
Early 5‐HT 6 receptor blockade prevents symptom onset in a model of adolescent cannabis abuse
Abstract
Cannabis abuse during adolescence confers an increased risk for developing later in life cognitive deficits reminiscent of those observed in schizophrenia, suggesting common pathological mechanisms that remain poorly characterized. In line with previous findings that revealed a role of 5‐HT 6 receptor‐operated mTOR activation in cognitive deficits of rodent developmental models of schizophrenia, we show that chronic administration of ∆9‐tetrahydrocannabinol (THC) to mice during adolescence induces a long‐lasting activation of mTOR in prefrontal cortex (PFC), alterations of excitatory/inhibitory balance, intrinsic properties of layer V pyramidal neurons, and long‐term depression, as well as cognitive deficits in adulthood. All are prevented by administrating a 5‐HT 6 receptor antagonist or rapamycin, during adolescence. In contrast, they are still present 2 weeks after the same treatments delivered at the adult stage. Collectively, these findings suggest a role of 5‐HT 6 receptor‐operated mTOR signaling in abnormalities of cortical network wiring elicited by THC at a critical period of PFC maturation and highlight the potential of 5‐HT 6 receptor antagonists as early therapy to prevent cognitive symptom onset in adolescent cannabis abusers.
Article type: Research Article
Keywords: 5, adolescent cannabis abusers, cognitive deficits, synaptic transmission, Chemical Biology, Development & Differentiation, Neuroscience
Affiliations: IGF, University of Montpellier CNRS, INSERM Montpellier France; Department of Adult Psychiatry Montpellier University Hospital Montpellier France; Department of Medicinal Chemistry Jagiellonian University Medical College Kraków Poland; Neuropharmacology Laboratory Department of Experimental and Health Sciences Pompeu Fabra University Barcelona Spain
License: © 2020 The Authors. Published under the terms of the CC BY 4.0 license CC BY 4.0 This is an open access article under the terms of the http://creativecommons.org/licenses/by/4.0/ License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Article links: DOI: 10.15252/emmm.201910605 | PubMed: 32329240 | PMC: PMC7207164
Relevance: Relevant: mentioned in keywords or abstract
Full text: PDF (76 KB)
Introduction
Cannabis is the most commonly used recreational drug worldwide, and the last 30 years have been marked by a dramatic increase in cannabis consumption at an increasingly early age by young people in most developed countries (Hall & Babor, 2000). Epidemiological studies suggest that cannabis abuse during adolescence confers an increased risk for developing later in life psychotic symptoms and neurocognitive alterations reminiscent of those observed in schizophrenia (D’Souza et al, 2009; Evins et al, 2012). Moreover, neuroimaging studies in adolescent cannabis users revealed structural abnormalities and altered neural activity in the prefrontal cortex (PFC) during resting state and several types of cognitive paradigms, suggesting a critical role of this brain region in the core cognitive symptoms associated with cannabis abuse during adolescence (Schweinsburg et al, 2008; Batalla et al, 2013). Likewise, chronic administration of Δ9‐tetrahydrocannabinol (THC), the main psychoactive constituent of cannabis, to adolescent rats, induces behavioral alterations and cognitive deficits in adulthood that reflect, at least in part, PFC dysfunction (Schneider & Koch, 2003; O’Shea et al, 2006; Malone et al, 2010; Renard et al, 2013, 2017; Rubino & Parolaro, 2013, 2016; Zamberletti et al, 2014).
The PFC undergoes highly orchestrated maturation processes during adolescence. These include sprouting and pruning of synapses, refinement of circuit connectivity and of various neurotransmitter systems, such as glutamatergic, GABAergic, dopaminergic, and endocannabinoid systems (Spear, 2000; Rubino & Parolaro, 2016). It is conceivable that any interference with these maturational events such as chronic cannabis consumption during adolescence would lead to irreversible alterations of PFC connectivity and functionality that represent a risk factor for neuropsychiatric disorders in adulthood (Arseneault et al, 2004; Stefanis et al, 2004; Renard et al, 2014). Consistent with this hypothesis and underscoring the vulnerability of the adolescent brain to cannabis exposure, chronic administration of THC to adult rats (at similar doses to those injected in adolescent animals) does not reproduce the long‐lasting cognitive deficits observed in THC‐injected adolescent animals (Spear, 2000; Rubino & Parolaro, 2013).
Among the targets currently under investigation to alleviate cognitive deficits associated with various neuropsychiatric disorders, including schizophrenia, the serotonin 5‐HT6 receptor still raises particular interest in view of its high expression level in brain regions involved in mnemonic functions and the pro‐cognitive effects of 5‐HT6 receptor blockade in a broad range of cognitive paradigms in rodents (Codony et al, 2011; Yun & Rhim, 2011). In an effort to identify signaling mechanisms underlying cognition control by the 5‐HT6 receptor, we previously demonstrated that receptor‐operated activation of mechanistic Target Of Rapamycin (mTOR) in PFC underlies cognitive deficits in two rat developmental models of schizophrenia, namely neonatal phencyclidine administration and rearing in social isolation after weaning (Meffre et al, 2012). These results are consistent with the role of non‐physiological mTOR activation in cognitive impairment observed in genetic forms of autism spectrum disorders (ASD) and Down’s syndrome (Ehninger et al, 2008; Ehninger & Silva, 2011; Troca‐Marin et al, 2012). Likewise, a previous study has shown that acute THC administration to adult mice induces an activation of mTOR that underlies the associated amnesic‐like effects (Puighermanal et al, 2009).
In addition to its well‐described role in cognition, the 5‐HT6 receptor has recently emerged as a key regulator of neurodevelopmental processes such as neuronal migration (Dayer et al, 2015), neurite growth (Duhr et al, 2014), and dendritic protrusion (Rahman et al, 2017). Though the role of mTOR, under the control of 5‐HT6 receptor, in these neurodevelopmental mechanisms remains to be established, a large body of evidence indicates that the proper structural and functional development of brain circuitry depends on the fine tuning of mTOR signaling that has also a key influence upon synaptic transmission and synaptic plasticity in the mature brain (Swiech et al, 2008; Kim et al, 2009; Bockaert & Marin, 2015).
In light of these findings, we hypothesized that chronic cannabis abuse during adolescence might induce a persistent non‐physiological activation of mTOR in adolescent brain that might lead to abnormalities in PFC maturation and cognitive impairment at the adult stage. The implication of 5‐HT6 receptors in these deficits and the underlying alterations of synaptic transmission remain to be established, an issue we have explored in the present study, using mice injected daily with THC between post‐natal days (PNDs) 30 and 45 as a model of cannabis abuse during adolescence. We show that this treatment induces a sustained activation of mTOR signaling in PFC, an alteration of both GABAergic and glutamatergic synaptic transmissions and impairment of LTD in the PFC at layer I/V synapses and deficits in novel object recognition, sociability, and social discrimination in adulthood, which are all prevented by early administration (during adolescence) of the 5‐HT6 receptor antagonist SB258585 or the mTOR inhibitor rapamycin. Additional experiments were undertaken to characterize THC‐induced modifications of intrinsic properties of layer V pyramidal neurons and whether they can be prevented by blocking 5‐HT6 receptor‐operated mTOR signaling during adolescence.
Results
5‐HT6 receptors mediate delayed mTOR activation elicited by THC administration to adolescent mice
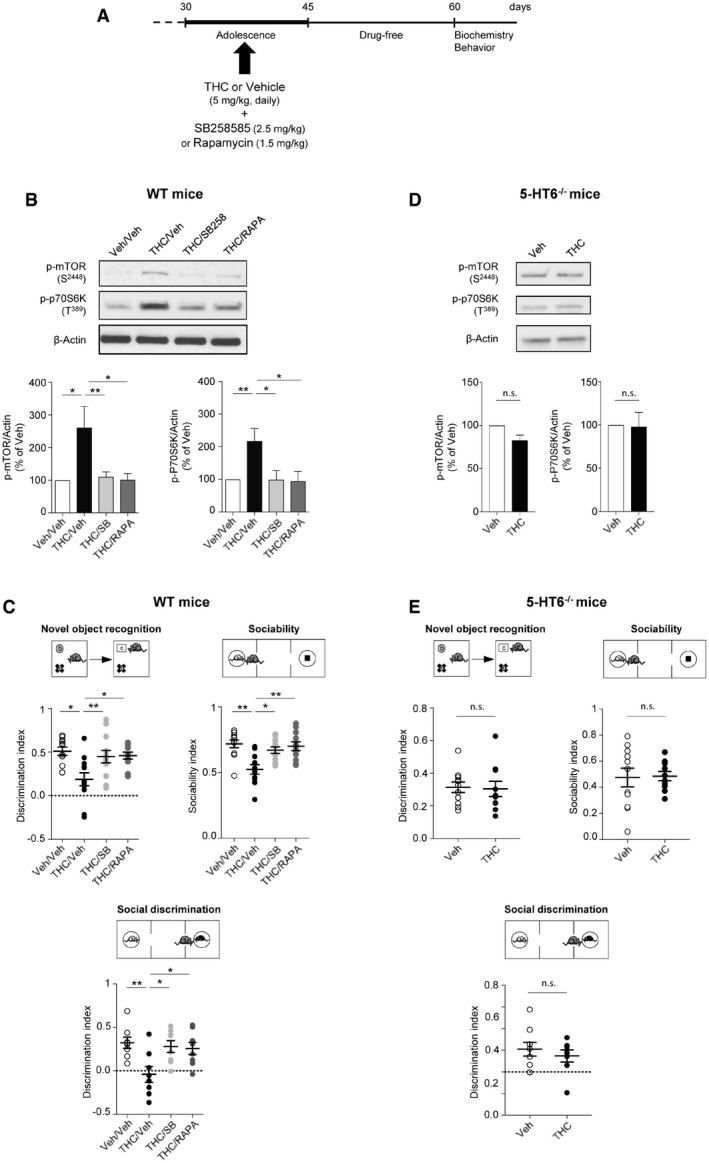
We used a preclinical model of cannabis abuse in adolescent mice, consisting of daily administration of THC (5 mg/kg, i.p.) between PNDs 30 and 45 (Fig 1A). This treatment induced in adulthood a marked increase in the phosphorylation level of mTOR at Ser2448 and of its substrate 70 kDa ribosomal protein S6 kinase (p70S6K) at Thr389 in the PFC (Fig 1B) but not in the hippocampus (Appendix Fig S1), compared to control mice injected with saline solution (vehicle), indicative of a sustained activation of mTOR signaling in PFC. As expected, THC administration did not promote mTOR activation in cannabinoid type 1 (CB1) receptor‐deficient mice (CB1 −/− mice, Fig EV1A), confirming that THC mediates its effects through CB1 receptors. The persistent elevation of phosphorylated mTOR and p70S6K elicited by THC administration during adolescence was prevented by the concomitant injection (between PNDs 30 and 45) of either SB258585 (2.5 mg/kg), a 5‐HT6 receptor antagonist (Hirst et al, 2000), or rapamycin (1.5 mg/kg), a pharmacological mTOR inhibitor (Fig 1B). Administration of SB258585 or rapamycin during adolescence did not significantly affect the basal phosphorylation state of mTOR and p70S6K in the PFC of vehicle‐injected mice (Appendix Fig S2A). Further supporting the role of 5‐HT6 receptors in the THC‐mediated effects, administration of THC during adolescence did not induce mTOR activation in the PFC of 5‐HT6 receptor‐deficient mice (5‐HT6 −/− mice, Fig 1D).
