
Repeated Tetrahydrocannabinol Injections Induce Tolerance but Do Not Disrupt Ongoing Behaviour Upon Withdrawal in Male and Female Rats With Inflammatory Pain
Abstract
Background:
Many people use cannabis to self‐medicate for pain. Little is known about the impact of pain on tolerance and spontaneous withdrawal to delta‐9‐tetrahydrocannabinal (THC), the primary psychoactive compound in cannabis. Our previous research with the opioid morphine suggests persistent pain will increase the magnitude and duration of THC withdrawal.
Methods:
Male and female Sprague–Dawley rats were individually housed in a cage with a running wheel to provide a continuous and objective measure of well‐being. All rats were injected with Complete Freund’s Adjuvant (CFA) into the right hindpaw to induce inflammatory pain. Beginning 1 day later, rats received twice daily THC or vehicle injections for 7 days followed by assessment of tolerance and spontaneous withdrawal.
Results:
Administration of CFA decreased wheel running. Twice daily injections of THC (3 mg/kg/injection) caused a further reduction in running in male and female rats. Tolerance to the THC‐induced decrease in running was more pronounced in male compared to female rats. There was no evidence of spontaneous withdrawal to THC despite continuous assessment for 6 days. Likewise, withdrawal had no effect on body weight.
Conclusion:
The lack of spontaneous withdrawal in rats with hindpaw inflammation is consistent with our recent study showing a lack of spontaneous withdrawal to THC in pain free rats, but opposite to our opioid research showing enhancement of spontaneous withdrawal to morphine in rats with hindpaw inflammation. In sum, persistent inflammatory pain does not appear to alter the effects of repeated THC injections in male or female rats.
Significance:
The use of THC as a treatment for pain is limited by side effects and tolerance but not by dependence associated withdrawal symptoms.
Article type: Research Article
Keywords: cannabis, dependence, sex differences, wheel running
Affiliations: Washington State University Vancouver Vancouver Washington USA
License: © 2026 The Author(s). European Journal of Pain published by John Wiley & Sons Ltd on behalf of European Pain Federation ‐ EFIC ®. CC BY 4.0 This is an open access article under the terms of the http://creativecommons.org/licenses/by/4.0/ License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Article links: DOI: 10.1002/ejp.70279 | PubMed: 42028989 | PMC: PMC13108161
Relevance: Relevant: mentioned in keywords or abstract
Full text: PDF (737 KB)
Introduction
Pain is the primary medical reason people take cannabis (Sexton et al. ref. 2016; Walsh et al. ref. 2013). The antinociceptive effects of delta‐9‐tetrahydrocannabinol (THC), the primary psychoactive compound in cannabis, have also been documented in animal research (Blanton et al. ref. 2021; Martin and Lichtman ref. 1998). Although the antinociceptive effects of THC are mild compared to that produced by opioids (Lötsch et al. ref. 2018), the safety profile is much better. Tolerance and dependence develop rapidly to repeated opioid administration (Bailey and Connor ref. 2005; Bobeck et al. ref. 2012; Kandasamy et al. ref. 2017; Morgan and Bobeck ref. 2025), and opioid side effects and withdrawal are severe (Gowing et al. ref. 2017; Hill and Canals ref. 2022). Although tolerance and withdrawal to THC have been reported, almost nothing is known about tolerance and withdrawal when THC is administered to treat pain.
Tolerance to the antinociceptive effects of THC has been reported in male and female rats with (Craft et al. ref. 2023) and without persistent pain (Marusich et al. ref. 2015; Wakley et al. ref. 2014). Dependence also has been reported, but withdrawal symptoms are mild. Typical withdrawal symptoms in humans consist of irritability, anxiety and insomnia (Budney et al. ref. 2004; Chesney et al. ref. 2024). Precipitation of withdrawal in animals by administration of a cannabinoid receptor antagonist is associated with a range of motor behaviours (e.g., shakes, tremors, licking, arched back) (Aceto et al. ref. 1995; Bruijnzeel et al. ref. 2016; Marusich et al. ref. 2015; Tsou et al. ref. 1995). In addition, anxiety has been reported weeks after precipitated withdrawal in rats made dependent on the cannabinoid receptor agonist WIN55212‐2 (Brewer et al. ref. 2024).
