
Biotechnological Fungal Platforms for the Production of Biosynthetic Cannabinoids
Abstract
Cannabinoids are bioactive meroterpenoids comprising prenylated polyketide molecules that can modulate a wide range of physiological processes. Cannabinoids have been shown to possess various medical/therapeutic effects, such as anti-convulsive, anti-anxiety, anti-psychotic, antinausea, and anti-microbial properties. The increasing interest in their beneficial effects and application as clinically useful drugs has promoted the development of heterologous biosynthetic platforms for the industrial production of these compounds. This approach can help circumvent the drawbacks associated with extraction from naturally occurring plants or chemical synthesis. In this review, we provide an overview of the fungal platforms developed by genetic engineering for the biosynthetic production of cannabinoids. Different yeast species, such as Komagataella phaffii (formerly P. pastoris) and Saccharomyces cerevisiae, have been genetically modified to include the cannabinoid biosynthetic pathway and to improve metabolic fluxes in order to increase cannabinoid titers. In addition, we engineered the filamentous fungus Penicillium chrysogenum for the first time as a host microorganism for the production of Δ9-tetrahydrocannabinolic acid from intermediates (cannabigerolic acid and olivetolic acid), thereby showing the potential of filamentous fungi as alternative platforms for cannabinoid biosynthesis upon optimization.
Article type: Review Article
Keywords: cannabinoids, fungal platforms, genetic engineering, yeasts
Affiliations: INBIOTEC (Instituto de Biotecnología de León), Av. Real 1, 24006 León, Spain; Área de Bioquímica y Biología Molecular, Departamento de Biología Molecular, Campus de Vegazana, Universidad de León, 24007 León, Spain; Departamento de Ciencias Biomédicas, Campus de Vegazana, Universidad de León, 24007 León, Spain
License: © 2023 by the authors. CC BY 4.0 Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Article links: DOI: 10.3390/jof9020234 | PubMed: 36836348 | PMC: PMC9963667
Relevance: Moderate: mentioned 3+ times in text
Full text: PDF (1.9 MB)
1. Introduction
Cannabis sativa produces about 100 (alkyl-)phytocannabinoids (meroterpenoids and their analogs derived from cannabis), generically known as cannabinoids [ref. 1,ref. 2,ref. 3]. Cannabis is supposedly of Asian origin and is one of humanity’s oldest cultivated crops [ref. 4,ref. 5,ref. 6]. It is an annual, usually dioecious and sporadically monoecious, wind-pollinated species that is extremely allogamous (cross-fertilized) in nature [ref. 2]. The taxonomy of the Cannabis genus is complex due to the existence of cultivated and wild varieties, as well as its economic importance and uses (hemp, recreational or medical). The current botanical nomenclature [ref. 7] includes just one species in the genus, C. sativa, and two subspecies (subsp. sativa (fibers and oil cultivars) and subsp. indica (“narcotic” cultivars)) based on the THC (tetrahydrocannabinol) content (hemp varieties should not contain more than 0.3% THC in dried female flowering tops). The domestication status is also used as a characteristic to distinguish the two varieties within each subspecies [ref. 4,ref. 8,ref. 9].
The fiber varieties (“hemp”: fiber extracted from the stems of C. sativa) present a high carbon-sequestering potential due to their rapid growth, and so can be used as building materials or biofuel. However, the traditional use of fiber includes the manufacture of rope, shoes, canvas, paper, clothing, and sails [ref. 4,ref. 9,ref. 10]. On the other hand, the cannabinoids present in the sticky resin produced by the female plant, including psychoactive compounds such as THC and (−)-trans-Δ9-tetrahydrocannabinol (Δ9-THC) and non-psychoactive components with potential therapeutic effects (e.g., reducing inflammation, chronic pain, and nausea) such as cannabidiol (CBD) [ref. 11,ref. 12,ref. 13], are other relevant products of C. sativa.
2. Chemical Structure of Cannabinoids, Types, and Metabolic Routes for Biosynthesis
Phytocannabinoids are bioactive natural meroterpenoids with a resorcinol core bearing a para-positioned isoprenyl, alkyl, or aralkyl side chain [ref. 14]. The side chain usually contains an odd number of carbon atoms (cannabinoids containing an even number of carbon atoms are rare). Phytocannabinoids are found in angiosperms, fungi, and liverworts and are produced in several plant organs, such as the flower and glandular trichome of C. sativa, the scales of Rhododendron, and the oil bodies of Radula species. Moreover, it has been found that the mammalian brain has receptors that respond to compounds found in C. sativa. These cannabinoid receptors form the endocannabinoid system and regulate several biological functions and metabolic processes. Therefore, these bioactive compounds can be beneficial for the treatment of pain, anxiety, and cachexia in humans [ref. 15,ref. 16].
The different cannabinoids produced by C. sativa [ref. 17] are classified into structural families [ref. 18,ref. 19], such as cannabidiols (CBDs), cannabigerols (CBGs), cannabicyclols (CBLs), cannabinodiols (CBNDs), cannabinols (CBNs), cannabitriols (CBTs), cannabichromenes (CBCs), Δ9-THC, and miscellaneous cannabinoids [ref. 20]. The acid forms are the final product of the cannabinoid biosynthetic pathways, although several spontaneous modifications such as oxidation, decarboxylation, and cyclization frequently take place because of the poor oxidative stability of phytocannabinoids, in particular Δ9-THC [ref. 14].
Phytocannabinoids are accumulated in glandular thricomes all over the aerial parts of the plant, especially in the female flowers [ref. 21,ref. 22]. The phytocannabinoid biosynthetic pathway is split between different cellular compartments and organelles: the cytosol of gland cells (polyketide pathway), the plastids (methylerythritol 4-phosphate (MEP) pathway for prenylation), and the extracellular storage cavity (oxidocyclization and storage) (Figure 1). It is still unclear how intermediates and precursors are transported between compartments, although it is most likely that vesicle trafficking and transport proteins are involved in the movements of these intermediates across the interface between the gland cells and the storage cavity.
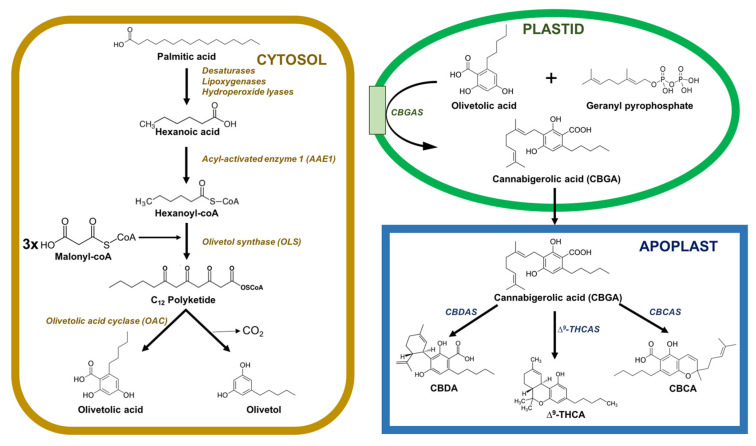
Inside the cytosol, the biosynthesis of cannabinoids involves the integration of several steps in polyketide and isoprenoid metabolism. C18 fatty acids are desaturated, preoxygenated, and cleaved into hexanoic acid, which is transformed into thioester hexanoyl-CoA in a reaction catalyzed by acyl-activated enzyme 1 (AAE1). Later, the hexanoyl-CoA is elongated with malonyl-CoA in a reaction catalyzed by olivetol synthase (OLS) and cyclized by olivetolic acid cyclase (OAC) to produce olivetolic acid (OA) [ref. 23,ref. 24,ref. 25].
