
Anti-inflammatory and analgesic potential of minor cannabinoids in vivo
Abstract
Background:
The cannabis plant produces many bioactive compounds, including the major cannabinoids THC and CBD, and many lesser studied “minor” phytocannabinoids including cannabinol (CBN), cannabichromene (CBC), cannabicyclol (CBL), and cannabigerol (CBG). These compounds are touted for various ailments, including pain, inflammation, and anxiety, but experimental data on their effects are lacking, especially that of CBL, which has yet to be assessed in vivo.
Methods:
To assess in vivo activity, adult male and female C57BL/6J mice were administered each compound and tested repeatedly in the tetrad battery. The potential analgesic effects in chronic pain states were assessed using the lipopolysaccharide (LPS)-induced hindpaw inflammatory pain and chronic constriction injury (CCI) neuropathic pain paradigms. Lastly, to address common psychological comorbidities of pain, CBN, CBL, and CBG were assessed in the tail suspension and marble burying tests.
Results:
Cannabinol (≥ 25 mg/kg) induced classic cannabinoid effects, including acute antinociception. These effects were differentially and partially blocked by selective antagonism of CB1, adenosine A2A, or TRPV1 receptors. CBL (≥ 50 mg/kg) induced hypothermia that was fully blocked by A2A antagonism but had no apparent CB1-mediated activity. LPS-induced edema and paw proinflammatory cytokine levels were reduced by either CBN or CBL (100 mg/kg). CCI-induced cold allodynia was attenuated by either CBN (≥ 50 mg/kg) or CBL (100 mg/kg), but only at high doses that also induce catalepsy and hypothermia. None of these minor cannabinoids displayed anxiolytic- or antidepressant-like activity without concomitant locomotor effects.
Conclusions:
Together, these findings suggest that CBN produces anti-inflammatory effects via cannabinoid receptor-dependent and -independent pathways, whereas CBL acts primarily through CB receptor-independent mechanisms.
Supplementary Information:
The online version contains supplementary material available at 10.1186/s42238-025-00384-7.
Article type: Research Article
Keywords: Minor cannabinoid, Cannabinol, Cannabichromene, Cannabicyclol, Cannabigerol, Pain, Inflammation
Affiliations: https://ror.org/02der9h97grid.63054.340000 0001 0860 4915School of Nursing, University of Connecticut, 231 Glenbrook Rd U-4026, Storrs, CT 06269 USA; https://ror.org/02der9h97grid.63054.340000 0001 0860 4915Department of Psychological Sciences, University of Connecticut, 231 Glenbrook Rd U-4026, Storrs, CT 06269 USA; https://ror.org/040kfrw16grid.411023.50000 0000 9159 4457Department of Neuroscience and Physiology, State University of New York Upstate Medical University, Syracuse, NY USA; https://ror.org/008zs3103grid.21940.3e0000 0004 1936 8278Department of Chemistry, Rice University, Houston, TX USA
License: © The Author(s) 2026 CC BY 4.0 Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
Article links: DOI: 10.1186/s42238-025-00384-7 | PubMed: 41680865 | PMC: PMC12998089
Relevance: Relevant: mentioned in keywords or abstract
Full text: PDF (4.2 MB)
Introduction
The endocannabinoid system regulates many biological processes, including metabolism, sleep, stress, immune function, and neuroendocrine activity (Finn ref. 2010; Lu and Mackie ref. 2021; Svíženská et al. ref. 2008) warranting its attraction as a research target for novel pharmacotherapies. Cannabis plants contain a complex combination of over 550 identified chemical compounds (Ahmed et al. ref. 2015; Rock and Parker ref. 2021), some of which act on the cannabinoid receptors (i.e., CB1 and CB2). Specifically, 125 cannabinoid compounds have been isolated from the cannabis plant (ElSohly and Gul ref. 2014; Radwan et al. ref. 2021). Delta-9-tetrahydrocannabinol (Δ9-THC) and cannabidiol (CBD) are the most abundant and well-studied of these cannabis phytoconstituents and are thus referred to as the ‘major cannabinoids.’
Beyond Δ9-THC and CBD, some of the most prevalent cannabinoids found in cannabis flowers are delta-8-tetrahydrocannabinol (Δ8-THC), cannabigerol (CBG), cannabinol (CBN), and cannabichromene (CBC) (Al Bakain et al. ref. 2020; ElSohly and Gul ref. 2014; Mechoulam ref. 2005). Additionally, cannabicyclol (CBL), cannabinodiol (CBND), cannabielsoin (CBE), and cannabitriol (CBT) may be present in negligible concentrations (Morales et al. ref. 2017). The marketing and popularity of minor cannabinoid products, especially in areas where Δ9-THC use is restricted, far outstrips empirical evidence to support the purported therapeutic effects, or potential physiological mechanisms of these minor cannabinoids.
CBN and CBC, which are natural breakdown products of Δ9-THC and cannabichromenic acid, respectively, have relatively higher in vitro receptor affinity for CB2 over CB1 (De Petrocellis et al. ref. 2011; Morales et al. ref. 2017; Rhee et al. ref. 1997; Rosenthaler et al. ref. 2014). However, the in vivo effects of CBN are inconsistent in the tetrad battery, a classic whole animal screen for CB1 agonist activity that includes tests of catalepsy, antinociception, body temperature, and locomotor activity. For example, CBN reduced locomotor activity but did not induce other tetrad effects in male Swiss mice (El-Alfy et al. ref. 2010), whereas all tetrad behaviors except for catalepsy were observed in male and female ICR mice (Schwarz et al. ref. 2024). These variable results obscure whether CBN induces cannabimimetic effects in vivo, or whether they may occur via CB1. In addition to acting on the cannabinoid receptors, CBN and CBC act on temperature-sensitive TRP channels, including TRPV1, consistent with analgesic potential (Cascio and Pertwee ref. 2014; De Petrocellis et al. ref. 2011). Indeed, CBN attenuates mechanical allodynia in a mouse chemotherapy-induced peripheral neuropathy model (Schwarz et al. ref. 2024). Similarly, CBC induces antinociceptive effects in the mouse tail-flick assay (Davis and Hatoum ref. 1983) attributed to activity at CB1, adenosine A1, and TRPA1, but not TRPV1 receptors (Maione et al. ref. 2011). In contrast, beyond the structure and biosynthetic pathway of CBL, a degrative product of CBC (Crombie et al. ref. 1968; Ferraro and Umstead ref. 2023; Kane et al. ref. 1984), very little is known about this compound. CBL appears to have antiviral effects against SARS-CoV-2 spike protein fusion in Vero cells (Classen et al. ref. 2024), and acts as a potent positive allosteric modulator of 5-HT1A in vitro (Haghdoost et al. ref. 2025). Yet, there are no reports of its pharmacological effects in vivo.
The overall goal of the present study was to determine the cannabimimetic, analgesic, anxiolytic and antidepressive effects of the minor cannabinoids CBN, CBC, and CBL. To this end, each compound was screened for cannabimimetic effects using the tetrad battery, and these effects were challenged via pharmacological blockade of CB1, adenosine A2A, and TRPV1 receptors. The analgesic and anti-inflammatory potential of each compound was assessed in the lipopolysaccharide (LPS) inflammatory pain and chronic constriction injury (CCI) neuropathic pain models. Chronic pain is comorbid with psychiatric conditions (Roughan et al. ref. 2021). Identifying drugs that reduce both physical and emotional aspects of pain would improve treatment outcomes and reduce reliance on polydrug therapy, thereby lowering the risk for side effects and harmful drug interactions. Thus, each compound was assessed using common anxiolytic and antidepressant drug screens.
Materials and methods
Animals
Adult (> 8-weeks-old) male and female C57BL/6J mice (The Jackson Laboratory; Bar Harbor, ME) were group-housed (4–5 per cage) in polysulfone plastic cages (Optimice: C79100PFF) on aspen chip bedding with food and water available ad libitum. All mice were drug- and experimentally-naïve unless explicitly stated otherwise. Mice were housed in a temperature (21 ± 1 °C) and humidity (40 ± 5%) controlled room, on a 12:12 h light: dark cycle (lights on at 0700) in an AAALAC accredited facility. Mice were stratified by sex before random assignment, within each cage, to treatment group, using a random number generator, such that each cage contained mice from at least two different treatment groups (i.e., no cage contains only vehicle- or drug-treated mice). A power analysis of preliminary data, using a power of 0.8, an alpha of 0.05, and medium effect sizes, indicated a required total sample size of 8 mice. All experiments were carried out during the light phase, starting at approximately 0800 by trained technicians who were blinded to treatment conditions. For the locomotor and marble burying assays, test apparatuses were placed inside sound-attenuating test chambers outfitted with a fan and white LED/infrared lighting. The Institutional Animal Care and Use Committee at the University of Connecticut approved all experimental protocols.
