH mRNA (forward: 5′‐GTCTCCTCTGACTTCAACAGCG −3′ and reverse: 5′‐ACCACCCTGTTGCTGTAGCCAA −3′). The sequences of the primers used were as follows: forward: 5′‐CAGACAGTTCCATCCAAGGGAG‐3′ and reverse: 5′‐CTTGACAACGCTCTCCAAAGCG‐3′. IHC staining for ANLN was accomplished by two professional pathologists. Formalin‐fixed human lung tumor tissue sections were sliced into serial sections with a thickness of 5 μm for IHC. An anti‐ANLN antibody (anti‐ANLN: catalog no., BS‐7738R; dilution, 1:200; Bioss Inc.) was used as the primary antibody. PBS was functioned a negative control to keep the specificity of staining. A 3,3′‐diaminobenzidine (DAB) and hematoxylin counterstain (ZSGB‐Bio, China) were used to visualize antibody staining. Finally, immunostained sections were photographed using a microscope (Olympus Corporation). This study was approved by the Ethics Committee of the hospital. Each participant had signed informed consent in accordance with the Declaration of Helsinki (Ethics Approval No. KL901198).
RESULTS
Pan‐cancer ANLN expression
Our study firstly analyzed the expression pattern of ANLN in tumor and normal samples from the TCGA pan‐cancer dataset. As shown in Figure 1A, ANLN expression was significantly increased in BLCA, BRCA, CESC, CHOL, COAD, ESCA, HNSC, KICH, KIRC, KIRP, LIHC, LUAD, LUSC, PAAD, PCPG, PRAD, READ, SARC, STAD, THCA, and UCEC. Next, we investigated ANLN protein expression based on the HPA database. ANLN expression was significantly higher in tumor samples than normal tissues in BLCA, CRC (including COAD and READ), LUAD, OV, and TGCT (Figure 1B). We further validated ANLN expression in LUAD patients. The immunohistochemical and mRNA results demonstrated that ANLN expression was significantly higher in tumor tissues than paracancerous samples (Figure 1C,D).