The symptoms of spontaneous THC withdrawal in rats are less pronounced if they occur at all (Aroni et al. ref. 2024; Harte‐Hargrove and Dow‐Edwards ref. 2012). There are two major challenges in assessing spontaneous withdrawal. Are the symptoms the same as in precipitated withdrawal and when do those symptoms occur? Our recent manuscript addressed these questions by assessing the impact of withdrawal on voluntary home cage wheel running for 6 days following termination of THC administration (Hickey et al. ref. 2025). That study found no obvious signs of spontaneous withdrawal in pain‐free male and female rats.
Previous research with opioids shows clear changes in voluntary wheel running during spontaneous withdrawal. Spontaneous withdrawal from morphine or fentanyl causes a decrease in dark phase wheel running that lasts approximately 2–3 days (Morgan, Hilgendorf, and Kandasamy ref. 2023; Morgan and Ataras ref. 2022) and an increase in light phase running that corresponds with insomnia (Morgan et al. ref. 2021; Morgan and Nguyen ref. 2024). The magnitude and duration of the opioid withdrawal‐induced decrease in running were enhanced in rats with inflammatory pain (Kandasamy et al. ref. 2017). The objective of the present experiment was to test the hypothesis that inflammatory pain will have a similar effect on spontaneous withdrawal to THC, essentially producing withdrawal symptoms where none were evident in our previous experiment using pain‐free rats (Hickey et al. ref. 2025).
Methods
Animals
A total of 32 adult Sprague–Dawley rats (Envigo Laboratories, California), 16 male and 16 female, were used. Rats were housed in same‐sex pairs for 1 week upon arrival and then housed individually in a standard Plexiglas cage fitted with a top‐mounted stainless steel running wheel (Tecniplast, Starr Life Sciences Corp; Pennsylvania). Animals were maintained on a 12:12‐h light–dark cycle (lights off at 10:30 or 11:00 AM depending on the cohort test room) in a temperature and humidity‐controlled room. Food and water were available ad libitum. Cages were changed once per week. This experiment was approved by the Washington State University (WSU) Institutional Animal Care and Use Committee (IACUC) and was conducted in a manner consistent with ARRIVE Guidelines and the Guide for the Care and Use of Laboratory Animals (ref. 2011).
Wheel Running
Rats had continuous access to a running wheel in the home cage. The wheel had a diameter of 33 cm so each revolution was approximately 1 m. Data were collected in 5‐min bins 24 h a day throughout the experiment. Rats were only removed from the cage for daily body weight measurements, injections and for the weekly cage change. These things occurred just prior to the beginning of the dark phase of the circadian cycle. The only exception was administration of a second injection 6 h into the dark phase for 7 days. No data were analysed during the 2 h preceding the light phase to avoid confounds produced by handling the rats or anticipation of handling.
Experimental Design
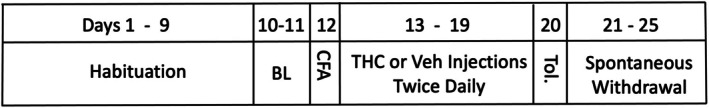
Rats were allowed to habituate to the running wheel cage for 11 days. The average of the data from the last 2 days was used as the baseline value. Rats were injected with CFA into the right hindpaw immediately prior to the beginning of the dark phase on Day 12. Beginning on Day 13, rats were injected twice daily with THC (3 mg/kg/injection, s.c.) or vehicle (saline:ethanol:cremophor, 18:1:1) for seven consecutive days. Injections were administered 5–10 min prior to the dark phase and again 6 h later. THC was diluted with the vehicle from a solution provided as a gift from the NIDA Drug Supply Program. Tolerance was assessed by injecting every rat with THC (3 mg/kg, s.c.) on the day following the last THC/vehicle injections (Morgan and Bobeck ref. 2025) and measuring the impact on wheel running. Spontaneous withdrawal was assessed continuously starting 1‐day after the tolerance test (Day 21) and ending 6 days after the last THC injection (Day 25). Rats were euthanized with isoflurane following the final day of withdrawal assessment. The timeline is presented in Figure 1.