Drugs
The minor phytocannabinoids cannabinol (CBN), cannabichromene (CBC), cannabicyclol (CBL), and cannabigerol (CBG) were synthesized by the Sarlah lab, as previously described (Dennis et al. ref. 2022; Yeom et al. ref. 2013) and purified by column chromatography to purity > 95% (HPLC and NMR). CBN and CBL were tested in all experimental paradigms, CBC was tested in all paradigms except the lipopolysaccharide (LPS) and anxiety-/depressive-like behavior experiments, and CBG was only tested in the anxiety-/depressive-like behavior experiments. The selective CB1 receptor antagonist SR141716A (rimonabant), the synthetic CB2-selective antagonist SR144528 (“SR2”), the adenosine A2A receptor antagonist istradefylline, the TRPV1-selective antagonist capsazepine, and the synthetic glucocorticoid dexamethasone were purchased from Cayman Chemical (Ann Arbor, MI). All drugs were dissolved in a vehicle solution of 5% ethanol, 5% Kolliphor EL (Sigma-Aldrich), and 90% normal saline (Kinsey and Cole ref. 2013). All solutions were injected at a volume of 10 µL/g body mass.
Dose selection
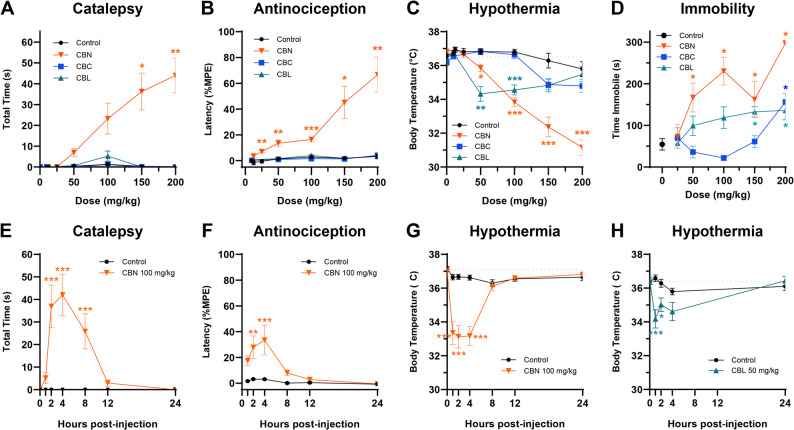
Based on previous literature, we used the highest dose of each antagonist that blocks the action of an agonist without inducing effects per se: rimonabant 3 mg/kg (Schlosburg et al. ref. 2011), istradefylline 3.2 mg/kg (LaVigne et al. ref. 2021), capsazepine 5 mg/kg (Nguyen et al. ref. 2010). In the dose response experiment (Fig. 1), we observed that 200 mg/kg of the minor cannabinoids was not well tolerated, thus doses of ≤ 150 mg/kg were used in subsequent testing.
Tetrad battery
The “tetrad” is a well-characterized battery of four assays used to evaluate the effects of cannabinoid agonists (Lichtman et al. ref. 2001). It consists of assessments of catalepsy, antinociception, core body temperature, and spontaneous locomotor activity. Mice were acclimated to the test room for a minimum of 1 h before testing (Grim et al. ref. 2016; Schlosburg et al. ref. 2009).
- Catalepsy was assessed first by gently laying the forepaws of each mouse over a metal bar elevated 3 cm above the benchtop. Total latency to move a forepaw off the bar was recorded over three trials, with a maximum cutoff of 60 s (Long et al. ref. 2009).
- Antinociception was measured immediately following the catalepsy test by gently placing each mouse headfirst into a small, padded bag and immersing the 1 cm distal tip of the tail into a 56 °C water bath (Falenski et al., 2010). Latency to withdraw the tail from the water was recorded, with a maximum cutoff of 10 s. Tail immersion data is represented as a percent of the maximum possible effect (%MPE; [(measured response – baseline response)/(10 s – baseline response)x100]) (Kaski et al. ref. 2019).
- Hypothermia was assessed immediately following the tail immersion test by taking rectal temperature using a microprobe thermocouple thermometer designed for use with mice (BAT-12 with RET-3 probe, Physitemp Instruments Inc., Clifton, NJ, USA).
- Spontaneous locomotor activity was measured last, by placing individual mice into a transparent plastic test chamber (18 cm W x 28 cm L x 12 cm H) inside a sound-attenuating chamber outfitted with a fan, white LED/infrared lighting, and an overhead video camera. Testing lasted 5 min and locomotor activity was scored in real-time using ANY-maze video tracking software (Stoelting, Wool Dale, IL, USA). Time immobile was determined by setting the tracking parameters to a latency of 1200 ms for 90% of the mouse image pixels (Trexler et al. ref. 2020). In some cases where mice were repeatedly tested (i.e., time course experiment), locomotor activity was not measured, because mice quickly habituate to locomotor testing.
Acute drug effects
To evaluate the dose-dependent acute effects of each compound, 4–5 male and 4–5 female mice (n = 9–10 for each compound) were baselined for catalepsy, tail withdrawal latency in the tail immersion test, and body temperature and then injected (i.p.) with cumulative doses of CBN (12.5, 12.5, 25, 50, 50, & 50 mg/kg), CBC (10, 40, 50, 50, & 50 mg/kg), or CBL (10, 40, 50, 50, & 50 mg/kg) to create a dose-response curve (CBN final doses: 12.5, 25, 50, 100, 150, & 200 mg/kg; CBC/CBL final doses: 10, 50, 100, 150, & 200 mg/kg) 1 h prior to subsequent tetrad testing (i.e., 60 min interval between each dose). Control mice received repeated injections of vehicle. The tetrad dose-response experiment was the only instance where cumulative dosing was used. Locomotor activity was measured in separate groups of naïve mice (i.e., each mouse only tested once; n = 8 for each dose; 128 total) to avoid habituation.
To determine the time course of cannabinol-induced cannabimimetic effects, a separate group of naïve mice (n = 10[5 M/5F] per group) were baseline.
tested for catalepsy, tail withdrawal latency in the tail immersion test, and body temperature and then injected (i.p.) with a single dose of CBN (100 mg/kg) or vehicle and tested repeatedly (1, 2, 4, 8, 12, 24 h post-injection) in a modified tetrad battery.
(i.e., bar test, tail immersion test, and body temperature). This procedure was also used to determine the time course of CBL-induced hypothermia (50 mg/kg, i.p.) wherein only body temperature was repeatedly tested at 0, 1, 2, and 4 h post-injection (n = 10[5 M/5F] per group).
Receptor mechanism challenge
To probe receptor mechanisms by which CBN, CBC, and CBL induce tetrad effects, a separate group of naïve mice (n = 9–10 [4–5 M/4–5 F]) was administered the CB1-selective antagonist rimonabant (3 mg/kg, i.p.), the A2A receptor-selective antagonist istradefylline (3.2 mg/kg) (LaVigne et al. ref. 2021), the TRPV1 antagonist capsazepine (5 mg/kg) (Nguyen et al. ref. 2010), or vehicle 30 min prior to administration (i.p.) of CBN, CBC, CBL (100–150 mg/kg) or vehicle. For the rimonabant and istradefylline challenges, testing was conducted 1 h after administration of the minor cannabinoids or vehicle. A 2 h pretreatment was used for the capsazepine challenge to circumvent the hyperreflexive effects of CBN observed in previous experiments.
Lipopolysaccharide (LPS)-induced paw edema
Inflammatory pain was induced by injecting 25 µg LPS from Escherichia coli 026:B6 (Sigma, St. Louis, MO, USA) in 50 µl of sterile normal saline into the plantar surface of each hind paw (DeLong et al. ref. 2010; Kanaan et al. ref. 1996). Control mice received 50 µl of saline into each hind paw. Paw thickness was measured both before and 24 h after LPS injection, using digital calipers (Mitutoyo, Japan) and expressed to the nearest 0.01 mm. Additionally, thermal preference in a thermal gradient ring were measured 24–26 h post-LPS injection. The synthetic glucocorticoid dexamethasone was used as a positive control. Experimental compounds (i.e., CBN, CBL, or dexamethasone) were injected 1 h before and 6 and 23 h after LPS administration (i.e., 3 injections total). CB1/CB2 antagonists (rimonabant or SR144528) were injected 30 min before each CBN injection.
Thermal Gradient Ring (TGR) test
An automated Thermal Gradient Ring (Ugo Basile, Italy) is an enclosed, conductive ring connecting two hot/cold plates that create a temperature differential across the ring (i.e., 20–40 °C) (Rodriguez et al. ref. 2024; Vanegas et al. ref. 2024). Individual mice were placed in the 23 ± 1 °C zone (i.e., room temperature) on alternating sides of the ring and allowed to freely explore for 16 min. Mouse location and movement were tracked using an overhead camera and analyzed in real-time using ANY-maze software. Dependent variables included the weighted average of temperature/zone occupancy (i.e., “thermal preference”) and distance traveled.
