
The endocrine disruptor DEHP and the ECS: analysis of a possible crosstalk
Abstract
Studies of the last decade associated the environmental contamination by di-(2-ethylhexyl)-phthalate (DEHP) with obesity and endocrine malfunction. DEHP was found to interact with several receptors – among them are receptors of the endocannabinoid system (ECS) with high expression levels in adipose tissue. Furthermore, the correlation for BMI and body fat to the serum endocannabinoid level raises the question if the obesogenic and endocrine-disrupting DEHP effects are mediated via the ECS. We therefore characterized the ECS in a human cell model of adipogenesis using the SGBS preadipocytes to subsequently investigate if DEHP exposure affects the intrinsic ECS. The receptors of the ECS and the endocannabinoid-metabolizing enzymes were upregulated during normal adipogenesis, accompanied by an increasing secretion of the adipokines adiponectin and leptin. DEHP affected the secretion of both adipokines but not the ECS, suggesting DEHP to alter the endocrine function of adipocytes without the involvement of the intrinsic ECS.
Article type: Research Article
Keywords: adipocytes, DEHP, endocannabinoid system, leptin, SGBS, endocrine disruptor
Affiliations: Department of Anatomy and Cell Biology, Faculty of Medicine, Martin Luther University Halle-Wittenberg, Halle (Saale), Germany
License: © 2020 The authors CC BY 4.0 This work is licensed under a Creative Commons Attribution 4.0 International License.
Article links: DOI: 10.1530/EC-19-0548 | PubMed: 31910153 | PMC: PMC6993259
Relevance: Moderate: mentioned 3+ times in text
Full text: PDF (967 KB)
Introduction
During the last decades, environmental pollution as well as the prevalence of obesity and metabolic syndrome have increased (ref. 1). So, an environmental link to obesity is barely surprising (ref. 2, ref. 3, ref. 4, ref. 5). Increasing evidence was found for an adverse impact on human health due to environmental agents, such as phthalates (ref. 6). Di-(2-ethylhexyl)-phthalate (DEHP) has belonged to the most abundant phthalates in industry and consumer goods for many years acting as an endocrine-disrupting chemical (ref. 7). As DEHP is not covalently bound to PVC, it easily migrates into the environment and by that accumulates in lipophilic products like cosmetics or food and binds to house dust particles (ref. 8). Associations to numerous health problems, including obesity, diabetes and the metabolic syndrome, have been observed by epidemiological studies (ref. 9, ref. 10, ref. 11, ref. 12, ref. 13). Furthermore, investigations in mice identified DEHP as an obesogen by increasing food intake, body weight, fat mass, serum leptin and decreasing serum adiponectin (ref. 14, ref. 15, ref. 16). Accordingly, data from in vitro models showed a DEHP-dependent impairment of adipogenesis and adipocyte function (ref. 14, ref. 17). Analyses on underlying mechanisms are difficult, because energy metabolism and endocrine homeostasis involve complex regulatory systems – among them is the endocannabinoid system (ECS). Endocannabinoids are endogenous ligands of the G-protein-coupled cannabinoid receptors, imitating several effects of the pharmacological active substance Δ9-tetrahydrocannabinol (THC) from cannabis sativa (ref. 18). Endocannabinoids like the N-arachidonoylethanolamine (AEA) and 2-arachidonoylglycerol (2-AG) mediate their effects via CB1 and CB2 cannabinoid receptors (ref. 19, ref. 20, ref. 21), but also via the recently identified GPR55 (ref. 22) and non-cannabinoid receptors like the transient receptor potential vanilloid 1 (TRPV1) (ref. 23) or the peroxisome proliferator-activated receptors (PPARs) (ref. 24). Endocannabinoid levels are regulated by the synthesizing enzymes N-acylphosphatidylethanolamine phospholipase D (NAPE-PLD) and diacylglycerol lipase (DAGL) (ref. 25, ref. 26) as well as by the metabolizing fatty acid amide hydrolase (FAAH) and monoacylglycerol lipase (MAGL) (ref. 27, ref. 28). The influence of phthalates on ECS in peripheral organs is poorly investigated. However, the effect of di-isononyl phthalate (DiNP) as one of the dominating alternatives to DEHP was studied in fish models demonstrating a deregulation of the intrinsic ECS in the gonads, the liver as well in the hepatic lipid metabolism (ref. 29, ref. 30, ref. 31). DiNP exhibited adipogenic activity in murine 3T3-derived adipocytes (ref. 32).
Adipose tissue expression of ECS components differs between lean and obese subjects as reflected by altered blood endocannabinoid levels (ref. 33, ref. 34, ref. 35, ref. 36, ref. 37). Literature in the field of ECS, adipogenesis and adipose tissue as a metabolic and endocrine organ still draw a blurred picture of possible interactions. As CB1 activation promotes adipocyte proliferation and differentiation, it furthermore positively affects insulin-stimulated but not basal glucose uptake in 3T3-derived adipocytes (ref. 38, ref. 39, ref. 40). Accordingly, an increase of glucose uptake after activation of CB1 was demonstrated in human primary adipocytes – accompanied by calcium influx and translocation of GLUT4. However, adiponectin and leptin were not altered (ref. 41). The inhibition of CB1R in adipocytes directly reduced the leptin secretion in mice. In line with these results, an in vitro study in 3T3-derived adipocytes confirmed increased leptin levels after treatment with different CB1R agonists that were inhibited by the employment of a CB1R inverse agonist (ref. 42). In human adipose tissue, no association was found between CNR1-mRNA level and adiponectin expression, its secretion or circulating adiponectin (ref. 43). After CB1 antagonism in rats, a higher adipose gene expression and serum level of adiponectin was detected. This finding was proposed as a consequence of reduced food intake (ref. 44). Contrary to the assumption of an indirect effect, the in vitro blockade of CB1 led to an upregulation of adiponectin in 3T3-derived adipocytes (ref. 38, ref. 39, ref. 45). THC also elevated adiponectin gene expression in this cell line. The authors discussed that the variety of different types and concentrations of CB1-manipulating agents may likely be responsible for the miscellaneous effects among studies (ref. 46). Comparing different ligands of the ECS in human bone marrow derived adipocytes, an exclusive activation of CB1-inhibited adipogenesis paralleled by a reduction of adiponectin. Nevertheless, these effects of CB1 activation were diminished when ligands not only bound to CB1 but also to the non-CB1/CB2 receptor PPARgamma, which is a crucial transcription factor of adipogenesis (ref. 47).