Statistics
The experiment employed a 2 × 2 factorial design with sex and THC administration as between‐subjects factors (8 rats/condition). Three aspects of wheel running were analysed: (1) Total wheel revolutions during the 12 h dark phase; (2) Total wheel revolutions during the first 10 h of the light phase; and (3) Maximum running speed. Maximum running speed was defined as the highest number of wheel revolutions during a 5‐min bin. Wheel running and body weight data were analysed with 2‐ and 3‐way ANOVAs (Sex × THC × Day) using SPSS. Analysis of sex differences in dark phase wheel running, maximum running speed and body weight required converting data to a percent of baseline because of significant differences in baseline values (Table 1). Light phase running was too low during the baseline period to accurately convert data to a percent of baseline. Data were only collected for the first 10 h of the 12‐h light phase because running levels increased dramatically during the last 2 h of the light phase and data collection was disrupted during the last hour to provide animal care, assess body weight and administer injections. Data were analysed separately for each section of the experiment: Baseline, CFA administration, THC administration, THC tolerance test and spontaneous withdrawal. Two female rats from each group were not tested on the tolerance test so the sample size for that part of the experiment was N = 6 instead of 8. Statistical significance was defined as a probability of < 5%.
TABLE 1: Sex differences in baseline data (Mean ± SEM).
| Body weight (g) | Dark running | Light running | Maximum speed | |
|---|---|---|---|---|
| Female | 185 ± 3.6 | 7230 ± 986 | 956 ± 253 | 208 ± 20 |
| Male | 265 ± 3.0 | 2311 ± 407 | 289 ± 99 | 124 ± 11 |
Note: Running is presented as number of wheel revolutions. Maximum speed is the number of revolutions in 5 min.
Results
Baseline Data
Male and female rats differed on all baseline measures (Table 1). Male rats were significantly heavier than female rats on the last day of the habituation period (t 30 = 16.979, p < 0.001). Female rats ran significantly further than male rats during the dark (t 30 = 4.611, p < 0.001) and light (t 30 = 2.453, p = 0.02) phases of the circadian cycle. In both cases, average male running was approximately 30% of the running in female rats. Female rats also attained a higher maximal speed compared to male rats (t 30 = 3.730, p < 0.001).
Hindpaw Inflammation
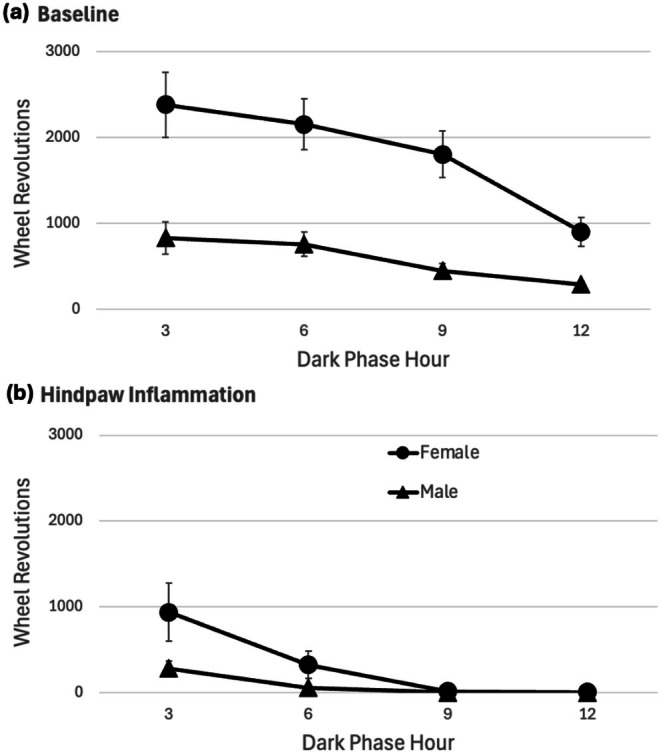
A mean decrease of 10.2 g of body weight for male rats and 2.6 g for female rats occurred 24 h after the CFA injection. Comparison of this weight loss as a percent of baseline revealed that male rats lost significantly more weight than female rats (96.2% vs. 98.5% of baseline weight, respectively) (t 30 = 3.634, p < 0.001). Administration of CFA also caused a profound decrease in wheel running. In the 12‐h dark phase following CFA administration, male rats ran 15.1% ± 3.4% of their baseline value and female rats ran 21.6% ± 5.8% of their baseline value. These decreases in running from baseline to CFA administration resulted in a significant main effect of Day (F(1,29) = 49.368, p < 0.001). There was no sex difference in this decrease when analysed as a percent change from baseline (t 29 = 0.959, p = 0.346). In contrast to the gradual decrease in running that occurs across the dark phase of the circadian cycle during baseline (Figure 2a), wheel running following CFA administration was greatly reduced within the first 3 h and was almost completely abolished during the remainder of the dark phase (Figure 2b). Most rats injected with CFA did not use the running wheel at all during the last 6 h of the dark phase or the subsequent light phase. None of the male rats achieved more than nine revolutions during the entire 10 h light phase measurement following the CFA injection, and 11 of the 16 female rats ran nine revolutions or less during the first 10 h of the light phase.