Paw cytokine quantification (Ella)
At the conclusion of all behavioral testing, hind paw soft tissue was collected, snap-frozen in liquid nitrogen, and stored at −80 °C until assay. Samples were homogenized using a Tissue Tearor (Bartlesville, OK) in 1 mL ice-cold phosphate-buffered saline and centrifuged at 4 °C and 7000 g for 15 min. The supernatant was collected and used for quantification of cytokines (IL-1β, IL-6, & TNF-α) in triplicate via Ella multiplex ELISA cartridges (ProteinSimple, Minneapolis, MN, USA). Cytokine levels were standardized to total wet weight (g) of each sample.
Chronic Constriction Injury (CCI) of the sciatic nerve
CCI surgery was performed as described previously (Crowe et al. ref. 2015; Crowe and Kinsey ref. 2017; Grim et al. ref. 2014). Briefly, under isoflurane anesthesia, an incision was made in the skin posterior to the femur, and the sciatic nerve was loosely ligated using a single sterile, 5 − 0 braided silk suture. The surrounding muscle and skin were sutured with sterile 6 − 0 nylon monofilament suture (Ethilon) with a P-1 11 mm 3/8c reverse cutting needle. Allodynia in the paw ipsilateral to CCI was evident 7–10 days post-surgery. Thus, nociceptive testing began no earlier than 10 days after surgery, as previously described (Crowe et al. ref. 2015; Crowe and Kinsey ref. 2017). To reduce animal numbers, a total of 40 (20 M/20F) mice were used in the CCI experiments. Mice received a maximum of three treatments, and a washout period of at least one week was given between treatments.
Mechanical and cold allodynia
Individual mice were placed on an aluminum wire mesh table and acclimated for at least 1 h before measuring cold allodynia in the acetone test. A small, fixed amount (10 µl) of acetone was pipetted onto the plantar surface of each hind paw, after which the total time spent lifting the paw off the mesh surface was recorded using a stopwatch for up to 30s. The average across three trials was used for analyses. Immediately following the acetone test, mechanical allodynia was assessed. The hind paws of each mouse were stimulated with calibrated von Frey filaments using a modified version of the simplified up-down (SUDO) method (Bonin et al. ref. 2014). The plantar surface of each hind paw was stimulated 3–5 times with each filament (0.07–6.0 g), starting with the 1 g filament and increasing or decreasing by two sequential filaments depending on whether the mouse responded by clutching and/or lifting the paw. Paw lifting in response to 3 or more stimulations was coded as a positive response. When there was a change in the response (i.e., paw lift in response to a higher filament or no response to a lighter filament), the next lightest or heaviest filament, respectively, was used to detect the sensory threshold. The paw withdrawal threshold was defined as the thickest filament used that did not elicit a positive response.
Minor cannabinoid effects on anxiety- and depressive-like behaviors
Pain is multifaceted and stress-inducing. Individuals with chronic pain conditions are more likely to develop mental health disorders, including anxiety and depression, which may exacerbate pain perception (Roughan et al. ref. 2021; Vinall et al. ref. 2016). In consideration of this significant comorbidity, the effects of minor cannabinoids were evaluated in the below anxiety- and depressive-like behavioral models. Of note, the effects of CBC (El-Alfy et al. ref. 2010), but not cannabigerol (CBG), have previously been studied in the tail suspension test. CBG increases rat open arm time in the elevated plus maze (Mendiguren et al. ref. 2023). Thus, we determined the effects of CBN, CBL, and CBG in the following screens for anxiolytic and antidepressant drugs.
Tail suspension test
The tail suspension test was performed as previously described (Trexler et al. ref. 2018). Mice were suspended by their tails using adhesive tape attached to a horizontal bar placed approximately 40 cm above the benchtop and video recorded for 6 min. The amount of time the mice actively struggled was hand-scored by a trained and blinded observer. A subset of 6 videos was scored by a second observer to ensure inter-rater reliability (r2 = 0.98). Active struggling was defined as one or more legs repeatedly kicking within one second or arching of the spine. Swinging motions or head movement per se was not scored as struggling. It is important to note that increased immobility while briefly suspended by the tail is associated with higher levels of depressive-like activity, which can be reversed using antidepressants (Steru et al. ref. 1985). Given that mice quickly habituate to this test, separate groups of experimentally naïve mice were used to test each dose (n = 8–10 per dose).
Marble burying test
The marble burying test was performed as a standalone measure as previously described (Broekkamp et al. ref. 1986; Kinsey et al. ref. 2011b; Trexler et al. ref. 2018). Clear, plastic cages (18 cm W x 28 cm L x 12 cm H) filled with 5 cm of Teklad Aspen Sani-Chip bedding (7090 A; Envigo, Indianapolis, IN, USA) and a 4 × 5 array of 20 clear glass marbles was laid gently across the top of the leveled bedding in a grid-like fashion. 60 min following injection of experimental compounds, each mouse was individually placed facing into the front, right-hand corner of the cage and allowed to explore freely for 20 min. At the end of the test, each mouse was quickly and carefully removed, and the number of unburied marbles (≥ 1/3 of the surface showing) was recorded and subtracted from the 20 total marbles. activity was simultaneously recorded for the duration of the test by a camera mounted on the top of the test chamber and analyzed in real time using ANY-maze (Stoelting, Wood Dale, IL, USA). Dependent variables included time immobile and the number of marbles buried. Time immobile was determined by setting the tracking parameters to a latency of 1200 ms for 90% of the mouse image pixels (Trexler et al. ref. 2020).
Data and statistical analyses
Time course and dose-response main effects of each target compound were compared using two-way mixed ANOVA, with drug treatment as a between-subjects variable and time or drug dose as a within-subject variable. Data from the receptor antagonist challenge and LPS experiments were analyzed using one-way ANOVA, followed by Bonferroni post hoc comparisons, with drug treatment as the between-subjects independent variable. Because this study was not designed to analyze sex effects, it was underpowered to detect true sex differences, and thus data are presented collapsed across sexes which may increase variability and mask some effects. All data are represented as mean ± SEM and results were considered significant if p < 0.05.
Results
Cannabimimetic effects of CBN persist for at least 4 h
To assess acute cannabimimetic effects, mice were administered CBN (12.5–200 mg/kg, i.p.), CBC (10–200 mg/kg, i.p.), or CBL (10–200 mg/kg, i.p.) 60 min prior to testing in the tetrad battery. Only CBN produced all four tetrad effects (Table S1). That is, CBN induced catalepsy at ≥ 150 mg/kg (Fig. 1A), antinociception at ≥ 50 mg/kg (Fig. 1B), hypothermia at ≥ 50 mg/kg (Fig. 1C), and immobility at 100 and 200 mg/kg (Fig. 1D). CBC induced immobility at the highest dose tested (200 mg/kg) (Fig. 1D) but had no effect on catalepsy, antinociception, or hypothermia. CBL induced hypothermia at 50–100 mg/kg (Fig. 1C), and immobility at ≥ 150 mg/kg (Fig. 1D), but had no effect on catalepsy or antinociception. Baseline measurements (mean ± SEM) for each test were: bar test (0 ± 0 s), tail immersion test (0.99 ± 0.09 s), body temperature (36.5 ± 0.56 °C), and time immobile (54.65 ± 41.44 s). CBN-induced (100 mg/kg, i.p.) catalepsy (Fig. 1E), antinociception (Fig. 1F), and hypothermia (Fig. 1G) peaked at 4 h and persisted for at least 4–8 h post-injection, whereas CBL-induced hypothermia (50 mg/kg, i.p.) peaked at 1 h and persisted at least 2 h post-injection (Table S2). The time course effects of immobility were not assessed for any of the cannabinoids, given that mice could not be repeatedly tested in the locomotor test.
CBN and CBL act via CB1-dependent and -independent mechanisms
To further probe potential CB1 receptor activity, a separate group of mice was pretreated (i.p.) with the CB1 antagonist rimonabant (3 mg/kg) (Kinsey et al. ref. 2009; Schlosburg et al. ref. 2011) 30 min before CBN (150 mg/kg), CBC (150 mg/kg), or CBL (100 mg/kg) and tested in the tetrad battery. One-way ANOVA revealed a main effect of treatment for each test: catalepsy (Fig. 2A), antinociception (Fig. 2B), hypothermia (Fig. 2C), and immobility (Fig. 2D). Rimonabant pretreatment blocked CBN-induced catalepsy (p < 0.0001, Table S3) and partially blocked the antinociceptive (p = 0.0008 vs. control, p < 0.0001 vs. CBN) and hypothermic effects of CBN (p < 0.0001 vs. control, p = 0.0267 vs. CBN) but had no effect on immobility (p = 0.34) indicating that the tetrad effects of CBN are not completely CB1-mediated. Rimonabant had no effect on CBL-induced hypothermia (p = 0.17) but potentiated CBC-induced immobility (p = 0.0245). CBC also induced hypothermia in this experiment, which was not affected by rimonabant pretreatment (p = 0.13).
