
Cannabidiol prevents cognitive and social deficits in a male rat model of Alzheimer’s disease through CB1 activation and inflammation modulation
Abstract
Cognitive decline is a hallmark of Alzheimer’s disease (AD). Cannabidiol (CBD), a non-intoxicating phytocannabinoid with immunomodulatory properties, shows promise in alleviating AD symptoms. This study examined the effects of chronic CBD treatment in a male rat model of sporadic AD induced by intracerebroventricular streptozotocin (ICV-STZ) and explored its impact on neuroinflammatory genes and cannabinoid signaling. STZ rats showed impaired performance in object location and recognition tasks, along with reduced social behavior. STZ exposure also affected AD-related hippocampal markers, leading to increased levels of amyloid β-protein (Aβ) and tau phosphorylation (p-Tau) and elevated mRNA levels of triggering receptor expressed on myeloid cells 2 (TREM2) and apolipoprotein E4 (APOEε4). Additionally, STZ increased hippocampal neuroinflammatory markers, including mRNA levels of Tumor Necrosis Factor α (TNF-α), nuclear factor kappa B subunit 1 (NF-κB1), and interleukin (IL)-1β. It also altered cannabinoid receptor expression, with cannabinoid receptor 1 (cnr1) and 2 (cnr2) genes upregulated in the dentate gyrus (DG), whereas in the CA1, cnr2 was upregulated and cnr1 downregulated. Chronic CBD treatment restored the STZ-induced behavioral deficits, reduced neuroinflammatory marker expression, and mitigated AD-associated changes. Importantly, the CB1 receptor antagonist AM251, but not CB2 antagonist AM630, blocked the beneficial effects of CBD on performance in object location and social tasks in STZ-treated rats, highlighting CB1 receptor activation as a key mechanism. These findings suggest that CBD holds promise as a therapeutic agent for inflammation-induced AD, with the potential to ameliorate cognitive deficits and prevent disease onset through mechanisms involving CB1 receptor activation and modulation of neuroinflammation.
Article type: Research Article
Keywords: Cognitive ageing, Neuroscience
Affiliations: https://ror.org/02f009v59grid.18098.380000 0004 1937 0562School of Psychological Sciences, Department of Psychology, University of Haifa, Haifa, 3498838 Israel; https://ror.org/02f009v59grid.18098.380000 0004 1937 0562The Integrated Brain and Behavior Research Center (IBBRC), University of Haifa, Haifa, 3498838 Israel
License: © The Author(s) 2025 CC BY 4.0 Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
Article links: DOI: 10.1038/s41386-025-02213-0 | PubMed: 40859005 | PMC: PMC12603115
Relevance: Relevant: mentioned in keywords or abstract
Full text: PDF (1.7 MB)
Introduction
Alzheimer’s disease (AD) is marked by progressive cognitive decline, amyloid β-protein (Aβ) accumulation, tau pathology, neuronal death, and brain inflammation, leading to neuropsychiatric symptoms like depression, anxiety, and social withdrawal [ref. 1, ref. 2].
Neuroinflammation, marked by elevated interleukins (IL-1, IL-6), Tumor Necrosis Factor α (TNF-α), and nuclear factor kappa B subunit 1 (NF-kB1), is a central feature of AD; it is frequently found in postmortem AD brains [ref. 3–ref. 5], often preceding Aβ and tau pathology [ref. 6]. Genetic factors like apolipoprotein E4 (APOEε4) and triggering receptor expressed on myeloid cells 2 (TREM2) also regulate inflammatory responses [ref. 7, ref. 8].
The intracerebroventricular (ICV) injection of streptozotocin (STZ) in rats effectively mimics key features of sporadic AD, inducing persistent neuroinflammation, glial activation, reduced hippocampal neurogenesis, and spatial memory deficits [ref. 9–ref. 11]. In transgenic AD models, ICV-STZ exacerbates neuroinflammation, cognitive deficits, plaque pathology, and tau hyperphosphorylation, increasing AD vulnerability [ref. 12–ref. 14]. The ICV-STZ model is induced in mature rats, rather than aged ones, to isolate Alzheimer’s-like symptoms from changes associated with aging [ref. 11].
Cannabidiol (CBD) shows promise in alleviating AD symptoms by immunomodulatory and neuroprotective properties in neurodegenerative diseases [ref. 15–ref. 18]. CBD prevents learning and memory deficits in Aβ-injected mice [ref. 19] and improves spatial learning and anxiety in female APPxPS1 mice [ref. 20].
CBD prevents neuroinflammation through multiple mechanisms, including the reduction of pro-inflammatory markers such as TNF-α, NF-κB1, IL-1β, and inducible nitric oxide synthase [ref. 21, ref. 22]. This effect is partly mediated by inhibiting fatty acid amide hydrolase (FAAH) and modulation of cannabinoid receptors. Specifically, CBD acts as a CB2 partial agonist and a CB1 negative allosteric modulator, influencing receptor function without direct activation [ref. 23, ref. 24].
This study investigated CBD’s potential to prevent cognitive impairments and AD-related markers in the ICV-STZ model. As this is an initial study aimed at investigating the underlying mechanisms of CBD’s preventive effects, only male subjects were used to minimize variability and ensure consistency. Also, Sprague-Dawley (SD) female rats display differences in some AD typical biomarkers compared to those observed in humans [ref. 25].
Additionally, we examined CBD’s effects on hippocampal neuroinflammatory markers and cannabinoid receptor expression, and the role of CB1r and CB2r antagonists (AM251 and AM630) in mediating CBD’s therapeutic outcomes. With an established safety profile, limited side effects, and accessibility, CBD has the potential for rapid clinical translation as a candidate treatment for AD.
Materials and methods
For elaborated procedures see supplementary information.
Animals
Adult male SD rats (Envigo Laboratories, Jerusalem) were used. The study was approved by the University of Haifa Ethics and Animal Care Committee (UoH-IL-2207-164-4) and adhered to NIH guidelines for minimizing pain and discomfort.
Pharmacology
Rats were chronically injected with vehicle, CBD (0.1 mg/kg or 1 mg/kg), the CB1r antagonist AM251 (0.3 mg/kg), or the CB2r antagonist AM630 (1 mg/kg) (Cayman Chemical, USA) daily for two weeks. All drugs were freshly prepared and administered intraperitoneally (i.p.) at a volume of 1 ml/kg. CBD was dissolved in vehicle containing 2% Tween-80 and 98% saline. AM251 and AM630 were prepared in vehicle containing 5% dimethylsulfoxide (DMSO), 5% Tween-80, and 90% saline. Doses were based on previous publications [ref. 26–ref. 29].
Surgical procedure and microinjection
The surgical procedure followed the protocol described in reference (25). Rats were anesthetized with ketamine/domitor (75/0.5 mg/kg, subcutaneously) and placed in a stereotaxic apparatus. On day 0, an ICV injection of STZ (3 mg/kg; 10 μL volume; 2 μL/min) (Sigma-Aldrich, Rehovot, Israel) or aCSF (dissolved in 10 μL DDW) (hello bio, Bristol, UK) was administered into the left ventricle, using a Hamilton syringe (Hamilton Co., USA) (coordinates: AP − 0.8 mm, ML + 1.5 mm, DV − 3.6 mm). The syringe was held for 10 min post-injection to prevent reflux. STZ dosing was based on [ref. 12, ref. 30].
Behavioral tests
The open field (OF) test assessed general locomotor function (total distance, cm) and novelty-induced anxiogenic behavior (time in arena center, first 5 min). The Object Location (OL) and Novel Object Recognition (NOR) tests, with an inter-trial interval (ITI) of 5 min, were used to measure spatial and visual recognition, as well as working memory. We assessed total exploration time (s) and the mean discrimination index (DI), calculated as TN/TN + TF (TN = novel place/object exploration time, TF = familiar place/object exploration time) in the test phase. The Social Interaction Test (SIT) assessed social and non-social behaviors. Time (s) spent on each behavior was measured, and the sociability index was calculated as the time spent engaging in social behaviors divided by the total test time (5 min).
Western blotting (WB)
Protein expression levels of Aβ, Tau, and p-Tau were assessed by Western blot, as previously described [ref. 31]. Protein quantification was performed using a BCA assay, and β-actin served as a loading control.
Quantitative real-time PCR (qRT-PCR)
cDNA preparation and qRT-PCR were performed using standard methodology as previously described [ref. 32, ref. 33]. We examined mRNA gene expression levels of TNF-α, NF-κB1, IL-1β, IL-6, CB1r, CB2r, ApoEɛ4, and TREM2 (Table S1).
Immunohistochemistry procedure
Anesthetized rats were perfused, and brains were fixated and frozen at −80 °C until sectioned with a cooled cryostat (40 μm) (Leica, CM1900). Sections were washed, blocked, and incubated overnight with primary antibodies against p-Tau (pS396) and Aβ. Following PBS washes, Alexa Fluor-conjugated secondary antibodies were applied. Sections were mounted with antifade medium and stored at 4 °C.
Statistical analysis
Results are presented as means ± SEM. Data were analyzed using one-way/two-way ANOVA and Pearson correlation, with post hoc Tukey’s test where applicable. Normality was assessed via Kolmogorov–Smirnov and Shapiro–Wilk tests. Statistical significance was set at p ≤ 0.05, and analysis was performed in SPSS 27 (IBM).
For experimental design see Fig. S1.