THC Injections
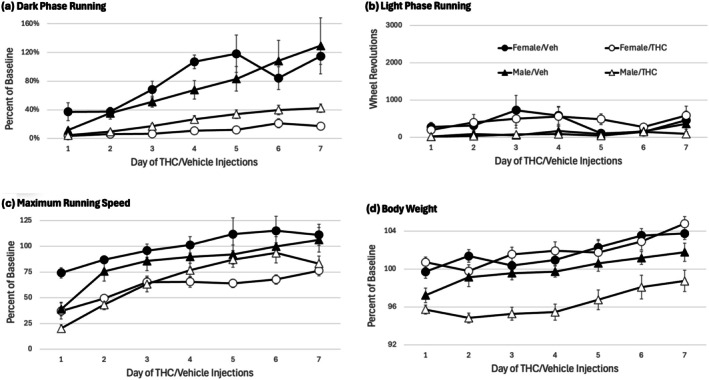
Twice daily THC or vehicle injections began 24 h after the CFA injection. Administration of THC caused a significant decrease in wheel running during the dark phase (Figure 3a) compared to vehicle treated rats (F(1,28) = 51.645, p < 0.001). Running increased each day as rats recovered from the CFA injection, but this increase was significantly greater in vehicle compared to THC treated rats (THC × Day interaction: F(6,168) = 5.797, p < 0.001). There was no significant difference in dark phase running between male and female rats (F(1,28) = 0.019, p = 0.891). Light phase running was very low for the entire 7‐day injection period (Figure 3b). Administration of THC also caused a significant reduction in maximum running speed (Figure 3c) compared to vehicle treated rats (F(1,28) = 21.585, p < 0.001). This reduction in running speed was comparable in male and female rats as indicated by the lack of a significant effect of Sex (F(1,28) = 0.61, p = 0.441). Repeated THC administration also caused a significant reduction in body weight (Figure 3d) in male, but not female rats as indicated by a significant THC × Sex interaction (F(1,28) =7.601. p = 0.01).
THC Tolerance
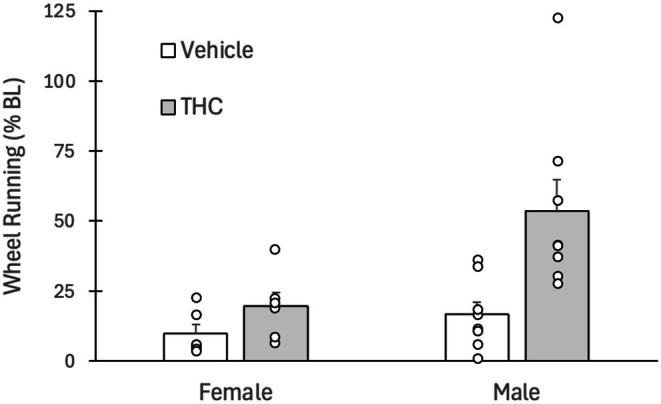
Tolerance was assessed by injecting all rats with 3 mg/kg of THC on the day after the last of the seven twice daily THC/vehicle injections. Administration of THC caused a large decrease in dark phase wheel running in vehicle rats receiving THC for the first time (Figure 4). A decrease in running also occurred in rats that had received twice daily injections of THC for 7 days compared to their baseline levels of running. However, administration of THC produced a significantly greater decrease in wheel running in the naïve vehicle rats compared to the THC experienced rats (F(1,24) = 9.988, p = 0.004). Moreover, male rats displayed greater tolerance than female rats as indicated by a significant main effect of Sex (F(1,24) = 7.757, p = 0.01).