To date there are no reports on the relationship between DEHP and the ECS in obesity. The known fact of interactions of DEHP with receptors of the ECS (ref. 48, ref. 49) raised the question whether obesogenic and endocrine-disrupting DEHP effects in adipocytes are mediated via the ECS. For the present study, we first characterized the intrinsic ECS in a human cell model of adipogenesis using the Simpson-Golabi-Behmel Syndrome (SGBS) preadipocytes followed by investigating the impact of DEHP on the ECS as an endocrine modulator of the adipokine system.
Materials and methods
Chemicals
DEHP was dissolved in dimethyl sulfoxide (DMSO), both purchased from Sigma-Aldrich, and stored as a 1000-fold stock solution until further use.
Cell culture
The SGBS preadipocytes were kindly provided by Prof M Wabitsch (Division of Pediatric Endocrinology and Diabetes, Department of Pediatrics and Adolescent Medicine, Ulm University Medical Center, Ulm, Germany). These preadipocytes are a non-immortalized cell model for adipogenesis cultured and differentiated as described previously without modifying the protocols (ref. 50, ref. 51). During the induction phase (day 0 to day 4), cells were exposed to a final DEHP concentration of 128 µM (50 µg/mL) and a concentration of 0.1% DMSO in the culture media, whereas controls were run as vehicle controls with 0.1% DMSO only. These experimental conditions are based on in-vitro investigations of our group previously performed in murine C3H10-T1/2 mesenchymal stem cells identifying the induction phase as a particularly vulnerable exposure window of adipogenesis (ref. 17, ref. 52) and in the SGBS cell model revealing effective but non-toxic concentrations of used substances (ref. 53). Furthermore, the herein applied DEHP concentration is considered as environmentally relevant (ref. 54, ref. 55, ref. 56). At day 8 of differentiation, all experiments have been finalized. Samples were taken at day 0, 4 and 8 of adipogenesis.
Quantitative real-time PCR
Quantitative realtime PCR (qRT-PCR) was performed to measure the mRNA expression levels in a StepOnePlus™ Real-Time PCR System (Applied Biosystems). Plasmid standards were generated based on a gene-specific target sequence. Absolute mRNA copies were calculated by quantitative standard curves using serial dilutions (106, 105, 104 and 103) of gene-specific plasmid standards. Assays were run with duplicates of each cDNA sample as well as a no template control (NTC) in a 96-well format for the following genes: ADIPOR1 and ADIPOR2, CNR1 and CNR2, DAGLalpha, FAAH, GLUT1 and GLUT4, GPR55, LEPR, MAGL, NAPE-PLD and TRPV1. For normalization, we analyzed the expression of the housekeeping gene TATA-box-binding protein (TBP). Absolute mRNA expression was calculated as copy number per 103 molecules TBP. The primers and amplicons were as shown in Table 1.
Table 1: Primers for quantitative RT-PCR.
| Gene | Accession number | Forward primer | Reverse primer | Tm (°C) | Amplicon (bp) |
|---|---|---|---|---|---|
| ADIPOR1 | NM_001290629 | TGCGGCGGGGAGTTTAGAAG | CGTGTCAGCTTCCCTGTTACT | 63 | 245 |
| ADIPOR2 | NM_024551 | GAGACACGCGGATCAACTCA | GTTGGTGCCCTTTTCTGAGC | 60 | 175 |
| CNR1 | NM_033181 | CTCAGTCATTTTGAGCTCAGCC | GCCATGTCACCTTTGATGTCTTC | 60 | 153 |
| CNR2 | NM_001841 | GCTCCTCATCTGTTGGTTCC | TGACCATGGAGTTGATGAGGC | 60 | 125 |
| DAGLa | NM_006133 | AGAATGTCACCCTCGGAATG | GGTTGTAGGTCCGCAGGTTA | 60 | 150 |
| FAAH | NM_001441 | TCAAGGAGTGCTTCACCTACAAG | GTCATAGCTGAACATGGACTGTG | 60 | 164 |
| GLUT1 | NM_006516 | TGGCATCAACGCTGTCTTCT | CTAGCGCGATGGTCATGAGT | 60 | 212 |
| GLUT4 | NM_001042 | ACTGGCCATTGTTATCGGCA | GTCAGGCGCTTCAGACTCTT | 60 | 213 |
| GPR55 | NM_005683 | GGTGCTCTCCCTCCCATT | GCTCACCAGTAGCGGGTAAC | 60 | 172 |
| LEPR | NM_002303 | ACACCAGAGTGATGCAGGTTT | ATGCTCAAACGTTTCTGGCTTC | 62 | 187 |
| MAGL | NM_007283 | ATCACCATTCCCCAAATTGA | GATGTACCAGCCCTTCTGGA | 60 | 204 |
| NAPE-PLD | NM_198990 | TCACGGATCCCATCTTTAGC | TCTCACAGCCACATTTTTGC | 60 | 243 |
| TBP | NM_003194 | TGTGCTCACCCACCAACAAT | AGTCGTCTTCCTGAATCCCT | 60 | 199 |
| TRPV1 | NM_080704 | TGACCCTCCTGGTGGAGA | CTGCAGCAGGAACTTCACG | 60 | 158 |
adiponectin receptor 1 and 2 (ADIPOR1 and ADIPOR2), cannabinoid receptor 1 and 2 (CNR1 and CNR2), diacylglycerol lipase alpha (DAGLa), fatty acid amide hydrolase (FAAH), glucose transporter 1 and 4 (GLUT1 and GLUT4), G protein-coupled receptor 55 (GPR55), leptin receptor (LEPR), monoacylglycerol lipase (MAGL), N-acylphosphatidylethanolamine phospholipase D (NAPE-PLD), TATA-box binding protein (TBP) and transient receptor potential vanilloid 1 (TRPV1).