Results
Effects of chronic CBD administration on behavior of rats with ICV-STZ-Induced AD
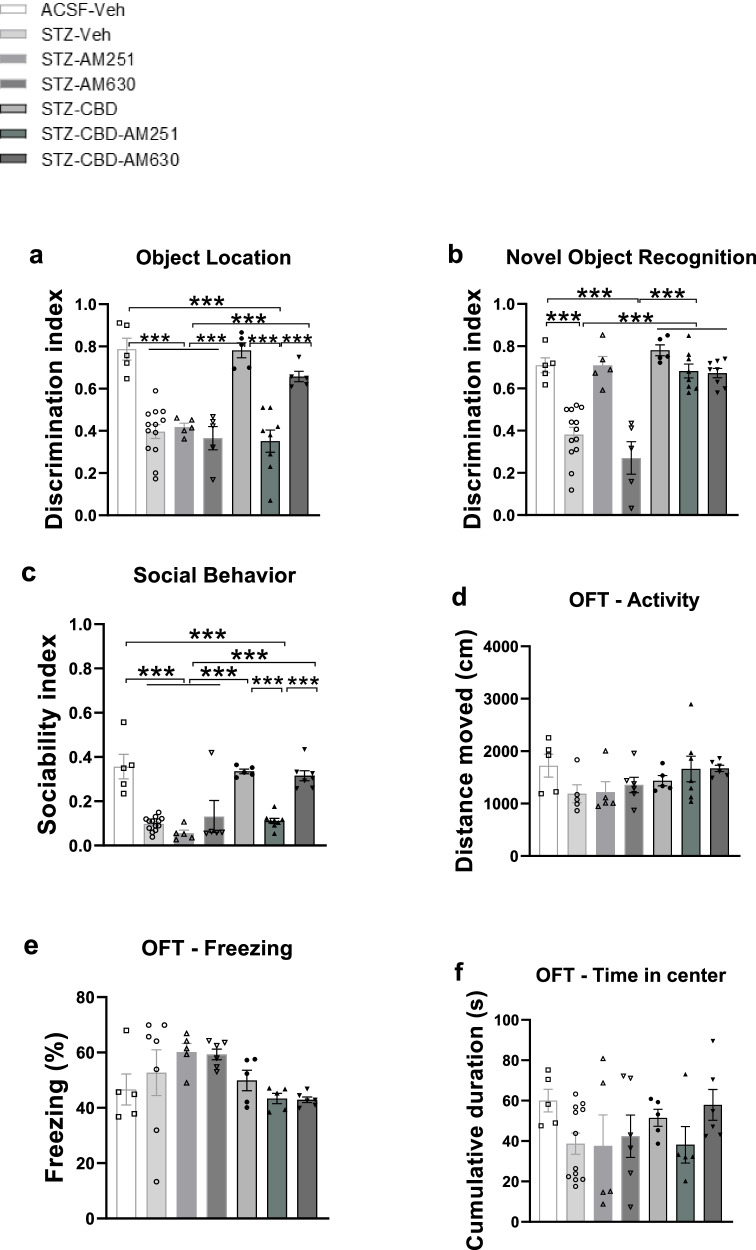
In the OL task (Fig. 1a), a two-way ANOVA revealed significant effects of STZ [F(1,40) = 55.512, p < 0.001], CBD [F(1,40) = 32.249, p < 0.001], and the STZ×CBD interaction [F(1,40) = 31.977, p < 0.001]. Post-hoc comparisons indicated a lower DI in the STZ+Veh group compared to the STZ + CBD (p < 0.01), aCSF+Veh (p < 0.001), and aCSF+CBD (p < 0.001) groups.
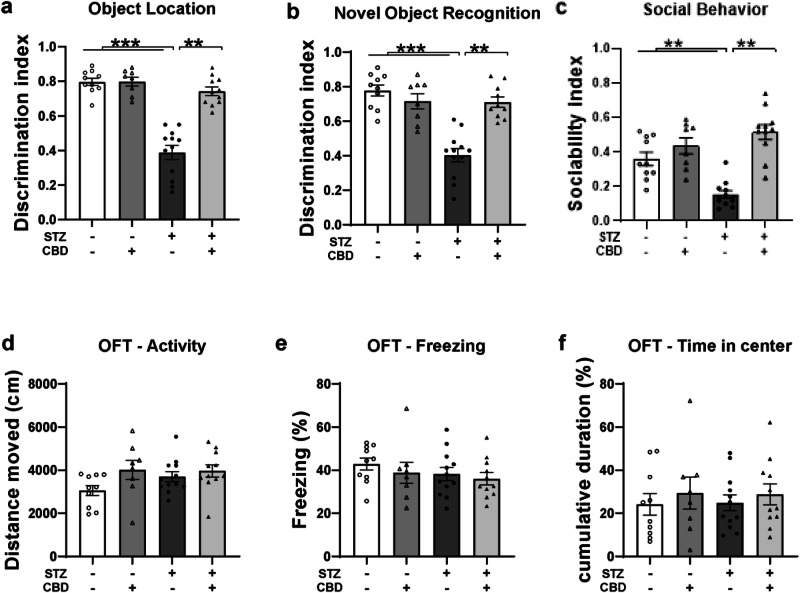
In the NOR task (Fig. 1b), a two-way ANOVA revealed significant effects of STZ [F(1,40) = 26.373, p < 0.001], CBD [F(1,40) = 11.05, p < 0.01], and the STZ×CBD interaction [F(1,40) = 24.936, p < 0.001]. Post-hoc comparisons showed a lower DI in the STZ+Veh group compared to the STZ + CBD (p < 0.01), aCSF+Veh (p < 0.001), and aCSF+CBD (p < 0.001) groups.
We also measured the total exploration time in the cognitive tests (Fig. S2).
In the SIT (Fig. 1c), a two-way ANOVA revealed significant effects of STZ [F(1,40) = 19.697, p < 0.01], CBD [F(1,40) = 34.394, p < 0.001], and the STZ×CBD interaction [F(1,40) = 14.799, p < 0.001]. Post-hoc comparisons showed a lower sociability index in the STZ+Veh group compared to the STZ + CBD (p < 0.01), aCSF+Veh (p < 0.01), and aCSF+CBD (p < 0.01) groups.
These findings suggest that CBD prevented STZ-induced impairment in the OL and NOR tasks, as well as in social behavior. Individual social behaviors are presented in Fig. S3.
In the OFT, no significant effects were observed for distance traveled (Fig. 1d), freezing levels (Fig. 1e), and time spent in the center (Fig. 1f) (see Supplementary Materials for full statistical details).
In a preliminary study, we examined the effect of two low doses of CBD (0.1 mg/kg, 1 mg/kg) on performance in emotional and cognitive tests in STZ-treated animals. We found similar therapeutic effects for both doses (Fig. S4); therefore, we proceeded with the lower dose of 0.1 mg/kg.
Effects of chronic CBD administration on hippocampal AD-related markers in rats with ICV-STZ-Induced AD
We assessed protein levels in the CA1 and DG regions.
Aβ protein and p-Tau
In the CA1, a two-way ANOVA revealed significant effects on Aβ protein levels (Fig. 2a) and p-Tau (Fig. 2c) of STZ [Aβ: F(1,24) = 10.44, p < 0.05; Tau: F(1,24) = 288.79, p < 0.001], CBD [Aβ: F(1,24) = 14.036, p < 0.001; Tau: F(1,24) = 214.423, p < 0.001], and the STZ×CBD interaction [Aβ: F(1,24) = 26.16, p < 0.001; Tau: F(1,24) = 295.315, p < 0.001]. Post-hoc comparisons showed a significant increase in Aβ protein and p-Tau in the STZ+Veh group compared to the STZ + CBD (Aβ: p < 0.001; Tau: p < 0.001), aCSF+Veh (Aβ: p < 0.001; Tau: p < 0.001), and aCSF+CBD (Aβ: p < 0.001; Tau: p < 0.001) groups.
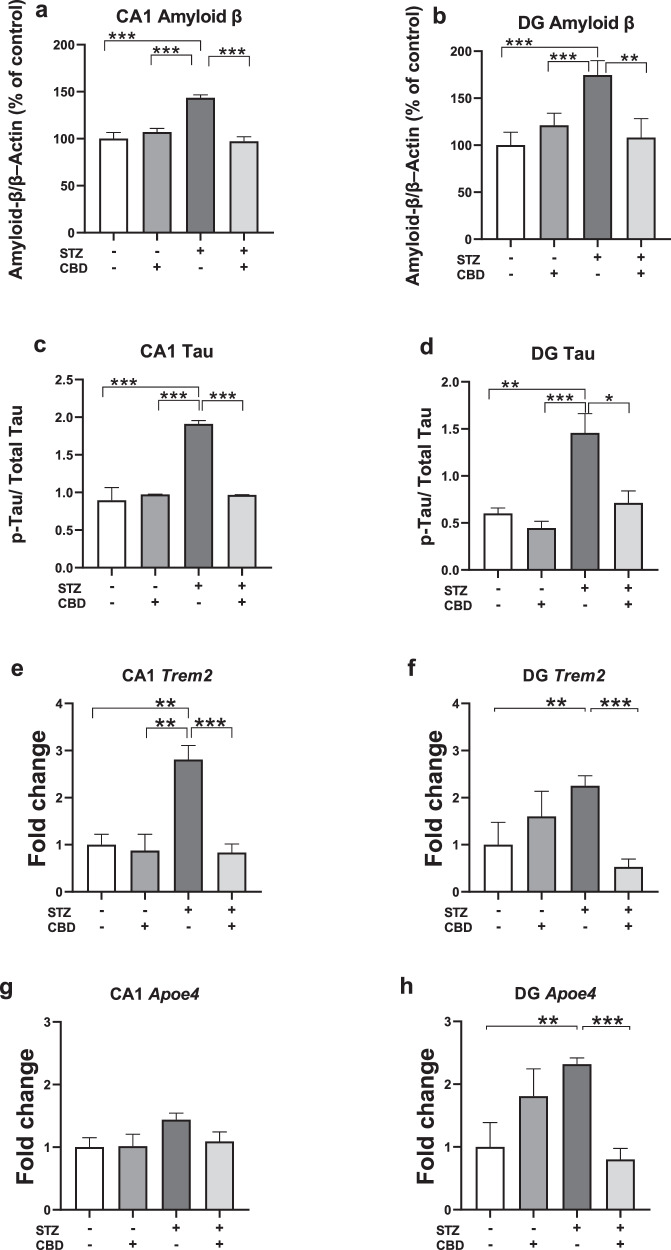
In the DG, a two-way ANOVA revealed significant effects on Aβ protein levels (Fig. 2b) and p-Tau (Fig. 2d) of STZ [Aβ: F(1,28) = 5.276, p < 0.05; Tau: F(1,21) = 34.059, p < 0.001], CBD [Aβ: F(1,28) = 6.137, p < 0.05; Tau: F(1,21) = 19.544, p < 0.001], and the STZ×CBD interaction [Aβ: F(1,28) = 9.861, p < 0.01; Tau: F(1,21) = 6.087, p < 0.05]. Post-hoc comparisons showed a significant increase in Aβ protein and p-Tau in the STZ+Veh group compared to the STZ + CBD (Aβ: p < 0.01; Tau: p < 0.05), aCSF+Veh (Aβ: p < 0.001; Tau: p < 0.01), and aCSF+CBD (Aβ: p < 0.05; Tau: p < 0.001) groups.