THC Withdrawal
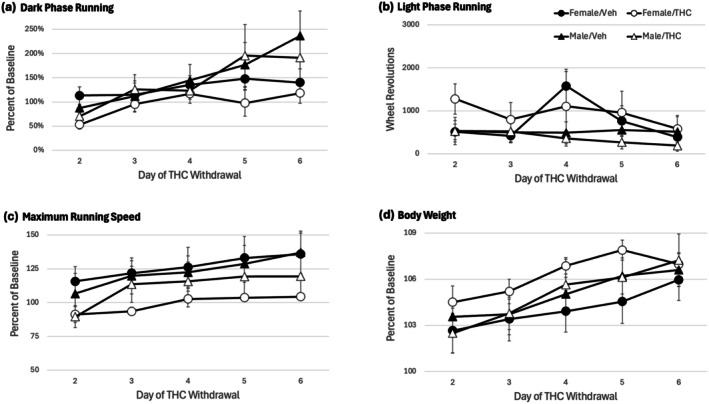
Withdrawal was assessed for 6 days following the THC injection for the tolerance test. The tolerance test (Figure 4) shows the data for the 12‐h dark phase immediately following this last THC injection. Figure 5 shows withdrawal Days 2–6. No change in dark phase wheel running is evident in rats withdrawn from THC compared to vehicle (Figure 5a) (F(1,28) = 0.764, p = 0.390). Although there was a significant increase in dark phase running across days (F(4,112) = 12.308, p < 0.001), this increase was not related to prior THC treatment as evident by the lack of a significant Day × THC interaction (F(4,112) = 0.509, p = 0.729). Likewise, there was no significant effect of THC withdrawal on light phase running (Figure 5b; F(1,28) = 0.014, p = 0.906), maximum running speed (Figure 5c; F(1,28) = 2.920, p = 0.099), or body weight (Figure 5d; F(1,24) = 1.106, p = 0.303).
Discussion
The two primary findings of this manuscript are that administration of 3 mg/kg of THC causes a decrease in wheel running and repeated THC administration produces greater tolerance to the decrease in wheel running in male compared to female rats. There was no evidence for spontaneous withdrawal measured as a change in wheel running or body weight. These data are almost identical to the results of our previous analysis of tolerance and withdrawal to THC in pain‐free rats and indicate that pain does not alter tolerance or dependence to THC (Hickey et al. ref. 2025).
The finding that baseline voluntary wheel running is greater in female compared to male rats is well‐known (Carrera et al. ref. 2011; Titchenal ref. 1988). The present study shows that male Sprague–Dawley rats run about 30% as much as female rats during both the dark and light phases of the circadian cycle. Likewise, the large decrease in wheel running caused by injection of CFA into the hindpaw has been reported previously (Kandasamy et al. ref. 2016). The present study shows a near complete inhibition of wheel running beginning approximately 6 h after the injection and persisting through the light phase before rats begin the gradual day‐by‐day recovery of running.
Antinociceptive effects of THC have been shown to occur using acute pain tests whether rats have inflammatory pain (Britch et al. ref. 2020; Craft et al. ref. 2023) or not (Lichtman and Martin ref. 1991; Tseng and Craft ref. 2001). However, caution must be used in interpreting these results because many of these studies use large doses of THC (≥ 3 mg/kg) known to inhibit movement (Wakley and Craft ref. 2011; Wiley and Burston ref. 2014). When nociception is assessed using pain‐depressed tests such as wheel running (Kandasamy and Morgan ref. 2021), relatively mild antinociceptive effects occur and only at a dose of 0.32 mg/kg (Kandasamy et al. ref. 2018; Morgan, Stickney, and Wilson‐Poe ref. 2023). Given that THC is known to reduce inflammatory pain (Britch et al. ref. 2020; Craft et al. ref. 2013, ref. 2023), there is little doubt the 3 mg/kg dose used in the present study produced antinociception, but this antinociception did not restore wheel running because of the direct inhibitory effect on locomotion. Our previous research shows that increasing the dose of THC from 0.32 mg/kg to 0.56 or 1 mg/kg is enough to depress wheel running (Kandasamy et al. ref. 2018; Morgan, Stickney, and Wilson‐Poe ref. 2023). A major advantage of pain‐depressed (e.g., wheel running) as opposed to pain‐evoked (e.g., von Frey, hot‐plate) tests is the ability to distinguish between doses that produce antinociception independent of disruptive side effects.