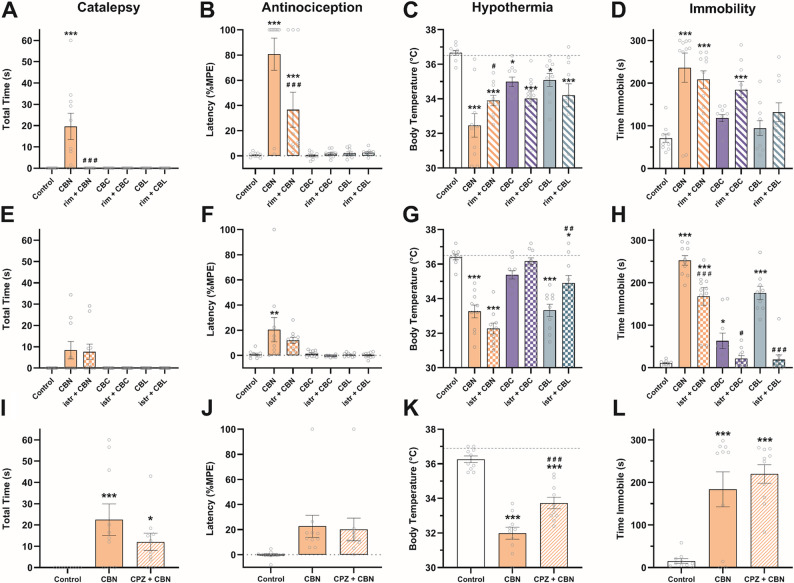
Because the adenosine A2A receptor mediates some tetrad effects of terpenoids found in cannabis (LaVigne et al. ref. 2021), a separate group of mice was pretreated (i.p.) with the A2A antagonist istradefylline (3.2 mg/kg) (LaVigne et al. ref. 2021) 30 min before CBN (100 mg/kg), CBC (100 mg/kg), or CBL (100 mg/kg). Istradefylline had no effect on catalepsy (p = 0.79; Fig. 2E) or antinociception (p = 0.13; Fig. 2F). However, istradefylline increased CBN-induced hypothermia (p = 0.0264; Fig. 2G) and partially blocked CBN-induced immobility (p < 0.0001; Fig. 2H). The hypothermic effects of CBL (p = 0.0005; Fig. 2G) and locomotor effects of CBC and CBL (p = 0.0344 & p < 0.0001; Fig. 2H) were also blocked by istradefylline.
The hypothermic effects of CBN were challenged by pretreatment with the TRPV1 antagonist capsazepine (5 mg/kg, i.p.) (Kinsey et al. ref. 2009; Nguyen et al. ref. 2010) 30 min prior to CBN (100 mg/kg) and tested in the tetrad battery. The main effect of drug treatment for each test are as follows: catalepsy (Fig. 2I), tail immersion (Fig. 2J), core body temperature (Fig. 2K), and time immobile (Fig. 2L). Capsazepine partially blocked CBN-induced hypothermia (p < 0.0001 vs. control, p = 0.0003 vs. CBN) but had no effect on CBN-induced catalepsy (p = 0.14) or immobility (p = 0.35).
CBN blocks LPS-induced paw inflammation and cytokine level increase via cannabinoid receptor-independent mechanisms
To assess anti-inflammatory potential, CBN (100 mg/kg), CBL (100 mg/kg), dexamethasone (2 mg/kg), or vehicle was administered (i.p.) 1 h before, and 6 and 23 h after, injecting lipopolysaccharide (LPS) into both hind paws (Table S4). At 24 h-post LPS, there was a main effect of treatment on paw thickness (Fig. 3A), thermal preference (Fig. 3B), and distance traveled in the thermal gradient ring (Fig. 3C). CBN prevented and CBL reduced LPS-induced paw edema (p < 0.0001). CBN-treated mice preferred the same temperatures as controls but also traveled less in the thermal gradient ring (TGR) (p < 0.0001), which may have increased variance in thermal preference. CBL also reduced distance traveled in the TGR (p < 0.0001) but did not block LPS-induced warmth preference (p > 0.99).
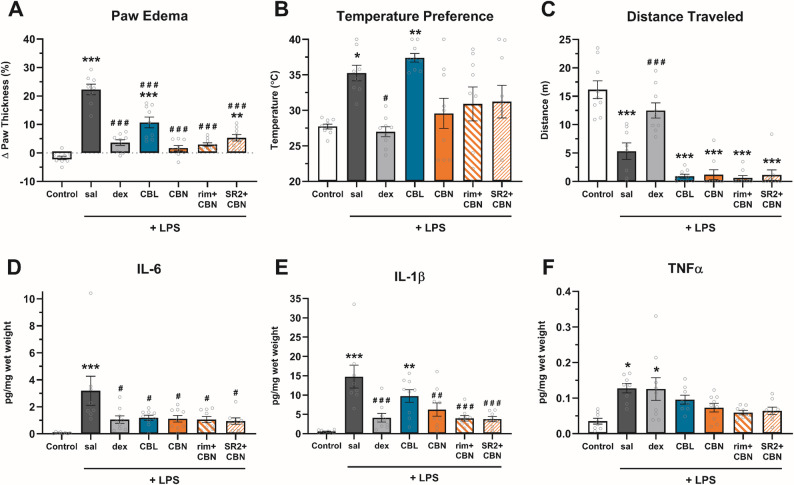
Either CBN (p = 0.0272) or CBL (p = 0.0403) blocked LPS-increased paw levels of IL-6 (Fig. 3D), but only CBN (p = 0.0048) blocked LPS-increased levels of IL-1β (Fig. 3E). There was a main effect of treatment for levels of TNF-α (Fig. 3F), and Bonferroni post hoc analyses revealed that neither CBN (p > 0.99) nor CBL (p = 0.29) differed significantly from controls.
To probe whether the anti-inflammatory effects of CBN are cannabinoid receptor-dependent, two separate groups of CBN-treated mice were pretreated (i.p.) with either rimonabant (3 mg/kg) or SR2 (3 mg/kg) 30 min prior to each CBN injection. SR2-treated mice differed from controls (p = 0.0027), indicating a partial block of CBN-attenuated paw edema (Fig. 3A). Neither rimonabant nor SR2 influenced any CBN effects in other measures.
CBN and CBL reverse CCI-induced cold allodynia at doses that induce hyperreflexia and hypothermia
Chronic neuropathic pain is particularly resistant to existing analgesics. So, in addition to inflammatory pain, we evaluated the analgesic efficacy of minor cannabinoids in a chronic neuropathic pain model. Mice were subjected to chronic constriction injury (CCI) of the sciatic nerve and CBN (25, 50, 100, 150 mg/kg), CBC (10, 30, 60, 100, 150 mg/kg), or CBL (10, 30, 60, 100, 150 mg/kg) was administered (i.p.) to assess acute antiallodynic potential (Table S5). CBN (≥ 100 mg/kg, i.p.) and CBL (≥ 30 mg/kg, i.p.) reduced CCI-increased paw lifting in the cold allodynia test (Fig. 4A). There was a main effect of CBC treatment on acetone-induced allodynia, although Bonferroni post hoc analyses revealed no statistically significant differences of individual doses vs. vehicle. Mechanical allodynia was unaffected by CBN, CBC, or CBL (Fig. 4B). However, it is noteworthy that CBN (≥ 100 mg/kg) also induced hyperreflexia (Fig. 4C), limiting interpretation of the von Frey data.