Hormone assay
Cell supernatants were collected to measure the concentrations of leptin (high sensitive Leptin ELISA, IBL, Hamburg, Germany) and adiponectin (Quantikine® ELISA Human Total Adiponectin/Acrp30, BioVendor, Kassel, Germany) by ELISA according to manufacturer’s manual. ELISA data were normalized to the protein concentration of individual samples. Protein was isolated using radioimmunoprecipitation assay (RIPA) buffer including protease and phosphatase inhibitors (Roche). Protein concentration was determined by the BioRad Protein Assay (BioRad).
Western blot
For protein analyses of ECS components, cells were harvested in lysis buffer containing 80 mM Tris, 70 mM sodium dodecyl sulfate (SDS), 0.3 M saccharose, 3 mM sodium orthovanadate and 0.5 mM phenylmethylsulfonyl floride (PMSF) at pH 7.4. Samples of 40 µg protein were separated by a 12.5% (w/v) SDS-polyacrylamid gel before blotting onto nitrocellulose membrane (Protran BA 85, GE Healthcare). Non-specific protein-binding sites were blocked for 30 min with 5% (w/v) milk (Carl Roth, Karlsruhe, Germany) or 10% (v/v) Roti-block solution (Carl Roth) in TBST. For protein detection, primary antibodies against CB1, DAGLalpha and DAGLbeta, FAAH, MAGL and NAPE-PLD as well as against beta-ACTIN and GAPDH as housekeeping proteins were used (Table 2). Membranes were incubated for 16 h at 4°C. They were subsequently washed and the horseradish peroxidase-conjugated secondary antibodies (Table 2) were applied for 1 h at room temperature. Chemiluminescence detection was performed by Luminata Forte (Millipore). ImageJ analysis software version 1.46r (National Institutes of Health, Laboratory for Optical and Computational Instrumentation, University of Wisconsin, Madison, WI, USA) was used for the analysis of the intensity of the immunoreactive bands.
Table 2: Antibodies for Western blot (WB) and immunohistochemistry (IHC).
| Dilution WB | Dilution IHC | Manufacturer | |
|---|---|---|---|
| Primary antibody | |||
| Rabbit polyclonal antibody against CB1 | 0.5 µg/mL | 1.65 µg/mL | Cayman, Mississippi, USA |
| Rabbit pig antibody against DAGLalpha | 1:2000 | 1:200 | Frontier Institute, Hokkaido, Japan |
| Rabbit antibody against DAGLbeta | 1:1000 | 1:100 | Thermo Scientific, Dreieich, Germany |
| Rabbit antibody against human FAAH | 1:1000 | 1:200 | Cayman |
| Rabbit polyclonal antibody against human MAGL | 1:1000 | 1:200 | Cayman |
| Rabbit polyclonal antibody against human NAPE-PLD | 1:1000 | 1:200 | Cayman |
| Mouse antibody against human beta-ACTIN | 1:5000 | Cell Signaling, Boston, USA | |
| Rabbit antibody against human GAPDHa | 1:1000 | Cell Signaling | |
| Secondary antibody | |||
| Anti-rabbit-IgG | 1:20,000 | Vektor laboratories, Burlingame, CA | |
| Anti-mouse-IgG | 1:10,000 | Vektor laboratories | |
| Anti-rabbit-IgG | 1:2 | DAKO, Hamburg, Germany | |
aThis primary antibody is already horseradish peroxidase-conjugated and does not require any secondary antibody.
cannabinoid receptor 1 (CB1), diacylglycerol lipase alpha and beta (DAGLalpha and DAGLbeta), fatty acid amide hydrolase (FAAH), glyceraldehyde 3-phosphate dehydrogenase (GAPDH), monoacylglycerol lipase (MAGL), N-acylphosphatidylethanolamine phospholipase D (NAPE-PLD).
Immunohistochemical staining
Fifty thousand SGBS cells were plated on PLL (Millipore) covered glass plates and treated according to the protocol (see ‘Cell culture’ section). On day 0, 4 and 8 cells were fixed with 4% paraformaldehyde for 25 min. Before staining, the cells were washed with 0.02 M PBS and incubated with goat serum (Sigma-Aldrich) for 30 min. Primary antibodies against CB1, DAGLalpha and DAGLbeta, FAAH, MAGL and NAPE-PLD (Table 2) were diluted in 0.05% BSA (Sigma-Aldrich) and incubated overnight. The next day, cells were washed three times with PBS/Triton and incubated with a HRP-labelled secondary antibody diluted in PBS for 1 h. After washing with PBS, cells were covered with 0.05 M Tris buffer and exposed to DAB (Sigma-Aldrich) for 5 min. Finally, hematoxylin (Carl Roth) staining was performed and the plates were covered with Entallan (Millipore).
Statistical analyses
At least four independent experiments (N) were performed for each group. Data was presented as mean ± s.e.m. For expression analyses during adipogenesis without DEHP exposure, the values of the DMSO control group at day 0 was defined as reference and the ANOVA with the Bonferroni’s post hoc test performed. To evaluate differences between the DMSO control group and the DEHP exposure group, an unpaired Student’s t-test or the Wilcoxon rank-sum test were used. Data differences were considered as statistically significant at P value ≤0.05.