Inside plastids, geranyl pyrophosphate (GPP) is synthesized through the MEP pathway. OA is prenylated using GPP by cannabigerolic acid synthase (CBGAS), thereby forming cannabigerolic acid (CBGA), the first cannabinoid. CBGA is an essential cannabinoid because it is the precursor of several other cannabinoids. Then, the CBGA is converted into Δ9-tetrahydrocannabinolic acid (Δ9-THCA) and cannabidiolic acid (CBDA) in the apoplastic space by the action of two enzymes, Δ9-THCA synthase (THCAS) and CBDA synthase (CBDAS), respectively. This conversion causes the reduction of O2 into hydrogen peroxide (H2O2) via oxidative cyclization reactions. Another important enzyme, cannabichromenic acid synthase (CBCAS), participates in the synthesis of cannabichromenic acid (CBCA) from CBGA using FAD and O2. THCAS, CBGAS, and CBDAS are also flavoproteins, which are strictly dependent on the presence of O2 as an electron acceptor. All these oxidocyclases carry a secretion signal peptide and are exported to the extracellular resin space. THCAS and CBDAS are active in the resin space, but it is still unknown whether their activity is exclusive to the extracellular space [ref. 23,ref. 26,ref. 27].
Δ9-THCA, CBDA, and CBDA with a pentyl side chain are the end-products of the enzymatic biosynthesis of cannabinoids and are synthesized in the apoplastic space. These active compounds undergo spontaneous rearrangement reactions when exposed to heat or radiation or during storage [ref. 23,ref. 26,ref. 28].
3. Importance, Applications, and Impact of Cannabinoids
Phytocannabinoids play several roles in human health, and cannabis preparations have been used in medicines since ancient times [ref. 29]. On the one hand, they exhibit anti-microbial activity against some bacteria and fungi, and they are effective against a wide range of infectious diseases in humans, acting as potent antibiotics. Cannabis extracts possess antimicrobial activity against some Gram-positive bacteria, such as Bacillus subtilis and Staphylococcus aureus, and the Gram-negative bacteria Escherichia coli and Pseudomonas aeruginosa. In contrast, no activity is displayed against Candida albicans and Aspergillus niger [ref. 30]. Phytocannabinoids (e.g., Δ9-THC, CBG, CBN, CBD, and CBC) display antibiotic activity against methicillin-resistant Staphylococcus aureus [ref. 31]. Δ9-THC and CBD exhibit bactericidal activity against streptococci and staphylococci but not against Gram-negative bacteria [ref. 32]. Because of their pharmacological potential, an increasing number of countries are relaxing their legislation around phytocannabinoids, and the global industry around cannabis-derived products is growing exponentially [ref. 33].
On the other hand, phytocannabinoids exert strong therapeutic potential in humans owing to their interaction with the G-protein-coupled cannabinoid receptors (GPCRs), such as CB1 and CB2; transient receptor potential (TRP) ion channels; and peroxisome proliferator-activated receptor (PPAR). CB1 is the most abundant GPCR in the central nervous system, and CB2 is located predominantly in the cells and tissues of the immune system [ref. 34]. Thus, cannabinoids play a key role in signaling, and the proper functioning of the immune and central nervous systems. Δ9-THC is the major psychoactive component of cannabis and displays pleiotropic effects in humans, including analgesic response, relaxation, pain tolerance, and dysphoria (anxiety disorder). Δ9-THC is also used by patients with insomnia and depression, since it improves sleep [ref. 35]. CBD exerts its function in humans through the CB1 and CB2 receptors in the CNS and the peripherical regions [ref. 36]. It is administered to patients with treatment-resistant epilepsy. CBD can also be delivered to patients receiving pharmacotherapy and can act as a potential cannabinoid to cure obesity, convulsive disorder, and rheumatoid arthritis. Furthermore, CBD also exhibits anti-psychotic, anti-nausea, and anti-anxiety properties [ref. 37].
In addition to the legal cannabis market (including medical and recreational uses), the EMCDDA (European Monitoring Centre for Drugs and Drug Addiction) estimated the illicit cannabis market in 2017 to be around 1.4–1.7 tones, valued at EUR 10.5–12.8 billion [ref. 38]. Several clinical uses of cannabinoids have been described, but just a few are legally recognized by international regulatory agencies. The possible clinical applications of CBD have been summarized in sixteen diseases, which included several cancer types through anti-proliferative and anti-invasive action; inflammatory bowel and Chron’s diseases; cardiovascular diseases through anti-oxidant and anti-inflammatory properties; and neurodegenerative diseases (e.g., Parkinson’s and Alzheimer’s) [ref. 13]. However, in January 2020 the US Food and Drug Administration (FDA), FDA and Cannabis: Research and Drug Approval Process: https://www.fda.gov/news-events/public-health-focus/fda-and-cannabis-research-and-drug-approval-process (accessed on 1 February 2023) indicated that up to that date just one cannabis-derived drug product had been approved, Epidiolex, containing a purified form of CBD involved in the treatment of seizures associated with Lennox–Gastaut syndrome or Dravet syndrome in patients 2 years of age and older. In addition, three synthetic cannabis-related drug products have been validated by the FDA: (i) Marinol (dronabinol); (ii) Syndros (dronabinol); and (iii) Cesamet (nabilone). On the one hand, Marinol and Syndros represent a synthetic version of THC, the psychoactive component of cannabis, for the treatment of nausea due to cancer chemotherapy and anorexia, thus preventing weight loss in AIDS patients. On the other hand, Cesamet has a chemical synthetic structure similar to THC as an active ingredient, which is used to treat nausea associated with cancer chemotherapy.
Thus, in summary, cannabis as a hemp fiber is an economical reality, but the most promising market comprises the medical and recreative uses of the plant. However, the botanical derivatives are scarce in the legal market due to the putative risk of co-purification with other cannabinoids during the extraction process from naturally occurring plants. Nowadays, synthetic products are on the market through companies such as AbbVie Inc.; Corbus Pharmaceuticals Holdings Inc.; INSYS Therapeutics Inc.; and Bausch Health, although chemical synthesis is a costly process involving the use of chemicals that are not environmentally friendly. Nevertheless, synthetic production has paved the way for the biotechnological production of single cannabinoid compounds or derivatives in microorganisms, mainly fungi, which have been endowed with the biochemical machinery to reproduce the plant pathways and products [ref. 39], an approach that has been promoted by the development of recent synthetic biology methodologies [ref. 40].
4. Development of Biotechnological Fungal Platforms for the Biosynthesis of Cannabinoids
Fungi produce a broad diversity of primary and secondary metabolites with different bioactivities. Different biotechnological processes have been implemented with the yeasts Saccharomyces cerevisiae [ref. 41,ref. 42,ref. 43], Yarrowia lipolitica [ref. 44,ref. 45], and Komagataella phaffii (formerly Pichia pastoris [ref. 46,ref. 47]) [ref. 48,ref. 49]. The production of high-value metabolites through different biotechnological processes is also achieved by means of filamentous fungi, such as Aspergilli and Penicillium species, Acremonium chrysogenum, Blakeslea trispora, and Trichoderma reesei [ref. 50,ref. 51,ref. 52]. Among these filamentous fungi, P. chrysogenum (P. rubens) stands out for its adaptability to the massive industrial overproduction of penicillin [ref. 53,ref. 54,ref. 55], which suggests the use of this microorganism as a cell factory for products other than β-lactam antibiotics [ref. 56,ref. 57,ref. 58]. The complete genome sequences of yeasts and filamentous fungi obtained in recent years have provided the basic information for the development of new approaches aimed at improving production strains, increasing metabolite yields, and designing new fungal production platforms for natural products with potential commercial value [ref. 51,ref. 59,ref. 60].