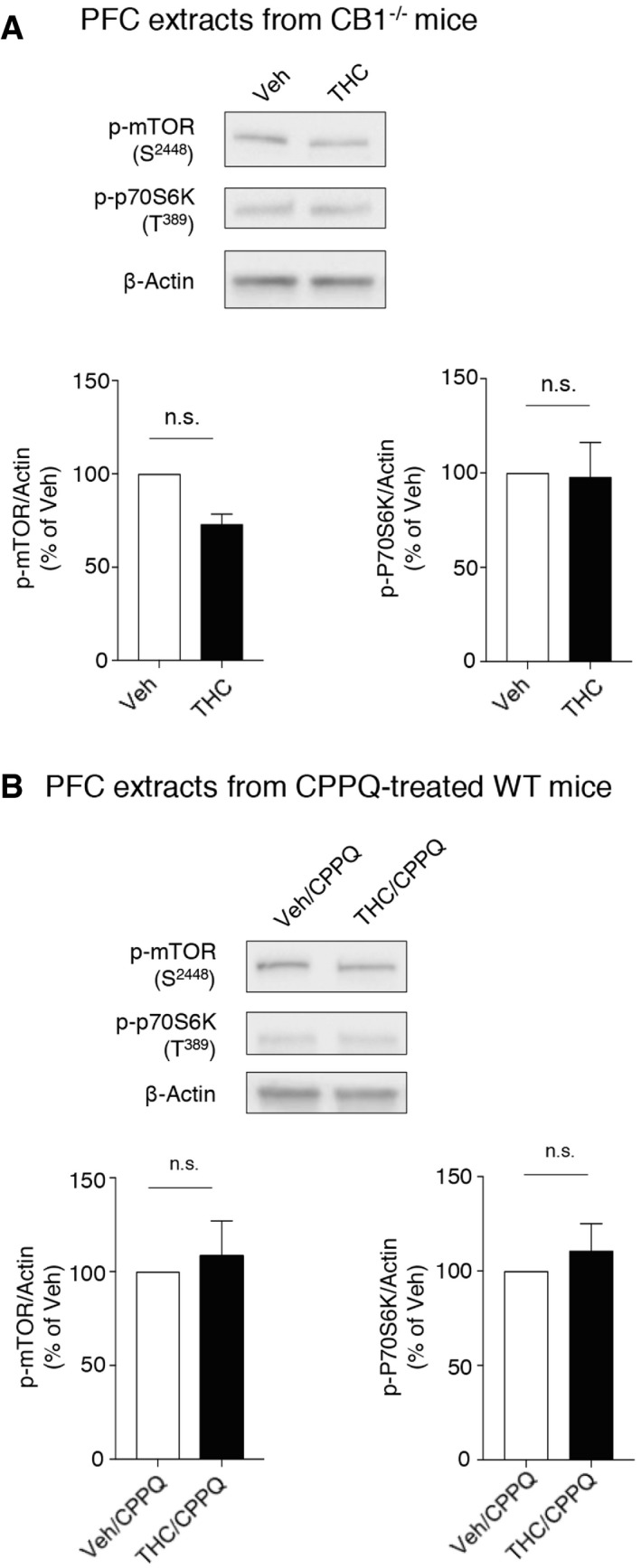
5HT6 receptors are known to exhibit a high level of constitutive activity both in vitro and in vivo (Kohen et al, 2001; Purohit et al, 2003; Duhr et al, 2014; Deraredj Nadim et al, 2016) that is inhibited by SB258585 (Duhr et al, 2014), which thus behaves as an inverse agonist. Administration of the recently characterized 5HT6 receptor neutral antagonist CPPQ ((S)‐1‐[(3‐chlorophenyl)sulfonyl]‐4‐(pyrrolidine‐3‐yl‐amino)‐1H‐pyrrolo[3,2‐c]quinolone, 2.5 mg/kg) (Deraredj Nadim et al, 2016; Grychowska et al, 2019) to mice injected with THC during adolescence prevented the persistent elevation of mTOR and p70S6K phosphorylation observed in adulthood (Fig EV1B), suggesting that mTOR activation is due to the activation of 5‐HT6 receptors by endogenously released 5‐HT rather than agonist‐independent activity.
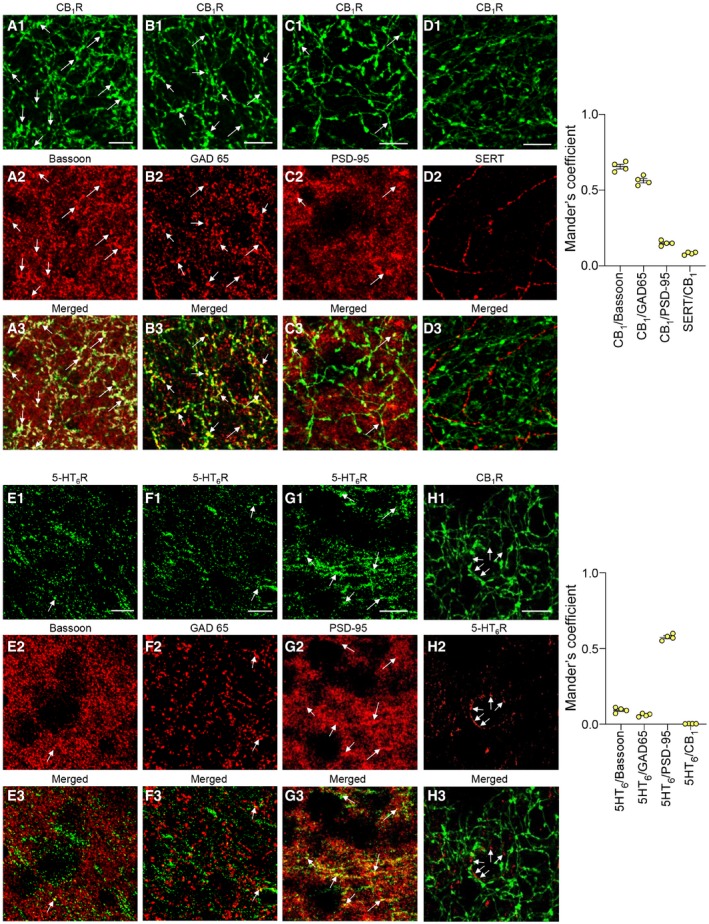
Immunostaining of CB1 and 5‐HT6 receptors showed distinct neuronal localizations of both receptors in the mouse PFC. Consistent with previous findings, CB1 receptors are mainly localized presynaptically on GABAergic neurons (as shown by their co‐localization with the presynaptic protein Bassoon, Fig 2A, and the GABAergic marker GAD65, Fig 2B; Eggan & Lewis, 2007; Cathel et al, 2014) while negligible receptor amounts were detected on 5‐HT terminals (Fig 2D). A fraction of CB1 receptors was also found at the postsynapse (co‐localization with PSD‐95, Fig 2C), corroborating previous findings (Maroso et al, 2016). Conversely, 5‐HT6 receptors are mostly localized at the postsynapse (Fig 2E–G). Furthermore, no co‐localization of CB1 and 5HT6 receptors was found within PFC neurons (Fig 2H). Collectively, these anatomical observations suggest that the long‐lasting activation of mTOR signaling in the PFC of THC‐injected mice does not result from a crosstalk between CB1 and 5‐HT6 receptor‐operated signaling but rather from a cross‐correlative action on the neuronal cortical network.