Results
DEHP alters the secretion of adiponectin and leptin during adipogenesis
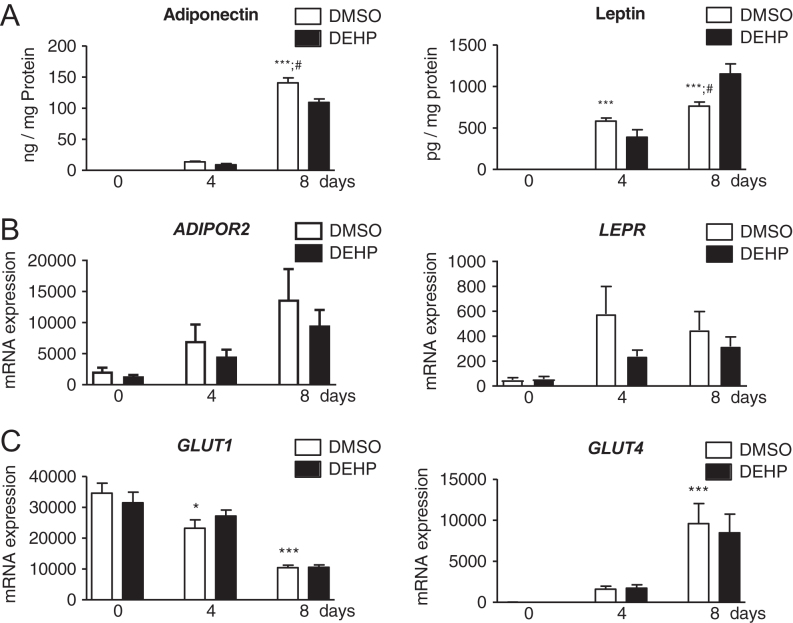
To evaluate the efficacy of our experimental conditions on adiponectin and leptin, their receptors and the glucose transporters were investigated as adipogenic markers (Fig. 1). During normal adipogenesis, the secretion level of both adipokines were significantly increased at day 4 and 8 for leptin and at day 8 for adiponectin compared to day 0. DEHP exposure led to significantly reduced adiponectin and increased leptin values at day 8 (Fig. 1A). Gene expression of receptors ADIPOR2 and LEPR was not significantly altered (both with P = 0.06 during normal adipogenesis). DEHP had no influence on their gene expression (Fig. 1B). ADIPOR1 was not expressed. During normal adipogenesis, both investigated glucose transporters showed alterations with a significant decrease of GLUT1 at day 4 and 8, whereas GLUT4 increased at day 8. No effect of DEHP exposure was detected for both glucose transporters (Fig. 1C).
The receptors and the metabolizing enzymes of the ECS were differentially expressed during normal adipogenesis
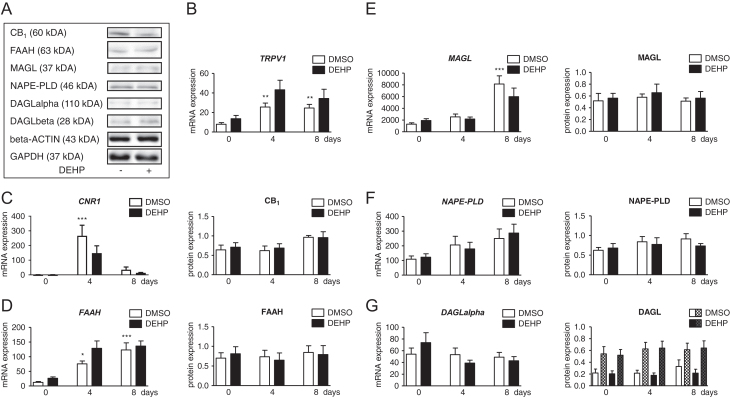
The expression of components of ECS was studied during adipogenesis in SGBS cells (Fig. 2). CNR2 and GPR55 were absent at detectable expression levels. The expression of the receptor TRPV1 was significantly upregulated at day 4 and remained at a high level until day 8 (Fig. 2B). Another receptor, CNR1, was significantly and transiently upregulated at day 4 and showed a decline at day 8 when compared to day 0 values (Fig. 2C). FAAH expression was increased at day 4 and day 8 (Fig. 2D). An elevation in MAGL expression was found at day 8 only (Fig. 2E). Data on endocannabinoid-synthesizing enzymes showed no alteration for NAPE-PLD and DAGLalpha expression (Fig. 2F and G).
DEHP did not affect the expression of the ECS
After investigating the expression of the ECS during normal adipogenesis, SGBS cells were exposed to DEHP within the induction phase, and its influence on mRNA and protein levels was measured (Fig. 2). DEHP did not significantly change the expression of TRPV1, CNR1, FAAH, MAGL, NAPE-PLD and DAGLalpha (Fig. 2B, C, D, E, F and G). This was verified for CB1, FAAH, MAGL, NAPE-PLD and DAGL at protein level (Fig. 2C, D, E, F and G). Western blots for CB2, GPR55 and TRPV1 could not be performed due to a lack of appropriate specific antibodies.
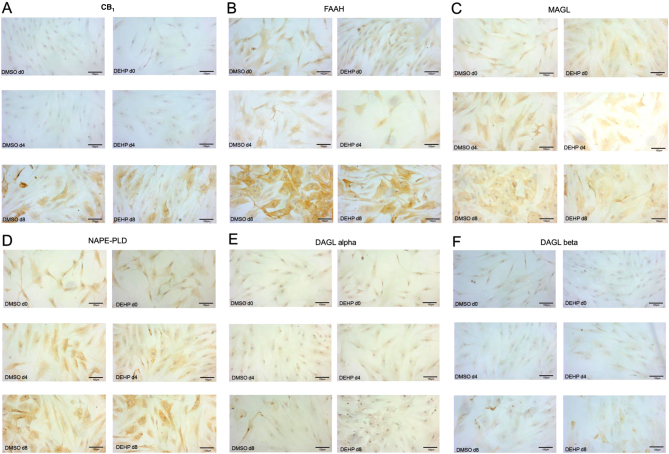
Furthermore, immunohistochemical studies were performed. During induction phase, SGBS cells grew mostly remaining spindle shaped. With increasing differentiation, cells became larger with expanded somata and visible lipid droplets. Additionally, only few isolated, very small cells were present in cultures (Fig. 3). All investigated ECS components were expressed in fully differentiated SGBS adipocytes. Comparing the data obtained from qRT-PCR, Western blot and immunohistochemistry describes an ECS component-specific characteristic pattern during adipogenesis with partial discrepancy. CB1 and FAAH immunoreactivities were evident in the somata of day 8-adipocytes with only very few positive cells at days 0 and 4 (Fig. 3A and B). A rather weak immunoreactivity was observed for MAGL at day 4. At day 8, MAGL was stained with stronger intensity mainly located around lipid droplets (Fig. 3C). NAPE-PLD immunoreactivity was found at day 4 and with much stronger intensity at day 8 (Fig. 3D). Whereas DAGLalpha positive cells were particularly abundant at days 0 and 4, a weak DAGLalpha immunoreactivity was observed at day 8 of adipogenesis (Fig. 3E). DAGLbeta-positive cells were more abundant and intensely stained than DAGLalpha positive cells (Fig. 3F). Notably, for all ECS components, no difference was detected between the DEHP-exposed and the DMSO-control group at all days investigated (Fig. 3A, B, C, D, E and F).