Although bacteria such as Escherichia coli, Bacillus subtilis, Streptomyces lividans, and Corynebacterium glutamicum are easier to manipulate genetically, these microorganisms are not always suitable for the heterologous expression of proteins of eukaryotic origin due to their lack of post-translational modifications and their limited ability to secrete heterologous proteins. On the contrary, some yeasts and filamentous fungi have the ability to biosynthesize biologically active proteins with a degree of glycosylation similar to the original proteins [ref. 52] and possess a great secreting capacity for different enzymes and metabolites. It should be noted that 82% of the commercial enzymes used in the food industry are manufactured using fungal hosts [ref. 61], which confirms the role of these microorganisms as cell factories for heterologous protein production [ref. 62].
The application of new genetic engineering techniques at the molecular level has allowed a considerable increase in the production of recombinant proteins and secondary metabolites in yeast and filamentous fungi [ref. 49,ref. 63,ref. 64,ref. 65,ref. 66,ref. 67]. This has contributed to the design of fungal hosts for the heterologous expression of biosynthetic gene clusters that could work as efficient platforms to produce high-added-value secondary metabolites, such as cannabinoids.
4.1. Biosynthesis of Cannabinoids in Yeasts
Cannabinoid biosynthesis by means of biotechnological approaches and independently of the plant has been boosted by: (i) its interest as pharmaceutical and bioactive compounds; (ii) the limiting production of synthetic THC and; (iii) the legal regulatory issues for the cultivation of C. sativa in most countries. Thus, numerous papers and patents describing biotechnological procedures for the microbial production of cannabinoids (mainly in yeasts) have been reported in the recent years, as it is summarized below.
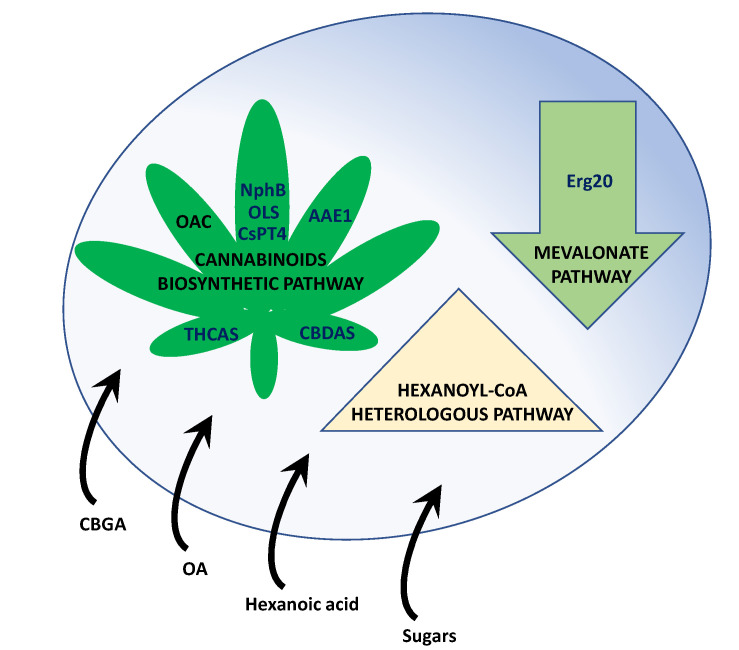
The identification of the cannabinoids biosynthetic genes and enzymes [ref. 24,ref. 68,ref. 69,ref. 70,ref. 71,ref. 72] paved the way to reconstruct the biosynthetic pathway and engineer the metabolism in suitable heterologous systems [ref. 39], mainly yeasts (Figure 2).
Early reports focused on the recombinant expression of THCAS in different hosts, such as tobacco hairy roots, insect cell cultures, and K. phaffii cultures [ref. 71,ref. 73]. The expression of THCAS in these hosts was very low, although it opened the door to the possibility of biotechnological Δ9-THCA production by means of biotechnological platforms. E. coli was initially used as a potential host microorganism, but it was quickly discarded, since no functional expression of THCAS was achieved in this microorganism [ref. 74,ref. 75], thereby suggesting that a eukaryotic expression system was necessary for appropriate enzyme folding and maturation. Initial drawbacks of the THCAS scale-up processes regarding the water solubility of CBGA and the membrane localization of CBGAS led scientists to also focus on the intracellular expression of THCAS in S. cerevisiae and K. phaffii cells and the possible application of a whole-cell production system for Δ9-THCA [ref. 75]. These authors used the signal peptide of the vacuolar proteinase A to target THCAS in the cell vacuole and found that the highest enzyme activity was obtained in cultures from proteinase A knockout strains of K. phaffii. The further optimization of THCAS expression in K. phaffii dramatically increased enzyme activity in relation to previous studies, and the bioconversion of CBGA in whole yeast cells led to the production of 1 mM Δ9-THCA [ref. 75].
The CBGAS expression problems related to its membrane localization led scientists to use the soluble aromatic prenyltransferase from Streptomyces sp. CL190 strain (StNphB) instead of plant prenyltransferase. The partial reconstruction of the Δ9-THCA biosynthetic pathway in K. phaffii gave rise to the successful synthesis of this compound from OA and GPP. Although the side-product 2-O-geranyl OA was also produced because of StNPhB activity, these results suggested that NphB was a promising candidate for the substitution of plant-derived membrane-bound prenyltransferases in the heterologous system [ref. 76].
Although K. phaffii was found to be a good host for the heterologous expression of cannabinoid biosynthetic enzymes, several bottlenecks attributed to THCAS protein folding were also observed. They were counteracted by the co-production of different proteins, including foldases and chaperones. Thus, the highest THCAS activity was obtained with the co-production of the spliced gene version of: (i) basic leucine zipper transcription factor Hac1s (involved in the regulation of several genes involved in protein folding, secretion, ER quality control, glycosylation, and ER-associated degradation); (ii) CNE1 (an ER membrane protein that functions as a constituent of the ER quality control apparatus); and (iii) FAD1 (FAD synthetase, involved in the production of FAD from FMN). This strain showed improved productivity in comparison to previous strains and reached 3.05 g/L THCA from the bioconversion of CBGA within 8 h of incubation [ref. 77]. The same authors also carried out site-directed mutagenesis in THCAS and CBDAS in order to elucidate the structure–function relationship and improve the enzyme properties for biotechnological cannabinoid production. After the recombinant expression of different mutant enzymes in K. phaffi, the enzyme bearing the double substitution N89Q + N499Q (lacking two glycosylation sites) gave rise to a 2.0-fold increase in THCAS activity, whereas the variant A414V (mutant in the active site) was able to increase the catalytic activity for the production of CBDA by 3.3 times [ref. 78].