The decrease in wheel running caused by THC administration in the present experiment is consistent with our previous research showing a pronounced and prolonged decrease in wheel running in pain‐free rats (Hickey et al. ref. 2025). Although wheel running was already reduced by hindpaw inflammation in the present study, administration of THC caused a further decrease in running. The decrease in running caused by THC was comparable in male and female rats and consistent with previous research in which locomotion was assessed in an open field or the rat’s home cage (Britch and Craft ref. 2023; Harte‐Hargrove and Dow‐Edwards ref. 2012; Wakley and Craft ref. 2011). Moreover, THC‐induced suppression of locomotion occurs at doses significantly lower than needed to induce catalepsy (Wakley and Craft ref. 2011).
Tolerance to the antinociceptive effects of THC has been documented in both human (Cuttler et al. ref. 2020) and animal (Craft et al. ref. 2023; Wakley et al. ref. 2014) research. Animal research tends to focus on tolerance to the antinociceptive effects of THC. These studies reveal greater tolerance in pain‐free female compared to male rats (Wakley et al. ref. 2014). Tolerance to the antinociceptive effect of THC was less pronounced in rats with hindpaw inflammation and no sex difference was evident in these rats (Britch et al. ref. 2020; Craft et al. ref. 2023). Likewise, there does not appear to be a sex difference in tolerance to THC‐induced locomotor suppression when measured with wheel running (Hickey et al. ref. 2025). The only effect of hindpaw inflammation was to limit the development of tolerance to locomotor suppression in female rats. The present data show that the one situation where tolerance may be greater in male than female rats is suppression of wheel running in rats with hindpaw inflammation. Although the smaller sample size in female rats tested for tolerance (N = 6 vs. N = 8 for males) makes the female data less reliable than the male data, these sample sizes were sufficient to result in statistically significant sex difference in tolerance.
The THC injection for the tolerance test decreased wheel running for the entire day so assessment of withdrawal‐induced changes in running began 24 h after this injection. Although withdrawal was assessed continuously using wheel running from the second to sixth day following the last THC injection, no significant changes were evident. Female rats undergoing withdrawal tended to have a lower maximal running speed than female controls or male rats, but that was the only potential change in wheel running. In contrast, withdrawal following twice daily injections of the opioid morphine for 5 days caused a large decrease in dark phase wheel running and an increase in light phase running in male and female rats (Morgan et al. ref. 2021; Morgan and Nguyen ref. 2024). Moreover, hindpaw inflammation enhanced the magnitude and duration of the decrease in wheel running in male rats undergoing morphine withdrawal (Kandasamy et al. ref. 2017). The morphine data show that wheel running is a sensitive measure of withdrawal.
The present data indicate that spontaneous withdrawal from THC is mild if evident at all following twice daily injections for 7 days. Previous studies in rats also failed to find statistically significant withdrawal symptoms following discontinuation of much larger doses of THC (> 10 mg/kg/day) (Aceto et al. ref. 1996; Aroni et al. ref. 2024), although mild symptoms have been reported in mice (Kesner et al. ref. 2022; Trexler et al. ref. 2018, ref. 2019). Our data assessing withdrawal continuously using home cage wheel running is consistent with this lack of obvious signs of withdrawal and shows that the lack of withdrawal is not caused by assessment at the wrong time.
In contrast, precipitation of THC withdrawal by administration of a cannabinoid receptor antagonist such as rimonabant produces clear withdrawal symptoms in rats (Aceto et al. ref. 1995; Bruijnzeel et al. ref. 2016; Marusich et al. ref. 2015; Tsou et al. ref. 1995). The problem is that the clinical relevance of studies in which withdrawal is precipitated can be questioned because humans using cannabis undergo spontaneous, not precipitated, withdrawal. Our previous study in pain‐free rats also failed to find consistent changes in wheel running following spontaneous withdrawal from THC (Hickey et al. ref. 2025). The present results extend this finding by showing that even when THC is used to treat pain, spontaneous withdrawal symptoms do not seem to be a problem. The problem of tolerance and dependence is even less likely with a lower antinociceptive dose. The 3 mg/kg dose was used in the present study to ensure a behavioural effect but is 10 times greater than that needed to produce antinociception in rats (Kandasamy et al. ref. 2018; Morgan, Stickney, and Wilson‐Poe ref. 2023).