Discussion
The present study aimed to investigate whether the obesogenic and endocrine-disrupting DEHP effects in adipocytes were mediated via the ECS. At first, we characterized the intrinsic ECS during normal adipogenic differentiation from SGBS preadipocytes to mature adipocytes. The receptors CNR1 and TRPV1 and the endocannabinoid-metabolizing enzymes FAAH and MAGL as well as the endocannabinoid-synthetizing enzymes NAPE-PLD and DAGLalpha were expressed. CNR2 and GPR55 – two additional receptors – were not detectable. The findings on CNR2 are in agreement with data from bone marrow derived adipocytes (ref. 47). Also, expression and binding assays performed in human s.c. adipose tissue revealed a functional expression for CB1 and TRPV1, but not for CB2 (ref. 57). The presence of CB2 in adipose tissue has been a matter of controversy. An earlier study postulated contaminations with vascular, blood and immune cells as a potential source of positive CB2-findings (ref. 41). In our cell model, a contamination can be ruled out. By further analysing the expression pattern of CNR1 and TRPV1, we found both receptors to be upregulated during the induction phase followed by a decrease during differentiation. For CB1, an increasing immunoreactivity was detectable throughout maturation. In human bone marrow derived adipocytes, CNR1 and TRPV1 also increased after induction of adipogenesis. Additionally, a CB1-dependent inhibition of the differentiation was demonstrated, while otherwise AEA promoted adipogenesis by transactivation of PPARgamma (ref. 47). Comparing cannabinoid receptor expression in human primary fat cells before and during adipogenic differentiation, it was shown that CNR1 was not expressed in preadipocytes, but rapidly appeared with differentiation, whereas CNR2 started at low levels to become undetectable (ref. 41). In contrast, in preadipocytes and mature adipocytes isolated from human omental and s.c. adipose tissue, CNR1 and CNR2 were present in both cell types, but were more abundant in mature adipocytes. Interestingly, CNR2 was much higher expressed than CNR1 with functional proteins for both (ref. 58). A functional expression of CB1, CB2 and TRPV1 in murine 3T3-derived adipocytes was demonstrated during adipogenesis with increasing CB1 but declining CB2, whereas TRPV1 was unchanged (ref. 40). Investigating the protein expression pattern of the enzymes responsible for synthesis or degradation of main endocannabinoids namely 2-AG and AEA in the SGBS cell model showed: (a.) a slight increase in immunoreactivity of both DAGLalpha and MAGL for metabolizing 2-AG and (b.) no change in NAPE-PLD accompanied by an increase in FAAH immunoreaction for AEA during adipogenesis. The findings might point to a more or less constant 2-AG and a decrease in AEA levels. Studies with activity and expression data demonstrated a functional expression of enzymes involved in biosynthesis and hydrolysis of endocannabinoids in human s.c. and abdominal adipose tissue (ref. 41, ref. 57). Differentiation of murine 3T3-derived adipocytes was accompanied by an increasing degradation of AEA by FAAH, whereas synthesis by NAPE-PLD was not influenced. Additionally, AEA was found to enhance – via CB1 – the insulin-regulated glucose uptake, that typical increases during adipogenic maturation (ref. 40). Furthermore, stimulation of murine 3T3-derived adipocytes with leptin decreased AEA and 2-AG levels (ref. 45). As adipogenic differentiation is paralleled by increasing leptin levels, the reduction of endocannabinoids by upregulation of endocannabinoid-metabolizing enzymes during adipogenesis may be the consequence of leptin-mediated regulation.
Concordant to previous data (ref. 50), the typical adipocyte markers leptin, adiponectin and GLUT- were upregulated during adipogenic maturation of SGBS adipocytes. Our investigations include not only the insulin-regulated glucose transporter GLUT4 but also the basal glucose transporter GLUT1. In contrast to the increase of GLUT4, we found GLUT1 to be reduced. This is in agreement with results of previous studies in human adipocyte precursor cells, assuming differentiation to be correlated with the development of insulin sensitivity in mature adipocytes (ref. 59, ref. 60). As already described, DEHP exposure did not influence cell proliferation, but impaired adipogenic differentiation and lowered the lipid content of mature adipocytes (ref. 53). In proof of the efficacy of the present experiments, we investigated the DEHP action on adipokines. In agreement to our recent data (ref. 53), adiponectin was decreased and leptin was increased after DEHP exposure in mature SGBS adipocytes. Notably, in murine 3T3-derived adipocytes, DEHP reduced the cellular lipid content and adiponectin but increased the cell proliferation (ref. 14). In contrast, the higher number of adipocytes and the enhanced adipogenic differentiation of the murine mesenchymal stem cell line C3H/10T1/2 by DEHP exposure were associated with an increase of adiponectin expression (ref. 17). In vivo, DEHP exposure of mice and rats caused a gain of fat mass and a decrease of the serum adiponectin but increase of serum leptin (ref. 14, ref. 15, ref. 16, ref. 61). Environmental contamination by DEHP is associated with human obesity (ref. 9, ref. 12, ref. 13). Interestingly, a positive correlation to the BMI and body fat was found for the circulating levels of endocannabinoids, whereas obesity was linked to a reduced adipose expression of Cnr1 and FAAH (ref. 34, ref. 35, ref. 45). DEHP was found to interact with receptors of the ECS, namely CB1 and PPARs (ref. 48, ref. 49). The herein investigated receptors and enzymes of the ECS were not altered by DEHP. As shown before, DEHP did not affect the protein amount of PPARalpha and PPARgamma in SGBS (ref. 53). However, DEHP-mediated alterations on PPARs was reported in both directions as assessed in rodent in vitro and in vivo experiments (ref. 14, ref. 16, ref. 17, ref. 61), which points toward species-specific effects. Taken together, the absence of any effect on the expression of ECS enzymes by DEHP in the present study does not mean that DEHP has no impact on endocannabinoid metabolism. Further functional studies, for example, on enzyme activity will be needed to adequately address this aspect.