These findings suggest that CBD prevented the STZ-induced increase in CA1- and DG-Aβ and CA1- and DG-p-Tau protein levels.
Representative Western blot images are provided in Supplementary Fig. S5.
TREM2
In the CA1, a two-way ANOVA revealed significant effects of STZ [F(1,21) = 7.125, p < 0.05], CBD [F(1,21) = 13.234, p < 0.01], and the STZ×CBD interaction [F(1,21) = 8.497, p < 0.01] (Fig. 2e). Post-hoc comparisons indicated a significant increase in TREM2 mRNA in the STZ+Veh group compared to the STZ + CBD (p < 0.001), aCSF+Veh (p < 0.01) and aCSF+CBD (p < 0.01) groups.
In the DG, a two-way ANOVA revealed a significant effect of STZ×CBD interaction [F(1,20) = 25.503, p < 0.001] (Fig. 2f) with no significant effects of STZ [F(1,20) = 0.079, ns] or CBD [F(1,20) = 2.283, ns]. Post-hoc comparisons showed a significant increase in TREM2 mRNA in the STZ+Veh group compared to the aCSF+Veh (p < 0.01) and STZ + CBD (p < 0.001) groups.
ApoEɛ4
In the CA1, a two-way ANOVA revealed no significant effects of STZ [F(1,21) = 0.1.958, ns], CBD [F(1,21) = 0.701, ns], or the STZ×CBD interaction [F(1,21) = 0.88, ns] (Fig. 2g).
In the DG, two-way ANOVA revealed significant effects of STZ×CBD interaction [F(1,20) = 18.13, p < 0.001] with no significant effect of STZ [F(1,20) = 0.106, ns] or CBD [F(1,20) = 1.612, ns] (Fig. 2h). Post-hoc comparisons showed a significant increase in ApoEɛ4 mRNA in the STZ+Veh group compared to the aCSF+Veh (p < 0.01) and STZ-CBD (p < 0.001) groups.
These findings suggest that CBD prevented the STZ-induced increase in CA1- and DG-TREM2 and DG- ApoEɛ4 mRNA levels.
Pearson bivariate correlation tests were conducted to examine the associations between AD-like behaviors and measures of mRNA and protein levels in the CA1 and DG (Table S2).
Effects of chronic CBD administration on mRNA expression of neuroinflammatory markers and CB1/CB2 receptors in the CA1 and DG of rats with ICV-STZ-Induced AD
TNF-α
In the CA1, a two-way ANOVA revealed significant effects of STZ [F(1,27) = 7.415, p < 0.01], CBD [F(1,27) = 13.908, p < 0.001], and the STZ×CBD interaction [F(1,27) = 6.585, p < 0.05] (Fig. 3a). Post-hoc comparisons showed a significant increase in TNF-α mRNA in the STZ+Veh group compared to the STZ + CBD (p < 0.001), aCSF+Veh (p < 0.01), and aCSF+CBD (p < 0.001) groups.
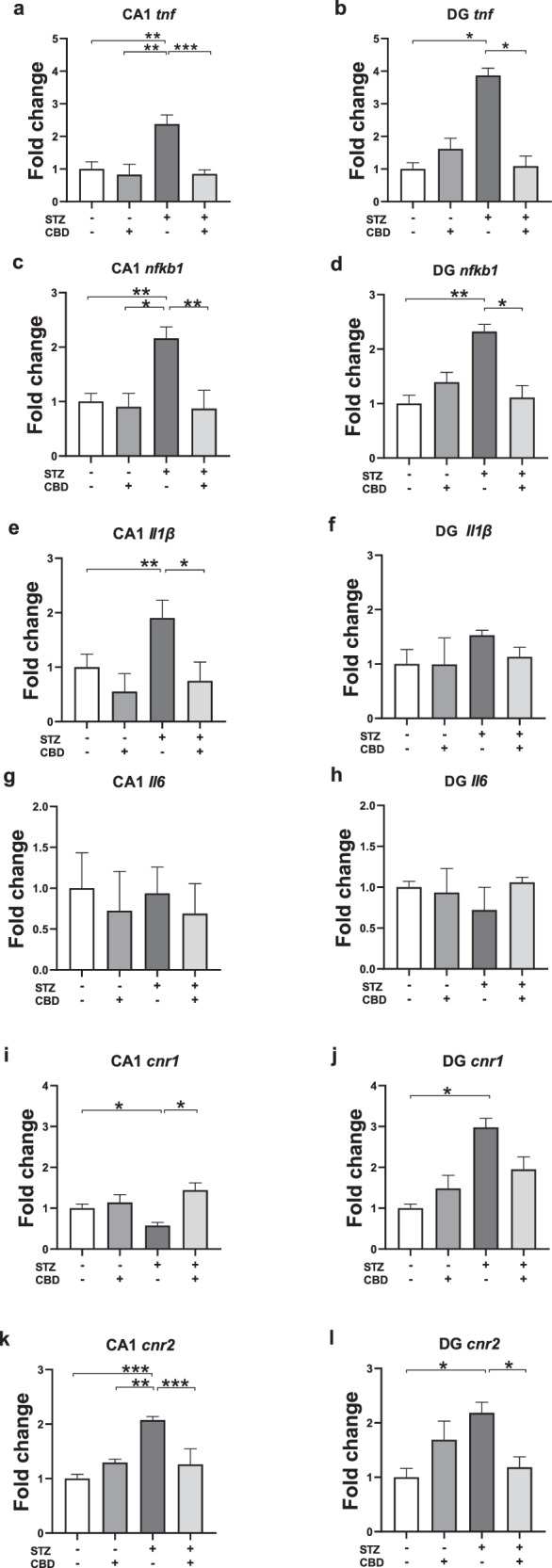
In the DG, a two-way ANOVA revealed significant effects of STZ×CBD interaction [F(1,20) = 12.278, p < 0.01] (Fig. 3b) with no significant effect of STZ [F(1,20) = 3.677, ns] or CBD [F(1,20) = 2.486, ns]. Post-hoc comparisons showed a significant increase in TNF-α mRNA in the STZ+Veh group compared to the STZ + CBD (p < 0.05) and aCSF+Veh (p < 0.05) groups.
These findings suggest that CBD prevented the STZ-induced increase of CA1- and DG-TNF-α.
NF-κB1
In the CA1, a two-way ANOVA revealed significant effects of STZ [F(1,24) = 4.649, p < 0.05], CBD [F(1,24) = 8.858, p < 0.01], and the STZ×CBD interaction [F(1,24) = 5.664, p < 0.05] (Fig. 3c). Post-hoc comparisons showed a significant increase in NF-κB1 mRNA in the STZ+Veh group compared to the STZ + CBD (p < 0.01), aCSF+Veh (p < 0.01), and aCSF+CBD (p < 0.05) groups.
In the DG, a two-way ANOVA revealed a significant effect of STZ×CBD interaction [F(1,20) = 11.668, p < 0.01] (Fig. 3d) with no significant effects of STZ [F(1,20) = 3.901, ns] or CBD [F(1,20) = 1.672, ns]. Post-hoc comparisons showed a significant increase in NF-κB1 mRNA in the STZ+Veh group compared to the STZ + CBD (p < 0.05) and aCSF+Veh (p < 0.05) groups.
These findings suggest that CBD prevented the STZ-induced increase in CA1- and DG-NF-κB1.
IL-1β
In the CA1, a two-way ANOVA revealed significant effects of STZ [F(1,23) = 4.784, p < 0.05], and CBD [F(1,23) = 12.449, p < 0.01], with no significant STZ×CBD interaction [F(1,23) = 0.622, ns] (Fig. 3e). Post-hoc comparisons showed a significant increase in IL-1β mRNA levels in the STZ+Veh group compared to both the STZ + CBD (p < 0.05) and aCSF+Veh (p < 0.01) groups, suggesting that CBD prevented the STZ-induced increase in CA1- IL-1β.
In the DG, a two-way ANOVA revealed no significant effects of STZ [F(1,20) = 0.64, ns], CBD [F(1,20) = 0.196, ns], or the STZ×CBD interaction [F(1,20) = 0.183, ns] (Fig. 3f).
IL-6
In the CA1 (Fig. 3g) and DG (Fig. 3h), a two-way ANOVA revealed no significant effects of STZ [CA1: F(1,21) = 0.000, ns; DG: F(1,20) = 0.034, ns], CBD [CA1: F(1,21) = 0.883, ns; DG: F(1,20) = 0.084, ns], or the STZ×CBD interaction [CA1: F(1,21) = 0.044, ns; DG: F(1,20) = 0.173, ns].