In parallel, S. cerevisiae was engineered to generate strains capable of performing the complete biosynthesis of CBGA, Δ9-THCA, CBDA, Δ9-tetrahydrocannabivarinic acid, and cannabidivarinic acid from the simple sugar galactose [ref. 23]. In this biosynthetic platform, hexanoyl-CoA was produced either by feeding hexanoic acid as a substrate for the acyl-activating enzyme (encoded by CsAAE1 from C. sativa) or from acetyl-CoA (derived from sugars) due to the introduction of a multiorganism heterologous biosynthetic pathway comprising RebktB from Ralstonia eutropha, CnpaaH1 from Cupriavidus necator, Cacrt from Clostridium acetobutylicum, and Tdter from Treponema denticola. In addition to this, the native mevalonate pathway was modified, including a mutant version of Erg20 (F69W/N127W) to overproduce GPP. The introduction of the OA biosynthetic genes CsTKS and CsOAC, a gene encoding a previously undiscovered enzyme with geranylpyrophosphate:olivetolate geranyltransferase activity (CsPT4) involved in CBGA biosynthesis, together with THCAS and CBDAS (with a vacuolar localization tag), allowed S. cerevisiae to produce 1.1 mg/L Δ9-THCA or 4.3 μg/L CBDA from hexanoic acid, or 2.3 mg/L Δ9-THCA or 4.2 μg/L CBDA from galactose. These strains also produced Δ9-tetrahydrocannabivarinic acid and cannabidivarinic acid. The titers of Δ9-THCA and Δ9-tetrahydrocannabivarinic acid were further increased up to 8.0 mg/L (Δ9-THCA) and 4.8 mg/L (Δ9-tetrahydrocannabivarinic acid) after the overexpression of additional single copies of the biosynthetic genes CsTKS, CsOAC, and THCAS. Moreover, unnatural cannabinoid analogs with modifications in the part of the molecule that is known to alter receptor binding affinity and potency were obtained by feeding different fatty acids (e.g., pentanoic acid, heptanoic acid, and 5-hexenoic acid) to the S. cerevisiae engineered strains [ref. 23].
The reconstruction of the complete Δ9-THCA and CBDA biosynthetic pathway in baker’s yeast represented a milestone in the field of the heterologous biosynthesis of cannabinoids, but the titers were considered low from an industrial point of view. Therefore, bioengineering studies and the modeling of the heterologous biosynthesis of Δ9-THCA were carried out in this yeast chassis. Several critical aspects that require systematic optimization were detected after the analysis of metabolic bottlenecks, such as: (i) insufficient hexanoic acid formation in the fatty acid biosynthesis; (ii) low acetyl-CoA precursor delivery to the hexanoic acid biosynthesis and the mevalonate pathway; (iii) the limited catalytic activity of NphB or CsPT and THCAS; (iv) insufficient ATP and NADPH regeneration and; (v) ethanol production by Crabtree effect [ref. 79]. In another study, S. cerevisiae was engineered by introducing plant uridine diphosphate glycosyltransferases into its genome in order to produce OA glucoside and cannabigerolic acid glucoside. This could be of interest due to the high hydrophobicity of naturally occurring cannabinoids, which affects their use as pharmaceuticals [ref. 80].
Due to the importance of yeast peroxisomes in different metabolic pathways, such as the β-oxidation of fatty acids leading to the creation of a pool of acetyl-CoA that can be used in the formation of derivatives [ref. 81] (e.g., isoprenoid via the mevalonate pathway), these organelles were engineered for the more efficient production of CBGA. Thus, GPP synthase Erg20p (N127W) and geranyldiphosphate:olivetolate geranyltransferase CsPT4 were targeted to the yeast peroxisome together with the mevalonate pathway proteins. Under galactose-induced conditions and after supplementation with OA, the production of CBGA reached 0.82 mg/L, thereby confirming that yeast organelles can act as synthetic biology devices [ref. 82].
In addition to the scientific papers indicated above, which represent some of the research carried out in this field, several patents have been published dealing with the biosynthesis of cannabinoids in yeast. Some of these patents are described below. In a patent application filed in 2016 [ref. 83], transgenic yeasts (S. cerevisiae and Kluyvermyces marxianus) capable of reconstituting the CBDA metabolic pathway of C. sativa were generated. This biosynthetic process was claimed to be sustainable and more environmentally friendly than synthetic production. To circumvent the low production of the precursor GPP in the yeasts, an improved mutant prenyltransferase Erg20 (K179E) that shifted the GPP:FPP ratio towards the first compound (70:30) was used in both yeasts. In S. cerevisiae, CBDA was produced upon the addition of hexanoic acid. In contrast, K. marxianus was engineered to produce hexanoyl-CoA from glucose, since the genes encoding the enzymes required for this route were introduced in this microorganism in addition to those involved in CBDA biosynthesis. In another patent, S. cerevisiae was engineered to incorporate the cannabinoid biosynthetic pathway, including mutant variants of Erg20. Depending on the feedstock used (OA or hexanoic acid), the Δ9-THCA titers were 84 mg/L and 23 mg/L, respectively. An improvement in CBGA production was achieved due to the greater availability of hexanoic acid when the endogenous FOX1 gene, which controls the β-oxidation of long-chain fatty acids, was disrupted in this yeast strain [ref. 84]. In order to produce water-soluble cannabinoids, thereby improving traditional cannabinoid production, other inventors generated a yeast expression system based on the methylotrophic yeast K. phaffii, which was engineered to express glycosyltransfereases from different organisms. The intracellular expression of NtGT4 (UGT 73-like glycosyltransferase from Nicotiana tabacum) led to the highest level of glycosylation of CBDA after the addition of 27 μM CBDA [ref. 85]. In another patent application, S. cerevisiae was genetically modified for the production of phytocannabinoids and phytocannabinoid analogs using a type I fatty acid synthase/PKS from the amoeba Dictyostelium discoideum (DiPKS). In this enzyme, which synthesizes 1-methylolivetol directly from malonyl-CoA, thereby representing an advantage regarding hexanoyl-CoA supply, point mutations were included to diminish the methylation of olivetol (G1516R) or to allow the biosynthesis of both methyl-olivetol and olivetol (G1516D/G1518A). Due to genetic engineering, the yeast strains produced 66.3 mg/L methyl-CBG, 74.26 mg/L olivetol, and 42.44 mg/L methyl-olivetol [ref. 86]. In another patent, the mevalonate pathway was overexpressed in S. cerevisiae. In addition, an ERG20 F96W/N127W mutant was added to provide a GPP precursor source in the cell, and pyruvate decarboxylase (PDC) from Zymomonas mobilis was chromosomally integrated to increase the flux from pyruvate towards acetyl-CoA. The inclusion of CsPT4 led to the production of 215.6 mg/L CBGA after feeding 1 mM OA [ref. 87]. Synthetic biology approaches in S. cerevisiae have also been reported in other patents [ref. 88,ref. 89], and a cell-free system for the production of cannabinoids has been patented [ref. 90].
Candida viswanathii was also tested as a host microorganism for the production of cannabinoids from fatty acids. The strains generated in this patent contained an engineered β-oxidation pathway and an engineered pathway for the production of CBGA, yielding 0.67 mg/L (supernatants) or 1.51 mg/L (lysates) CBGA and 5.86 mg/L (supernatants) or 12.23 mg/L (lysates) OA from oleic acid [ref. 91]. The oleaginous yeast Y. lipolytica has also been genetically engineered to produce the cannabinoid precursor OA. The co-expression of different enzymes, such as acetyl-CoA carboxylase, pyruvate dehydrogenase bypass, and the malic enzyme, as well as the activation of the peroxisomal β-oxidation pathway and the ATP export pathway, allowed the production of 9.18 mg/L OA in a shake-flask culture [ref. 92], which makes this yeast a promising microbial host cell for the biosynthesis of cannabinoids.
4.2. Biosynthesis of Cannabinoids in Filamentous Fungi: The Paradigm of P. chrysogenum
To date, there have been no reports on the biosynthetic production of cannabinoids in filamentous fungi. To the best of our knowledge, there is only one report describing the biosynthesis of OA in Aspergillus nidulans using fungal tandem polyketide synthases without the requirement of CsOLS and CsOAC [ref. 93], which suggested that new synthetic biology strategies can be implemented for the biosynthesis of microbial cannabinoids in filamentous fungi.