Summing up, in the human SGBS cell model an upregulation of the ECS receptors CNR1 and TRPV1 as well as the endocannabinoid-metabolizing enzymes FAAH and MAGL – presumably for reducing the endocannabinoid level in the differentiation process – was found during normal adipogenesis. As expected, the secretion of adiponectin and leptin was simultaneously increased. These data implicate the ECS to play a role in normal adipogenesis. As DEHP altered the level of adipokines secreted by mature adipocytes without affecting the intrinsic ECS, we conclude this DEHP-mediated endocrine impairment to be independent of the intrinsic ECS as endocrine modulator.
Declaration of interest
The authors declare that there is no conflict of interest that could be perceived as prejudicing the impartiality of the research reported.
Funding
Jana Ernst, Kristina Schädlich and Urszula Grabiec were supported by the Roux Programme of the Faculty of Medicine, Martin Luther University Halle-Wittenberg (JE FKZ 29/11, KS FKZ 26/06, UG FKZ 29/18). We acknowledge the financial support within the funding programme Open Access Publishing by the German Research Foundation (DFG).
Author contribution statement
JE performed acquisition, analysis and interpretation of data, conception and design of the study, drafted the article and approved the final manuscript. UG performed acquisition, analysis and interpretation of data and approved the final manuscript. KF performed acquisition, analysis and interpretation of data and approved the final manuscript. FD was a project leader and performed revision and approved the final manuscript. KS was a project leader and performed conception and design of the study and revision and approved the final manuscript.
References
- PF Baillie-Hamilton. Chemical toxins: a hypothesis to explain the global obesity epidemic.. Journal of Alternative and Complementary Medicine, 2002. [DOI | PubMed]
- F Grün. Obesogens.. Current Opinion in Endocrinology, Diabetes, and Obesity, 2010. [DOI]
- F Grün, B Blumberg. Endocrine disrupters as obesogens.. Molecular and Cellular Endocrinology, 2009. [DOI]
- W Holtcamp. Obesogens: an environmental link to obesity.. Environmental Health Perspectives, 2012. [DOI | PubMed]
- TT Schug, A Janesick, B Blumberg, JJ Heindel. Endocrine disrupting chemicals and disease susceptibility.. Journal of Steroid Biochemistry and Molecular Biology, 2011. [DOI | PubMed]
- S Benjamin, E Masai, N Kamimura, K Takahashi, RC Anderson, PA Faisal. Phthalates impact human health: epidemiological evidences and plausible mechanism of action.. Journal of Hazardous Materials, 2017. [DOI]
- U Heudorf, V Mersch-Sundermann, J Angerer. Phthalates: toxicology and exposure.. International Journal of Hygiene and Environmental Health, 2007. [DOI | PubMed]
- R Kavlock, D Barr, K Boekelheide, W Breslin, P Breysse, R Chapin, K Gaido, E Hodgson, M Marcus, K Shea. NTP-CERHR expert panel update on the reproductive and developmental toxicity of di(2-ethylhexyl) phthalate.. Reproductive Toxicology, 2006. [DOI | PubMed]
- MC Buser, HE Murray, F Scinicariello. Age and sex differences in childhood and adulthood obesity association with phthalates: analyses of NHANES 2007–2010.. International Journal of Hygiene and Environmental Health, 2014. [DOI | PubMed]
- TM James-Todd, T Huang, EW Seely, AR Saxena. The association between phthalates and metabolic syndrome: the National Health and Nutrition Examination Survey 2001–2010.. Environmental Health: A Global Access Science Source, 2016. [DOI | PubMed]
- JH Kim, HY Park, S Bae, YH Lim, YC Hong. Diethylhexyl phthalates is associated with insulin resistance via oxidative stress in the elderly: a panel study.. PLoS ONE, 2013. [DOI | PubMed]
- JH Kim, H Park, J Lee, G Cho, S Choi, G Choi, SY Kim, SH Eun, E Suh, SK Kim. Association of diethylhexyl phthalate with obesity-related markers and body mass change from birth to 3 months of age.. Journal of Epidemiology and Community Health, 2016. [DOI | PubMed]
- A Smerieri, C Testa, P Lazzeroni, F Nuti, E Grossi, S Cesari, L Montanini, G Latini, S Bernasconi, AM Papini. Di-(2-ethylhexyl) phthalate metabolites in urine show age-related changes and associations with adiposity and parameters of insulin sensitivity in childhood.. PLoS ONE, 2015. [DOI | PubMed]
- N Klöting, N Hesselbarth, M Gericke, A Kunath, R Biemann, R Chakaroun, J Kosacka, P Kovacs, M Kern, M Stumvoll. Di-(2-ethylhexyl)-phthalate (DEHP) causes impaired adipocyte function and alters serum metabolites.. PLoS ONE, 2015. [DOI | PubMed]
- Z Lv, J Cheng, S Huang, Y Zhang, S Wu, Y Qiu, Y Geng, Q Zhang, G Huang, Q Ma. DEHP induces obesity and hypothyroidism through both central and peripheral pathways in C3H/He mice.. Obesity, 2016. [DOI | PubMed]
- JS Schmidt, K Schaedlich, N Fiandanese, P Pocar, B Fischer. Effects of di(2-ethylhexyl) phthalate (DEHP) on female fertility and adipogenesis in C3H/N mice.. Environmental Health Perspectives, 2012. [DOI | PubMed]