CB1r
In the CA1, a two-way ANOVA revealed significant effects of CBD [F(1,24) = 8.535, p < 0.01] and the STZ×CBD interaction [F(1,24) = 4.441, p < 0.05], with no significant effect of STZ [F(1,24) = 1.633, ns] (Fig. 3i). Post-hoc comparisons showed a significant decrease in CB1r mRNA in the STZ+Veh group compared to the STZ + CBD (p < 0.01) and aCSF+Veh (p < 0.05) groups, suggesting that CBD prevented the STZ-induced decrease in CA1-CB1r.
In the DG, a two-way ANOVA revealed a significant effect of STZ [F(1,20) = 8.199, p < 0.05], with no significant effects of CBD [F(1,20) = 0.003, ns], or STZ×CBD interaction [F(1,20) = 2.939, ns] (Fig. 3j). Post-hoc comparisons indicated a significant increase in CB1r mRNA in the STZ+Veh group compared to the aCSF+Veh group (p < 0.05), suggesting an effect of the STZ model.
CB2r
In the CA1, a two-way ANOVA revealed significant effects of STZ [F(1,26) = 7.798, p < 0.05], CBD [F(1,26) = 5.909, p < 0.05], and the STZ×CBD interaction [F(1,26) = 18.454, p < 0.001] (Fig. 5f). Post-hoc comparisons indicated a significant increase in CB2r mRNA in the STZ+Veh group compared to the STZ + CBD (p < 0.001), aCSF+Veh (p < 0.001), and aCSF+CBD (p < 0.01) groups.
In the DG, a two-way ANOVA revealed a significant effect of STZ×CBD interaction [F(1,20) = 10.758, p < 0.005] (Fig. 3l), with no significant effects of STZ [F(1,20) = 1.514, ns] or CBD [F(1,20) = 0.063, ns]. Post-hoc comparisons indicated a significant increase in CB2r mRNA in the STZ+Veh group compared to the aCSF+Veh (p < 0.05) and STZ + CBD (p < 0.05) groups.
These findings suggest that CBD prevented the STZ-induced increase in CA1- and DG-CB2r.
Pearson bivariate correlation tests were conducted to explore the associations between AD-like behaviors and mRNA expression levels of neuroinflammatory markers and CB1/CB2 genes in the CA1 and DG (Tables S3, 4).
Effects of CB1 and CB2 receptor blockade on behavior following chronic CBD administration in rats with ICV-STZ-induced AD
Given the changes in CB1r and CB2r gene expression, we next examined the impact of receptor blockers on CBD’s therapeutic effects on behavior. The experiment followed the same procedure as the first, with the addition of AM251 (CB1r antagonist) and AM630 (CB2r antagonist), administered with or without CBD. Animals were then tested in the same behavioral assays as previously described (Fig. S1).
In the OL task (Fig. 4a), a one-way ANOVA revealed significant group effects [F(6,42) = 18.49, p < 0.001]. Post-hoc comparisons indicated that the DI was significantly lower in the STZ + CBD + AM251 group compared to the STZ + CBD (p < 0.001), STZ + CBD + AM630 (p < 0.001), and aCSF+Veh (p < 0.001) groups. Additionally, the DI in the STZ + CBD + AM630 group was significantly higher than in the STZ+Veh (p < 0.001), STZ + AM251 (p < 0.001), STZ + AM630 (p < 0.001), and STZ + CBD + AM251 (p < 0.001) groups. These findings suggest that AM251, but not AM630, blocked the effects of CBD on performance in the OL task in STZ rats.
In the NOR task (Fig. 4b), a one-way ANOVA revealed significant group effects [F(6,42) = 22.584, p < 0.001]. Post-hoc comparisons showed that the DI was significantly higher in the STZ + CBD + AM251 and STZ + CBD + AM630 groups compared to the STZ+Veh (p < 0.001) and STZ + AM630 (p < 0.001) groups.
We also measured total exploration time during the cognitive tests (Fig. S6).
In the SIT (Fig. 4c), a one-way ANOVA revealed significant group effects [F(6,42) = 42.968, p < 0.001]. Post-hoc comparisons showed that the sociability index was significantly lower in the STZ + CBD + AM251 group compared to the STZ + CBD (p < 0.001), STZ + CBD + AM630 (p < 0.001), and aCSF+Veh (p < 0.001) groups. Additionally, the sociability index in the STZ + CBD + AM630 group was significantly higher than in the STZ+Veh (p < 0.001), STZ + AM251 (p < 0.001), STZ + AM630 (p < 0.001), and STZ + CBD + AM251 (p < 0.001) groups. Individual social behaviors are presented in Fig. S7.
These findings suggest that in STZ rats, AM251—but not AM630—blocked the effects of CBD in the OL and SIT tasks, while neither antagonist blocked CBD’s effects in the NOR task. Moreover, AM251 alone appeared to enhance NOR performance in STZ rats.
In the OFT, no significant group effects were observed for distance traveled (Fig. 4d), freezing levels (Fig. 4e), or time spent in the center (Fig. 4f) (see Supplementary Materials for full statistical details).
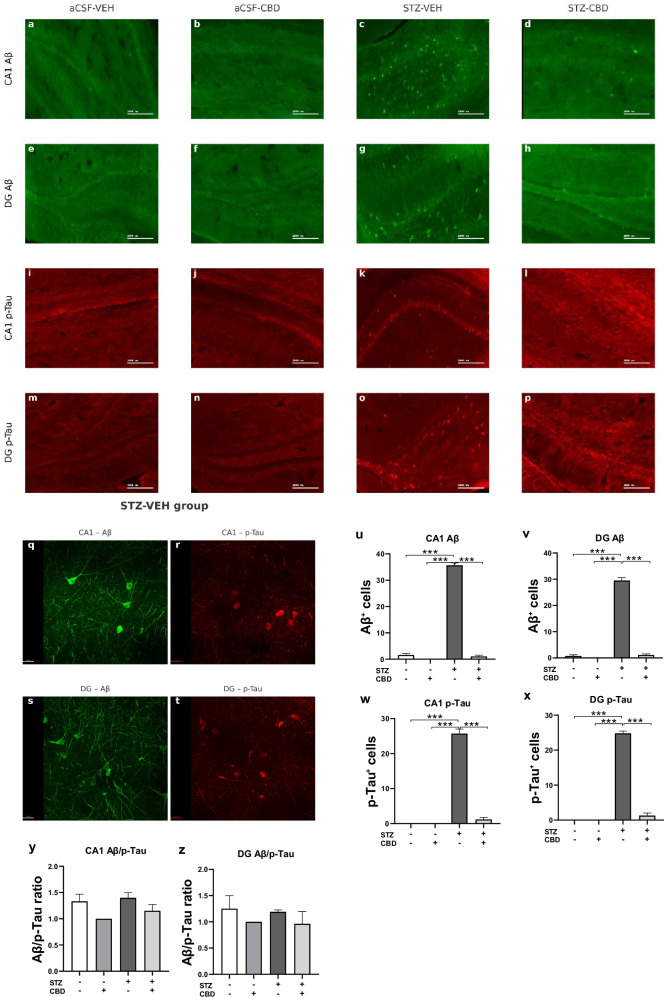
Effects of chronic CBD administration on Aβ and p-Tau accumulation in the hippocampus of rats with ICV-STZ-Induced AD
Immunohistochemical analysis was used to quantify Aβ-positive and p-Tau cells in the CA1 and DG.
A two-way ANOVA conducted on Aβ expression in the CA1 and DG revealed significant effects of STZ [CA1: F(1,16) = 837.537, p < 0.001; DG: F(1,16) = 520.745, p < 0.001], CBD [CA1: F(1,16) = 890.022, p < 0.001; DG: F(1,16) = 496.341, p < 0.001], and the STZ×CBD interaction [CA1:F(1,16) = 741.087, p < 0.001; DG: F(1,16) = 446.594, p < 0.001] (Fig. 5u–v). Consistent with the findings for Aβ, the analysis of p-Tau yielded similar results; a two-way ANOVA conducted on p-Tau expression in the CA1 and DG regions revealed significant effects of STZ [CA1: F(1,16) = 366.313, p < 0.001; DG: F(1,16) = 696.853, p < 0.001], CBD [CA1: F(1,16) = 304.539, p < 0.001; DG: F(1,16) = 565.753, p < 0.001], and the STZ×CBD interaction [CA1: F(1,16) = 304.539, p < 0.001; DG: F(1,16) = 565.753, p < 0.001] (Fig. 5w, x).
Post-hoc analysis revealed a significant increase in Aβ-positive cells in the STZ-Veh group compared to control groups. This increase was attenuated in the STZ-CBD group, with expression levels restored close to near-control values (p < 0.001) in both CA1 and DG regions. Similarly, in both the CA1 and DG regions, STZ-Veh rats exhibited a significantly higher number of p-Tau-positive cells compared to all other groups (p < 0.001).
A two-way ANOVA of the Aβ/p-Tau cell ratio in the CA1 and DG subregions revealed no significant effects of STZ [CA1: F(1,16) = 1.14, ns; DG: F(1,16) = 0.079, ns], CBD [CA1: F(1,16) = 3.14, ns; DG: F(1,16) = 1.909, ns], or STZ×CBD interaction [CA1: F(1,16) = 0.174, ns; DG:F(1,16) = 0.004, ns], suggesting that the Aβ/p-Tau ratio remained stable across both ACSF and STZ conditions, regardless of CBD administration (Fig. 5y, z).
Discussion
We demonstrate that CBD prevents cognitive and social impairments, as well as hippocampal AD-related markers (Aβ accumulation, p-tau, TREM2, and ApoEɛ4) induced by STZ. It also reduces STZ-induced increases in hippocampal pro-inflammatory markers (TNF-α, NFkB1, IL-1β) and alters CB1/CB2 mRNA levels. Blockade of CB1r with AM251, but not CB2r with AM630, prevents CBD’s beneficial effects in object location and social tasks in STZ-treated rats. This suggests that CBD’s protective effects in the STZ AD model are likely due to its impact on CB1r activity.