Early blockade of 5‐HT6 receptor‐operated mTOR signaling prevents cognitive deficits induced by THC administration during adolescence
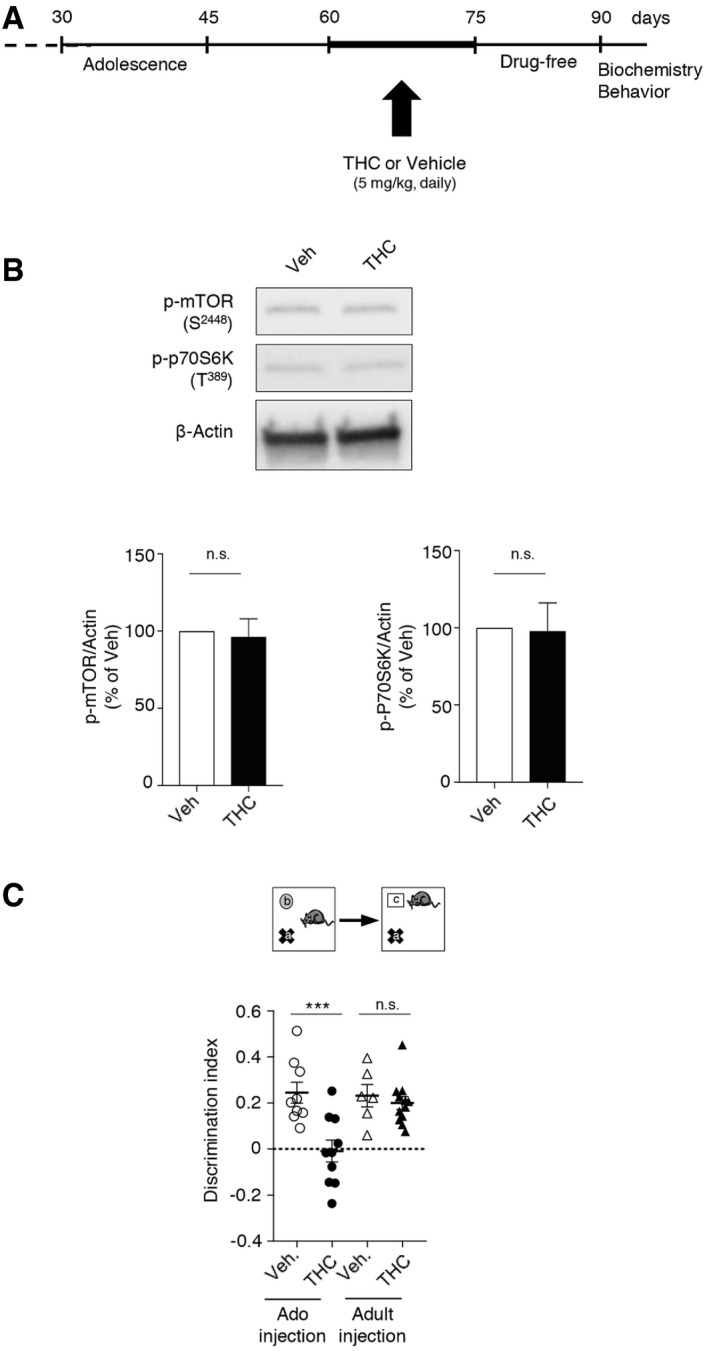
As previously observed in the rat (O’Shea et al, 2006), mice exposed to THC during adolescence showed later in life deficits in behavioral tasks assessing cognitive functions that depend on PFC among other regions: the novel object recognition test (discrimination index: 0.19 ± 0.07, N = 12, and 0.51 ± 0.05, N = 11, for THC + vehicle and for vehicle + vehicle conditions, respectively, P < 0.001, Fig 1C, left panel), the 3‐chamber social preference test (sociability index: 0.71 ± 0.03, N = 11, and 0.52 ± 0.03, N = 11, for THC + vehicle and for vehicle + vehicle conditions, respectively, P < 0.001, Fig 1C, right panel) and the social discrimination test (discrimination index: −0.04 ± 0.09, N = 8, and 0.32 ± 0.06, N = 8, for THC + vehicle and for vehicle + vehicle conditions, respectively, P < 0.01, Fig 1C, bottom panel). In contrast, administration of THC to mice during adolescence did not alter locomotion nor induced an anxiety phenotype in adulthood (Fig EV2).
Given the deleterious influence of non‐physiological mTOR activation upon cognition in various neuropsychiatric conditions (Hoeffer & Klann, 2010; Bockaert & Marin, 2015) and its role in cognitive deficits induced by cannabis intake, we next explored whether blocking 5‐HT6 receptor‐elicited mTOR elevation in adolescent mice exposed to THC prevents the associated cognitive impairments in adulthood. THC‐injected mice treated with SB258585 or rapamycin during adolescence showed a similar performance as vehicle‐injected animals in the novel object recognition task (discrimination index: 0.45 ± 0.07, N = 12, and 0.46 ± 0.04, N = 11, for THC + SB and THC + Rapa conditions, respectively, P > 0.05 vs. vehicle/vehicle mice, Fig 1C, left panel), the social preference test (social index: 0.67 ± 0.02, N = 12, and 0.70 ± 0.03, N = 11, for THC + SB and for THC + Rapa conditions, respectively, P > 0.05 vs. vehicle mice, Fig 1C, right panel) and the social discrimination test (discrimination index: 0.28 ± 0.06, N = 8, and 0.25 ± 0.07, N = 8, for THC + SB and for THC + Rapa conditions, respectively, P > 0.05 vs. vehicle mice, Fig 1C, bottom panel). SB258585 or rapamycin administration to vehicle‐treated mice did not alter their performance in each of these tests (Appendix Fig S2B). Likewise, THC administration to 5‐HT6 −/−mice during adolescence did not alter their performance in these three behavioral tasks (Fig 1E). Collectively, these results demonstrate that the chronic intake of THC during adolescence induces a 5‐HT6 receptor‐dependent long‐lasting activation of mTOR signaling in the PFC that causes cognitive deficits in adulthood.
Effects of THC administration and of blocking the 5‐HT6/mTOR pathway in adulthood
To explore whether blocking 5‐HT6 receptor‐mediated mTOR activation during adulthood could also have a delayed beneficial influence upon cognition, mice injected with THC during adolescence were treated with SB258585 (2.5 mg/kg) or rapamycin (1.5 mg/kg) during 2 weeks at the adult stage (daily injections from PND 60 to 75). Biochemical analysis and behavioral studies were performed 2 weeks after the last injection of the 5‐HT6 receptor antagonist or rapamycin (PND 90, Fig 3A). A significant increase in phosphorylated mTOR and p70S6K was observed at PND 90 in THC‐injected mice, compared with vehicle‐injected mice, and this mTOR overactivation was not affected by SB258585 or rapamycin administration at the adult stage (Fig 3B). Moreover, performances were similar in the THC‐injected mice treated or not with SB258585 or rapamycin in adulthood in the novel object recognition task (Fig 3C). These results demonstrate that blocking the 5‐HT6/mTOR signaling pathway at the adult stage in mice injected with THC during adolescence does not abolish the long‐lasting activation of mTOR and, consequently, does not induce persistent cognitive improvements.
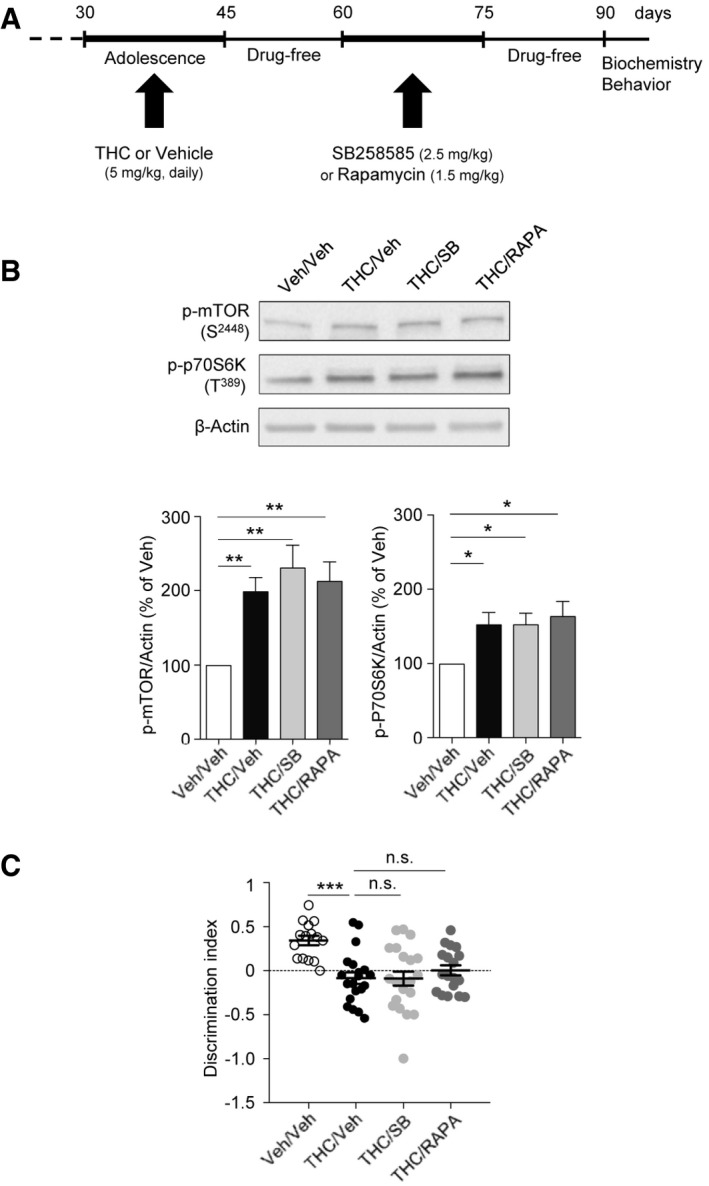
We also administered THC from PNDs 60 to 75 (Fig EV3A) to determine whether its long‐term deleterious effects upon mTOR signaling and cognition are really restricted to the juvenile period. Analysis of mTOR signaling 15 days after the last THC injection (PND 90) did not present any change in the phosphorylation state of mTOR and p70S6K in THC‐treated mice (Fig EV3B). Correspondingly, THC administration at the adult stage did not induce a persistent deficit in novelty discrimination in the novel object recognition test in the same time frame (Fig EV3C).