- R Biemann, A Navarrete Santos, A Navarrete Santos, D Riemann, J Knelangen, M Blüher, H Koch, B Fischer. Endocrine disrupting chemicals affect the adipogenic differentiation of mesenchymal stem cells in distinct ontogenetic windows.. Biochemical and Biophysical Research Communications, 2012. [DOI]
- V Di Marzo. ‘Endocannabinoids’ and other fatty acid derivatives with cannabimimetic properties: biochemistry and possible physiopathological relevance.. Biochimica and Biophysica Acta, 1998. [DOI]
- LA Matsuda, SJ Lolait, MJ Brownstein, AC Young, TI Bonner. Structure of a cannabinoid receptor and functional expression of the cloned cDNA.. Nature, 1990. [DOI]
- S Munro, KL Thomas, M Abu-Shaar. Molecular characterization of a peripheral receptor for cannabinoids.. Nature, 1993. [DOI]
- RG Pertwee, RA Ross. Cannabinoid receptors and their ligands.. Prostaglandins, Leukotrienes, and Essential Fatty Acids, 2002. [DOI]
- RA Ross. The enigmatic pharmacology of GPR55.. Trends in Pharmacological Sciences, 2009. [DOI | PubMed]
- V Di Marzo, L de Petrocellis. Endocannabinoids as regulators of transient receptor potential (TRP) channels: a further opportunity to develop new endocannabinoid-based therapeutic drugs.. Current Medicinal Chemistry, 2010. [DOI | PubMed]
- M Pistis, M Melis. From surface to nuclear receptors: the endocannabinoid family extends its assets.. Current Medicinal Chemistry, 2010. [DOI | PubMed]
- Y Okamoto, K Tsuboi, N Ueda. Enzymatic formation of anandamide.. Vitamins and Hormones, 2009. [DOI]
- N Ueda, K Tsuboi, T Uyama, T Ohnishi. Biosynthesis and degradation of the endocannabinoid 2-arachidonoylglycerol.. BioFactors, 2011. [DOI]
- TP Dinh, D Carpenter, FM Leslie, TF Freund, I Katona, SL Sensi, S Kathuria, D Piomelli. Brain monoglyceride lipase participating in endocannabinoid inactivation.. PNAS, 2002. [DOI | PubMed]
- F Fezza, C de Simone, D Amadio, M Maccarrone. Fatty acid amide hydrolase: a gate-keeper of the endocannabinoid system.. Sub-Cellular Biochemistry, 2008. [DOI]
- I Forner-Piquer, F Maradonna, G Gioacchini, S Santangeli, M Allarà, F Piscitelli, HR Habibi, V Di Marzo, O Carnevali. Dose-specific effects of di-isononyl phthalate on the endocannabinoid system and on liver of female zebrafish.. Endocrinology, 2017. [DOI | PubMed]
- I Forner-Piquer, CC Mylonas, J Calduch-Giner, F Maradonna, G Gioacchini, M Allarà, F Piscitelli, V Di Marzo, J Pérez-Sánchez, O Carnevali. Endocrine disruptors in the diet of male Sparus aurata: modulation of the endocannabinoid system at the hepatic and central level by di-isononyl phthalate and bisphenol A.. Environment International, 2018. [DOI]
- I Forner-Piquer, S Santangeli, F Maradonna, A Rabbito, F Piscitelli, HR Habibi, V Di Marzo, O Carnevali. Disruption of the gonadal endocannabinoid system in zebrafish exposed to diisononyl phthalate.. Environmental Pollution, 2018. [DOI]
- V Pomatto, E Cottone, P Cocci, M Mozzicafreddo, G Mosconi, ER Nelson, FA Palermo, P Bovolin. Plasticizers used in food-contact materials affect adipogenesis in 3T3-L1 cells.. Journal of Steroid Biochemistry and Molecular Biology, 2018. [DOI | PubMed]
- MF Bennetzen, TS Nielsen, SK Paulsen, J Bendix, S Fisker, N Jessen, S Lund, B Richelsen, SB Pedersen. Reduced cannabinoid receptor 1 protein in subcutaneous adipose tissue of obese.. European Journal of Clinical Investigation, 2010. [DOI | PubMed]
- M Blüher, S Engeli, N Klöting, J Berndt, M Fasshauer, S Bátkai, P Pacher, MR Schön, J Jordan, M Stumvoll. Dysregulation of the peripheral and adipose tissue endocannabinoid system in human abdominal obesity.. Diabetes, 2006. [DOI | PubMed]
- S Engeli, J Böhnke, M Feldpausch, K Gorzelniak, J Janke, S Bátkai, P Pacher, J Harvey-White, FC Luft, AM Sharma. Activation of the peripheral endocannabinoid system in human obesity.. Diabetes, 2005. [DOI | PubMed]
- S Engeli, AC Lehmann, J Kaminski, V Haas, J Janke, AA Zoerner, FC Luft, D Tsikas, J Jordan. Influence of dietary fat intake on the endocannabinoid system in lean and obese subjects.. Obesity, 2014. [DOI | PubMed]
- K Kempf, J Hector, T Strate, B Schwarzloh, B Rose, C Herder, S Martin, P Algenstaedt. Immune-mediated activation of the endocannabinoid system in visceral adipose tissue in obesity.. Hormone and Metabolic Research, 2007. [DOI | PubMed]
- L Bellocchio, C Cervino, V Vicennati, R Pasquali, U Pagotto. Cannabinoid type 1 receptor: another arrow in the adipocytes’ bow.. Journal of Neuroendocrinology, 2008. [DOI | PubMed]
- M Gary-Bobo, G Elachouri, B Scatton, G Le Fur, F Oury-Donat, M Bensaid. The cannabinoid CB1 receptor antagonist Rimonabant (SR141716) inhibits cell proliferation and increases markers of adipocyte maturation in cultured mouse 3T3 F442A preadipocytes.. Molecular Pharmacology, 2006. [DOI | PubMed]
- V Gasperi, F Fezza, N Pasquariello, M Bari, S Oddi, AF Agrò, M Maccarrone. Endocannabinoids in adipocytes during differentiation and their role in glucose uptake.. Cellular and Molecular Life Sciences, 2007. [DOI | PubMed]