As an initial step, this study examined the involvement of neuroinflammatory and endocannabinoid markers in the STZ model and was therefore conducted in males. Yet, we plan to investigate CBD treatment in females in future studies. The fact that the STZ model is typically applied only to males is a bias that limits the model’s representativeness [ref. 34].
The effects of CBD on the behavioral phenotypes in STZ rats
Chronic CBD treatment prevents STZ-induced cognitive and social deficits, consistent with prior research showing similar outcomes in various models [ref. 35]. CBD has protective effects in neurodegenerative diseases, potentially reducing AD-related pathology, alleviating AD symptoms, and slowing cognitive decline [ref. 34, ref. 35]. CBD also prevents memory deficits in Aβ-injected mice [ref. 19, ref. 36], improves learning and reduces Aβ accumulation in female APPxPS1 mice [ref. 37, ref. 38], and enhances recognition memory in pharmacological AD rat models [ref. 39].
No differences in anxiety-like behavior or motor function were observed. The rat model used here represents sporadic AD, which mimics late-onset AD. Anxiety and depression are more pronounced in early-onset AD [ref. 40]. Conflicting results on motor function have been reported in AD rodent models with some studies showing no locomotion deficits in the ICV-STZ model [ref. 41, ref. 42].
The effects of CBD on AD-related markers in STZ rats
Chronic CBD treatment reduced STZ-induced increases in Aβ protein levels, p-Tau, TREM2, and ApoEɛ4 mRNA expression, suggesting preventive effects on AD-related markers. While Aβ and tau pathology are key neuropathological hallmarks of AD [ref. 43], and their elevation has been documented in the STZ model [ref. 43–ref. 45], research on CBD’s impact on these markers is limited, mainly to in vitro models [ref. 46] [ref. 47, ref. 48]. Our study provides the first evidence of CBD’s protective effects on Aβ and Tau in the STZ model.
TREM2 and ApoEɛ4 are markers of disease-associated microglia while some AD models report increased TREM2 expression [ref. 49–ref. 51], others show a decrease [ref. 52, ref. 53]. These discrepancies likely reflect TREM’s stage-dependent role in AD, with low levels reducing proinflammatory cytokines in early stages and exacerbating neuroinflammation in later stages [ref. 54], as observed in our study.
Hippocampal expression of AD-related markers was significantly correlated with cognitive and social impairments, as previously suggested in AD and MCI patients [ref. 55, ref. 56] and animal models [ref. 57–ref. 59]. Increased Aβ and p-tau levels in the CA1 and DG regions were associated with both cognitive and social deficits. Additionally, elevated TREM2 and ApoEε4 expression showed region-specific associations—correlating with cognitive deficits in the CA1 and social deficits in the DG. Evidence from rodent studies supports a role for TREM2 in modulating social behavior, likely through its effects on microglial function and synaptic development [ref. 60–ref. 62].
In our study, CBD modulated ApoEɛ4 expression in the DG, with no significant effect in the CA1. The ApoE gene, particularly the ε4 allele, is well-established as a major risk factor for AD, influencing amyloid and tau pathology, cognitive function, neurodegeneration, and immune responses [ref. 63, ref. 64]. However, research on the interaction between ApoE and CBD is limited, with no studies directly examining CBD’s impact on ApoE levels in AD models.
Importantly, our immunohistochemical findings demonstrate that STZ-induced pathology is associated with increased Aβ and p-Tau accumulation in both the CA1 and DG and that CBD treatment reduced the expression of both markers, suggesting a potential neuroprotective effect.
The effects of CBD on neuroinflammation in STZ rats
CBD prevented the STZ-induced increase in pro-inflammatory markers TNF-α and NFkB1 in the CA1 and DG and IL-1β in the CA1, highlighting the role of neuroinflammation in its therapeutic effects. TNF-α is typically low in healthy adults but significantly elevated in AD, where it accelerates cognitive decline [ref. 65–ref. 67]. NF-kB1 regulates pro-inflammatory gene expression and is implicated in inflammatory and neurodegenerative diseases [ref. 68, ref. 69]. IL-1β, a central pro-inflammatory marker in AD, also influences disease risk [ref. 66, ref. 70].
Changes in hippocampal neuroinflammatory markers were correlated with cognitive impairments (CA1: TNF-α, NF-κB1, IL-1β; DG: TNF-α, NF-κB1) and social deficits (CA1: TNF-α,IL-1β; DG: TNF-α, NF-κB1). Negative social interactions increase pro-inflammatory cytokines such as TNF-α [ref. 71], while positive social support reduces them [ref. 72]. Interestingly, while CA1 NF-κB1 mRNA expression was not significantly associated with social behavior, a significant effect was observed in the DG, suggesting a region-specific role of NF-κB1 in modulating social deficits in the STZ model. The DG is increasingly recognized for its role not only in cognitive functions but also in affective and social behavior, including stress regulation, novelty processing, and social memory [ref. 73, ref. 74]. NF-κB signaling in the hippocampus has been linked to inflammatory responses that impair synaptic plasticity and behavior, and its upregulation in the DG may directly affect circuits underlying social behavior [ref. 75].
Clinical studies have linked IL-1β gene variants to cognitive performance in the elderly [ref. 76], and elevated IL-1β levels are associated with impaired social behavior in rats after proinflammatory stress [ref. 77]. The observed negative correlation between IL-1β levels in the CA1 region and both cognitive performance and social behavior aligns with previous studies demonstrating that elevated CA1 IL-1β contributes to hippocampal neuroinflammation, impairs memory function [ref. 78], and promotes inflammation-induced social withdrawal [ref. 79].
IL-1β mRNA expression in the DG was not significantly altered across experimental conditions, reflecting region-specific regulation of IL-1β signaling within the hippocampus. Recent studies have highlighted that, while the DG expresses neuronal IL-1 receptors, its response to IL-1β differs from other hippocampal subregions [ref. 80, ref. 81]. Our study found no effect of STZ on IL-6 levels, consistent with findings that IL-6 is less frequently elevated in AD [ref. 82].
The effects of CBD on CB1 and CB2 genes in STZ rats
In the CA1, CBD prevented the STZ-induced CB1 downregulation and CB2 upregulation, aligning with studies showing decreased CB1 and increased CB2 expression in postmortem AD brain tissues [ref. 83]. CB1r and CB2r play distinct roles in AD, with CB1 deficiency linked to cognitive impairment and exacerbated symptoms in mouse AD models [ref. 84–ref. 86]. CB1r protects against Aβ-induced toxicity, reduces gliosis, and improves spatial memory [ref. 87].
Conversely, research and post-mortem studies of Alzheimer’s patients show increased CB2r expression associated with Aβ and neuroinflammation [ref. 88, ref. 89]. CB2 genetic deletion has been linked to reduced microglia-mediated neuroinflammation and improved spatial memory in mice [ref. 90, ref. 91].
Interestingly, STZ upregulated CB1r mRNA expression in the DG. This opposing pattern may reflect a region-specific compensatory response, as CB1r upregulation in the DG is thought to reduce hyperexcitability and promote resilience under neural activity or damage [ref. 92, ref. 93]. Additionally, inherent differences in endocannabinoid signaling and receptor distribution across hippocampal subregions may further contribute to these distinct expression profiles.
Changes in CA1 cannabinoid receptor expression correlated with impaired cognitive function (CB1 and CB2) and social deficits (CB2). These findings align with studies linking reduced CB1 expression to cognitive dysfunction in Parkinson’s models and AD postmortem brain tissues [ref. 94, ref. 95] and highlighting CB2’s role in cognitive [ref. 96] and social [ref. 97, ref. 98] functions.
The effects of CB1 and CB2 receptor blockade following chronic CBD administration on the behavioral phenotypes in STZ rats
AM251, but not AM630, blocked CBD’s preventive effects on OL and social interaction tasks in STZ rats, though neither affected CBD’s impact on NOR. AM251 alone enhanced performance in STZ rats, consistent with previous studies showing improved NOR performance without affecting social behavior or anxiety [ref. 99–ref. 101]. Its therapeutic effects on memory may relate to the high density of PFC-CB1r [ref. 102, ref. 103]. In AD models, AM251 has been shown to block the protective memory effects of ACEA [ref. 104], impair spatial memory, and increase tau mRNA expression in the Aβ toxin model of AD [ref. 105]. Conversely, it enhances spatial memory in ischemia‐induced cognitive impairment models [ref. 106]. These varied effects likely depend on dosage, model, timing, co-administration, brain region, and age [ref. 103, ref. 107]. In contrast, AM630 consistently impairs cognitive and social functions [ref. 98, ref. 106, ref. 108].
The perirhinal cortex (PRC) plays a pivotal role in object recognition and is implicated in AD [ref. 109]. The STZ model is known to disrupt PRC function, which in turn impairs performance on learning-related tasks [ref. 110]. Also, in aged rats, the expression of CB1r is increased in the PRC and related brain areas [ref. 111]. Hence, the involvement of different brain structures encoding location (e.g., the hippocampus) and recognition (e.g., the PFC and the PRC) memories may account for the differential effects of AM251 on these types of memory [ref. 9].
Low dose CBD administration
The doses of CBD (0.1 or 1 mg/kg) used in this study were relatively low compared to those commonly reported in the literature. Previous studies have employed a wide range of CBD doses from 0.05–0.25 mg/kg [ref. 26], 1 mg/kg [ref. 112] up to 50 mg/kg [ref. 29] in behavioral and biochemical experiments. CBD exhibits a biphasic effect, where low and high doses can lead to different, and sometimes opposing, physiological and behavioral outcomes [ref. 113, ref. 114]. Studies indicate that lower doses of CBD tend to exert anti-inflammatory effects, whereas higher doses are often required for pronounced analgesic effects [ref. 115]. Additionally, dose-dependent interactions with different receptor systems, including CB1, CB2, TRPV1, and 5-HT1A, may contribute to variations in efficacy [ref. 116]. Future studies should further explore the therapeutic window of CBD to optimize its potential benefits and minimize variability in experimental outcomes.