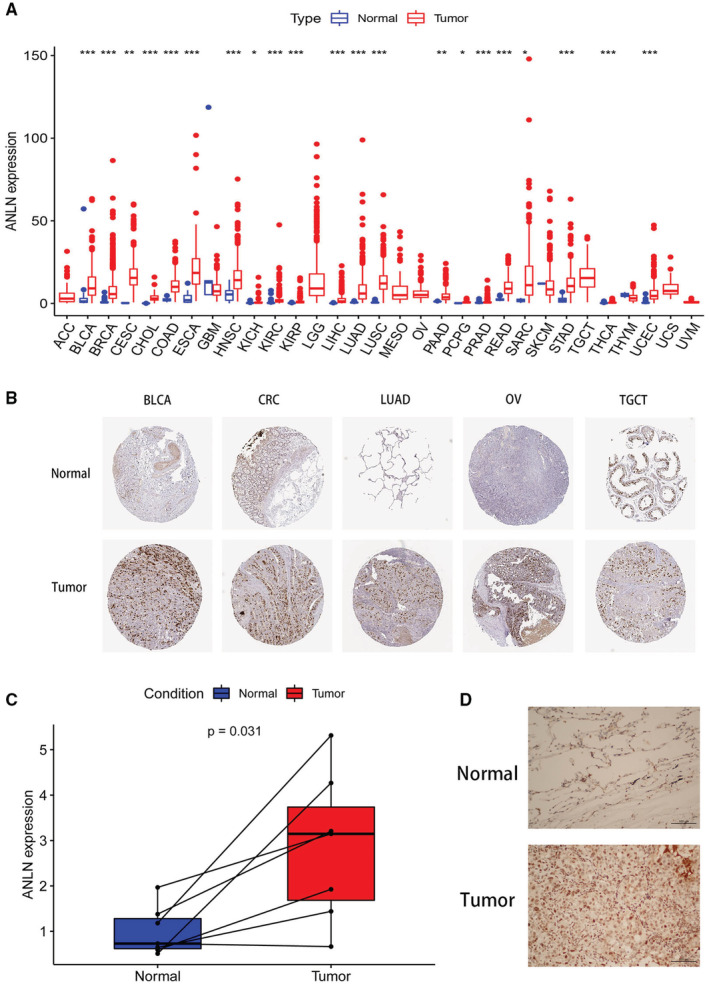
Survival analysis of ANLN
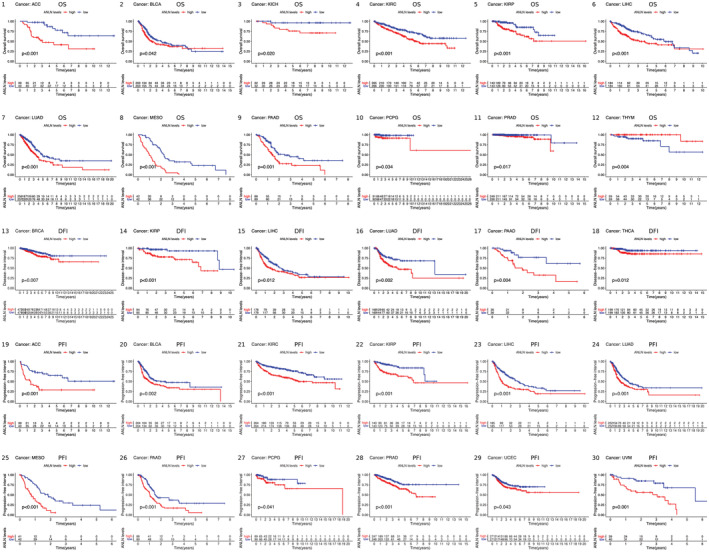
Our study investigated the correlation between ANLN expression and patients’ OS, DFI, PFI using the TCGA cohort. Kaplan–Meier curves showed that high expression of ANLN was significantly associated with poor OS in ACC, BLCA, KICH, KIRC, KIRP, LIHC, LUAD, MESO, PAAD, PCPG, and PRAD. Particularly, low expression of ANLN was linked to poor OS in THYM. In addition, high ANLN expression was correlated with poor DFI in BRCA, KIRP, LIHC, LUAD, PAAD, and THCA. Results of the Kaplan–Meier plot also suggested that increased level of ANLN was distinctly connected with worse PFI in patients with ACC, BLCA, KIRC, KIRP, LIHC, LUAD, MESO, PAAD, PCPG, PRAD, UCEC, and UVM (Figure 2).
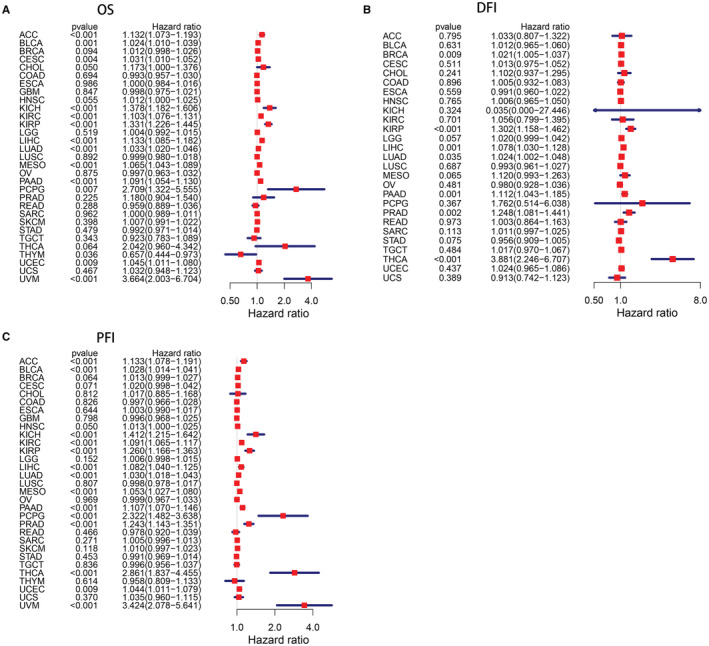
In Cox regression analysis, high ANLN expression was an unfavorable factor for patients’ OS in ACC, BLCA, CESC, CHOL, KICH, KIRC, KIRP, LIHC, LUAD, MESO, PAAD, PCPG, UCEC, and UVM. While patients with low ANLN expression showed shortened OS in THYM. For DFI, elevated level of ANLN was significantly correlated with shorter DFI in BRCA, KIRP, LIHC, LUAD, PAAD, PRAD, and THCA. Additionally, increased level of ANLN was strongly correlated with worse PFI in ACC, BLCA, HNSC, KICH, KIRC, KIRP, LIHC, LUAD, MESO, PAAD, PCPG, PRAD, THCA, UCEC, and UVM (Figure 3).