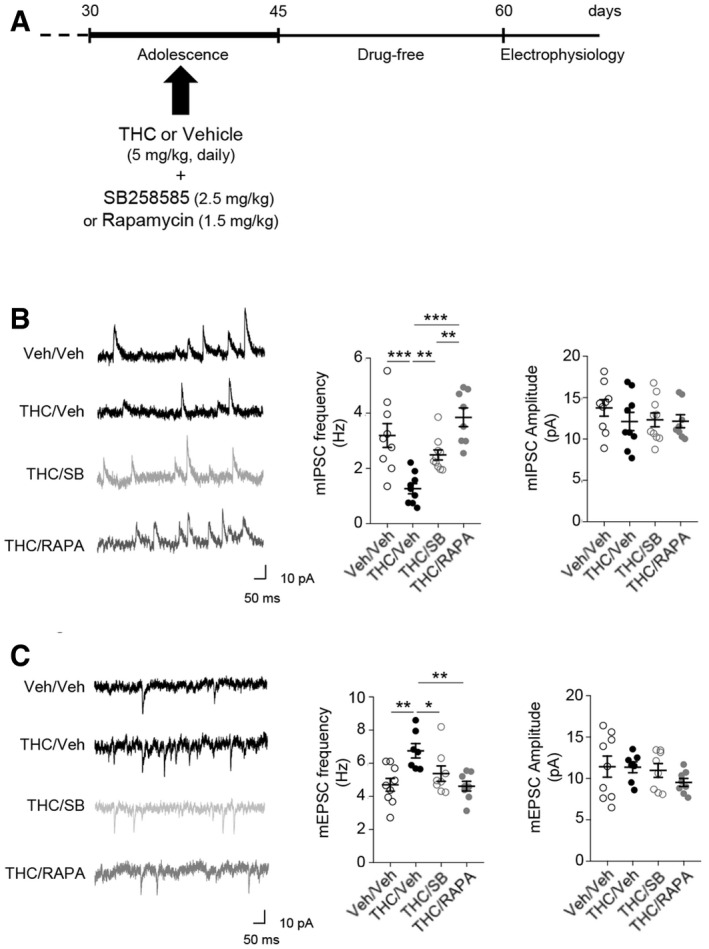
Alterations of GABAergic and glutamatergic synaptic transmissions in THC‐injected mice during adolescence are prevented by early blockade of 5‐HT6‐elicited mTOR activation
To determine whether chronic THC consumption during adolescence affects prefrontal synaptic transmission in adulthood, we analyzed the excitatory and inhibitory synaptic transmissions in acute slices of medial PFC. We performed whole‐cell patch‐clamp recordings of layer V pyramidal neurons, which integrate excitatory inputs from cortical and sub‐cortical areas and measured the amplitude and the frequency of both GABA receptor‐mediated miniature inhibitory postsynaptic currents (mIPSCs) and AMPA receptor‐mediated miniature excitatory postsynaptic currents (mEPSCs). THC‐injected mice showed a robust decrease in mIPSC frequency, whereas their amplitude was not affected (Fig 4B), suggesting a defect in GABA release. Furthermore, in experimental conditions where the inhibitory transmission was initially blocked, mEPSC frequency was significantly increased with no change in their amplitude (Fig 4C). THC‐induced alterations of GABAergic and glutamatergic transmissions were abolished by SB258585 or rapamycin administration during adolescence (Fig 4B and C). In mice injected with vehicle during adolescence, neither SB258585 nor rapamycin administration significantly affected mIPSC or mEPSC frequency and amplitude (Appendix Fig S3). These results suggest that cannabis abuse during adolescence induces a sustained alteration of GABAergic and glutamatergic synaptic transmissions in PFC. This leads to a disruption in the excitatory/inhibitory (E/I) balance that can be prevented by blocking 5‐HT6 receptor‐operated mTOR signaling.
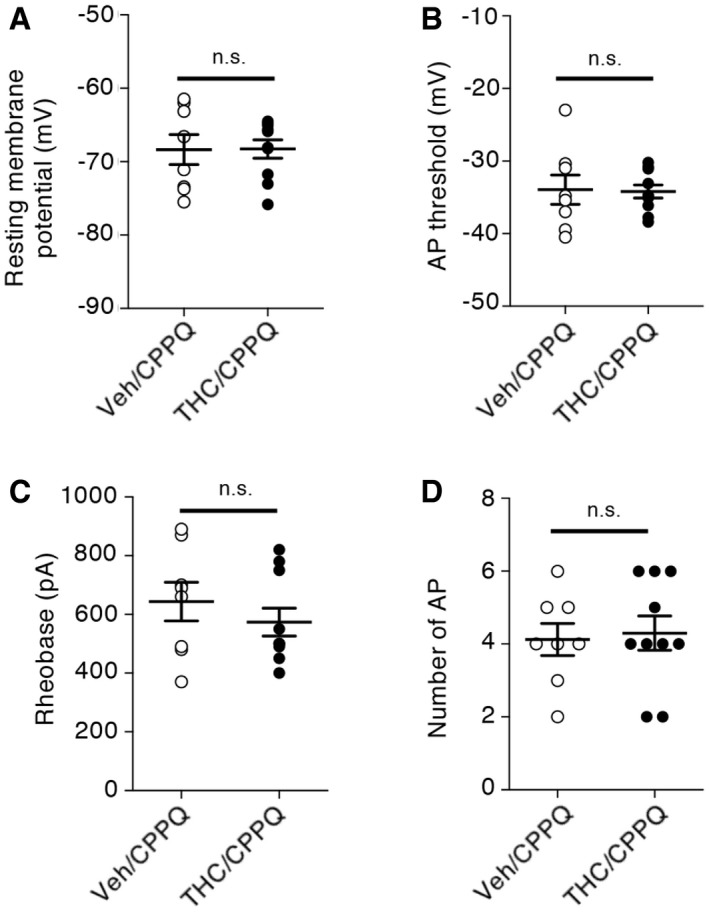
THC administration during adolescence induces changes in the intrinsic properties of layer V PFC pyramidal neurons through modulation of HCN1 channels
Spike timing results from dynamic interactions between synaptic activity and intrinsic neuronal excitability. We assessed whether persistent activation of mTOR elicited by THC administration during adolescence might affect the intrinsic electrophysiological properties and the firing rate of layer V pyramidal neurons by monitoring four different parameters: the resting membrane potential (RMP), the action potential (AP) threshold, the rheobase (i.e., minimal current required to induce neuronal firing), and the firing rate (i.e., number of APs for a 150 pA injected current during 250 ms). THC administration during adolescence significantly increased the resting membrane potential (Fig 5A) and lowered the AP threshold (Fig 5B) and the rheobase (Fig 5C), without affecting the firing rate, in adulthood (Fig 5D). Moreover, the concomitant administration of SB258585 or rapamycin prevented these changes (Fig 5A–C) but had no effect on the intrinsic neuronal properties of layer V PFC pyramidal neurons from vehicle‐injected mice (Appendix Fig S4). Mimicking the effects of SB258585, concomitant administration of the neutral 5‐HT6 receptor antagonist, CPPQ, to THC‐injected mice during adolescence also prevented the observed alterations of the resting membrane potential, the rheobase, and the AP threshold (Fig EV4A–C) of PFC pyramidal neurons, without affecting their firing rate (Fig EV4D). Collectively, these results indicate that THC intake during adolescence induces long‐lasting changes in the intrinsic neuronal properties of prefrontal layer V pyramidal neurons that are prevented by the early blockade of the 5‐HT6/mTOR pathway.
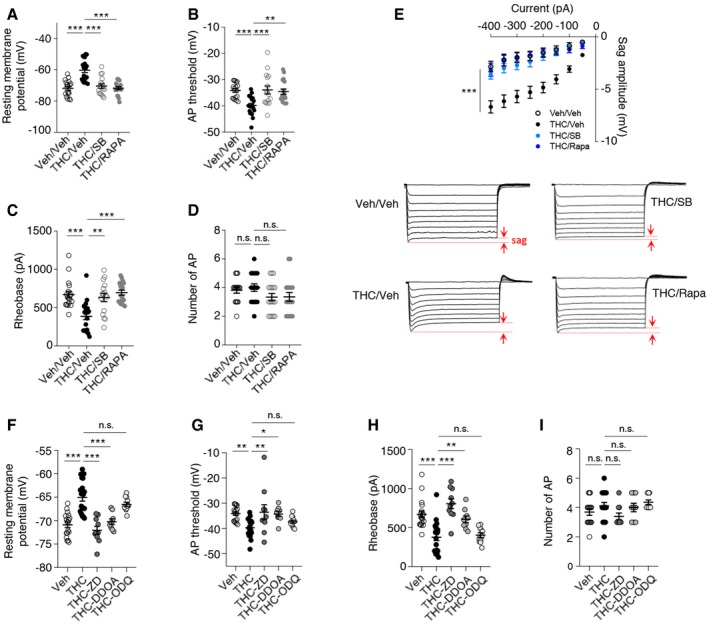
Hyperpolarization‐activated cyclic nucleotide‐gated channel 1 (HCN1) is the predominant isoform of HCN channels, a family of voltage‐gated ion channels responsible for the hyperpolarization‐activated current (I h) that modulates spike firing and synaptic potential integration (He et al, 2014; Shah, 2014) by influencing the resting membrane potential of pyramidal neurons. Notably, HCN1 channels enable intrinsic persistent firing of prefrontal layer V pyramidal neurons and are necessary for PFC‐dependent behavioral tasks such as executive function during working memory episodes (Thuault et al, 2013). Furthermore, HCN1 channels have been involved in CB1 receptor‐induced deficits in LTP in hippocampal pyramidal neurons located in the superficial portion of the CA1 pyramidal cell layer and in spatial memory formation (Maroso et al, 2016). We thus examined whether THC administration during adolescence likewise modifies the activity of HCN1 channels in PFC neurons and measured the voltage sag as an index of postsynaptic I h in layer V pyramidal neurons, using whole‐cell patch‐clamp recordings in a current‐clamp configuration. Following injection of incremental negative currents (50‐pA increment from −400 to 0 pA), the sag amplitude was increased in layer V pyramidal neurons from THC‐injected mice, compared with vehicle‐injected mice (Fig 5E). Furthermore, administration of SB258585 or rapamycin during adolescence restored a normal sag potential amplitude in THC‐injected mice (Fig 5E).
To confirm that HCN1 channels contribute to modifications of the intrinsic neuronal properties, we bath applied onto the PFC slices an organic HCN1 antagonist ZD7288 at a concentration of 10 μM that minimizes non‐specific effects on synaptic transmission (Chevaleyre & Castillo, 2002). Application of ZD7288 restored the resting membrane potential, the AP threshold and the rheobase of layer V pyramidal neurons from THC‐injected mice to values close to those observed in vehicle‐treated animals (Fig 5F–I), reminiscent of the effects induced by administration of SB258585 or rapamycin during adolescence, whereas it did not modify the intrinsic properties nor the firing pattern of neurons from vehicle‐treated mice (Appendix Fig S5). As previously shown (Thuault et al, 2013), the pharmacological inhibition of HCN1 did not significantly alter the firing rate in layer V pyramidal neurons from vehicle‐injected animals (Fig 4I and Appendix Fig S5).
HCN1 are gated by both cAMP and cGMP (He et al, 2014). Any increase in cAMP or cGMP would thus cause a depolarizing shift in the activation curve for I h. Consistent with this hypothesis, a recent study has shown that CB1 receptors modulate HCN1 activity and increase postsynaptic I h through a cGMP‐dependent pathway (Maroso et al, 2016). In order to determine whether the alteration of HCN1 activity is mediated by an increase in intracellular cGMP, we perfused 1H‐[1,2,4]oxadiazolo[4,3‐a]quinoxalin‐1‐one (ODQ, 10 μM), a selective inhibitor of guanylate cyclase (the enzyme responsible for cGMP production) (Boulton et al, 1995) into the recording pipette. Postsynaptic application of ODQ did not modify the intrinsic neuronal properties of neurons from THC‐injected mice (Fig 5F–I). In contrast, perfusion of the selective adenylate cyclase inhibitor 2′,3′‐dideoxyadenosine (DDOA, 15 μM; Pelkey et al, 2008) restored the resting membrane potential (Fig 5F), the AP threshold (Fig 5G) and the rheobase (Fig 5H) of layer V pyramidal neurons from THC‐injected mice to values similar to those observed in vehicle‐treated animals. Application of DDOA or ODQ did not modify the intrinsic properties nor the firing pattern of neurons from vehicle‐injected mice (Appendix Fig S5). Collectively, these results suggest a role for cAMP in the HCN1‐mediated alterations of neuronal intrinsic properties of PFC pyramidal neurons.