Conclusions
Our findings suggest that CBD protects against STZ-induced cognitive and social deficits, hippocampal neuroinflammation, and AD-related pathology, with CB1r playing a key role in its therapeutic effects. As current AD treatments are limited, our study highlights CBD as a promising candidate, demonstrating for the first time that a low dose can prevent behavioral and molecular deficits in a rodent model of sporadic AD. By targeting neuroinflammation and endocannabinoid pathways, CBD may help prevent cognitive decline and neuropathological changes in AD.
Supplementary Materials
References
- CPC Galts, LEB Bettio, DC Jewett, CC Yang, PS Brocardo, ALS Rodrigues. Depression in neurodegenerative diseases: Common mechanisms and current treatment options. Neurosci Biobehav Rev, 2019. [DOI | PubMed]
- MF Mendez. The relationship between anxiety and Alzheimer’s disease1. J Alzheimers Dis Rep, 2021. [DOI | PubMed]
- FL Heppner, RM Ransohoff, B Becher. Immune attack: the role of inflammation in Alzheimer disease. Nat Rev Neurosci, 2015. [DOI | PubMed]
- C Sala Frigerio, L Wolfs, N Fattorelli, N Thrupp, I Voytyuk, I Schmidt. The major risk factors for Alzheimer’s disease: age, sex, and genes modulate the microglia response to Aβ Plaques. Cell Rep, 2019. [DOI | PubMed]
- PL McGeer, J Rogers, EG McGeer. Inflammation, antiinflammatory agents, and Alzheimer’s disease: the last 22 years. J Alzheimer’s Dis, 2016. [DOI | PubMed]
- E Zotova, V Bharambe, M Cheaveau, W Morgan, C Holmes, S Harris. Inflammatory components in human Alzheimer’s disease and after active amyloid-β42 immunization. Brain, 2013. [DOI | PubMed]
- R Guerreiro, A Wojtas, J Bras, M Carrasquillo, E Rogaeva, E Majounie. TREM2 Variants in Alzheimer’s Disease. N Engl J Med, 2013. [DOI | PubMed]
- A Serrano-Pozo, Z Li, A Noori, HN Nguyen, A Mezlini, L Li. Effect of APOE alleles on the glial transcriptome in normal aging and Alzheimer’s disease. Nat Aging, 2021. [DOI | PubMed]
- TB Bassani, JM Turnes, ELR Moura, JM Bonato, V Cóppola-Segovia, SM Zanata. Effects of curcumin on short-term spatial and recognition memory, adult neurogenesis and neuroinflammation in a streptozotocin-induced rat model of dementia of Alzheimer’s type. Behavi Brain Res, 2017. [DOI]
- 10.Bassani TB, Bonato JM, Machado MMF, Cóppola-Segovia V, Moura ELR, Zanata SM, et al. Decrease in Adult Neurogenesis and Neuroinflammation Are Involved in Spatial Memory Impairment in the Streptozotocin-Induced Model of Sporadic Alzheimer’s Disease in Rats. Mol Neurobiol. 2017. 10.1007/s12035-017-0645-9.
- J Homolak, AB Perhoc, A Knezovic, J Osmanovic Barilar, M Salkovic-Petrisic. Additional methodological considerations regarding optimization of the dose of intracerebroventricular streptozotocin A response to: “Optimization of intracerebroventricular streptozotocin dose for the induction of neuroinflammation and memory impairments in rats” by Ghosh et al., Metab Brain Dis 2020 July 21. Metab Brain Dis, 2021. [DOI | PubMed]
- Y Chen, Z Liang, Z Tian, J Blanchard, C Dai, S Chalbot. Intracerebroventricular streptozotocin exacerbates Alzheimer-Like Changes of 3xTg-AD Mice. Mol Neurobiol, 2014. [DOI | PubMed]
- YD Ke, F Delerue, A Gladbach, J Götz, LM Ittner. Experimental diabetes mellitus exacerbates tau pathology in a transgenic mouse model of Alzheimer’s disease. PLoS ONE, 2009. [DOI | PubMed]
- K Plaschke, J Kopitz, M Siegelin, R Schliebs, M Salkovic-Petrisic, P Riederer. Insulin-Resistant Brain State after Intracerebroventricular Streptozotocin Injection Exacerbates Alzheimer-like Changes in Tg2576 AβPP-Overexpressing Mice. J Alzheimer’s Dis, 2010. [DOI | PubMed]
- S Bhunia, N Kolishetti, AY Arias, A Vashist, M Nair. Cannabidiol for neurodegenerative disorders: A comprehensive review. Front Pharm, 2022. [DOI]
- S Elsaid, S Kloiber, B Le Foll. Effects of cannabidiol (CBD) in neuropsychiatric disorders: A review of pre-clinical and clinical findings. Prog Mol Biol Transl Sci, 2019. [DOI | PubMed]
- MV Fogaça, FMCV Reis, AC Campos, FS Guimarães. Effects of intra-prelimbic prefrontal cortex injection of cannabidiol on anxiety-like behavior: Involvement of 5HT1A receptors and previous stressful experience. Eur Neuropsychopharmacol, 2014. [DOI | PubMed]
- R Li, M Singh. Sex differences in cognitive impairment and Alzheimer’s disease. Front Neuroendocrinol, 2014. [DOI | PubMed]
- AM Martín-Moreno, D Reigada, BG Ramírez, R Mechoulam, N Innamorato, A Cuadrado. Cannabidiol and other cannabinoids reduce microglial activation in vitro and in vivo: relevance to Alzheimer’s disease. Mol Pharm, 2011. [DOI]
- R Chesworth, D Cheng, C Staub, T Karl. Effect of long-term cannabidiol on learning and anxiety in a female Alzheimer’s disease mouse model. Front Pharm, 2022. [DOI]
- G Esposito, C Scuderi, C Savani, L Steardo, D De Filippis, P Cottone. Cannabidiol in vivo blunts β‐amyloid induced neuroinflammation by suppressing IL‐1β and iNOS expression. Br J Pharm, 2007. [DOI]
- S Burstein. Cannabidiol (CBD) and its analogs: a review of their effects on inflammation. Bioorg Med Chem, 2015. [DOI | PubMed]
- A Ligresti, L De Petrocellis, V Di Marzo. From phytocannabinoids to cannabinoid receptors and endocannabinoids: pleiotropic physiological and pathological roles through complex pharmacology. Physiol Rev, 2016. [DOI | PubMed]
- M Tham, O Yilmaz, M Alaverdashvili, MEM Kelly, EM Denovan‐Wright, RB Laprairie. Allosteric and orthosteric pharmacology of cannabidiol and cannabidiol‐dimethylheptyl at the type 1 and type 2 cannabinoid receptors. Br J Pharm, 2019. [DOI]
- D Moreira-Silva, R Vizin, T Martins, T Ferreira, M Almeida, D Carrettiero. Intracerebral injection of streptozotocin to model Alzheimer disease in rats. Bio Protoc, 2019. [DOI | PubMed]
- D De Gregorio, RJ McLaughlin, L Posa, R Ochoa-Sanchez, J Enns, M Lopez-Canul. Cannabidiol modulates serotonergic transmission and reverses both allodynia and anxiety-like behavior in a model of neuropathic pain. Pain, 2019. [DOI | PubMed]
- M García‐Gutiérrez, J Pérez‐Ortiz, A Gutiérrez‐Adán, J Manzanares. Depression‐resistant endophenotype in mice overexpressing cannabinoid CB 2 receptors. Br J Pharm, 2010. [DOI]
- S Fidelman, T Mizrachi Zer-Aviv, R Lange, CJ Hillard, I Akirav. Chronic treatment with URB597 ameliorates post-stress symptoms in a rat model of PTSD. Eur Neuropsychopharmacol, 2018. [DOI | PubMed]
- LE Long, R Chesworth, X-F Huang, IS McGregor, JC Arnold, T Karl. A behavioural comparison of acute and chronic Δ9-tetrahydrocannabinol and cannabidiol in C57BL/6JArc mice. Int J Neuropsychopharmacol, 2010. [DOI | PubMed]
- A Kraska, MD Santin, O Dorieux, N Joseph-Mathurin, E Bourrin, F Petit. In vivo cross-sectional characterization of cerebral alterations induced by intracerebroventricular administration of streptozotocin. PLoS ONE, 2012. [DOI | PubMed]
- T Mizrachi Zer-Aviv, L Islami, PJ Hamilton, EM Parise, EJ Nestler, B Sbarski. Enhancing endocannabinoid signaling via β-catenin in the nucleus accumbens attenuates PTSD- and depression-like behavior of male rats. Biomedicines, 2022. [DOI | PubMed]
- U Bright, I Akirav. Modulation of endocannabinoid system components in depression: pre-clinical and clinical evidence. Int J Mol Sci, 2022. [DOI | PubMed]
- A Portugalov, H Zaidan, I Gaisler-Salomon, CJ Hillard, I Akirav. FAAH Inhibition restores early life stress-induced alterations in PFC microRNAs Associated with depressive-like behavior in male and female rats. Int J Mol Sci, 2022. [DOI | PubMed]
- J Bao, YAR Mahaman, R Liu, J-Z Wang, Z Zhang, B Zhang. Sex differences in the cognitive and hippocampal effects of streptozotocin in an animal model of sporadic AD. Front Aging Neurosci, 2017. [DOI | PubMed]
- G Watt, T Karl. In vivo evidence for therapeutic properties of cannabidiol (CBD) for Alzheimer’s disease. Front Pharm, 2017. [DOI]
- H Li, Y Liu, D Tian, L Tian, X Ju, L Qi. Overview of cannabidiol (CBD) and its analogues: Structures, biological activities, and neuroprotective mechanisms in epilepsy and Alzheimer’s disease. Eur J Med Chem, 2020. [DOI | PubMed]
- D Cheng, JK Low, W Logge, B Garner, T Karl. Chronic cannabidiol treatment improves social and object recognition in double transgenic APPswe/PS1∆E9 mice. Psychopharmacology, 2014. [DOI | PubMed]
- M Coles, G Watt, F Kreilaus, T Karl. Medium-dose chronic cannabidiol treatment reverses object recognition memory Deficits of APPSwe/PS1ΔE9 Transgenic Female Mice. Front Pharm, 2020. [DOI]
- D de Paula Faria, L Estessi de Souza, FL Duran, S de, CA Buchpiguel, LR Britto. Cannabidiol treatment improves glucose metabolism and memory in streptozotocin-induced Alzheimer’s disease rat model: a proof-of-concept study. Int J Mol Sci, 2022. [DOI | PubMed]
- R Botto, N Callai, A Cermelli, L Causarano, I Rainero. Anxiety and depression in Alzheimer’s disease: a systematic review of pathogenetic mechanisms and relation to cognitive decline. Neurol Sci, 2022. [DOI | PubMed]
- TP O’Leary, A Robertson, PH Chipman, VF Rafuse, RE Brown. Motor function deficits in the 12 month-old female 5xFAD mouse model of Alzheimer’s disease. Behav Brain Res, 2018. [DOI | PubMed]
- KG Ravelli, BdosA Rosário, R Camarini, MS Hernandes, LR Britto. Intracerebroventricular streptozotocin as a model of Alzheimer’s disease: neurochemical and behavioral characterization in mice. Neurotox Res, 2017. [DOI | PubMed]
- RJ Bateman, C Xiong, TLS Benzinger, AM Fagan, A Goate, NC Fox. Clinical and biomarker changes in dominantly inherited Alzheimer’s disease. N. Engl J Med, 2012. [DOI | PubMed]
- CL Bitel, C Kasinathan, RH Kaswala, WL Klein, PH Frederikse. Amyloid-β and Tau Pathology of Alzheimer’s disease induced by diabetes in a rabbit animal model. J Alzheimer’s Dis, 2012. [DOI | PubMed]
- 45.Ermiş M, Çiftci G Role of curcumin on beta-amyloid protein, tau protein, and biochemical and oxidative changes in streptozotocin-induced diabetic rats. Naunyn Schmiedebergs Arch Pharmacol. 2024. 10.1007/s00210-024-03231-3.