Author Contributions
This experiment was conceived and designed by C.M.H. and M.M.M. Data collection was overseen by C.M.H. Data were compiled and analysed by M.M.M. The manuscript was written by M.M.M. and edited by C.M.H. Both authors have approved the final version of the manuscript and agree to be accountable for all aspects of the work.
Funding
Washington State Alcohol and Drug Abuse Research Program.
Disclosure
No AI tool is generated.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Cannabinoid Precipitated Withdrawal by the Selective Cannabinoid Receptor Antagonist, SR 141716A.”. European Journal of Pharmacology, 1995. [PubMed]
- Dependence on Delta 9‐Tetrahydrocannabinol: Studies on Precipitated and Abrupt Withdrawal.”. Journal of Pharmacology and Experimental Therapeutics, 1996. [PubMed]
- Functional Adaptation in the Brain Habenulo‐Mesencephalic Pathway During Cannabinoid Withdrawal.”. Cells, 2024. [PubMed]
- Opioids: Cellular Mechanisms of Tolerance and Physical Dependence.”. Current Opinion in Pharmacology, 2005. [PubMed]
- Sex Differences and the Endocannabinoid System in Pain.”. Pharmacology, Biochemistry, and Behavior, 2021. [PubMed]
- Differential Development of Antinociceptive Tolerance to Morphine and Fentanyl Is not Linked to Efficacy in the Ventrolateral Periaqueductal Gray of the Rat.”. Journal of Pain, 2012. [PubMed]
- Somatic and Anxiety‐Like Behaviors in Male and Female Rats During Withdrawal From the Non‐Selective Cannabinoid Agonist WIN 55,212‐2.”. Pharmacology, Biochemistry, and Behavior, 2024. [PubMed]
- Cannabidiol and Delta‐9‐Tetrahydrocannabinol Interactions in Male and Female Rats With Persistent Inflammatory Pain.”. Journal of Pain, 2023. [PubMed]
- Antinociceptive and Immune Effects of Delta‐9‐Tetrahydrocannabinol or Cannabidiol in Male Versus Female Rats With Persistent Inflammatory Pain.”. Journal of Pharmacology and Experimental Therapeutics, 2020. [PubMed]
- Behavioral Characterization of the Effects of Cannabis Smoke and Anandamide in Rats.”. PLoS One, 2016. [PubMed]
- Review of the Validity and Significance of Cannabis Withdrawal Syndrome.”. American Journal of Psychiatry, 2004. [PubMed]
- Gender Dimorphic Effects of Voluntary Running in Laboratory Rats Depends on Maturational Status.”. Quarterly Journal of Experimental Psychology, 2011
- Psychosis Associated With Cannabis Withdrawal: Systematic Review and Case Series.”. British Journal of Psychiatry, 2024
- Vaporized Cannabis Extract‐Induced Antinociception in Male vs Female Rats With Persistent Inflammatory Pain.”. Pain, 2023. [PubMed]
- Sex Differences in Anti‐Allodynic, Anti‐Hyperalgesic and Anti‐Edema Effects of Δ(9)‐Tetrahydrocannabinol in the Rat.”. Pain, 2013. [PubMed]
- Short‐ and Long‐Term Effects of Cannabis on Headache and Migraine.”. Journal of Pain, 2020. [PubMed]
- Opioid Antagonists With Minimal Sedation for Opioid Withdrawal.”. Cochrane Database of Systematic Reviews, 2017. [PubMed]
- 2011. Guide for the Care and Use of Laboratory Animals. National Academies Press.