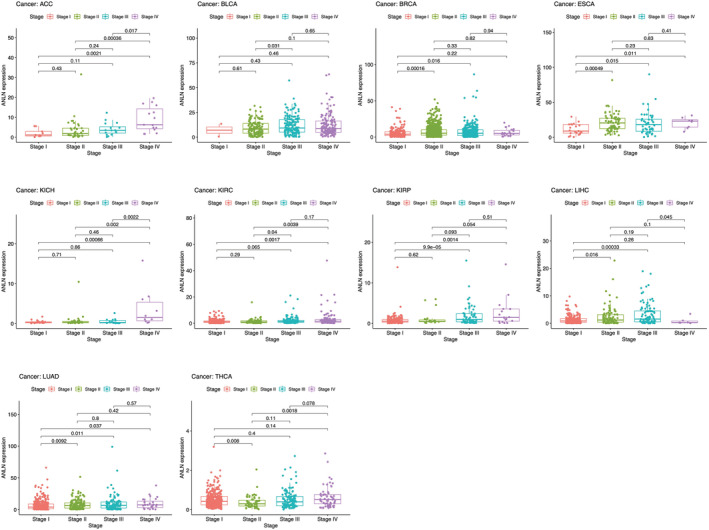
Furthermore, we explored the correlation between ANLN expression and different clinical features across pan‐cancer. Results revealed that higher expression of ANLN was significantly associated with advanced TNM stage in ACC, BLCA, BRCA, ESCA, KICH, KIRC, KIRP, LUAD, and reversely in LIHC. Interestingly, ANLN expression was higher in stage I compared with stage II (p = 0.008), but lower in stage II compared with stage IV (p = 0.0018) in THCA (Figure 4). ANLN expression also differs greatly in age and gender across different tumors (Figure S1).
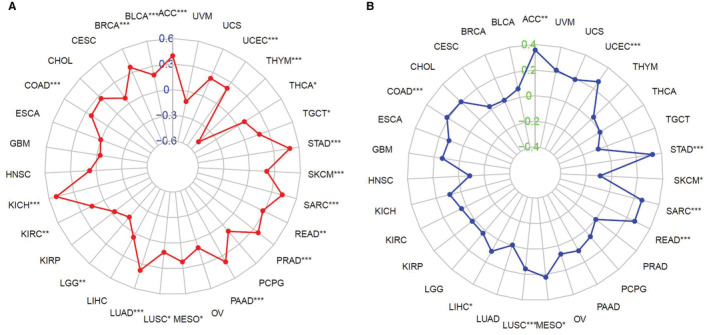
Tumor mutational burden and microsatellite instability analysis data
TMB and MSI are essential in the process of tumor formation and development. We assessed the association between ANLN expression and TMB/MSI across 31 cancers. Results suggested that elevated ANLN expression was significantly associated with higher TMB in ACC, BLCA, BRCA, COAD, KICH, KIRC, LUAD, LUSC, MESO, PAAD, PRAD, READ, SARC, SKCM, STAD, TGCT, THCA, and UCEC, while increased ANLN level was inversely related to TMB in LGG and THYM (Figure 5). In addition, ANLN expression was positively associated with MSI in ACC, COAD, LIHC, LUSC, MESO, READ, SARC, STAD, UCEC, and negatively linked to SKCM (Figure 5).