- T Imamura, YT Yanagihara, Y Ohyagi, N Nakamura, KM Iinuma, R Yamasaki. Insulin deficiency promotes formation of toxic amyloid-β42 conformer co-aggregating with hyper-phosphorylated tau oligomer in an Alzheimer’s disease model. Neurobiol Dis, 2020. [DOI | PubMed]
- S Alali, G Riazi, MR Ashrafi-Kooshk, S Meknatkhah, S Ahmadian, M Hooshyari Ardakani. Cannabidiol inhibits tau aggregation in vitro. Cells, 2021. [DOI | PubMed]
- G Esposito, D De Filippis, R Carnuccio, AA Izzo, T Iuvone. The marijuana component cannabidiol inhibits β-amyloid-induced tau protein hyperphosphorylation through Wnt/β-catenin pathway rescue in PC12 cells. J Mol Med, 2006. [DOI | PubMed]
- R Fol, J Braudeau, S Ludewig, T Abel, SW Weyer, J-P Roederer. Viral gene transfer of APPsα rescues synaptic failure in an Alzheimer’s disease mouse model. Acta Neuropathol, 2016. [DOI | PubMed]
- G Carbajosa, K Malki, N Lawless, H Wang, JW Ryder, E Wozniak. Loss of Trem2 in microglia leads to widespread disruption of cell coexpression networks in mouse brain. Neurobiol Aging, 2018. [DOI | PubMed]
- C Caldeira, C Cunha, AR Vaz, AS Falcão, A Barateiro, E Seixas. Key aging-associated alterations in primary microglia response to beta-amyloid stimulation. Front Aging Neurosci, 2017. [DOI | PubMed]
- TR Jay, AM Hirsch, ML Broihier, CM Miller, LE Neilson, RM Ransohoff. Disease Progression-Dependent Effects of TREM2 Deficiency in a Mouse Model of Alzheimer’s Disease. J Neurosci, 2017. [DOI | PubMed]
- TK Ulland, M Colonna. TREM2 — a key player in microglial biology and Alzheimer disease. Nat Rev Neurol, 2018. [DOI | PubMed]
- L Karanfilian, MG Tosto, K Malki. The role of TREM2 in Alzheimer’s disease; evidence from transgenic mouse models. Neurobiol Aging, 2020. [DOI | PubMed]
- J King-Robson, H Wilson, M Politis. Associations Between Amyloid and Tau Pathology, and Connectome Alterations, in Alzheimer’s Disease and Mild Cognitive Impairment. J Alzheimer’s Dis, 2021. [DOI | PubMed]
- RA Sperling, EC Mormino, AP Schultz, RA Betensky, KV Papp, RE Amariglio. The impact of amyloid‐beta and tau on prospective cognitive decline in older individuals. Ann Neurol, 2019. [DOI | PubMed]
- H Fujiwara, S Takayama, K Iwasaki, M Tabuchi, T Yamaguchi, K Sekiguchi. Yokukansan, a traditional Japanese medicine, ameliorates memory disturbance and abnormal social interaction with anti-aggregation effect of cerebral amyloid β proteins in amyloid precursor protein transgenic mice. Neuroscience, 2011. [DOI | PubMed]
- C Samaey, A Schreurs, S Stroobants, D Balschun. Early Cognitive and Behavioral Deficits in Mouse Models for Tauopathy and Alzheimer’s Disease. Front Aging Neurosci, 2019. [DOI | PubMed]
- YJ Tan, ASL Ng, A Vipin, JKW Lim, RJ Chander, F Ji. Higher Peripheral TREM2 mRNA Levels Relate to Cognitive Deficits and Hippocampal Atrophy in Alzheimer’s Disease and Amnestic Mild Cognitive Impairment. J Alzheimer’s Dis, 2017. [DOI | PubMed]
- F Filipello, R Morini, I Corradini, V Zerbi, A Canzi, B Michalski. The microglial innate immune receptor TREM2 Is required for synapse elimination and normal brain connectivity. Immunity, 2018. [DOI | PubMed]
- Y Tian, X Xiao, W Liu, S Cheng, N Qian, L Wang. TREM2 improves microglia function and synaptic development in autism spectrum disorders by regulating P38 MAPK signaling pathway. Mol Brain, 2024. [DOI | PubMed]
- Y Tian, R Ai, X Xiao, W Liu, S Cheng, X Zhu. Mechanism of the effect of TREM2 on cognitive function in autistic mice. Cell Mol Biol, 2024. [DOI | PubMed]
- L Arnaud, P Benech, L Greetham, D Stephan, A Jimenez, N Jullien. APOE4 drives inflammation in human astrocytes via TAGLN3 repression and NF-κB activation. Cell Rep, 2022. [DOI | PubMed]
- IK Malashenkova, SA Krynskiy, MV Mamoshina, NA Didkovskiy. Apoe gene polymorphism: the impact of APOE4 allele on systemic inflammation and its role in the pathogenesis of Alzheimer’s disease. Med Immunol, 2018. [DOI]
- B Decourt, DK Lahiri, MN Sabbagh. Targeting tumor necrosis factor alpha for Alzheimer’s Disease. Curr Alzheimer Res, 2017. [DOI | PubMed]
- MT Heneka, MJ Carson, J Khoury, El, GE Landreth, F Brosseron. Neuroinflammation in Alzheimer’s disease. Lancet Neurol, 2015. [DOI | PubMed]
- C Holmes, C Cunningham, E Zotova, J Woolford, C Dean, S Kerr. Systemic inflammation and disease progression in Alzheimer disease. Neurology, 2009. [DOI | PubMed]
- S Singh, TG Singh. Role of Nuclear Factor Kappa B (NF-κB) Signalling in neurodegenerative diseases: an mechanistic approach. Curr Neuropharmacol, 2020. [DOI | PubMed]
- E Sun, A Motolani, L Campos, T Lu. The Pivotal Role of NF-kB in the pathogenesis and therapeutics of Alzheimer’s disease. Int J Mol Sci, 2022. [DOI | PubMed]
- F Su, F Bai, Z Zhang. Inflammatory cytokines and alzheimer’s disease: a review from the perspective of genetic polymorphisms. Neurosci Bull, 2016. [DOI | PubMed]
- JJ Chiang, NI Eisenberger, TE Seeman, SE Taylor. Negative and competitive social interactions are related to heightened proinflammatory cytokine activity. Proc Natl Acad Sci, 2012. [DOI | PubMed]
- BN Uchino, R Trettevik, RG Kent de Grey, S Cronan, J Hogan, BRW Baucom. Social support, social integration, and inflammatory cytokines: A meta-analysis. Health Psychol, 2018. [DOI | PubMed]
- MA Kheirbek, LJ Drew, NS Burghardt, DO Costantini, L Tannenholz, SE Ahmari. Differential control of learning and anxiety along the dorsoventral axis of the dentate gyrus. Neuron, 2013. [DOI | PubMed]
- E Tunc-Ozcan, C-Y Peng, Y Zhu, SR Dunlop, A Contractor, JA Kessler. Activating newborn neurons suppresses depression and anxiety-like behaviors. Nat Commun, 2019. [DOI | PubMed]
- EC Dresselhaus, MK Meffert. Cellular specificity of NF-κB function in the nervous system. Front Immunol, 2019. [DOI | PubMed]
- 76.Kicik A, Tuzun E, Erdogdu E, Bilgic B, Tufekcioglu Z, Ozturk-Isik E, et al. Neuroinflammation mediators are reduced in sera of Parkinson? disease patients with mild cognitive impairment. Archi Neuropsychiatry. 2019. 2019. 10.29399/npa.23624.