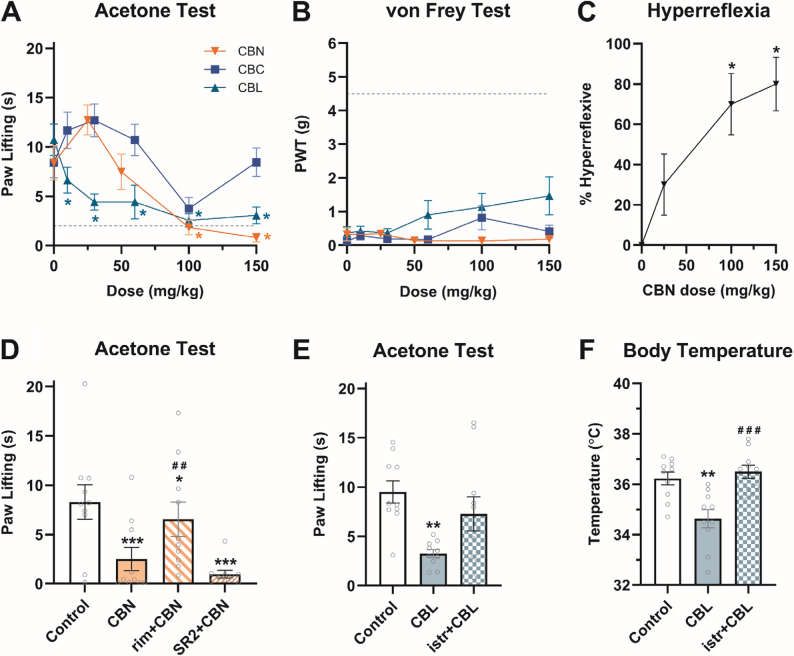
The antiallodynic effects of CBN (100 mg/kg) in the cold allodynia test were blocked by pretreatment (i.p.) of the CB1-selective antagonist rimonabant (3 mg/kg) but not the CB2-selective antagonist SR2 (3 mg/kg) (Fig. 4D). In contrast, the A2A-selective antagonist istradefylline (3.2 mg/kg, i.p.) blocked the antiallodynic effects CBL (30 mg/kg, i.p.) in the acetone-induced cold allodynia test (Fig. 4E). This dose of CBL caused hypothermia in the tetrad, which could interfere with perception of a cold stimulus. Therefore, hypothermia was assessed in these same mice and was also blocked by istradefylline (Fig. 4F). Given that these effects were apparent at high doses, these data suggest that CBN reduces CCI-induced cold allodynia via a CB1 mechanism that overlaps with locomotor deficits, while CBL anti-allodynic effects occur at hypothermic doses that are mediated via A2A receptors.
No effect of minor cannabinoids on anxiety- or depressive-like behaviors
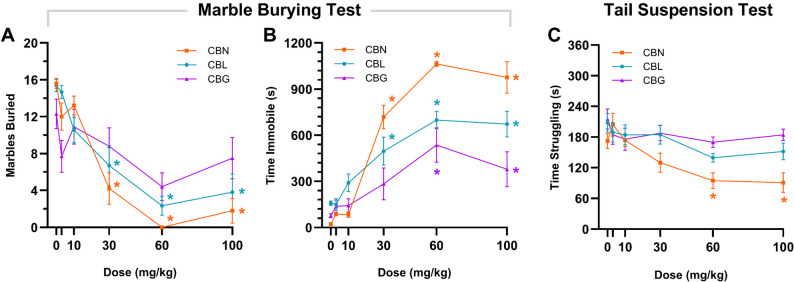
Mice were administered (i.p.) a single dose of CBN, CBL, CBG (3, 10, 30, 60, or 100 mg/kg), or vehicle and then assessed 1 h later in the marble-burying behavior and tail suspension tests. CBN and CBL, but not CBG, reduced the number of marbles buried (Fig. 5 A). However, CBN, CBL, and CBG also increased immobility (Fig. 5B), inconsistent with anxiolysis. CBN dose-dependently reduced struggling in the tail suspension test (Fig. 5 C), but neither CBL nor CBG affected time struggling in the tail suspension test.
Discussion
The goal of the present study was to assess the effects of a subset of minor cannabinoids using established screens for cannabimimetic effects in the tetrad battery, as well as anti-inflammatory and analgesic effects in models of endotoxin-induced inflammatory pain and surgically-induced neuropathic pain. Cannabinol (CBN) was the only minor cannabinoid that induced all four tetrad behaviors, indicative of psychoactive properties consistent with those of CB1 agonists, including the major cannabinoid Δ9-THC. Relatively high doses of CBN (≥ 50 mg/kg) were required to induce THC-like effects in the tetrad, whereas the tetrad effects of Δ9-THC are typically observed at ≥ 10 mg/kg in mice (Vanegas et al. 2022; Wang et al. 2020). Unlike canonical CB1 agonists, these acute effects were only partially and differentially blocked by pretreatment with receptor-selective antagonists of CB1, A2A, or TRPV1. These data align with previous reports that CBN activates both CB1 and TRPV1 receptors in vitro (De Petrocellis et al. ref. 2011; Pertwee & Cascio ref. 2014). Thus, it is likely that CBN activates multiple, as yet to be identified receptor systems to produce the antinociception, hypothermia, and immobility observed in the present study. Given the established role of dopaminergic and adrenergic signaling in motor function and their crosstalk with the endocannabinoid system (Li et al. ref. 2023; Mendiguren et al. ref. 2023; Peters et al. ref. 2021; Zikereya et al. ref. 2025), we speculate that D2 or α−2 receptors are favorable targets for further investigation. In support of this idea, CBN inhibits the release of dopamine and norepinephrine in the rat brain, suggesting potential indirect effects on motor activity (Poddar and Dewey ref. 1980).
It is possible that higher doses of rimonabant, istradefylline, and capsazepine were needed to fully block the effects of the high CBN doses used here. Although the antagonist doses selected are well established as sufficient to block the effects of potent agonists, and higher doses would likely introduce confounding effects of the antagonists themselves (LaVigne et al. ref. 2021; Nguyen et al. ref. 2010; Schlosburg et al. ref. 2011). Additionally, while capsazepine is relatively selective and competitive for TRPV1, it has known activity at other receptors and TRP channels. Future work may therefore benefit from using the more selective TRPV1 antagonist SB-366,791 to challenge the effects of CBN (Varga et al. 2005). Another limitation of this work is that the post-injection interval to observe the effects of CBN in the tetrad needed to be extended from 1 to 2 h to minimize hyperreflexive confounds in the catalepsy test. Because hyperreflexia at the 1-hour time point was intermittent in early experiments, the need for this adjustment only became clear after most studies had been completed. As a result, the optimized 2-hour pretreatment interval was implemented only in the final tetrad experiment (i.e., the capsazepine challenge; Fig. 2I–L). Differences in the pretreatment interval of CBN may also contribute to contrasts between the present data and previous reports that did not observe CBN-induced catalepsy (El-Alfy et al. ref. 2010; Schwarz et al. ref. 2024). Specifically, the previous studies measured catalepsy ≤ 30 min post-injection using the slightly different ring test (versus the bar test used here). Our findings indicate that catalepsy is most pronounced between 2 and 4 h post-CBN administration, which may explain why CBN-induced catalepsy has not been observed previously. It is noteworthy that there were some minor differences between the cumulative dosing and the antagonist challenge experiments. For example, CBC induced hypothermia and increased immobility that was statistically significant only in the latter experiment. We speculate this difference may be due to handling stress in the former or from delivering a large bolus of drug in the latter experiment.CBN blocked both lipopolysaccharide (LPS)-increased paw swelling and proinflammatory cytokine levels. SR144528, but not rimonabant, partially blocked the antiedematous effect of CBN, consistent with a partial CB2 mechanism. These in vivo data are consistent with CBN having higher affinity for CB2 over CB1 in vitro (Pertwee & Cascio ref. 2014). Moreover, CB2 activation reduces inflammation and release of TNF-α, IL-1β, and IL-6 from immune cells (Capozzi et al. ref. 2022; Kinsey et al. ref. 2011a, ref. b; Nass et al. ref. 2021; Tang et al. ref. 2018). However, blocking either CB receptor had no effect on cytokine levels in hindpaw tissue, suggesting the involvement of additional, CB2-independent mechanisms mediating the anti-inflammatory effects of CBN. It is worth noting that a relatively high concentration of LPS was used in the present study, with the goal of inducing not only allodynia but also edema, which may have masked a potential CB2 effect. In further support of this idea, the anti-inflammatory effects of other phytocannabinoids have been linked to multiple parallel receptor systems pathways including peroxisome proliferator-activated receptors (PPARs) (Palomares et al. ref. 2020), GPR55 (Balenga et al. ref. 2011; Yang et al. ref. 2016), 5-HT1A (Landucci et al. ref. 2022), several TRP channels (Muller et al. ref. 2018), cyclooxygenase (COX-1/−2) inhibition (Ruhaak et al. ref. 2011; Takeda et al. ref. 2014), and reduced nitric oxide production (Borrelli et al. ref. 2013; Costa et al. ref. 2007), so it is plausible that CBN also acts through one or more of these receptors in addition to cannabinoid receptors. Similarly, CBC inconsistently produced hypothermic effects, and the hypothermic effect of CBL was fully blocked by istradefylline, so we decided that challenging these compounds with capsazepine was not justified.
We observed acute analgesia with CBN in the tetrad battery, so we next evaluated its efficacy in a chronic pain state. CBN reduced chronic constriction injury (CCI)-induced cold allodynia, but only at doses that also caused catalepsy and hypothermia. The effect of CBN on cold allodynia in the CCI model was blocked by rimonabant, but not SR144528, consistent with a CB1-mediated mechanism. Surprisingly, CBN had no effect on CCI-induced mechanical allodynia, contrasting previous work that found CBN attenuates CIPN-induced mechanical allodynia in mice (Schwarz et al. ref. 2024). Of note, locomotor effects were also observed at the dose used in the study, aligning with the effect of CBN observed here. In the present study, measures of mechanical allodynia were limited by hyperreflexia induced by CBN. Hyperreflexia is decreased muscle control that often precedes catalepsy induced by various cannabinoid agonists. The partial effects of CBN at CB1 are consistent with partial agonism previously reported in vitro (Booker et al. ref. 2009). Regardless of the limitations induced by the observed hyperreflexia, it is unlikely that CBN will reduce mechanical allodynia at doses that do not also induce overt locomotor confounds.