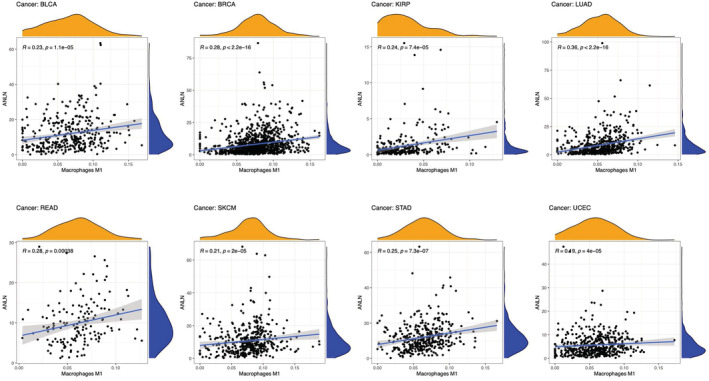
Correlation between ANLN expression and immune infiltration
Tumor‐infiltrating immune cells are important components for the tumor microenvironment, and are closely linked to tumorigenesis and progression.ref. cam45177-bib-0021 Infiltrating stromal and immune cells in tumor microenvironment play essential roles in disturbing tumor signaling and cancer biology.ref. cam45177-bib-0006 Hence, we firstly explored the correlation between ANLN expression and tumor‐infiltrating immune cells. Our results demonstrated that ANLN expression was significantly associated with the infiltration levels of macrophages M1 in 18 cancer types (Figure 6, those with p < 0.00001 were shown), CD4 + T cells in 12 cancer types, CD8+ T cells in 9 cancer types, T cells regulatory (Tregs) in 13 cancer types, B cells in 8 cancer types, M2 in 7 cancer types, NK cells in 9 cancer types, and neutrophils in 7 cancer types (those with p < 0.00001 were shown in Figure S2). For instance, in BLCA, ANLN expression was positively correlated with M1 macrophages and neutrophil infiltration, but negatively correlated with Tregs. In HNSC, ANLN expression was negatively correlated with CD8+ cells, Tregs, but positively correlated with CD4 + T cells memory resting.
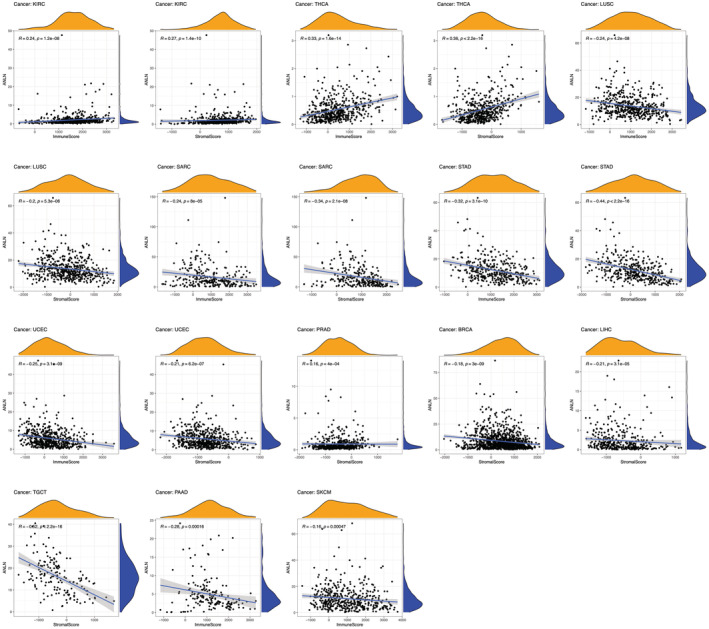
Secondly, our study investigated the association between ANLN expression and the immune and stromal cell ratio. Results showed that ANLN expression is positively correlated with the stromal score and immune score in KIRC, THCA, while negatively correlated with in LUSC, SARC, STAD, and UCEC (Figure 7). In addition, our study observed a statistical positive connection between ANLN expression and stromal score in PRAD but noted a negative connection in BRCA, LIHC, TGCT. Moreover, ANLN expression was negatively linked to immune score in PAAD and SKCM (Figure 7).