- Withdrawal From THC During Adolescence: Sex Differences in Locomotor Activity and Anxiety.”. Behavioural Brain Research, 2012. [PubMed]
- Tolerance but no Spontaneous Withdrawal Following Repeated THC Injections in Male and Female Rats.”. Cannabis and Cannabinoid Research, 2025. [PubMed]
- Experimental Considerations for the Assessment of In Vivo and In Vitro Opioid Pharmacology.”. Pharmacology & Therapeutics, 2022. [PubMed]
- Home Cage Wheel Running Is an Objective and Clinically Relevant Method to Assess Inflammatory Pain in Male and Female Rats.”. Journal of Neuroscience Methods, 2016. [PubMed]
- Anti‐Migraine Effect of ∆9‐Tetrahydrocannabinol in the Female Rat.”. European Journal of Pharmacology, 2018. [PubMed]
- Depression of Home Cage Wheel Running Is an Objective Measure of Spontaneous Morphine Withdrawal in Rats With and Without Persistent Pain.”. Pharmacology, Biochemistry, and Behavior, 2017. [PubMed]
- “Reinventing the Wheel” to Advance the Development of Pain Therapeutics.”. Behavioural Pharmacology, 2021. [PubMed]
- Changes in Striatal Dopamine Release, Sleep, and Behavior During Spontaneous Δ‐9‐Tetrahydrocannabinol Abstinence in Male and Female Mice.”. Neuropsychopharmacology, 2022. [PubMed]
- Spinal and Supraspinal Components of Cannabinoid‐Induced Antinociception.”. Journal of Pharmacology and Experimental Therapeutics, 1991. [PubMed]
- Current Evidence of Cannabinoid‐Based Analgesia Obtained in Preclinical and Human Experimental Settings.”. European Journal of Pain, 2018. [PubMed]
- Cannabinoid Transmission and Pain Perception.”. Neurobiology of Disease, 1998. [PubMed]
- The Impact of Gonadal Hormones on Cannabinoid Dependence.”. Experimental and Clinical Psychopharmacology, 2015. [PubMed]
- Sex Differences in the Impact of Pain, Morphine Administration and Morphine Withdrawal on Quality of Life in Rats.”. Pharmacology, Biochemistry, and Behavior, 2022. [PubMed]
- Narrative Review of the Scientific and Methodological Explanations for 101 Mechanisms of Opioid Tolerance.”. Pain Reports, 2025. [PubMed]
- Continuous Fentanyl Administration and Spontaneous Withdrawal Decreases Home Cage Wheel Running in Rats With and Without Hindpaw Inflammation.”. Physiology and Behavior, 2023. [PubMed]
- Diurnal Sex Differences in Morphine Withdrawal Revealed by Continuous Assessment of Voluntary Home Cage Wheel Running in the Rat.”. Behavioural Brain Research, 2024. [PubMed]
- Use of Home Cage Wheel Running to Assess the Behavioural Effects of Administering a Mu/Delta Opioid Receptor Heterodimer Antagonist for Spontaneous Morphine Withdrawal in the Rat.”. Behavioural Brain Research, 2021. [PubMed]
- Low‐Dose Δ9‐THC Produces Antinociception in Female, but Not Male Rats.”. Cannabis and Cannabinoid Research, 2023. [PubMed]
- A Cross‐Sectional Survey of Medical Cannabis Users: Patterns of Use and Perceived Efficacy.”. Cannabis and Cannabinoid Research, 2016. [PubMed]
- Exercise and Food Intake. What Is the Relationship?”. Sports Medicine, 1988. [PubMed]
- CB1 Positive Allosteric Modulation Attenuates Δ9‐THC Withdrawal and NSAID‐Induced Gastric Inflammation.”. Pharmacology, Biochemistry, and Behavior, 2019. [PubMed]
- Novel Behavioral Assays of Spontaneous and Precipitated THC Withdrawal in Mice.”. Drug and Alcohol Dependence, 2018. [PubMed]
- Sex Differences in Antinociceptive and Motoric Effects of Cannabinoids.”. European Journal of Pharmacology, 2001. [PubMed]
- Physical Withdrawal in Rats Tolerant to Delta 9‐Tetrahydrocannabinol Precipitated by a Cannabinoid Receptor Antagonist.”. European Journal of Pharmacology, 1995. [PubMed]
- THC‐Methadone and THC‐Naltrexone Interactions on Discrimination, Antinociception, and Locomotion in Rats.”. Behavioural Pharmacology, 2011. [PubMed]
- Sex Differences in Antinociceptive Tolerance to Delta‐9‐Tetrahydrocannabinol in the Rat.”. Drug and Alcohol Dependence, 2014. [PubMed]
- Cannabis for Therapeutic Purposes: Patient Characteristics, Access, and Reasons for Use.”. International Journal on Drug Policy, 2013. [PubMed]
- Sex Differences in Δ(9)‐Tetrahydrocannabinol Metabolism and In Vivo Pharmacology Following Acute and Repeated Dosing in Adolescent Rats.”. Neuroscience Letters, 2014. [PubMed]