- ND Broshevitskaya, IV Pavlova, MI Zaichenko. Early proinflammatory stress affects the social behavior of adult rats: effects of sex and the basal level of interleukin 1-beta in the blood. Neurochem J, 2022. [DOI]
- S Dadsetan, T Balzano, J Forteza, A Cabrera-Pastor, L Taoro-Gonzalez, V Hernandez-Rabaza. Reducing peripheral inflammation with infliximab reduces neuroinflammation and improves cognition in rats with hepatic encephalopathy. Front Mol Neurosci, 2016. [DOI | PubMed]
- J Abraham, RW Johnson. Central inhibition of interleukin-1β ameliorates sickness behavior in aged mice. Brain Behav Immun, 2009. [DOI | PubMed]
- C Rincón-López, A Tlapa-Pale, J-S Medel-Matus, J Martínez-Quiroz, JF Rodríguez-Landa, M-L López-Meraz. Interleukin-1β increases neuronal death in the hippocampal dentate gyrus associated with status epilepticus in the developing rat. Neurolía, 2017
- DP Nemeth, X Liu, MC Monet, H Niu, G Maxey, MS Schrier. Localization of brain neuronal IL-1R1 reveals specific neural circuitries responsive to immune signaling. J Neuroinflammation, 2024. [DOI | PubMed]
- A Ng, WW Tam, MW Zhang, CS Ho, SF Husain, RS McIntyre. IL-1β, IL-6, TNF- α and CRP in Elderly Patients with Depression or Alzheimer’s disease: Systematic Review and Meta-Analysis. Sci Rep, 2018. [DOI | PubMed]
- M Solas, PT Francis, R Franco, MJ Ramirez. CB2 receptor and amyloid pathology in frontal cortex of Alzheimer’s disease patients. Neurobiol Aging, 2013. [DOI | PubMed]
- E Aso, P Andrés-Benito, I Ferrer. Genetic deletion of CB1 cannabinoid receptors exacerbates the Alzheimer-like symptoms in a transgenic animal model. Biochem Pharm, 2018. [DOI | PubMed]
- G Bedse, A Romano, S Cianci, AM Lavecchia, P Lorenzo, MR Elphick. Altered expression of the CB1 cannabinoid receptor in the triple transgenic mouse model of Alzheimer’s disease. J Alzheimer’s Dis, 2014. [DOI | PubMed]
- C Stumm, C Hiebel, R Hanstein, M Purrio, H Nagel, A Conrad. Cannabinoid receptor 1 deficiency in a mouse model of Alzheimer’s disease leads to enhanced cognitive impairment despite of a reduction in amyloid deposition. Neurobiol Aging, 2013. [DOI | PubMed]
- A Patricio-Martínez, R Sánchez-Zavaleta, I Angulo-Cruz, L Gutierrez-Praxedis, E Ramírez, I Martínez-García. The Acute Activation of the CB1 Receptor in the hippocampus decreases neurotoxicity and prevents spatial memory impairment in rats lesioned with β-Amyloid 25–35. Neuroscience, 2019. [DOI | PubMed]
- C Benito, E Núñez, RM Tolón, EJ Carrier, A Rábano, CJ Hillard. Cannabinoid CB 2 receptors and fatty acid amide hydrolase are selectively overexpressed in neuritic plaque-associated glia in Alzheimer’s disease brains. J Neurosci, 2003. [DOI | PubMed]
- JP Lopez, A Kos, G Turecki. Major depression and its treatment. Curr Opin Psychiatry, 2018. [DOI | PubMed]
- Y Li, J Kim. CB2 Cannabinoid receptor knockout in mice impairs contextual long-term memory and enhances spatial working memory. Neural Plast, 2016
- A-C Schmöle, R Lundt, S Ternes, Ö Albayram, T Ulas, JL Schultze. Cannabinoid receptor 2 deficiency results in reduced neuroinflammation in an Alzheimer’s disease mouse model. Neurobiol Aging, 2015. [DOI | PubMed]
- S Ruehle, AA Rey, F Remmers, B Lutz. The endocannabinoid system in anxiety, fear memory and habituation. J Psychopharmacol, 2012. [DOI | PubMed]
- SA Wolf, A Bick-Sander, K Fabel, P Leal-Galicia, S Tauber, G Ramirez-Rodriguez. Cannabinoid receptor CB1 mediates baseline and activity-induced survival of new neurons in adult hippocampal neurogenesis. Cell Commun Signal, 2010. [DOI | PubMed]
- J Ceccarini, C Casteels, R Ahmad, M Crabbé, L Van de Vliet, H Vanhaute. Regional changes in the type 1 cannabinoid receptor are associated with cognitive dysfunction in Parkinson’s disease. Eur J Nucl Med Mol Imaging, 2019. [DOI | PubMed]
- JH Lee, G Agacinski, JH Williams, GK Wilcock, MM Esiri, PT Francis. Intact cannabinoid CB1 receptors in the Alzheimer’s disease cortex. Neurochem Int, 2010. [DOI | PubMed]
- CJ Jordan, Z-X Xi. Progress in brain cannabinoid CB2 receptor research: From genes to behavior. Neurosci Biobehav Rev, 2019. [DOI | PubMed]
- JA Komorowska-Müller, KA Ravichandran, A Zimmer, B Schürmann. Cannabinoid receptor 2 deletion influences social memory and synaptic architecture in the hippocampus. Sci Rep, 2021. [DOI | PubMed]
- M Rodríguez-Arias, F Navarrete, MC Blanco-Gandia, MC Arenas, MA Aguilar, A Bartoll-Andrés. Role of CB2 receptors in social and aggressive behavior in male mice. Psychopharmacology, 2015. [DOI | PubMed]
- I Bialuk, MM Winnicka. AM251, cannabinoids receptors ligand, improves recognition memory in rats. Pharm Rep, 2011. [DOI]
- C Guidali, D Viganò, S Petrosino, E Zamberletti, N Realini, G Binelli. Cannabinoid CB1 receptor antagonism prevents neurochemical and behavioural deficits induced by chronic phencyclidine. Int J Neuropsychopharmacol, 2011. [DOI | PubMed]
- P Lunardi, LW de Souza, B dos Santos, B Popik, L de Oliveira Alvares. Effect of the endocannabinoid system in memory updating and forgetting. Neuroscience, 2020. [DOI | PubMed]
- A Egerton, C Allison, RR Brett, JA Pratt. Cannabinoids and prefrontal cortical function: Insights from preclinical studies. Neurosci Biobehav Rev, 2006. [DOI | PubMed]
- M Häring, N Kaiser, K Monory, B Lutz. Circuit specific functions of cannabinoid CB1 receptor in the balance of investigatory drive and exploration. PLoS ONE, 2011. [DOI | PubMed]
- HD Pawar, Y Patil, A Patil, KT Nakhate, YO Agrawal, K Suchal. Cardioprotective effect of CB1 receptor antagonist AM251 against β receptor-stimulated myocardial infarction via modulation of NF-kB signaling pathway in diabetic mice. Heliyon, 2024. [DOI | PubMed]
- 105.Tavakoli-Far B, Zeraati M, Choopani S, Falah P, Darabi P, Mazloom R, et al. A cannabinoid type 1 receptor antagonist impairs spatial memory and increases the tau gene expression in an Animal Model of the Alzheimer’s disease. Arch. Razi Inst 2024;79:935
- MA Mori, E Meyer, FF da Silva, H Milani, FS Guimarães, RMW Oliveira. Differential contribution of CB1, CB2, 5‐HT1A, and PPAR‐γ receptors to cannabidiol effects on ischemia‐induced emotional and cognitive impairments. Eur J Neurosci, 2021. [DOI | PubMed]
- F Higuchi, S Uchida, H Yamagata, N Abe-Higuchi, T Hobara, K Hara. Hippocampal MicroRNA-124 Enhances Chronic Stress Resilience in Mice. J Neurosci, 2016. [DOI | PubMed]
- L Sun, R Dong, X Xu, X Yang, M Peng. Activation of cannabinoid receptor type 2 attenuates surgery-induced cognitive impairment in mice through anti-inflammatory activity. J Neuroinflammation, 2017. [DOI | PubMed]
- K Okada, K Hashimoto, K Kobayashi. Cholinergic regulation of object recognition memory. Front Behav Neurosci, 2022. [DOI | PubMed]
- DA Mandour, MA Bendary, AE Alsemeh. Histological and imunohistochemical alterations of hippocampus and prefrontal cortex in a rat model of Alzheimer like-disease with a preferential role of the flavonoid “hesperidin”. J Mol Histol, 2021. [DOI | PubMed]
- P Liu, DK Bilkey, CL Darlington, PF Smith. Cannabinoid CB1 receptor protein expression in the rat hippocampus and entorhinal, perirhinal, postrhinal and temporal cortices: regional variations and age-related changes. Brain Res, 2003. [DOI | PubMed]
- 112.Schwotzer D, Kulpa J, Gigliotti A, Dye W, Trexler K, Irshad H, et al. Biological response after 14-Day cannabidiol and propylene glycol inhalation in Sprague–Dawley rats. Cannabis Cannabinoid Res. 2024. October. 10.1089/can.2023.0132.
- AW Zuardi. Cannabidiol: from an inactive cannabinoid to a drug with wide spectrum of action. Rev Brasileira de Psiquiatria, 2008. [DOI]
- R Gallily, Z Yekhtin, LO Hanuš. Overcoming the bell-shaped dose-response of cannabidiol by using cannabis extract enriched in cannabidiol. Pharmacy, 2015
- M Machado Bergamaschi, R Helena Costa Queiroz, A Waldo Zuardi, J Alexandre S. Crippa. Safety and side effects of cannabidiol, a cannabis sativa constituent. Curr Drug Saf, 2011. [DOI | PubMed]
- K Iffland, F Grotenhermen. An update on safety and side effects of cannabidiol: a review of clinical data and relevant animal studies. Cannabis Cannabinoid Res, 2017. [DOI | PubMed]