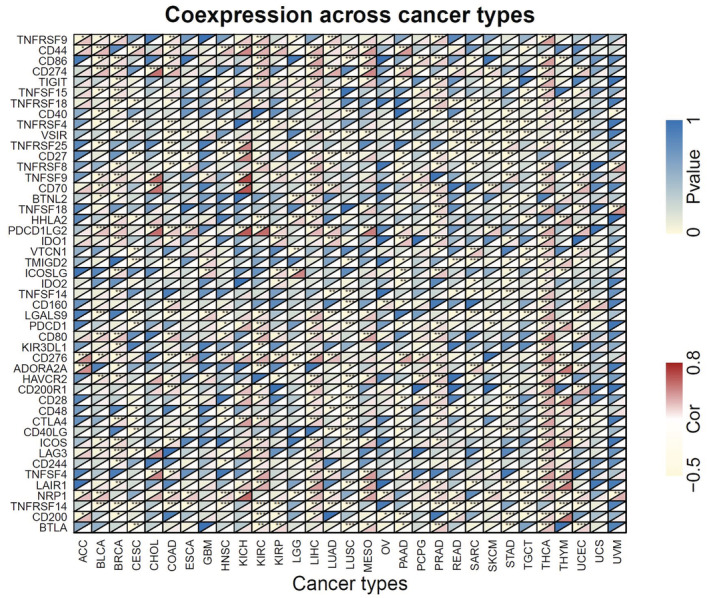
Immune checkpoint genes (ICGs) represent a novel target in cancer treatment. Therefore, we further examined the association between ANLN expression and the common immune checkpoint genes. As shown in Figure 8, cytotoxic T lymphocyte‐associated antigen 4 (CTLA‐4), programed death 1 (PD‐1), and PD‐L1 were positively associated with ANLN expression in multiple cancers, including BRCA, KICH, KIRC, LIHC, PRAD, and THCA. On the contrary, there was a negative connection between ANLN expression and CTLA‐4, PD‐1 in CESC, HNSC, and LUSC. Moreover, ANLN expression was positively linked to the majority of immune checkpoint genes in THCA. However, ANLN expression demonstrated a negative correlation with most of the immune checkpoint genes in GBM, UCS, and UVM.
Immunotherapeutic response prediction
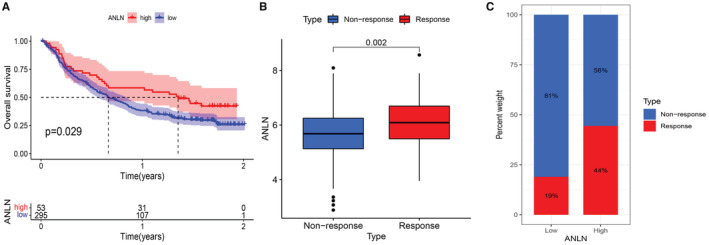
Considering the strong correlation between ANLN expression and tumor immune landscape, our study next explored the immunotherapeutic predictive ability of ANLN in an anti‐PD‐L1 cohort (IMvigor210). The Kaplan–Meier curves suggested that increased level of ANLN was significantly connected with prolonged OS (Figure 9A). In addition, patients with higher ANLN expression level exhibited better immune response (Figure 9B,C).
GO and KEGG‐based gene set enrichment analysis
Next, GSEA was applied to explore the functional enrichment of ANLN expression between high expression and low expression cohorts. GO enrichment term demonstrated that high expression of ANLN was mainly associated with cell immunity and cell proliferation (Figure S3). KEGG enrichment term exhibited that high expression of ANLN was positively involved “cell cycle” pathways in ACC, BRCA, KICH, HHSC, KICH, LUAD. MESO, OV, PAAD, PCPG, SARC, and UVM (Figure S4).
DISCUSSION
The pan‐cancer analysis project launched by TCGA was aimed to illustrate an integrated picture of commonalities and heterogeneities across different tumors.ref. cam45177-bib-0022 The project interpreted molecular aberrations from genomic, transcriptional, proteomic, and epigenetic levels in large patient cohorts, providing comprehensive information for early diagnosis and treatment strategy.ref. cam45177-bib-0023 Our study is the first to comprehensively analyze the expression pattern of ANLN and its correlation with patients’ prognosis and immune features in 31 different types of cancers.
Our study found that ANLN was upregulated in BLCA, BRCA, CESC, CHOL, COAD, ESCA, HNSC, KICH, KIRC, KIRP, LIHC, LUAD, LUSC, PAAD, PCPG, PRAD, READ, SARC, STAD, THCA, and UCEC. Cox regression and Kaplan–Meier analysis demonstrated that high ANLN expression was associated with shorter OS, DFI, and PFS in KIRP, LIHC, LUAD, and PAAD. This is in line with a recent study that ANLN is a prognostic biomarker for LUAD patients.ref. cam45177-bib-0024 ANLN is primarily involved in the process of cytokinesis and plays vital roles in tumorigenesis and progression.ref. cam45177-bib-0002 ANLN could significantly promote cancer cell growth and migration via targeting miR‐217ref. cam45177-bib-0025 and HMGA2.ref. cam45177-bib-0026 Upregulation of ANLN could activate RHOA, leading to the activation of phosphatidylinositol 3‐kinase‐AKT (PI3K/AKT), which promotes cell proliferation and progression.ref. cam45177-bib-0027 Another study revealed that ANLN might be involved in the metastasis of lung adenocarcinoma by promoting epithelial mesenchymal transformation of tumor cells.ref. cam45177-bib-0028 In addition, ANLN expression is also positively related to Wnt/β‐catenin signaling in gastric cancer.ref. cam45177-bib-0029 All these biological functions of ANLN may explain its oncogenic role in the above‐mentioned cancers. Notably, over‐expression of ANLN was correlated with better prognosis in THYM. Future studies are needed to explore its controversial role in human cancer.