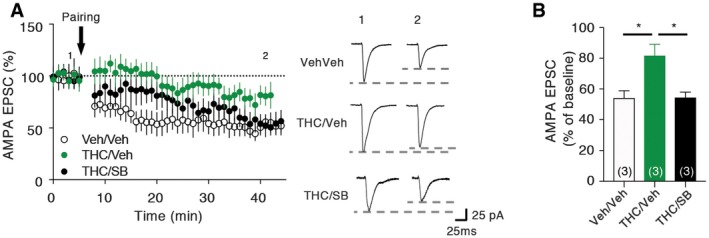
LTD impairment induced by adolescent THC exposure is prevented by early blockade of 5‐HT6 receptors
Previous studies have shown that exposure of rats to THC during adolescence disrupts endocannabinoid‐dependent long‐term depression (LTD) in the PFC at adulthood (Rubino et al, 2015). Corroborating these findings, we found that electrically induced LTD at PFC layer I/V synapses was impaired in mice treated with THC during adolescence (Fig 6A and B). As expected, the concomitant administration of SB258586 during adolescence restored normal LTD at these synapses (Fig 6A and B), indicating that long‐term modifications of synaptic plasticity induced by adolescent exposure to THC depend on 5‐HT6 receptor activation during this critical period of PFC maturation.
Discussion
Previous studies have shown that an acute administration of THC in adult mice induces a transient mTOR activation in the hippocampus that leads to an alteration in protein translation in this brain area and amnesic‐like effects in memory tasks depending of hippocampal function (Puighermanal et al, 2009, 2013). In contrast, in the present study, we showed that chronic exposure to THC during adolescence induces a sustained activation of mTOR signaling in the PFC but not in the hippocampus, reminiscent of previous observations in two neurodevelopmental models of schizophrenia, rats treated with phencyclidine at the neonatal stage or reared in social isolation after the weaning (Meffre et al, 2012). These findings are consistent with the hypothesis that common signaling mechanisms might contribute to cognitive symptoms in patients with schizophrenia and cannabis abusers during adolescence. They also suggest that THC intake might differentially influence mTOR activity depending on its mode of administration and the age of delivery: whereas an acute administration at the adult stage induces a transient elevation of mTOR signaling in the hippocampus (Puighermanal et al, 2009, 2013), chronic intake during adolescence leads to a long‐lasting mTOR activation in the PFC that persists in adulthood. The mechanisms underlying THC‐elicited mTOR activation following acute administration at the adult stage or chronic intake during adolescence also differ. Activation of mTOR induced by THC in adult mice is mainly mediated by CB1 receptors located in GABAergic terminals, leading to an inhibition of GABAergic tone in the hippocampus. This in turn enhances pyramidal glutamatergic inputs and promotes NMDA receptor‐dependent mTOR activation in pyramidal neurons (Puighermanal et al, 2009, 2013). Though such a mechanism might likewise contribute to the initial phase of prefrontal mTOR activation in mice exposed chronically to THC during adolescence, the activation of mTOR during the entire adolescent period clearly depends on 5‐HT6 receptor activation (Fig 7). Indeed, it was not observed in 5‐HT6 −/− mice and was prevented by the administration of a 5‐HT6 receptor neutral antagonist during the adolescence period, but not by the 5‐HT6 receptor blockade at the adult stage. The latter observation indicates it might result from a non‐physiological 5‐HT6 receptor activation by endogenously released 5‐HT rather than constitutive activity, which might be caused by CB1 receptor‐mediated decrease in GABA release and the disinhibition of 5‐HT terminals (Fig 7) in the PFC.
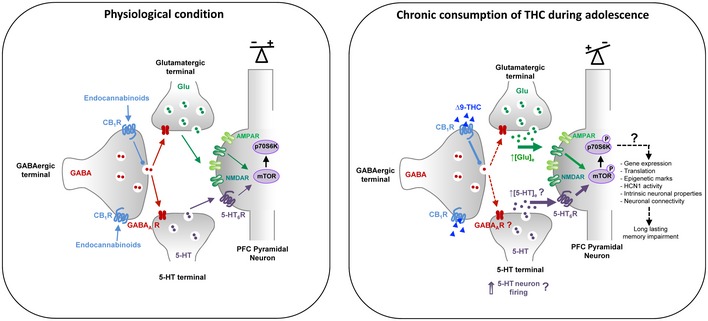
The mechanism underlying the persistent activation of mTOR in adulthood remains more uncertain. It is conceivable that adolescent exposure to THC and the resulting non‐physiological mTOR activation interfere with the maturation of the GABAergic system that undergoes refinement in the PFC until the end of adolescence (Kilb, 2012; Zamberletti et al, 2014). This might lead to a sustained increase in PFC serotonergic tone (Teissier et al, 2017) and, consequently, a persistent 5‐HT6 receptor activation by endogenously released 5‐HT.
A recent RNA‐seq study in rats treated with THC during adolescence showed a decreased expression of Raptor mRNA 2 weeks after the THC treatment, while the level of mTOR mRNA was not affected (Miller et al, 2019). This suggests that the enhanced mTOR activity in THC‐treated animals does not result from an increased expression of proteins of the mTOR complex 1 (mTORC1), but rather from an increase in mTOR catalytic activity, consistent with the observed increase in Ser2448 phosphorylation state.
The sustained increase mTOR signaling in PFC of THC‐treated mice plays a key role in the emergence of cognitive deficits in adulthood. Indeed, the cognitive deficits induced by THC intake during adolescence were abolished by the concomitant administration of either 5‐HT6 receptor antagonists or rapamycin and were absent in the 5‐HT6 −/− mice. Notably, the same treatments (followed by the same washout period) administered in adulthood did not induce a prolonged rescue of these deficits. Furthermore, THC administration in adulthood did not induce such a prolonged activation of mTOR signaling and associated cognitive deficits. Collectively, these observations confirm that the adolescence is a period of vulnerability to extrinsic risk factors such as drug abuse that irreversibly affect brain signaling processes crucial for cognitive functions (Higuera‐Matas et al, 2015; Renard et al, 2016; Saravia et al, 2019).
The sustained activation of mTOR signaling, under the control of 5‐HT6 receptors, elicited in PFC by adolescent THC exposure might induce cognitive deficits in adulthood through different mechanisms. First, mTOR plays a critical role in various neurodevelopmental processes, including neuronal progenitor proliferation, neuronal migration, growth of dendrites and axons, and synaptogenesis (Bockaert & Marin, 2015). Corroborating these observations, a large body of evidence indicates a deleterious influence of aberrant mTOR signaling upon cognition in rodent models of neurodevelopmental disorders (Ehninger et al, 2008; Sharma et al, 2010; Ricciardi et al, 2011; Meffre et al, 2012; Troca‐Marin et al, 2012; Huber et al, 2015), indicating that the deregulation of mTOR signaling in specific brain areas at critical developmental periods can compromise cognition later in life. Second, mTOR activation level finely controls dendritic spine pruning and shaping of synaptic connections at later stages of brain maturation but contrasting effects of mTOR overactivation upon spine density and morphology have been reported in autism models (Tavazoie et al, 2005; Tang et al, 2014). THC exposure during adolescence results in premature pruning of spines and protracted atrophy of distal apical trees associated with alteration of synaptic markers that might lead to a reduction in the complexity of pyramidal neurons and a reduced capacity for plasticity in neural circuits central for normal adult behavior (Rubino et al, 2015; Miller et al, 2019). The role of mTOR in structural abnormalities induced by adolescent exposure to THC remains to be established. Finally, a recent study revealed that exposure of adolescent rats to THC affects selective histone modifications that impact the expression of genes associated with synaptic plasticity. Again, changes in histone modifications are more widespread and pronounced after adolescent exposure than after adult exposure, suggesting specific adolescent vulnerability (Miller et al, 2019; Prini et al, 2018). As mTOR activity controls the phosphorylation state of proteins involved in histone post‐translational modifications and chromatin remodeling (Citro et al, 2015; Zhang et al, 2017), the sustained mTOR activation in PFC of mice exposed to THC during adolescence might also contribute to some epigenetic mechanisms underlying the development of cognitive deficits (Prini et al, 2018).
Irrespective of the structural changes and the molecular mechanisms elicited by the sustained non‐physiological mTOR activation induced by chronic THC consumption during adolescence, we showed that it induces a disruption in the excitatory–inhibitory balance in PFC. This alteration in the E/I balance, which is a landmark of neurodevelopmental disorders (Dani et al, 2005; Bateup et al, 2013; Berryer et al, 2016) such as schizophrenia or autism (Benes et al, 1991; Vogels & Abbott, 2009; Marin, 2012), persisted until adulthood and resulted from a decrease in GABAergic transmission and an increase in glutamatergic transmission. It might also result from the 5‐HT6 receptor‐dependent overactivation of mTOR, as the blockade of this pathway during adolescence restored a normal E/I balance in adulthood in THC‐injected mice.
In an effort to identify the molecular mechanisms underlying the disruption of the E/I balance following adolescent exposure to THC, we showed that this treatment induces an increase in HCN1 activity that leads to changes in intrinsic properties of PFC layer V pyramidal neurons characterized by a more depolarized resting membrane potential and a lower firing threshold. Previous RNA‐seq studies demonstrated a decrease in HCN1 mRNA level 2 weeks after the chronic administration of THC to adolescent rats, suggesting that the observed increase in HCN1 activity does not result from an increase in HCN1 channel expression (Miller et al, 2019) but is rather reminiscent of previous findings indicating that CB1 receptor activation induces deficits in hippocampal LTP and spatial memory formation through HCN channels (Maroso et al, 2016). The signaling cascade underlying CB1 receptor‐mediated activation of I h in the hippocampus involves a c‐Jun‐N‐terminal‐kinase (JNK) that in turn increases cGMP formation through the activation of nitric oxide synthase (Maroso et al, 2016). In contrast, HCN1‐dependent alterations of intrinsic properties of PFC pyramidal neurons elicited by administration of THC during adolescence were independent of cGMP, but rather involved a cAMP‐dependent mechanism that might itself result from the sustained activation of adenylyl cyclase mediated by prefrontal 5‐HT6 receptors. Previous studies have also suggested that HCN1 channels regulate synaptic plasticity by dendritic integration of synaptic inputs to pyramidal neurons and thus play a role in learning and memory (Magee, 1999; Nolan et al, 2004). Indeed, alteration in HCN1 activity might also explain the impairment of electrically induced LTD observed at PFC layer I/V synapses in mice injected with THC during adolescence.