The present study is the first to evaluate the in vivo effects of cannabicyclol (CBL), a degradative product of CBC formed via environmental irradiation (Crombie et al. ref. 1968). In the present study, CBL induced hypothermia and hypolocomotion, each of which was blocked by adenosine A2A receptor antagonism. These data indicate that CBL functions as a novel adenosine A2A agonist in vivo. Similarly, CBL attenuation of CCI-induced cold allodynia occurred through a mechanism requiring A2A activation. It is worth noting that contralateral paw lifting was also positively correlated with body temperature in the CBL-treated group (r2 = 0.56). Therefore, it cannot be ruled out that the observed anti-allodynia may be related to the hypothermic effects of CBL given that these effects were observed at doses that also induced mild hypothermia. That is, we cannot exclude the possibility that the lower body temperature of the mice may have impaired perception of the cold stimulus. Yet, CBL also blocked hind paw inflammation and the increase in paw tissue cytokine levels in the LPS model. A2A receptors have well-established roles in immunity, being expressed on many immune cell types such as neutrophils, eosinophils, monocytes, lymphocytes, epithelial and endothelial cells, and macrophages (Guerrero ref. 2018). It is a strong possibility that the anti-inflammatory effects of CBL could be mediated via A2A activation. Although considerable effort has been devoted to developing adenosine receptor agonists as treatments for a number of diseases, many ultimately fail in clinical trials due to the production of undesirable effects. CBL, on the other hand, may have fewer or milder side effects, but more work is needed to determine its safety and usefulness in human health conditions.
Although CBN reduced cold allodynia, it is noteworthy that none of the minor cannabinoids tested affected CCI-induced mechanical allodynia. Cannabichromene (CBC), which previously exhibited weak antinociception in the tail-flick assay (Maione et al. ref. 2011; Zagzoog et al. ref. 2020), was ineffective here. In contrast, CBC attenuated cisplatin-induced mechanical and cold allodynia, formalin-induced spontaneous pain behaviors, and thermal hyperalgesia in the tail-flick assay (Raup-Konsavage et al. ref. 2024). These discrepancies may reflect mechanistic differences between chemotherapy-induced peripheral neuropathy and CCI. Chemotherapy-induced peripheral neuropathy is a chemically-induced condition characterized by mitochondrial dysfunction, oxidative stress, and peripheral sensory neuron toxicity, whereas CCI is surgically-induced and involves nerve injury and central nociceptive pathways (Jali et al. ref. 2024). Additional differences in experimental design, including route of administration, timing of treatment, or pain stimulus modality, may also contribute to the divergent effects of CBC. For example, one study delivered microinjections of CBC into the ventrolateral periaqueductal grey and measured tail-flick latency in response to radiant heat (Maione et al. ref. 2011), whereas another study measured tail withdrawal latency from 52 °C water 15 min after i.p. injection (Zagzoog et al. ref. 2020). Thus, it is plausible that CBC-induced analgesia is dependent upon local administration, a shorter incubation period, or type of thermal stimuli. Furthermore, the lack of efficacy in the CCI model observed here may suggest that CBC is more effective at treating chemically-induced peripheral sensitization than physical injury accompanied by central neuroimmune processes.
In the marble burying test, either CBN or CBL reduced the number of marbles buried (Thomas et al. ref. 2009) (i.e., a proxy measure of digging behavior) but also induced immobility at the same doses, confounding any clear conclusion about potential anxiolysis. That is, CBN- and CBL-treated mice may have dug less during the test due to reduced locomotor activity. These data are consistent with previously reported effect of Δ9-THC in depression of both marble burying and locomotor activity (Kinsey et al. ref. 2011a, ref. b). Similarly, none of the minor cannabinoids tested displayed anti-depressive-like effects in the tail suspension test, even at relatively high doses. CBG increased the time male Sprague-Dawley rats spent in the open arms of the elevated plus maze (Mendiguren et al. ref. 2023), interpreted as an anxiolytic-like effect. The elevated plus maze was not used here, due to its susceptibility to confound by myriad environmental factors (Shoji and Miyakawa ref. 2021). For instance, increasing endocannabinoid levels via FAAH deletion increases time in the open arms, but this effect is dependent on test room lighting (Naidu et al. ref. 2007). Regardless, the lack of an antidepressant-like effect in the present study aligns with a previous report that neither CBN nor CBG increases time struggling in the forced swim test (El-Alfy et al. ref. 2010), a model of learned helplessness.
In the present study, no drug x sex interactions were observed in our preliminary analyses, with two exceptions. Female mice treated with CBL buried fewer marbles [F(5,46) = 2.55; p = 0.0407], and male mice treated with CBN struggled more in the tail suspension test [F(5,35) = 2.73; p = 0.0347]. These data may be interpreted as relative sex-based differences in sensitivity to either compound’s hypolocomotive effects but do not indicate a clear anxiolytic-like or anti-depressant-like pattern. Given the lack of a clear pattern across tests and the small sample size (n = 5/sex), we have reservations about the biological relevance of these potentially spurious results. Future studies powered to detect sex differences are needed.
Click or tap here to enter text.Click or tap here to enter text.Click or tap here to enter text.Availability and use of minor cannabinoid isolates have grown exponentially in recent years, although there remains a paucity of data evaluating their therapeutic efficacy. The antiedematous and anti-inflammatory potential of cannabinol and cannabicyclol reported here are encouraging, but much additional study, particularly regarding safety, is needed before appropriate recommendations can be made for any clinical application in people. Furthermore, CBN may be limited by the induction of psychoactive effects at high doses. As with many plant-derived compounds, these chemicals activate multiple different receptor systems and may have unpredictable effects, especially at higher doses or after repeated administration. For example, although some of the effects of CBN were partially blocked by CB1, adenosine A2A, or TRPV1 receptor antagonism, none of these receptors appear to fully mediate the antinociceptive, hypothermic, or locomotor effects of CBN. Thus, the potential side effect and drug interaction profile of minor cannabinoids is yet to be established, and more work is needed to uncover the pharmacological activity and molecular targets of these minor constituents found in cannabis.
Supplementary Materials
References
- SA Ahmed, SA Ross, D Slade, MM Radwan, IA Khan, MA ElSohly. Minor oxygenated cannabinoids from high potency Cannabis sativa L. Phytochemistry, 2015. [DOI | PubMed]
- RZ Al Bakain, YS Al-Degs, JV Cizdziel, MA Elsohly. Comprehensive classification of USA cannabis samples based on chemical profiles of major cannabinoids and terpenoids. J Liq Chromatogr Relat Technol, 2020. [DOI]
- NAB Balenga, E Aflaki, J Kargl, W Platzer, R Schröder, S Blättermann, E Kostenis, AJ Brown, A Heinemann, M Waldhoer. GPR55 regulates cannabinoid 2 receptor-mediated responses in human neutrophils. Cell Res, 2011. [DOI | PubMed]
- RP Bonin, C Bories, Y De Koninck. A simplified up-down method (SUDO) for measuring mechanical nociception in rodents using von Frey filaments. Mol Pain, 2014. [DOI | PubMed]
- L Booker, PS Naidu, RK Razdan, A Mahadevan, AH Lichtman. Evaluation of prevalent phytocannabinoids in the acetic acid model of visceral nociception. Drug Alcohol Depend, 2009. [DOI | PubMed]
- F Borrelli, I Fasolino, B Romano, R Capasso, F Maiello, D Coppola. Beneficial effect of the non-psychotropic plant cannabinoid cannabigerol on experimental inflammatory bowel disease. Biochem Pharmacol, 2013. [DOI | PubMed]
- CL Broekkamp, HW Rijk, D Joly-Gelouin, KL Lloyd. Major tranquillizers can be distinguished from minor tranquillizers on the basis of effects on marble burying and swim-induced grooming in mice. Eur J Pharmacol, 1986. [DOI | PubMed]
- Capozzi A, Caissutti D, Mattei V, Gado F, Martellucci S, Longo A, Recalchi S, Manganelli V, Riitano G, Garofalo T, Sorice M, Manera C, Misasi R. Anti-inflammatory activity of a cb2 selective cannabinoid receptor agonist: signaling and cytokines release in blood mononuclear cells. Molecules. 2022;27(1). 10.3390/molecules27010064.