TMB is emerging as a promising biomarker for immunotherapy. It refers to the total number of somatic coding mutations, gene insertion, base substitutions, and deletion errors detected per million bases.ref. cam45177-bib-0030 TMB presents as a useful estimation of tumor neoantigens, which could activate tumor‐specific T cells to recognize and kill tumor cells.ref. cam45177-bib-0030 High TMB are associated with prolonged survival in melanoma,ref. cam45177-bib-0031 NSCLC,ref. cam45177-bib-0032 and breast cancer.ref. cam45177-bib-0033 MSI is a molecular tumor phenotype caused by genomic hypermutability.ref. cam45177-bib-0034 Recent studies suggested that MSI could serve as an efficient biomarker for immunotherapy.ref. cam45177-bib-0035 Clinical trials showed that MSI‐H patients have better outcomes than microsatellite stable patients, owning to the activation of T lymphocyte recognition of neoantigens.ref. cam45177-bib-0035, ref. cam45177-bib-0036 Additionally, nearly all patients with MSI‐H have high TMB level and could respond more strongly to immunotherapy.ref. cam45177-bib-0017 Our study found that ANLN expression was positively associated with TMB or MSI in multiple cancers, including BLCA, which was consistent with the results from the BLCA immunotherapy cohort that patients with higher ANLN level had better immune responses and thus demonstrated longer OS. All the above findings suggested that ANLN expression in these tumors may help identify patients who are more likely to benefit from immunotherapy.
TME is a complex ecology consisting of tumor cells, tumor vasculature, surrounding immune and inflammatory cells, as well as adipocytes and fibroblasts.ref. cam45177-bib-0006 Tumor immune microenvironment (iTME), as a core part of the TME, consists of tumor‐infiltrating lymphocytes (TILs) and other immune cells, such as macrophages, neutrophils, and dendritic cells.ref. cam45177-bib-0037 Macrophages is a group of heterogeneous cells in the iTME and is strongly correlated with poor survival in diverse cancers.ref. cam45177-bib-0038 Tumor‐associated macrophages (TAMs) play essential roles in carcinogenesis and progression by providing cytokines and inducing tumor angiogenesis.ref. cam45177-bib-0039 TAMs could be polarized into two distinct phenotypes: (1) M1 macrophages, which act as an anti‐tumor defender; (2) M2 macrophages, which could promote tumor invasion and metastasis.ref. cam45177-bib-0039 Our study found that ANLN expression was positively associated with M1 macrophage infiltration, but negatively linked to M2 macrophage infiltration. Thus, we assumed that ANLN might participate in the host anti‐tumor immune response by inducing M1 macrophages infiltration. However, we also found that ANLN expression was negatively connected to CD8+ T cells in some cancer types. Cytotoxic CD8+ memory T cells in the TME is positively linked to cancer prognosis in diverse cancers.ref. cam45177-bib-0040, ref. cam45177-bib-0041, ref. cam45177-bib-0042 It could contribute to cancer prognoses by killing tumor cells.ref. cam45177-bib-0043 This phenomenon could be explained by the existence of other important regulators in the interaction between ANLN and CD8+ T cells. Further studies are needed to investigate the exact mechanism during this anti‐tumor progress. Single‐cell RNA sequencing (scRNA‐seq), which allows for transcriptome‐wide analyses of individual cells may help offer new insights in the future. A previous study showed that ANLN along with other regulators are significantly associated with the expression of naïve B cells, regulatory T cells, and neutrophils in head and neck squamous cell carcinoma (HNSCC).ref. cam45177-bib-0044 However, studies focusing on the functional mechanism of ANLN in immune cell infiltration is rare. Our GO and KEGG pathway analyses revealed that ANLN was mainly enriched in cell cycle, immunity, and metabolic‐related pathway, which may explain its oncogenic role in different tumors. Further studies are needed to find the exact functional mechanism between ANLN expression and different immune cells. Moreover, our analyses demonstrated that ANLN expression had a significant correlation with most of the common immune checkpoint markers. We further validated the immunotherapeutic predictive ability of ANLN in an anti‐PD‐L1 cohort (IMvigor210). Results showed that high ANLN expression was associated with prolonged OS and better immune response, which was consistent with the above definitions. All these above findings confirmed our hypothesis that ANLN could be a robust biomarker for prognosis and immune response prediction.