The successful trajectory of P. chrysogenum as industrial producer of penicillin, and the large titers achieved by this microorganism under industrial conditions [ref. 94], together with the availability of different molecular and omics tools for the genetic engineering of this filamentous fungus [ref. 54,ref. 55,ref. 57], make it very attractive as cell factory for secondary metabolite production. Based on this, we decided to use P. chrysogenum as a platform for cannabinoid biosynthesis and production. Therefore, we aimed to test the feasibility of filamentous fungi as cannabinoid producers by developing a P. chrysogenum-based biotechnological platform that allows the production of cannabinoids in submerged cultures. In order to divert the high metabolic flux resulting from penicillin biosynthesis through the heterologous cannabinoid biosynthetic pathway, a strain lacking a region that includes the penicillin biosynthetic gene cluster (pcbAB, pcbC, and penDE genes) and several ORFs flanking this cluster, was chosen as the parental strain. In addition, to facilitate selection upon transformation, a uridine auxotroph of this strain was used (P. chrysogenum Wis54-1255 npe10 pyrG−(Δpen)) [ref. 95,ref. 96]. Due to the availability of commercial CBGA and OA, we proposed a synthetic biology strategy consisting of the gradual introduction of heterologous genes required for Δ9-THCA biosynthesis, starting with the late biosynthetic gene THCAS (Figure 3a).
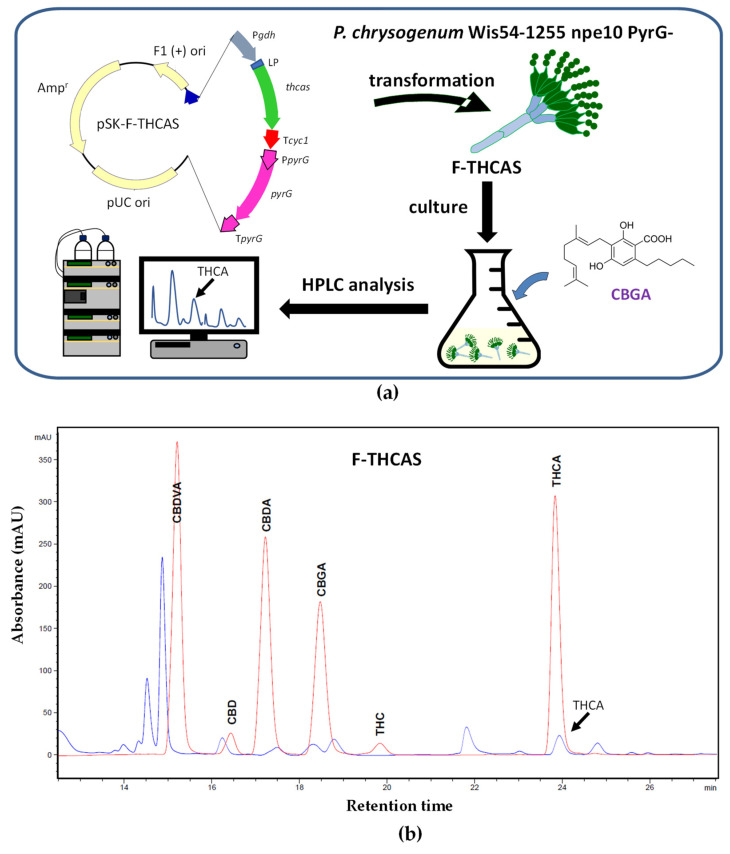
The sequence of the gene encoding the THCAS of C. sativa was optimized from the point of view of fungal codon usage. Previous studies executed in silico with the SignalP-5.0 program [ref. 97] indicated that the first 28 amino acids of this enzyme constitute a signal peptide. This signal peptide was replaced with the 19 amino acid signal peptide of the P. chrysogenum Pc21g02370 (with a strong similarity to aspergillopepsin apnS from Aspergillus phoenicis) [ref. 98]. Therefore, the cDNA of THCAS was designed to include 57 nucleotides encoding the above-mentioned signal peptide and was subcloned into the pBluescript II SK plasmid under the control of the A. awamori gdhA (glutamate dehydrogenase) gene promoter and the transcriptional terminator of the S. cerevisiae cyc1 gene. This construct also included the PyrG expression cassette to allow selection upon transformation and was named pSK-F-THCAS (Figure 3a). The transformants were tested by PCR to confirm the integration of the THCAS expression cassette (not shown), and genetically modified P. chrysogenum transformants were named F-THCAS. These transformants were grown in PMMY medium [ref. 99] for 48 h at 26 °C and 250 rpm and supplemented with 0.1 mM CBGA. After 48 h of additional growth, mycelia were collected, broken down in liquid nitrogen to a fine powder, and resuspended in methanol. HPLC analysis was carried out following the protocol previously described in [ref. 100] with some modifications. Chromatographic separations were achieved using a Core-Shell Kinetex® EVO C18 (Phenomenex, Torrance, CA, USA) analytical column (3 μm, 150 mm × 4.6 mm i.d.). The mobile phase consisted of a gradient of (methanol: acetonitrile 75:25)/water, containing 0.1% formic acid. The flow rate was set to 0.5 mL/min, and the injection volume was 20 μL. All experiments were carried out at 50 °C. Full spectra were recorded in the range of 200–400 nm. As a result of the bioconversion of CBGA, Δ9-THCA was detected intracellularly (Figure 3b). The authenticity of Δ9-THCA was confirmed by mass spectrometry using a Triple Quad 6500 System (AB Sciex LLC, Framingham, MA, USA).
In a second step, we evaluated the ability of the F-THCAS strain to carry out the prenylation of OA and biosynthesize Δ9-THCA (Figure 4a). To achieve this goal, we codon-optimized and overexpressed in these strains a mutant variant (Y288A/G286S) of the soluble aromatic prenyltransferase from Streptomyces sp. CL190 strain (StNphB), which has been reported to show improved enzyme kinetics [ref. 101], together with a codon-optimized mutant version of Erg20 (F69W/N127W), the latter leading to GPP overproduction [ref. 23].
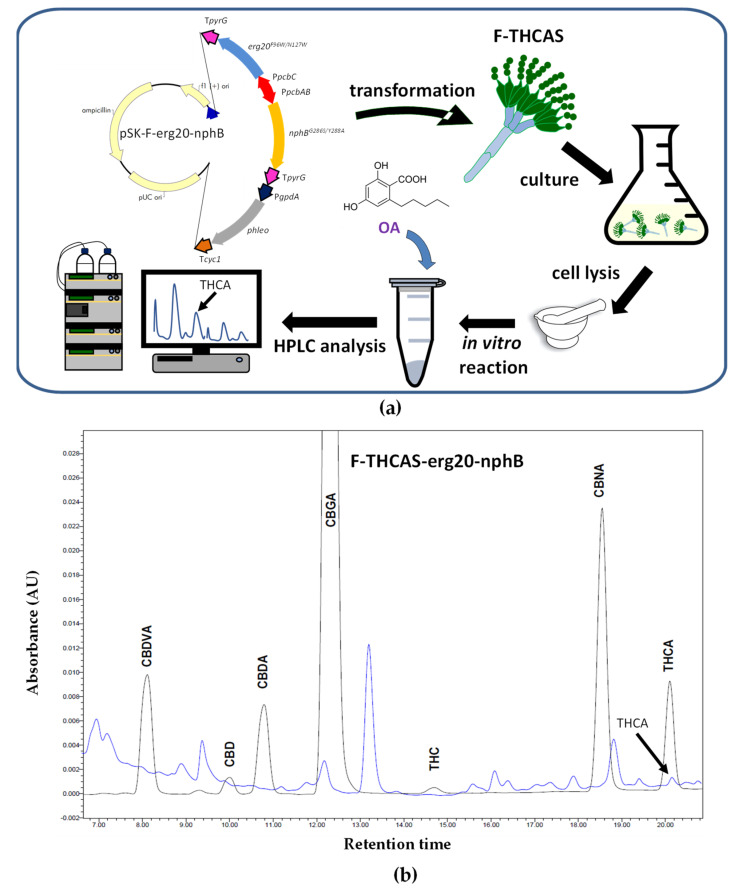
The cDNAs encoding these two proteins were included in the pBluescript II SK plasmid under the control of the bidirectional pcbAB-pcbC gene promoter (controlling the expression of the pcbAB and pcbC genes of the penicillin biosynthetic gene cluster and absent in the P. chrysogenum Wis54-1255 npe10 pyrG−strain) and the transcriptional terminator of the pyrG gene. For the selection of transformants, this construct also included the phleomycin resistance cassette, thus giving rise to pSK-F-erg20-nphB (Figure 4a).