- N Classen, T Pitakbut, M Schöfbänker, J Kühn, ER Hrincius, S Ludwig, A Hensel, O Kayser. Cannabigerol and cannabicyclol block SARS-CoV-2 cell fusion. Planta Med, 2024. [DOI]
- Cham, Switzerland Rock EM, Parker LA. Constituents of Cannabis Sativa. In: Murillo-Rodriguez E, Pandi-Perumal SR, Monti JM. (eds) Cannabinoids and Neuropsychiatric Disorders. Advances in Experimental Medicine and Biology, vol 1264. Springer, Cham. 2021. 10.1007/978-3-030-57369-0_1.
- B Costa, AE Trovato, F Comelli, G Giagnoni, M Colleoni. The non-psychoactive cannabis constituent cannabidiol is an orally effective therapeutic agent in rat chronic inflammatory and neuropathic pain. Eur J Pharmacol, 2007. [DOI | PubMed]
- L Crombie, R Ponsford, A Shani, B Yagnitinsky, R Mechoulam. Hashish components. Photochemical production of cannabicyclol from cannabichromene. Tetrahedron Lett, 1968. [DOI]
- MS Crowe, SG Kinsey. MAGL inhibition modulates gastric secretion and motility following NSAID exposure in mice. Eur J Pharmacol, 2017. [DOI | PubMed]
- MS Crowe, E Leishman, ML Banks, R Gujjar, A Mahadevan, HB Bradshaw. Combined inhibition of monoacylglycerol lipase and cyclooxygenases synergistically reduces neuropathic pain in mice. Br J Pharmacol, 2015. [DOI | PubMed]
- WM Davis, NS Hatoum. Neurobehavioral actions of cannabichromene and interactions with ∆9-tetrahydrocannabinol. Gen Pharmacol, 1983. [DOI | PubMed]
- L De Petrocellis, A Ligresti, AS Moriello, M Allarà, T Bisogno, S Petrosino. Effects of cannabinoids and cannabinoid-enriched Cannabis extracts on TRP channels and endocannabinoid metabolic enzymes. Br J Pharmacol, 2011. [DOI | PubMed]
- GT DeLong, CE Wolf, A Poklis, AH Lichtman. Pharmacological evaluation of the natural constituent of cannabis sativa, cannabichromene and its modulation by ∆9-tetrahydrocannabinol. Drug Alcohol Depend, 2010. [DOI | PubMed]
- DG Dennis, SD Anand, AJ Lopez, J Petrovčič, A Das, D Sarlah. Synthesis of the cannabimovone and cannabifuran class of minor phytocannabinoids and their anti-inflammatory activity. J Org Chem, 2022. [DOI | PubMed]
- AT El-Alfy, K Ivey, K Robinson, S Ahmed, M Radwan, D Slade. Antidepressant-like effect of Δ9-tetrahydrocannabinol and other cannabinoids isolated from Cannabis sativa L.. Pharmacol Biochem Behav, 2010. [DOI | PubMed]
- ElSohly MA, Gul W. Constituents of cannabis sativa. In: Pertwee R, editor. Handbook of cannabis. Oxford University Press; 2014. pp. 3–22. 10.1093/acprof:oso/9780199662685.003.0001.
- JM Ferraro, WJ Umstead. Chiral separation of cannabichromene, cannabicyclol, and their acidic analogs on polysaccharide chiral stationary phases. Molecules (Basel, Switzerland), 2023. [DOI]
- DP Finn. Endocannabinoid-mediated modulation of stress responses: physiological and pathophysiological significance. Immunobiology, 2010. [DOI | PubMed]
- TW Grim, S Ghosh, KL Hsu, BF Cravatt, SG Kinsey, AH Lichtman. Combined inhibition of FAAH and COX produces enhanced anti-allodynic effects in mouse neuropathic and inflammatory pain models. Pharmacol Biochem Behav, 2014. [DOI | PubMed]
- TW Grim, KL Samano, B Ignatowska-Jankowska, Q Tao, LJ Sim-Selly, DE Selley, LE Wise, A Poklis, AH Lichtman. Pharmacological characterization of repeated administration of the first generation abused synthetic cannabinoid CP47,497. J Basic Clin Physiol Pharmacol, 2016. [DOI | PubMed]
- A Guerrero. A2A adenosine receptor agonists and their potential therapeutic applications. An update. Curr Med Chem, 2018. [DOI | PubMed]
- M Haghdoost, Y DePorre, M Figi, S Young, C Krebs, MO Bonn-Miller. An unexpected activity of a minor cannabinoid: cannabicyclol (CBL) is a potent positive allosteric modulator of serotonin 5-HT1A receptor. J Nat Prod, 2025. [DOI | PubMed]
- AM Jali, D Banji, OJF Banji, KY Hurubi, FY Tawhari, AA Alameer, AS Dohal, RA Zanqoti. Navigating preclinical models and medications for peripheral neuropathy: a review. Pharmaceuticals, 2024. [DOI]
- SA Kanaan, NE Saadé, JJ Haddad, AM Abdelnoor, SF Atweh, SJ Jabbur, B Safieh-Garabedian. Endotoxin-induced local inflammation and hyperalgesia in rats and mice: A new model for inflammatory pain. Pain, 1996. [DOI | PubMed]
- VV Kane, AR Martin, JA Peters. Carbon-13 nuclear magnetic resonance spectra of Cannabichromene, Cannabicitran, and Cannabicyclol and their analogues. J Org Chem, 1984. [DOI]
- SW Kaski, AN White, JD Gross, KR Trexler, K Wix, AA Harland, TE Prisinzano, J Aubé, SG Kinsey, T Kenakin, DP Siderovski, V Setola. Preclinical testing of nalfurafine as an opioid-sparing adjuvant that potentiates analgesia by the mu opioid Receptor-targeting agonist morphine. J Pharmacol Exp Ther, 2019. [DOI | PubMed]
- SG Kinsey, EC Cole. Acute ∆9-tetrahydrocannabinol blocks gastric hemorrhages induced by the nonsteroidal anti-inflammatory drug diclofenac sodium in mice. Eur J Pharmacol, 2013. [DOI | PubMed]
- SG Kinsey, JZ Long, ST O’Neal, RA Abdullah, JL Poklis, DL Boger, BF Cravatt, AH Lichtman. Blockade of endocannabinoid-degrading enzymes attenuates neuropathic pain. J Pharmacol Exp Ther, 2009. [DOI | PubMed]
- SG Kinsey, PS Naidu, BF Cravatt, DT Dudley, AH Lichtman. Fatty acid amide hydrolase Blockade attenuates the development of collagen-induced arthritis and related thermal hyperalgesia in mice. Pharmacol Biochem Behav, 2011. [DOI | PubMed]
- SG Kinsey, ST O’Neal, JZ Long, BF Cravatt, AH Lichtman. Inhibition of endocannabinoid catabolic enzymes elicits anxiolytic-like effects in the marble burying assay. Pharmacol Biochem Behav, 2011. [DOI | PubMed]
- E Landucci, C Mazzantini, D Lana, M Calvani, G Magni, MG Giovannini. Cannabidiol inhibits microglia activation and mitigates neuronal damage induced by kainate in an in-vitro seizure model. Neurobiol Dis, 2022. [DOI]
- JE LaVigne, R Hecksel, A Keresztes, JM Streicher. Cannabis sativa terpenes are cannabimimetic and selectively enhance cannabinoid activity. Sci Rep, 2021. [DOI | PubMed]
- X Li, C Zhang, H Tao, X Yang, X Qian, J Zhou. Dexmedetomidine alleviates osteoarthritis inflammation and pain through the CB2 pathway in rats. Int Immunopharmacol, 2023. [DOI]
- AH Lichtman, JL Poklis, A Poklis, DM Wilson, BR Martin. The pharmacological activity of inhalation exposure to marijuana smoke in mice. Drug Alcohol Depend, 2001. [DOI | PubMed]
- JZ Long, W Li, L Booker, JJ Burston, SG Kinsey, JE Schlosburg. Selective blockade of 2-arachidonoylglycerol hydrolysis produces cannabinoid behavioral effects. Nat Chem Biol, 2009. [DOI | PubMed]
- HC Lu, K Mackie. Review of the endocannabinoid system. Biol Psychiatry Cogn Neurosci Neuroimaging, 2021. [DOI | PubMed]
- S Maione, F Piscitelli, L Gatta, D Vita, L De Petrocellis, E Palazzo. Non-psychoactive cannabinoids modulate the descending pathway of antinociception in anaesthetized rats through several mechanisms of action. Br J Pharmacol, 2011. [DOI | PubMed]
- R Mechoulam. Plant cannabinoids: a neglected pharmacological treasure trove. Br J Pharmacol, 2005. [DOI | PubMed]
- A Mendiguren, E Aostri, I Rodilla, I Pujana, E Noskova, J Pineda. Cannabigerol modulates α2-adrenoceptor and 5-HT1A receptor-mediated electrophysiological effects on dorsal raphe nucleus and locus coeruleus neurons and anxiety behavior in rat. Front Pharmacol, 2023. [DOI]
- P Morales, DP Hurst, PH Reggio. Molecular targets of the phytocannabinoids-a complex picture. Prog Chem Org Nat Prod, 2017. [DOI | PubMed]
- C Muller, P Morales, PH Reggio. Cannabinoid ligands targeting TRP channels. Front Mol Neurosci, 2018. [DOI]
- PS Naidu, SA Varvel, K Ahn, BF Cravatt, BR Martin, AH Lichtman. Evaluation of fatty acid amide hydrolase inhibition in murine models of emotionality. Psychopharmacology, 2007. [DOI | PubMed]
- SR Nass, FF Steele, TB Ware, AH Libby, KL Hsu, SG Kinsey. Monoacylglycerol lipase inhibition using JZL184 attenuates paw inflammation and functional deficits in a mouse model of inflammatory arthritis. Cannabis Cannabinoid Res, 2021. [DOI | PubMed]
- National Research Council. Guide for the care and use of laboratory animals: eighth edition. Washington, DC: The National Academies Press; 2011.