It is well accepted that HCN1 are important during neuronal development and network wiring (Bender & Baram, 2008). Developing neurons have to modulate both their intrinsic properties as well as their firing to constantly adapt to their changing environment. Factors that interfere with this process and alter HCN1 activity may have long‐lasting deleterious effects that lead to network dysfunction underlying cognitive impairment observed in several brain pathologies (Chen et al, 2001; Brewster et al, 2002). For instance, an alteration in HCN1 activity has been found in mouse models of Rett syndrome (Mecp2−/y mice) (Balakrishnan & Mironov, 2018) and Fragile X syndrome (Fmr1−/y mice) (Brager et al, 2012) as well as in epilepsy (Dube et al, 2006).
In conclusion, the present study shows that a sustained, non‐physiological mTOR activation under the control of 5‐HT6 receptors plays a key role in the alteration of intrinsic neuronal properties, E/I balance and synaptic plasticity in layer V pyramidal neurons of the PFC of adult mice exposed to cannabis during adolescence. It also shows that blocking 5‐HT6 receptor‐elicited mTOR activation during this critical period of PFC maturation definitively prevents emergence of cognitive deficits in mice exposed to cannabis during adolescence, whereas its blockade in adulthood does not induce such a long‐term pro‐cognitive effect. It suggests that 5‐HT6 receptor antagonists, which recently failed in Phase III clinical trials as symptomatic treatment of cognitive symptoms in Alzheimer’s disease (Atri et al, 2018; Khoury et al, 2018), might be repositioned as “disease‐modifying” treatment to prevent emergence of cognitive deficits in adolescent cannabis abusers. Such a strategy based on early administration of 5‐HT6 receptor antagonists is certainly more relevant than mTOR blockade by pharmacological inhibitors, as it will specifically prevent the non‐physiological activation of prefrontal mTOR without affecting physiological cerebral mTOR activity, which plays a key role in numerous physiological processes such as synaptic plasticity (Bockaert & Marin, 2015; Younts et al, 2016; Switon et al, 2017; Ryskalin et al, 2018). Accordingly, 5‐HT6 receptor antagonism will not reproduce the severe side effects induced by mTOR inhibitors such as rapamycin, which limit their clinical development for the treatment of psychiatric diseases. Given the deleterious influence of aberrant mTOR activation in several genetic forms of autism spectrum disorders (Ehninger et al, 2008; Ehninger & Silva, 2011; Huber et al, 2015; Sato, 2016; Ryskalin et al, 2018), administration of 5‐HT6 receptor antagonists as soon as adolescence might profitably be evaluated in autism. Finally, the recent progress in the identification of patients with high risk of transition to schizophrenia (Millan et al, 2016) suggests that such a strategy might also be extended to the more numerous population of patients with schizophrenia.
Materials and Methods
Animals
Wild‐type male and female C57BL/6JRj mice were purchased from Janvier Laboratories. 5‐HT6 −/− mice have a C57BL/6JRj background. Mice from both sexes were indifferently used in behavioral, electrophysiological, and biochemical experiments. Mice were received at PND 26 and housed under standardized conditions with a 12‐h light/dark cycle, stable temperature (22 ± 1°C), controlled humidity (55 ± 10%), and free access to food and water. CB1−/− mice and their littermates (background C57BL/6JRj) were housed in a specific area under standardized conditions as described above. Animal husbandry and experimental procedures were performed in compliance with the animal use and care guidelines of the University of Montpellier, the French Agriculture Ministry, and the European Council Directive (86/609/EEC). All the experiments were conducted from PND 60 or from PND 90 depending on the experimental conditions.
Drugs and treatments
Δ‐9‐tetrahydrocannabinol (THC) was purchased from THC Pharm. Picrotoxin (Sigma P1675), 6‐Cyano‐7‐nitroquinoxaline‐2,3‐dione disodium salt hydrate (CNQX; Sigma C239), and N‐ethyl‐1,6‐dihydro‐1,2‐dimethyl‐6‐(methylimino)‐N‐phenyl‐4‐pyrimidinamine hydrochloride (ZD7288; Sigma Z3777) were obtained from Sigma‐Aldrich. 4‐Iodo‐N‐[4‐methoxy‐3‐(4‐methylpiperazin‐1‐yl)‐phenyl]benzene‐sulfonamide (SB258585; Tocris #1961), tetrodotoxin (Tocris #1078) and D,L‐2‐Amino‐5‐phosphonopentanoic acid (D,L‐AP5; Tocris #0106) were purchased from Tocris and rapamycin from LC Laboratories (R5000). THC was dissolved in 5% ethanol and 5% cremophor in NaCl. Vehicle groups received this solution. THC was injected daily (5 mg/kg, i.p.) between PNDs 30 and 45. SB258585 (injected at 2.5 mg/kg, i.p.), rapamycin (injected at 1.5 mg/kg, i.p.), and CPPQ (injected at 2.5 mg/kg, i.p.) were dissolved in 5% DMSO and 5% Tween‐80 in NaCl. All mice received the same number of injections. Correspondingly, control mice were successively injected with the vehicle used for THC and the vehicle used for SB258585/Rapamycin and are referred as Veh/Veh on the figures. Likewise, mice treated with the vehicle used for THC and SB/Rapa were referred as Veh/SB and Veh/Rapa, respectively, while mice treated with THC and the vehicle used for SB/Rapa were both referred to THC/Veh.
Slice preparation
Animals were anesthetized with isoflurane before sacrifice. Slices were performed as previously described (Barre et al, 2016; Berthoux et al, 2019). Brains were removed and rapidly transferred into ice‐cold dissection buffer maintained in 5% CO2/95% O2 and containing 25 mM NaHCO3 (Sigma S5761), 1.25 mM NaH2PO4 (Sigma S8282), 2.5 mM KCl (Sigma P3911), 0.5 mM CaCl2 (Sigma C5080), 7 mM MgCl2 (Sigma M2670), 25 mM glucose (Sigma G7021), 110 mM choline chloride (Sigma C1879), 11.6 mM ascorbic acid (Sigma A4034), and 3.1 mM pyruvic acid (Sigma P3256). Coronal brain slices (300 μm) were cut in ice‐cold dissection buffer using a vibratome (Leica VT1200S). Slices were then transferred to artificial cerebrospinal fluid (aCSF, containing 118 mM NaCl, 2.5 mM KCl, 26.2 mM NaHCO3, 1 mM NaH2PO4, 11 mM glucose, 1.3 mM MgCl2, 2.4 mM CaCl2, and maintained in 5% CO2/95% O2), at room temperature (22–25°C).
Electrophysiological recordings
Patch recording pipettes (3–5 MΩ) were filled with intracellular solution (115 mM Cs‐gluconate, 20 mM CsCl (Sigma C3011), 10 mM HEPES (Sigma H3375), 2.5 mM MgCl2, 4 mM Na2ATP (Sigma A2383), 0.4 mM NaGTP (Sigma G8877), 10 mM sodium phosphocreatine (Sigma P7936), and 0.6 mM EGTA (Sigma E4378), pH 7.3). Whole‐cell recordings were obtained from layer V pyramidal neurons (300–400 μm from pial surface) of prelimbic PFC using a Multiclamp 700B amplifier (Axon Instruments) under an Axioscope2 microscope (Zeiss) equipped with infrared differential interference contrast optics. Data were filtered at 2 kHz and sampled at 10 kHz using Digidata 1440A (Molecular Devices) under the control of pClamp 10 (Axon Instruments). There were no significant differences in input or series resistance among groups. For miniature postsynaptic currents, the recording chamber was perfused with ACSF supplemented with 1 μM tetrodoxin at 22–25°C. mEPSC and mIPSC recordings were performed in the presence of 0.1 mM picrotoxin and 0.1 mM CNQX with 0.1 mM D, L‐AP5, respectively. Currents were monitored for 10 min, and the quantification was made during the last 2 min of recording with Clampfit 10.2 (Axon Instruments).
Intrinsic electrophysiological properties were assessed by current‐clamp recordings. Patch recording pipettes were filled with intracellular solution (120 mM K‐gluconate, 10 mM KCl, 10 mM HEPES, 1.8 mM MgCl2, 4 mM Na2ATP, 0.3 mM NaGTP, 14 mM Na‐ phosphocreatine, 0.2 mM EGTA, pH 7.2). Resting membrane potential of each cell was measured immediately after the whole‐cell patch formation. Action potentials (APs) were evoked by a ramp current injection with 10‐pA increments for 2 ms, and only the first AP was used to determine the AP threshold and the minimal current necessary to induce firing (rheobase). The firing rate was recorded by a current injection at 150 pA for 250 ms.
Hyperpolarization‐activated cation current (I h) was measured as the voltage sag to a 500 ms hyperpolarizing current injection (50‐pA increment from −400 pA to 0). Synaptic transmissions were blocked by adding 0.1 mM picrotoxin, 0.01 mM CNQX, and 0.1 mM D,L‐AP5.
Electrically induced LTD was generated by a pairing protocol consisting in seven trains of 50 Hz stimuli (100 pulses per train), delivered at 0.1 Hz, as previously described (Berthoux et al, 2019). Currents were monitored for 6 min before tetanic stimulation. Quantification of AMPA EPSC amplitude was made during the last 5‐min period before tetanic stimulation and during the last 5‐min period of recording, using Clampfit 10.2 (Axon Instruments).