The transformants were tested by PCR to confirm the integration of the DNA fragment including the StNphBY288A/G286S and Erg20F69W/N127W genes and the bidirectional pcbAB-pcbC promoter (not shown). The genetically modified P. chrysogenum transformants obtained in this step were named F-THCAS-erg20-nphB and were grown in PMMY medium. After 80 h of growth at 26 °C and 250 rpm, the mycelia were collected and broken down in liquid nitrogen to a fine powder to assess the formation of Δ9-THCA in the lysates in the presence of 1 mM OA and 0.1 mM GPP in a reaction carried out as previously described [ref. 76]. As a result of the bioconversion of OA into CBGA and the latter into Δ9-THCA, this cannabinoid was detected by HPLC analysis (Figure 4b) following a protocol similar to that indicated above with a different gradient. The authenticity of Δ9-THCA was confirmed by mass spectrometry using a Triple Quad 6500 System (SCIEX).
These preliminary results confirm that P. chrysogenum can biosynthesize cannabinoids using the codon-optimized plant biosynthetic genes, thus demonstrating the feasibility of filamentous fungi as cannabinoid producers. Due to the strong background of P. chrysogenum in the pharmaceutical industry as penicillin producer, the genetic engineering of this Ascomycete with the rest of the cannabinoid biosynthetic genes could give rise to an interesting platform for the industrial production of these compounds, although several optimization steps are still required to make it attractive for this purpose.
5. Conclusions
The biosynthetic production of cannabinoids in fungal hosts presents great advantages in terms of the cost and productivity of therapeutic cannabinoids. This strategy could yield greater amounts of pure products with increased stability and robustness and at a lower cost in terms of the activities and resources required compared to those necessary for the traditional process of extraction from the plant or chemical synthesis. Yeasts have emerged as suitable platforms for the industrial production of cannabinoids, and the optimization of the production process can lead to increased titers. Based upon the experience of previous industrial processes for the production of secondary metabolites, filamentous fungi could also represent an alternative host for the cannabinoid industry. Since P. chrysogenum has demonstrated its robustness under extreme industrial conditions in the production of penicillin, it could constitute a good candidate for the biosynthesis of other secondary metabolites. Herein, we provided evidence that this filamentous fungus could be genetically modified to produce small amounts Δ9-THCA after feeding cultures with CBGA or after the addition of OA to cell lysates, thereby confirming the feasibility of using this type of fungal host for this purpose. Although there is still much to be achieved in this field, including the optimization of expression cassettes, biosynthetic enzymes, and culture conditions and the redirection of metabolic fluxes, we believe that P. chrysogenum could become a suitable biotechnological fungal platform for the production of cannabinoids in the near future.
References
- Z. Atakan. Cannabis, a complex plant: Different compounds and different effects on individuals. Ther. Adv. Psychopharmacol., 2012. [DOI | PubMed]
- B.F. Thomas, M.A. ElSohly. The Botany of Cannabis sativa L.. The Analytical Chemistry of Cannabis, 2016. [DOI]
- J.A. Hartsel, J. Eades, B. Hickory, A. Makriyannis. Cannabis sativa and Hemp. Nutraceuticals: Efficacy, Safety and Toxicity, 2016. [DOI]
- B. Warf. High points: An historical geography of cannabis. Geogr. Rev., 2014. [DOI]
- H.-L. Li. An archaeological and historical account of cannabis in China. Econ. Bot., 1974. [DOI]
- G. Ren, X. Zhang, Y. Li, K. Ridout, M.L. Serrano-Serrano, Y. Yang, A. Liu, G. Ravikanth, M.A. Nawaz, A.S. Mumtaz. Large-scale whole-genome resequencing unravels the domestication history of Cannabis sativa. Sci. Adv., 2021. [DOI | PubMed]
- E. Small, A. Cronquist. A practical and natural taxonomy for cannabis. Taxon, 1976. [DOI]
- J.M. McPartland. Cannabis systematics at the levels of Family, Genus, and Species. Cannabis Cannabinoid Res., 2018. [DOI | PubMed]
- A. Pollio. The name of Cannabis: A short guide for nonbotanists. Cannabis Cannabinoid Res., 2016. [DOI | PubMed]
- S. Schilling, C.A. Dowling, J. Shi, L. Ryan, D.J.L. Hunt, E. O’Reilly, A.S. Perry, O. Kinnane, P.F. McCabe, R. Melzer. The cream of the crop: Biology, breeding, and applications of Cannabis sativa. Annu. Plant Rev. Online, 2021. [DOI]
- A. Iftikhar, U. Zafar, W. Ahmed, M.A. Shabbir, A. Sameen, A. Sahar, Z.F. Bhat, P.Ł. Kowalczewski, M. Jarzębski, R.M. Aadil. Applications of Cannabis sativa L. in Food and Its Therapeutic Potential: From a Prohibited Drug to a Nutritional Supplement. Molecules, 2021. [DOI | PubMed]
- S.C. Britch, S. Babalonis, S.L. Walsh. Cannabidiol: Pharmacology and therapeutic targets. Psychopharmacology, 2021. [DOI | PubMed]
- S. Pisanti, A.M. Malfitano, E. Ciaglia, A. Lamberti, R. Ranieri, G. Cuomo, M. Abate, G. Faggiana, M.C. Proto, D. Fiore. Cannabidiol: State of the art and new challenges for therapeutic applications. Pharmacol. Ther., 2017. [DOI | PubMed]
- L.O. Hanuš, S.M. Meyer, E. Muñoz, O. Taglialatela-Scafati, G. Appendino. Phytocannabinoids: A Unified Critical Inventory. Nat. Prod. Rep., 2016. [DOI | PubMed]
- D. Chanda, D. Neumann, J.F.C. Glatz. The endocannabinoid system: Overview of an emerging multi-faceted therapeutic target. Prostaglandins. Leukot. Essent. Fatty Acids, 2019. [DOI | PubMed]
- M. Dei Cas, E. Casagni, A. Casiraghi, P. Minghetti, D.M.M. Fornasari, F. Ferri, S. Arnoldi, V. Gambaro, G. Roda. Phytocannabinoids profile in medicinal cannabis oils: The impact of plant varieties and preparation methods. Front. Pharmacol., 2020. [DOI | PubMed]
- F. Taura, S. Sirikantaramas, Y. Shoyama, Y. Shoyama, S. Morimoto. Phytocannabinoids in Cannabis sativa: Recent studies on biosynthetic enzymes. Chem. Biodivers., 2007. [DOI | PubMed]