- TL Nguyen, YS Nam, SY Lee, HC Kim, CG Jang. Effects of capsazepine, a transient receptor potential vanilloid type 1 antagonist, on morphine-induced antinociception, tolerance, and dependence in mice. Br J Anaesth, 2010. [DOI | PubMed]
- B Palomares, M Garrido-Rodriguez, C Gonzalo-Consuegra, M Gómez-Cañas, S Saen-oon, R Soliva, JA Collado, J Fernández-Ruiz, G Morello, MA Calzado, G Appendino, E Muñoz. ∆9-tetrahydrocannabinolic acid alleviates collagen‐induced arthritis: role of PPARγ and CB1 receptors. Br J Pharmacol, 2020. [DOI | PubMed]
- Pertwee RG, Cascio MG. Known Pharmacological actions of Delta-9-Tetrahydrocannabinoland of four other chemical constituents of cannabis that activate cannabinoid receptors. Handbook of cannabis: Oxford University Press; 2014. pp. 115–36.
- KZ Peters, JF Cheer, R Tonini. Modulating the neuromodulators: dopamine, serotonin, and the endocannabinoid system. Trends Neurosci, 2021. [DOI | PubMed]
- MK Poddar, WL Dewey. Effects of cannabinoids on catecholamine uptake and release in hypothalamic and striatal synaptosomes. J Pharmacol Exp Ther, 1980. [DOI | PubMed]
- MM Radwan, S Chandra, S Gul, MA Elsohly. Cannabinoids, phenolics, terpenes and alkaloids of cannabis. Molecules, 2021. [DOI]
- Raup-Konsavage WM, Sepulveda DE, Wang J, Dokholyan NV, Vrana KE, Graziane NM. Antinociceptive effects of Cannabichromene (CBC) in mice: insights from von Frey, Tail-Flick, Formalin, and acetone tests. Biomedicines. 2024;12(1). 10.3390/biomedicines12010083.
- Rhee M-H, Vogel Z, Barg J, Bayewitch M, Levy R, Hanuš L, Breuer A, Mechoulam R. Cannabinol Derivatives: Binding to Cannabinoid Receptors and Inhibition of Adenylylcyclase. 1997. https://pubs.acs.org/sharingguidelines
- Rodriguez CEB, Vanegas SO, Reck AM, Schrom Y, Kinsey SG. Combined endocannabinoid and cyclooxygenase inhibition additively attenuates post-surgical pain. Cannabis Cannabinoid Res. 2025;10(1):e101–e111. 10.1089/can.2024.0088
- S Rosenthaler, B Pöhn, C Kolmanz, CN Huu, C Krewenka, A Huber, B Kranner, W-D Rausch, R Moldzio. Differences in receptor binding affinity of several phytocannabinoids do not explain their effects on neural cell cultures. Neurotoxicol Teratol, 2014. [DOI | PubMed]
- WH Roughan, AI Campos, LM García-Marín, G Cuéllar-Partida, MK Lupton, IB Hickie, SE Medland, NR Wray, EM Byrne, TT Ngo, NG Martin, ME Rentería. Comorbid chronic pain and depression: shared risk factors and differential antidepressant effectiveness. Front Psychiatry, 2021. [DOI | PubMed]
- LR Ruhaak, J Felth, PC Karlsson, JJ Rafter, R Verpoorte, L Bohlin. Evaluation of the cyclooxygenase inhibiting effects of six major cannabinoids isolated from cannabis sativa. Biol Pharm Bull, 2011. [DOI | PubMed]
- JE Schlosburg, BLA Carlson, D Ramesh, RA Abdullah, JZ Long, BF Cravatt, AH Lichtman. Inhibitors of endocannabinoid-metabolizing enzymes reduce precipitated withdrawal responses in THC-dependent mice. AAPS J, 2009. [DOI | PubMed]
- JE Schlosburg, ST O, DH Conrad, AH Lichtman. CB 1 receptors mediate rimonabant-induced pruritic responses in mice: investigation of locus of action. Psychopharmacology, 2011. [DOI | PubMed]
- AM Schwarz, D Kobeci, J Mancuso, V Moreno-Rodriguez, C Seekins, T Bui, A Welborn, J Carr, JM Streicher. Select minor cannabinoids from cannabis sativa are cannabimimetic and antinociceptive in a mouse model of chronic neuropathic pain. J Pharmacol Exp Ther, 2024. [DOI]
- H Shoji, T Miyakawa. Effects of test experience, closed-arm wall color, and illumination level on behavior and plasma corticosterone response in an elevated plus maze in male C57BL/6J mice: a challenge against conventional interpretation of the test. Mol Brain, 2021. [DOI | PubMed]
- L Steru, R Chermat, B Thierry, P Simon. The tail suspension test: a new method for screening antidepressants in mice. Psychopharmacology, 1985. [DOI]
- I Svíženská, P Dubový, A Šulcová. Cannabinoid receptors 1 and 2 (CB1 and CB2), their distribution, ligands and functional involvement in nervous system structures – a short review. Pharmacol Biochem Behav, 2008. [DOI | PubMed]
- S Takeda, H Okazaki, E Ikeda, S Abe, Y Yoshioka, K Watanabe. Down-regulation of cyclooxygenase-2 (COX-2) by cannabidiolic acid in human breast cancer cells. J Toxicol Sci, 2014. [DOI | PubMed]
- Y Tang, B Wolk, DA Kendall. Effects of a CB2 subtype selective agonist ABK5-1 on cytokine production in microglia. Physiol Behav, 2018. [DOI]
- A Thomas, A Burant, N Bui, D Graham, LA Yuva-Paylor, R Paylor. Marble burying reflects a repetitive and perseverative behavior more than novelty-induced anxiety. Psychopharmacology, 2009. [DOI | PubMed]
- KR Trexler, SR Nass, MS Crowe, JD Gross, MS Jones, AW McKitrick, DP Siderovski, SG Kinsey. Novel behavioral assays of spontaneous and precipitated THC withdrawal in mice. Drug Alcohol Depend, 2018. [DOI | PubMed]
- KR Trexler, SO Vanegas, JL Poklis, SG Kinsey. The short-acting synthetic cannabinoid AB-FUBINACA induces physical dependence in mice. Drug Alcohol Depend, 2020. [DOI | PubMed]
- SO Vanegas, A Zaki, CN Dealy, SG Kinsey. The minor phytocannabinoid delta-8-tetrahydrocannabinol attenuates collagen-induced arthritic inflammation and pain-depressed behaviors. J Pharmacol Exp Ther, 2024. [DOI | PubMed]
- Vinall J, Pavlova M, Asmundson G, Rasic GJ, N., Noel M. children Mental Health Comorbidities in Pediatric Chronic Pain: A Narrative Review of Epidemiology, Models, Neurobiological Mechanisms and Treatment. 2016. 10.3390/children3040040.
- H Yang, J Zhou, C Lehmann. GPR55 – A putative “type 3” cannabinoid receptor in inflammation. J Basic Clin Physiol Pharmacol, 2016. [DOI | PubMed]
- HS Yeom, H Li, Y Tang, RP Hsung. Total syntheses of cannabicyclol, clusiacyclol A and B, iso-eriobrucinol A and B, and eriobrucinol. Org Lett, 2013. [DOI | PubMed]
- A Zagzoog, KA Mohamed, HJJ Kim, ED Kim, CS Frank, T Black. In vitro and in vivo pharmacological activity of minor cannabinoids isolated from Cannabis sativa. Sci Rep, 2020. [DOI | PubMed]
- T Zikereya, C Liu, L Wei, Y Wang, Z Zhang, C Han. The cannabinoid receptor 1 mediates exercise-induced improvements of motor skill learning and performance in parkinsonian mouse. Exp Neurol, 2025. [DOI]