- C Pagano, C Pilon, A Calcagno, R Urbanet, M Rossato, G Milan, K Bianchi, R Rizzuto, P Bernante, G Federspil. The endogenous cannabinoid system stimulates glucose uptake in human fat cells via phosphatidylinositol 3-kinase and calcium-dependent mechanisms.. Journal of Clinical Endocrinology and Metabolism, 2007. [DOI | PubMed]
- J Tam, R Cinar, J Liu, G Godlewski, D Wesley, T Jourdan, G Szanda, B Mukhopadhyay, L Chedester, JS Liow. Peripheral cannabinoid-1 receptor inverse agonism reduces obesity by reversing leptin resistance.. Cell Metabolism, 2012. [DOI | PubMed]
- P Löfgren, E Sjölin, K Wåhlen, J Hoffstedt. Human adipose tissue cannabinoid receptor 1 gene expression is not related to fat cell function or adiponectin level.. Journal of Clinical Endocrinology and Metabolism, 2007. [DOI | PubMed]
- ZD Thornton-Jones, GA Kennett, KR Benwell, DF Revell, A Misra, DM Sellwood, SP Vickers, PG Clifton. The cannabinoid CB1 receptor inverse agonist, Rimonabant, modifies body weight and adiponectin function in diet-induced obese rats as a consequence of reduced food intake.. Pharmacology, Biochemistry, and Behavior, 2006. [DOI]
- I Matias, MP Gonthier, P Orlando, V Martiadis, L de Petrocellis, C Cervino, S Petrosino, L Hoareau, F Festy, R Pasquali. Regulation, function, and dysregulation of endocannabinoids in models of adipose and beta-pancreatic cells and in obesity and hyperglycemia.. Journal of Clinical Endocrinology and Metabolism, 2006. [DOI | PubMed]
- D Teixeira, D Pestana, A Faria, C Calhau, I Azevedo, R Monteiro. Modulation of adipocyte biology by δ(9)-tetrahydrocannabinol.. Obesity, 2010. [DOI | PubMed]
- S Ahn, S Yi, WJ Seo, MJ Lee, YK Song, SY Baek, J Yu, SH Hong, J Lee, DW Shin. A cannabinoid receptor agonist N-arachidonoyl dopamine inhibits adipocyte differentiation in human mesenchymal stem cells.. Biomolecules and Therapeutics, 2015. [DOI | PubMed]
- KM Bisset, AS Dhopeshwarkar, C Liao, RA Nicholson. The G protein-coupled cannabinoid-1 (CB1) receptor of mammalian brain: inhibition by phthalate esters in vitro.. Neurochemistry International, 2011. [DOI | PubMed]
- MK Sarath Josh, S Pradeep, KS Vijayalekshmi Amma, S Balachandran, UC Abdul Jaleel, M Doble, F Spener, S Benjamin. Phthalates efficiently bind to human peroxisome proliferator activated receptor and retinoid X receptor α, β, γ subtypes: an in silico approach.. Journal of Applied Toxicology, 2014. [DOI | PubMed]
- P Fischer-Posovszky, FS Newell, M Wabitsch, HE Tornqvist. Human SGBS cells – a unique tool for studies of human fat cell biology.. Obesity Facts, 2008. [DOI | PubMed]
- M Wabitsch, RE Brenner, I Melzner, M Braun, P Möller, E Heinze, KM Debatin, H Hauner. Characterization of a human preadipocyte cell strain with high capacity for adipose differentiation.. International Journal of Obesity and Related Metabolic Disorders, 2001. [DOI | PubMed]
- R Biemann, B Fischer, A Navarrete Santos. Adipogenic effects of a combination of the endocrine-disrupting compounds bisphenol A, diethylhexylphthalate, and tributyltin.. Obesity Facts, 2014. [DOI | PubMed]
- K Schaedlich, S Gebauer, L Hunger, LS Beier, HM Koch, M Wabitsch, B Fischer, J Ernst. DEHP deregulates adipokine levels and impairs fatty acid storage in human SGBS-adipocytes.. Scientific Reports, 2018. [DOI | PubMed]
- 54OECD. Health at a Glance 2011: OECD Indicators. Paris, France: OECD Publishing, 2011 (10.1787/health_glance-2011-en)
- R Kavlock, K Boekelheide, R Chapin, M Cunningham, E Faustman, P Foster, M Golub, R Henderson, I Hinberg, R Little. NTP Center for the Evaluation of Risks to Human Reproduction: phthalates expert panel report on the reproductive and developmental toxicity of di(2-ethylhexyl) phthalate.. Reproductive Toxicology, 2002. [DOI | PubMed]
- N Gillum, Z Karabekian, LM Swift, RP Brown, MW Kay, N Sarvazyan. Clinically relevant concentrations of di (2-ethylhexyl) phthalate (DEHP) uncouple cardiac syncytium.. Toxicology and Applied Pharmacology, 2009. [DOI]
- B Spoto, F Fezza, G Parlongo, N Battista, E Sgro’, V Gasperi, C Zoccali, M Maccarrone. Human adipose tissue binds and metabolizes the endocannabinoids anandamide and 2-arachidonoylglycerol.. Biochimie, 2006. [DOI | PubMed]
- R Roche, L Hoareau, S Bes-Houtmann, MP Gonthier, C Laborde, JF Baron, Y Haffaf, M Cesari, F Festy. Presence of the cannabinoid receptors, CB1 and CB2, in human omental and subcutaneous adipocytes.. Histochemistry and Cell Biology, 2006. [DOI | PubMed]
- H Hauner, K Röhrig, M Spelleken, LS Liu, J Eckel. Development of insulin-responsive glucose uptake and GLUT4 expression in differentiating human adipocyte precursor cells.. International Journal of Obesity and Related Metabolic Disorders, 1998. [DOI | PubMed]
- T Pederson, CM Rondinone. Regulation of proteins involved in insulin signaling pathways in differentiating human adipocytes.. Biochemical and Biophysical Research Communications, 2000. [DOI]
- Y Jia, T Liu, L Zhou, J Zhu, J Wu, D Sun, J Xu, Q Wang, H Chen, F Xu. Effects of di-(2-ethylhexyl) phthalate on lipid metabolism by the JAK/STAT pathway in rats.. International Journal of Environmental Research and Public Health, 2016. [DOI]