Analysis of mTOR activity in mouse PFC
Mice PFC were rapidly dissected and homogenized in ice‐cold buffer containing 0.32 M sucrose, 10 mM HEPES, pH 7.4, and a cocktail of protease and phosphatase inhibitors (Roche). Homogenates were centrifuged at 1,000 × g for 10 min to remove nuclei and large debris. Protein concentration in supernatants (PFC protein extracts) was determined by the bicinchoninic acid method. Proteins were resolved on 4–15% gradient gels (Bio‐Rad) and transferred electrophoretically onto nitrocellulose membranes (Bio‐Rad). Membranes were incubated in blocking buffer (Tris–HCl, 50 mM, pH 7.5; NaCl, 200 mM; Tween‐20, 0.1%, and skimmed dried milk, 5%) for 1 h at room temperature and overnight with primary antibodies in blocking buffer: rabbit anti‐phospho‐S2448 mTOR 1:1,000 (Cell Signaling #2971), rabbit anti‐phospho‐T389 p70S6K 1:1,000 (Cell Signaling #9205), and mouse anti‐panActin 1:2,000 (Lab Vision MS‐1295‐B). Membranes were then washed and incubated with horseradish peroxidase‐conjugated anti‐rabbit or anti‐mouse antibodies (1:4,000 in blocking buffer, Millipore #12‐348 and 12‐349, respectively) for 1 h at room temperature. Immunoreactivity was detected with an enhanced chemiluminescence method (ECL detection reagent, GE Healthcare) using a ChemiDoc™ Touch Imaging System (Bio‐Rad).
Immunohistochemistry
Mice were anesthetized with pentobarbital (100 mg/kg, i.p.; CevaSanteAnimale) and transcardially perfused with 4% paraformaldehyde, 0.1 M sodium phosphate buffer (PBS, pH 7.4). Brains were post‐fixed overnight in the same solution and stored at 4°C. Coronal slices (50 μm) were cut in the PFC with a vibratome (Leica VT1000S) and stored at −20°C in a cryoprotectant solution until being processed for immunohistochemistry. Briefly, brain sections were blocked in 1% Triton X‐100, 10% goat serum, 0.1 M PBS for 2 h, and immunostained overnight with primary antibodies: rabbit anti‐5‐HT6 receptor 1:500 (ab103016, Abcam), guinea‐pig anti‐CB1 receptor 1:500 (Frontier Institute #ab2571593), mouse anti‐PSD95 1:500 (NeuroMab #75‐028), mouse anti‐Bassoon 1:1,000 (Enzo Life Sciences, #ADI‐VAM‐P5003), mouse anti‐GAD65 1:500 (Millipore #MAB351R), and rabbit anti‐SERT 1:500 (Sigma #PC177L). Slices were then incubated with Alexa 594‐ or Alexa 488‐conjugated goat anti‐rabbit (1:1,000, Jackson immunoresearch), Alexa 488‐conjugated anti‐guinea‐pig IgG (1:1,000, Jackson immunoresearch) or Alexa 594‐conjugated goat anti‐mouse (1,000, Jackson immunoresearch). Sections were mounted and were analyzed under a 40× oil‐immersion objective using a confocal microscope (Carl Zeiss LSM 780). Briefly, for each image, we determined a precise ROI that was applied for all images. Then, we performed segmentation by thresholding to generate binary images from each selection using the « Otsu » auto‐threshold function. For co‐localization analysis, we used Mander’s split coefficients to identify the fraction of CB1 receptors or 5‐HT6 receptors that co‐localizes with synaptic markers and the fraction of CB1 that co‐localizes with 5‐HT6 receptors using the macro « coloc 2 » from ImageJ.
Behavioral analysis
Novel object recognition task
One week before the test, mice were extensively handled: The first 2 days, the operator put his hand on the home cage of the animals to familiarize the animals to his presence, and the following days, the animals were handled few minutes per day by the operator. Testing was carried out in a Plexiglas box placed in a dimly lit room with clearly visible contextual cues (black on white patterns) on the surrounding walls. Mice were habituated to the arena for 10 min per day, on days 1 and 2. On days 3, mice performed the novel object recognition task. The mice had a 10‐min training session, a 30‐min retention interval during which they were transferred back to the home cage, and a 10‐min test session. The objects were plastic toys (approximately 3 cm width, 3 cm length, 5 cm height) and were cleaned with 10% ethanol between sessions. The experiments were video‐recorded, and exploration times (nose in contact or sniffing at < 1 cm) were measured by a blinded observer. Mice with total time exploration of less than 3 s in test session were excluded. Discrimination indexes [(exploration time of novel object − exploration time of familiar object)/total object exploration time] were compared between groups.
Social preference task
Testing was carried out in a rectangular, three‐chamber box with dividing walls made of clear Plexiglas and an open middle section (60 cm length × 40 cm width in total, each compartment has a length of 20 cm), which allows free access to each chamber (Kaidanovich‐Beilin et al, 2011). Before the test, each tested mouse was placed for 10 min in the middle compartment a three‐chamber device, without the walls between the compartments, to allow free access to the three compartments. Then, a control mouse (same age, same sex as tested mice) was placed inside a wire containment jail, located in one chamber. In the second chamber, an object was placed inside a wire containment jail. Then, the walls were removed between the compartments, to allow free access for the test mouse to explore each of the three chambers. Duration of direct contacts between the tested mouse and the containment jail or cup housing the conspecific or object was recorded for 10 min. The experiments were video‐recorded, and exploration times (nose in contact or sniffing at < 1 cm) were measured by a blinded observer. Sociability indexes [(exploration time of congener − exploration time of object)/total exploration time] were compared between groups.
Social discrimination task
For the social novelty discrimination test (ability of an adult mouse to discriminate novel from familiar congeners), the same procedure as for the social preference task was used but the object in the wire containment jail was replaced by a mouse (same age, same sex as the tested mouse) during the second session. The walls between the compartments were then removed to allow free access to each of the three chambers for the tested mouse. The time of interaction the test mice with each congener was recorded for 10 min. Mice with total time exploration of less than 3 s in test session were excluded. Social discrimination indexes [(exploration time of the novel congener − exploration time of familiar congener)/total exploration time] were compared between groups.
Elevated Plus Maze
Anxiety‐related behavior was assessed in an Elevated Plus Maze (EPM) consisting in a black plexiglass apparatus with four elevated (50 cm above the floor) arms (26 cm long × 5 cm wide) set in cross from a neutral central square (5 × 5 cm). Two opposing arms, chipboard walls, and two opposing arms were devoid of walls (open arms), under dim lighting conditions (> 50 lux). Cumulative time spent in open (aversive) and closed (non‐aversive) arms was recorded during a 5‐min session.
Circular corridor
Locomotor activity was measured for 60 min in a circular corridor (14 cm wide, 18 cm in diameter) with four infrared beams placed at 90° angles (Imetronic, Pessac, France) in a low luminosity environ.
Statistical data analysis
For biochemistry, immunohistochemistry, and electrophysiology, a minimum of three animals was used in each group. Mice from both sexes were indifferently used, and they were submitted to behavioral experiments in a randomized sequential order. Behavioral recordings were blindly analyzed. Data were analyzed using GraphPad Prism software (v. 7.0). The homogeneity of sample variance was tested using Brown–Forsythe’s test. As long as no variance among groups was detected, ANOVA was performed. Statistical significance was determined by one‐way ANOVA followed by Newman–Keuls test for electrophysiological and biochemical experiments, or by one‐way ANOVA followed by Bonferroni test for behavioral experiments. Detailed statistics for figures are in the Appendix Table S1.
Author contributions
CBer performed electrophysiology experiments. CBer, CC and AR performed biochemistry experiments; AMH designed, performed and analyzed behavioral experiments. AR and ELD performed and analyzed behavioral experiments; AR and FA performed immunohistochemistry. PZ and KG synthesized and provided CPPQ. SCD provided the 5‐HT6 −/− mouse cohort. RM provided the CB1−/− mouse cohort. CBéc, JB and PM designed the experiments and supervised the study; CBéc, PM and CBer wrote the manuscript.
Conflict of interest
The authors declare that they have no conflict of interest.