Several limitations should be considered in our study. First, because of the use of microarray and sequencing data from different databases, systematic bias may occur. Hence, as mentioned above, scRNA‐seq or other higher resolution methods could be performed. Second, our study was a retrospective study, so future prospective researches are needed to verify the correlation between ANLN expression and patients’ prognosis and tumor immunity. Finally, the precise mechanisms by which ANLN facilitates tumor progression and tumor immunity remain largely obscure. Further mechanistic studies on ANLN at both cellular and molecular levels are needed to fully elucidate its functions.
In conclusion, our study found that ANLN was upregulated in diverse cancers and its aberrant expression was associated with pan‐cancer prognosis, MSI, TMB, the tumor immune microenvironment, and immune checkpoint genes. Therefore, ANLN may prove to be a promising biomarker for prognostic judgment and immunotherapy.
AUTHOR CONTRIBUTIONS
JC, QYZ, and XZ conceived and designed this study. LZ, YW, and YH obtained the data. XZ, LZ, YW, and YH analyzed the data. XPW and ZBH helped discuss the results. LZ drafted the manuscript. All authors contributed to the article and approved the submitted version.
CONFLICT OF INTEREST
The authors declare that there are no conflicts of interest.
ETHICS APPROVAL AND CONSENTS TO PARTICIPATE
This study was approved by the Ethics Committee of The Affiliated Suzhou Hospital of Nanjing Medical University and each participant had signed informed consent (Ethics Approval No. KL901198).
Supplementary Materials
References
- Role of anillin in tumour: from a prognostic biomarker to a novel target.. Cancers (Basel)., 2020. [PubMed]
- Importin beta2 mediates the Spatio‐temporal regulation of anillin through a noncanonical nuclear localization signal.. J Biol Chem., 2015. [PubMed]
- Overexpression of anillin (ANLN) is correlated with colorectal cancer progression and poor prognosis.. Cancer Biomark., 2016. [PubMed]
- ANLN and KDR are jointly prognostic of breast cancer survival and can be modulated for triple negative breast cancer control.. Front Genet., 2019. [PubMed]
- Screening hub genes as prognostic biomarkers of hepatocellular carcinoma by bioinformatics analysis.. Cell Transplant., 2019. [PubMed]
- The immune contexture in human tumours: impact on clinical outcome.. Nat Rev Cancer., 2012. [PubMed]
- Immunological role and prognostic value of APBB1IP in Pan‐cancer analysis.. J Cancer., 2021. [PubMed]
- A comprehensive prognostic and immune analysis of SLC41A3 in Pan‐cancer.. Front Oncol., 2020. [PubMed]
- An integrative Pan‐cancer analysis revealing LCN2 as an oncogenic immune protein in tumor microenvironment.. Front Oncol., 2020. [PubMed]
- Prognostic and immunological role of mRNA ac4C regulator NAT10 in Pan‐cancer: new territory for cancer research?. Front Oncol., 2021. [PubMed]
- Identification of prognostic genes in adrenocortical carcinoma microenvironment based on bioinformatic methods.. Cancer Med., 2020. [PubMed]
- Genomics of response to immune checkpoint therapies for cancer: implications for precision medicine.. Genome Med., 2018. [PubMed]
- Updated analysis from KEYNOTE‐189: pembrolizumab or placebo plus pemetrexed and platinum for previously untreated metastatic nonsquamous non‐small‐cell lung cancer.. J Clin Oncol., 2020. [PubMed]
- KEYNOTE‐590: phase III study of first‐line chemotherapy with or without pembrolizumab for advanced esophageal cancer.. Future Oncol., 2019. [PubMed]
- Pembrolizumab in microsatellite‐instability‐high advanced colorectal cancer.. N Engl J Med., 2020. [PubMed]