- B. Zirpel. Recombinant Expression and Functional Characterization of Cannabinoid Producing Enzymes in Komagataella phaffii. Ph.D. Thesis, 2018
- A.D. Kinghorn, H. Falk, S. Gibbons, J. Kobayashi. Phytocannabinoids. Unraveling the Complex Chemistry and Pharmacology of Cannabis sativa, 2017
- M.A. Elsohly, D. Slade. Chemical constituents of marijuana: The complex mixture of natural cannabinoids. Life Sci., 2005. [DOI | PubMed]
- I.J. Flores-Sanchez, R. Verpoorte. Secondary metabolism in cannabis. Phytochem. Rev., 2008. [DOI]
- N. Happyana, S. Agnolet, R. Muntendam, A. Van Dam, B. Schneider, O. Kayser. Analysis of cannabinoids in laser-microdissected trichomes of medicinal Cannabis sativa using LCMS and cryogenic NMR. Phytochemistry, 2013. [DOI | PubMed]
- X. Luo, M.A. Reiter, L. D’Espaux, J. Wong, C.M. Denby, A. Lechner, Y. Zhang, A.T. Grzybowski, S. Harth, W. Lin. Complete biosynthesis of cannabinoids and their unnatural analogues in yeast. Nature, 2019. [DOI | PubMed]
- S.J. Gagne, J.M. Stout, E. Liu, Z. Boubakir, S.M. Clark, J.E. Page. Identification of olivetolic acid cyclase from Cannabis sativa reveals a unique catalytic route to plant polyketides. Proc. Natl. Acad. Sci. USA, 2012. [DOI | PubMed]
- T.J. Raharjo, W.-T. Chang, Y.H. Choi, A.M. Peltenburg-Looman, R. Verpoorte. Olivetol as product of a polyketide synthase in Cannabis sativa L.. Plant Sci., 2004. [DOI]
- T. Gülck, B.L. Møller. Phytocannabinoids: Origins and biosynthesis. Trends Plant Sci., 2020. [DOI | PubMed]
- M. Geissler, J. Volk, F. Stehle, O. Kayser, H. Warzecha. Subcellular localization defines modification and production of Δ9-tetrahydrocannabinolic acid synthase in transiently transformed Nicotiana benthamiana. Biotechnol. Lett., 2018. [DOI | PubMed]
- F. Pollastro, D. Caprioglio, D. Del Prete, F. Rogati, A. Minassi, O. Taglialatela-Scafati, E. Munoz, G. Appendino. Cannabichromene. Nat. Prod. Commun., 2018. [DOI]
- E.B. Russo. Taming THC: Potential cannabis synergy and phytocannabinoid-terpenoid entourage effects. Br. J. Pharmacol., 2011. [DOI | PubMed]
- E.M.M. Ali, A.Z.I. Almagboul, S.M.E. Khogali, U.M.A. Gergeir. Antimicrobial activity of Cannabis sativa L.. Chin. Med., 2012. [DOI]
- G. Appendino, S. Gibbons, A. Giana, A. Pagani, G. Grassi, M. Stavri, E. Smith, M.M. Rahman. Antibacterial cannabinoids from Cannabis sativa: A structure-activity study. J. Nat. Prod., 2008. [DOI | PubMed]
- B. Van Klingeren, M. Ten Ham. Antibacterial activity of delta9-tetrahydrocannabinol and cannabidiol. Antonie Van Leeuwenhoek, 1976. [DOI | PubMed]
- T. Pellechia. Legal Cannabis Industry Poised for Big Growth, in North America and around the World. Forbes. 2018
- D.A. Kendall, G.A. Yudowski. Cannabinoid receptors in the central nervous system: Their signaling and roles in disease. Front. Cell. Neurosci., 2016. [DOI | PubMed]
- R. Mechoulam, L.A. Parker. The endocannabinoid system and the brain. Annu. Rev. Psychol., 2013. [DOI | PubMed]
- R. Mechoulam, M. Peters, E. Murillo-Rodriguez, L.O. Hanus. Cannabidiol—Recent advances. Chem. Biodivers., 2007. [DOI | PubMed]
- M.A. Huestis, R. Solimini, S. Pichini, R. Pacifici, J. Carlier, F.P. Busardò. Cannabidiol adverse effects and toxicity. Curr. Neuropharmacol., 2019. [DOI | PubMed]
- 38.European Monitoring Centre for Drugs and Drug Addiction (EMCDDA) Estimating the Size of the Main Illicit Drug Markets in Europe: An UpdateEMCDDALisbon, Portugal20199789294974440
- K. Blatt-Janmaat, Y. Qu. The biochemistry of phytocannabinoids and metabolic engineering of their production in heterologous systems. Int. J. Mol. Sci., 2021. [DOI | PubMed]
- C. Barreiro, C. García-Estrada. Recent developments in genome design and assembly tools. New Frontiers and Applications of Synthetic Biology, 2022
- J. Wang, Y. Yang, Y. Yan. Bioproduction of resveratrol. Biotechnology of Natural Products, 2018
- G. Wang, M. Huang, J. Nielsen. Exploring the potential of Saccharomyces cerevisiae for biopharmaceutical protein production. Curr. Opin. Biotechnol., 2017. [DOI | PubMed]
- J.L. Martínez, L. Liu, D. Petranovic, J. Nielsen. Pharmaceutical protein production by yeast: Towards production of human blood proteins by microbial fermentation. Curr. Opin. Biotechnol., 2012. [DOI | PubMed]
- C. Madzak. Yarrowia lipolytica: Recent achievements in heterologous protein expression and pathway engineering. Appl. Microbiol. Biotechnol., 2015. [DOI | PubMed]
- C. Madzak. Engineering Yarrowia lipolytica for Use in Biotechnological Applications: A Review of Major Achievements and Recent Innovations. Mol. Biotechnol., 2018. [DOI | PubMed]
- G.I. Naumov, E.S. Naumova, O.V. Tyurin, D.G. Kozlov. Komagataella kurtzmanii sp. nov., a new sibling species of Komagataella (Pichia) pastoris based on multigene sequence analysis. Antonie Van Leeuwenhoek, 2013. [DOI | PubMed]
- C.P. Kurtzman. Description of Komagataella phaffii sp. nov. and the transfer of Pichia pseudopastoris to the methylotrophic yeast genus Komagataella. Int. J. Syst. Evol. Microbiol., 2005. [DOI | PubMed]
- L. Gao, M. Cai, W. Shen, S. Xiao, X. Zhou, Y. Zhang. Engineered fungal polyketide biosynthesis in Pichia pastoris: A potential excellent host for polyketide production. Microb. Cell Fact., 2013. [DOI | PubMed]
- C. Liu, J.-S. Gong, C. Su, H. Li, H. Li, Z.-M. Rao, Z.-H. Xu, J.-S. Shi. Pathway engineering facilitates efficient protein expression in Pichia pastoris. Appl. Microbiol. Biotechnol., 2022. [DOI | PubMed]
- N.P. Keller. Fungal secondary metabolism: Regulation, function and drug discovery. Nat. Rev. Microbiol., 2019. [DOI | PubMed]
- V. Meyer. Genetic engineering of filamentous fungi—Progress, obstacles and future trends. Biotechnol. Adv., 2008. [DOI | PubMed]
- A.A. Sakekar, S.R. Gaikwad, N.S. Punekar. Protein expression and secretion by filamentous fungi. J. Biosci., 2021. [DOI | PubMed]