Supplementary Materials
References
- GABAergic spill‐over transmission onto hippocampal mossy fiber boutons.. J Neurosci, 2007. [PubMed]
- Causal association between cannabis and psychosis: examination of the evidence.. Br J Psychiatry, 2004. [PubMed]
- Effect of idalopirdine as adjunct to cholinesterase inhibitors on change in cognition in patients with Alzheimer disease: three randomized clinical trials.. JAMA, 2018. [PubMed]
- Rescue of hyperexcitability in hippocampal CA1 neurons from Mecp2 (−/y) mouse through surface potential neutralization.. PLoS One, 2018. [PubMed]
- Presynaptic serotonin 2A receptors modulate thalamocortical plasticity and associative learning.. Proc Natl Acad Sci USA, 2016. [PubMed]
- Structural and functional imaging studies in chronic cannabis users: a systematic review of adolescent and adult findings.. PLoS One, 2013. [PubMed]
- Excitatory/inhibitory synaptic imbalance leads to hippocampal hyperexcitability in mouse models of tuberous sclerosis.. Neuron, 2013. [PubMed]
- Hyperpolarization activated cyclic‐nucleotide gated (HCN) channels in developing neuronal networks.. Prog Neurobiol, 2008. [PubMed]
- Deficits in small interneurons in prefrontal and cingulate cortices of schizophrenic and schizoaffective patients.. Arch Gen Psychiatry, 1991. [PubMed]
- Decrease of SYNGAP1 in GABAergic cells impairs inhibitory synapse connectivity, synaptic inhibition and cognitive function.. Nat Commun, 2016. [PubMed]
- Sustained activation of postsynaptic 5‐HT2A receptors gates plasticity at prefrontal cortex synapses.. Cereb Cortex, 2019. [PubMed]
- mTOR in brain physiology and pathologies.. Physiol Rev, 2015. [PubMed]
- Nitric oxide‐dependent long‐term potentiation is blocked by a specific inhibitor of soluble guanylyl cyclase.. Neuroscience, 1995. [PubMed]
- Impaired dendritic expression and plasticity of h‐channels in the fmr1(−/y) mouse model of fragile X syndrome.. Cell Rep, 2012. [PubMed]
- Developmental febrile seizures modulate hippocampal gene expression of hyperpolarization‐activated channels in an isoform‐ and cell‐specific manner.. J Neurosci, 2002. [PubMed]
- Cannabinoid modulation of alpha2 adrenergic receptor function in rodent medial prefrontal cortex.. Eur J Neurosci, 2014. [PubMed]
- A new GABA‐A receptor subtype coupled with Ca++/Cl− synporter modulates aminergic release from rat brain neuron terminals.. J Neurosci Res, 1998. [PubMed]
- Persistently modified h‐channels after complex febrile seizures convert the seizure‐induced enhancement of inhibition to hyperexcitability.. Nat Med, 2001. [PubMed]
- Assessing the role of Ih channels in synaptic transmission and mossy fiber LTP.. Proc Natl Acad Sci USA, 2002. [PubMed]
- PI3K/mTOR mediate mitogen‐dependent HDAC1 phosphorylation in breast cancer: a novel regulation of estrogen receptor expression.. J Mol Cell Biol, 2015. [PubMed]
- 5‐HT(6) receptor and cognition.. Curr Opin Pharmacol, 2011. [PubMed]
- Reduced cortical activity due to a shift in the balance between excitation and inhibition in a mouse model of Rett syndrome.. Proc Natl Acad Sci USA, 2005. [PubMed]
- 5‐HT6 receptor: a new player controlling the development of neural circuits.. ACS Chem Neurosci, 2015. [PubMed]
- Physical interaction between neurofibromin and serotonin 5‐HT6 receptor promotes receptor constitutive activity.. Proc Natl Acad Sci USA, 2016. [PubMed]
- Cannabis and psychosis/schizophrenia: human studies.. Eur Arch Psychiatry Clin Neurosci, 2009. [PubMed]
- Temporal lobe epilepsy after experimental prolonged febrile seizures: prospective analysis.. Brain, 2006. [PubMed]
- Cdk5 induces constitutive activation of 5‐HT6 receptors to promote neurite growth.. Nat Chem Biol, 2014. [PubMed]
- Immunocytochemical distribution of the cannabinoid CB1 receptor in the primate neocortex: a regional and laminar analysis.. Cereb Cortex, 2007. [PubMed]
- Reversal of learning deficits in a Tsc2+/− mouse model of tuberous sclerosis.. Nat Med, 2008. [PubMed]
- Rapamycin for treating tuberous sclerosis and autism spectrum disorders.. Trends Mol Med, 2011. [PubMed]
- The effect of marijuana use on the risk for schizophrenia.. J Clin Psychiatry, 2012. [PubMed]
- Dual 5‐HT6 and D3 receptor antagonists in a group of 1H‐Pyrrolo[3,2‐c]quinolines with neuroprotective and procognitive activity.. ACS Chem Neurosci, 2019. [PubMed]
- Cannabis use and public health: assessing the burden.. Addiction, 2000. [PubMed]
- Neurophysiology of HCN channels: from cellular functions to multiple regulations.. Prog Neurobiol, 2014. [PubMed]
- Long‐term consequences of perinatal and adolescent cannabinoid exposure on neural and psychological processes.. Neurosci Biobehav Rev, 2015. [PubMed]
- Characterization of [(125)I]‐SB‐258585 binding to human recombinant and native 5‐HT(6) receptors in rat, pig and human brain tissue.. Br J Pharmacol, 2000. [PubMed]
- mTOR signaling: at the crossroads of plasticity, memory and disease.. Trends Neurosci, 2010. [PubMed]
- Dysregulation of mammalian target of rapamycin signaling in mouse models of autism.. J Neurosci, 2015. [PubMed]
- Assessment of social interaction behaviors.. J Vis Exp, 2011
- The role of 5 HT6‐receptor antagonists in Alzheimer’s disease: an update.. Expert Opin Investig Drugs, 2018
- Development of the GABAergic system from birth to adolescence.. Neuroscientist, 2012. [PubMed]
- DISC1 regulates new neuron development in the adult brain via modulation of AKT‐mTOR signaling through KIAA1212.. Neuron, 2009. [PubMed]
- Cloning of the mouse 5‐HT6 serotonin receptor and mutagenesis studies of the third cytoplasmic loop.. Brain Res Mol Brain Res, 2001. [PubMed]
- Dendritic lh normalizes temporal summation in hippocampal CA1 neurons.. Nat Neurosci, 1999. [PubMed]
- Adolescent cannabis use and psychosis: epidemiology and neurodevelopmental models.. Br J Pharmacol, 2010. [PubMed]
- Interneuron dysfunction in psychiatric disorders.. Nat Rev Neurosci, 2012. [PubMed]
- Cannabinoid control of learning and memory through HCN channels.. Neuron, 2016. [PubMed]
- 5‐HT(6) receptor recruitment of mTOR as a mechanism for perturbed cognition in schizophrenia.. EMBO Mol Med, 2012. [PubMed]
- Altering the course of schizophrenia: progress and perspectives.. Nat Rev Drug Discov, 2016. [PubMed]
- Adolescent exposure to Delta(9)‐tetrahydrocannabinol alters the transcriptional trajectory and dendritic architecture of prefrontal pyramidal neurons.. Mol Psychiatry, 2019. [PubMed]
- A behavioral role for dendritic integration: HCN1 channels constrain spatial memory and plasticity at inputs to distal dendrites of CA1 pyramidal neurons.. Cell, 2004. [PubMed]
- Repeated cannabinoid exposure during perinatal, adolescent or early adult ages produces similar longlasting deficits in object recognition and reduced social interaction in rats.. J Psychopharmacol, 2006. [PubMed]
- State‐dependent cAMP sensitivity of presynaptic function underlies metaplasticity in a hippocampal feedforward inhibitory circuit.. Neuron, 2008. [PubMed]
- Adolescent THC exposure in female rats leads to cognitive deficits through a mechanism involving chromatin modifications in the prefrontal cortex.. J Psychiatry Neurosci, 2018. [PubMed]
- Cannabinoid modulation of hippocampal long‐term memory is mediated by mTOR signaling.. Nat Neurosci, 2009. [PubMed]
- Dissociation of the pharmacological effects of THC by mTOR blockade.. Neuropsychopharmacology, 2013. [PubMed]
- Creation, expression, and characterization of a constitutively active mutant of the human serotonin 5‐HT6 receptor.. Synapse, 2003. [PubMed]
- 5‐Hydroxytryptamine 6 Receptor (5‐HT6R)‐mediated morphological changes via RhoA‐dependent pathways.. Mol Cells, 2017. [PubMed]
- Long‐term cognitive impairments induced by chronic cannabinoid exposure during adolescence in rats: a strain comparison.. Psychopharmacology, 2013. [PubMed]
- Long‐term consequences of adolescent cannabinoid exposure in adult psychopathology.. Front Neurosci, 2014. [PubMed]
- What can rats tell us about adolescent cannabis exposure? Insights from preclinical research.. Can J Psychiatry, 2016. [PubMed]
- Adolescent cannabinoid exposure induces a persistent sub‐cortical hyper‐dopaminergic state and associated molecular adaptations in the prefrontal cortex.. Cereb Cortex, 2017. [PubMed]
- Reduced AKT/mTOR signaling and protein synthesis dysregulation in a Rett syndrome animal model.. Hum Mol Genet, 2011. [PubMed]
- Cannabis abuse in adolescence and the risk of psychosis: a brief review of the preclinical evidence.. Prog Neuropsychopharmacol Biol Psychiatry, 2013. [PubMed]
- Adolescent exposure to THC in female rats disrupts developmental changes in the prefrontal cortex.. Neurobiol Dis, 2015. [PubMed]
- The impact of exposure to cannabinoids in adolescence: insights from animal models.. Biol Psychiatry, 2016. [PubMed]
- Presynaptic GABAA receptors enhance transmission and LTP induction at hippocampal mossy fiber synapses.. Nat Neurosci, 2010. [PubMed]
- mTOR‐related brain dysfunctions in neuropsychiatric disorders.. Int J Mol Sci, 2018. [PubMed]
- Concomitant THC and stress adolescent exposure induces impaired fear extinction and related neurobiological changes in adulthood.. Neuropharmacology, 2019. [PubMed]
- mTOR, a potential target to treat autism spectrum disorder.. CNS Neurol Disord Drug Targets, 2016. [PubMed]
- Chronic pubertal, but not adult chronic cannabinoid treatment impairs sensorimotor gating, recognition memory, and the performance in a progressive ratio task in adult rats.. Neuropsychopharmacology, 2003. [PubMed]
- The influence of marijuana use on neurocognitive functioning in adolescents.. Curr Drug Abuse Rev, 2008. [PubMed]
- Cortical HCN channels: function, trafficking and plasticity.. J Physiol, 2014. [PubMed]
- Dysregulation of mTOR signaling in fragile X syndrome.. J Neurosci, 2010. [PubMed]
- The adolescent brain and age‐related behavioral manifestations.. Neurosci Biobehav Rev, 2000. [PubMed]
- Early adolescent cannabis exposure and positive and negative dimensions of psychosis.. Addiction, 2004. [PubMed]
- Role of mTOR in physiology and pathology of the nervous system.. Biochim Biophys Acta, 2008. [PubMed]
- Molecular neurobiology of mTOR.. Neuroscience, 2017. [PubMed]
- Loss of mTOR‐dependent macroautophagy causes autistic‐like synaptic pruning deficits.. Neuron, 2014. [PubMed]
- Regulation of neuronal morphology and function by the tumor suppressors Tsc1 and Tsc2.. Nat Neurosci, 2005. [PubMed]
- Refining the role of 5‐HT in postnatal development of brain circuits.. Front Cell Neurosci, 2017. [PubMed]
- Prefrontal cortex HCN1 channels enable intrinsic persistent neural firing and executive memory function.. J Neurosci, 2013. [PubMed]
- Deregulated mTOR‐mediated translation in intellectual disability.. Prog Neurobiol, 2012. [PubMed]
- Gating multiple signals through detailed balance of excitation and inhibition in spiking networks.. Nat Neurosci, 2009. [PubMed]
- GABA(A) receptor‐mediated presynaptic inhibition on glutamatergic transmission.. Brain Res Bull, 2011. [PubMed]
- Presynaptic protein synthesis is required for long‐term plasticity of GABA release.. Neuron, 2016. [PubMed]
- The serotonin‐6 receptor as a novel therapeutic target.. Exp Neurobiol, 2011. [PubMed]
- Alterations of prefrontal cortex GABAergic transmission in the complex psychotic‐like phenotype induced by adolescent delta‐9‐tetrahydrocannabinol exposure in rats.. Neurobiol Dis, 2014. [PubMed]
- PI3K/AKT/mTOR signaling mediates valproic acid‐induced neuronal differentiation of neural stem cells through epigenetic modifications.. Stem Cell Reports, 2017. [PubMed]