- Immune checkpoint blockade: a common denominator approach to cancer therapy.. Cancer Cell., 2015. [PubMed]
- Analysis of 100,000 human cancer genomes reveals the landscape of tumor mutational burden.. Genome Med., 2017. [PubMed]
- Microsatellite instability in cancer of the proximal colon.. Science., 1993. [PubMed]
- Inferring tumour purity and stromal and immune cell admixture from expression data.. Nat Commun., 2013. [PubMed]
- TGFbeta attenuates tumour response to PD‐L1 blockade by contributing to exclusion of T cells.. Nature., 2018. [PubMed]
- Clinical relevance of host immunity in breast cancer: from TILs to the clinic.. Nat Rev Clin Oncol., 2016. [PubMed]
- The cancer genome atlas Pan‐cancer analysis project.. Nat Genet., 2013. [PubMed]
- Pan‐cancer analysis of whole genomes.. Nature., 2020. [PubMed]
- A combined two‐mRNA signature associated with PD‐L1 and tumor mutational burden for prognosis of lung adenocarcinoma.. Front Cell Dev Biol., 2021. [PubMed]
- Regulation of Actin‐binding protein ANLN by antitumor miR‐217 inhibits cancer cell aggressiveness in pancreatic ductal adenocarcinoma.. Oncotarget., 2017. [PubMed]
- High mobility group AT‐hook 2 promotes tumorigenicity of pancreatic cancer cells via upregulating ANLN.. Exp Cell Res., 2020. [PubMed]
- ANLN plays a critical role in human lung carcinogenesis through the activation of RHOA and by involvement in the phosphoinositide 3‐kinase/AKT pathway.. Cancer Res., 2005. [PubMed]
- Overexpression of ANLN in lung adenocarcinoma is associated with metastasis.. Thorac Cancer., 2019. [PubMed]
- In silico analysis of expression pattern of a Wnt/beta‐catenin responsive gene ANLN in gastric cancer.. Gene., 2014. [PubMed]
- Tumor mutational load predicts survival after immunotherapy across multiple cancer types.. Nat Genet., 2019. [PubMed]
- Significance of tumor mutation burden in immune infiltration and prognosis in cutaneous melanoma.. Front Oncol., 2020. [PubMed]
- Blood‐based tumor mutational burden as a predictor of clinical benefit in non‐small‐cell lung cancer patients treated with atezolizumab.. Nat Med., 2018. [PubMed]
- Tumor mutational burden is a determinant of immune‐mediated survival in breast cancer.. Onco Targets Ther., 2018
- Classification and characterization of microsatellite instability across 18 cancer types.. Nat Med., 2016. [PubMed]
- PD‐1 blockade in tumors with mismatch‐repair deficiency.. N Engl J Med., 2015. [PubMed]
- Somatic mutation profiles of MSI and MSS colorectal cancer identified by whole exome next generation sequencing and bioinformatics analysis.. PLoS One., 2010. [PubMed]
- The tumor microenvironment innately modulates cancer progression.. Cancer Res., 2019. [PubMed]
- Macrophages as key drivers of cancer progression and metastasis.. Mediators Inflamm., 2017. [PubMed]
- Prognostic value of a nine‐gene signature in glioma patients based on tumor‐associated macrophages expression profiling.. Clin Immunol., 2020. [PubMed]
- CD4/CD8 + T cells, DC subsets, Foxp3, and IDO expression are predictive indictors of gastric cancer prognosis.. Cancer Med., 2019. [PubMed]
- Tertiary lymphoid structures improve immunotherapy and survival in melanoma.. Nature., 2020. [PubMed]
- Tumor‐infiltrating lymphocytes and CD8(+) T cells predict survival of triple‐negative breast cancer.. J Cancer Res Clin Oncol., 2019. [PubMed]
- CD8(+) T cell states in human cancer: insights from single‐cell analysis.. Nat Rev Cancer., 2020. [PubMed]
- Identification novel prognostic signatures for head and neck squamous cell carcinoma based on ceRNA network construction and immune infiltration analysis.. Int J Med Sci., 2021. [PubMed]