- M.A. Van Den Berg, R. Albang, K. Albermann, J.H. Badger, J.M. Daran, A.J.M. Driessen, C. Garcia-Estrada, N.D. Fedorova, D.M. Harris, W.H.M. Heijne. Genome sequencing and analysis of the filamentous fungus Penicillium chrysogenum. Nat. Biotechnol., 2008. [DOI | PubMed]
- C. Barreiro, C. García-Estrada. Proteomics and Penicillium chrysogenum: Unveiling the secrets behind penicillin production. J. Proteom., 2019. [DOI]
- C. García-Estrada, J.F. Martín, L. Cueto, C. Barreiro. Omics approaches applied to Penicillium chrysogenum and penicillin production: Revealing the secrets of improved productivity. Genes, 2020. [DOI | PubMed]
- L. García-Calvo, R. Rodríguez-Castro, R.V. Ullán, S.M. Albillos, M. Fernández-Aguado, C.M. Vicente, K.F. Degnes, H. Sletta, C. Barreiro. Penicillium chrysogenum as a fungal factory for feruloyl esterases. Appl. Microbiol. Biotechnol., 2023. [DOI | PubMed]
- F. Fierro, I. Vaca, N.I. Castillo, R.O. García-Rico, R. Chávez. Penicillium chrysogenum, a Vintage Model with a Cutting-Edge Profile in Biotechnology. Microorganisms, 2022. [DOI | PubMed]
- M.A. Van den Berg. Impact of the Penicillium chrysogenum genome on industrial production of metabolites. Appl. Microbiol. Biotechnol., 2011. [DOI | PubMed]
- K.D. Clevenger, J.W. Bok, R. Ye, G.P. Miley, M.H. Verdan, T. Velk, C. Chen, K. Yang, M.T. Robey, P. Gao. A scalable platform to identify fungal secondary metabolites and their gene clusters. Nat. Chem. Biol., 2017. [DOI | PubMed]
- F. Guzmán-Chávez, R.D. Zwahlen, R.A.L. Bovenberg, A.J.M. Driessen. Engineering of the Filamentous Fungus Penicillium chrysogenum as Cell Factory for Natural Products. Front. Microbiol., 2018. [DOI | PubMed]
- J. Arnau, D. Yaver, C.M. Hjort. Strategies and challenges for the development of industrial enzymes using fungal cell factories. Grand Challenges in Fungal Biotechnology, 2020
- P.J. Punt, N. van Biezen, A. Conesa, A. Albers, J. Mangnus, C. van den Hondel. Filamentous fungi as cell factories for heterologous protein production. Trends Biotechnol., 2002. [DOI | PubMed]
- J.-L. Adrio, A.L. Demain. Recombinant organisms for production of industrial products. Bioeng. Bugs, 2010. [DOI | PubMed]
- Y.-M. Chiang, C.E. Oakley, M. Ahuja, R. Entwistle, A. Schultz, S.-L. Chang, C.T. Sung, C.C.C. Wang, B.R. Oakley. An Efficient System for Heterologous Expression of Secondary Metabolite Genes in Aspergillus nidulans. J. Am. Chem. Soc., 2013. [DOI | PubMed]
- D.C. Anyaogu, U.H. Mortensen. Heterologous production of fungal secondary metabolites in Aspergilli. Front. Microbiol., 2015. [DOI | PubMed]
- Q. Wang, C. Zhong, H. Xiao. Genetic Engineering of Filamentous Fungi for Efficient Protein Expression and Secretion. Front. Bioeng. Biotechnol., 2020. [DOI | PubMed]
- W. Li, L. Cui, J. Mai, T.-Q. Shi, L. Lin, Z.-G. Zhang, R. Ledesma-Amaro, W. Dong, X.-J. Ji. Advances in Metabolic Engineering Paving the Way for the Efficient Biosynthesis of Terpenes in Yeasts. J. Agric. Food Chem., 2022. [DOI | PubMed]
- F. Taura, S. Tanaka, C. Taguchi, T. Fukamizu, H. Tanaka, Y. Shoyama, S. Morimoto. Characterization of olivetol synthase, a polyketide synthase putatively involved in cannabinoid biosynthetic pathway. FEBS Lett., 2009. [DOI | PubMed]
- F. Taura, S. Morimoto, Y. Shoyama. Purification and characterization of cannabidiolic-acid synthase from Cannabis sativa L. Biochemical analysis of a novel enzyme that catalyzes the oxidocyclization of cannabigerolic acid to cannabidiolic acid. J. Biol. Chem., 1996. [DOI | PubMed]
- M. Fellermeier, M.H. Zenk. Prenylation of olivetolate by a hemp transferase yields cannabigerolic acid, the precursor of tetrahydrocannabinol. FEBS Lett., 1998. [DOI | PubMed]
- S. Sirikantaramas, S. Morimoto, Y. Shoyama, Y. Ishikawa, Y. Wada, Y. Shoyama, F. Taura. The gene controlling marijuana psychoactivity: Molecular cloning and heterologous expression of Delta1-tetrahydrocannabinolic acid synthase from Cannabis sativa L.. J. Biol. Chem., 2004. [DOI | PubMed]
- J.M. Stout, Z. Boubakir, S.J. Ambrose, R.W. Purves, J.E. Page. The hexanoyl-CoA precursor for cannabinoid biosynthesis is formed by an acyl-activating enzyme in Cannabis sativa trichomes. Plant J., 2012. [DOI | PubMed]
- F. Taura, E. Dono, S. Sirikantaramas, K. Yoshimura, Y. Shoyama, S. Morimoto. Production of Delta(1)-tetrahydrocannabinolic acid by the biosynthetic enzyme secreted from transgenic Pichia pastoris. Biochem. Biophys. Res. Commun., 2007. [DOI | PubMed]
- K. Lange, A. Schmid, M.K. Julsing. Δ(9)-Tetrahydrocannabinolic acid synthase production in Pichia pastoris enables chemical synthesis of cannabinoids. J. Biotechnol., 2015. [DOI | PubMed]
- B. Zirpel, F. Stehle, O. Kayser. Production of Δ9-tetrahydrocannabinolic acid from cannabigerolic acid by whole cells of Pichia (Komagataella) pastoris expressing Δ9-tetrahydrocannabinolic acid synthase from Cannabis sativa L.. Biotechnol. Lett., 2015. [DOI | PubMed]
- B. Zirpel, F. Degenhardt, C. Martin, O. Kayser, F. Stehle. Engineering yeasts as platform organisms for cannabinoid biosynthesis. J. Biotechnol., 2017. [DOI | PubMed]
- B. Zirpel, F. Degenhardt, C. Zammarelli, D. Wibberg, J. Kalinowski, F. Stehle, O. Kayser. Optimization of Δ9-tetrahydrocannabinolic acid synthase production in Komagataella phaffii via post-translational bottleneck identification. J. Biotechnol., 2018. [DOI]
- B. Zirpel, O. Kayser, F. Stehle. Elucidation of structure-function relationship of THCA and CBDA synthase from Cannabis sativa L.. J. Biotechnol., 2018. [DOI | PubMed]
- F. Thomas, C. Schmidt, O. Kayser. Bioengineering studies and pathway modeling of the heterologous biosynthesis of tetrahydrocannabinolic acid in yeast. Appl. Microbiol. Biotechnol., 2020. [DOI | PubMed]
- T. Gülck, J.K. Booth, Â. Carvalho, B. Khakimov, C. Crocoll, M.S. Motawia, B.L. Møller, J. Bohlmann, N.J. Gallage. Synthetic Biology of Cannabinoids and Cannabinoid Glucosides in Nicotiana benthamiana and Saccharomyces cerevisiae. J. Nat. Prod., 2020. [DOI | PubMed]
- Q. Zhang, W. Zeng, S. Xu, J. Zhou. Metabolism and strategies for enhanced supply of acetyl-CoA in Saccharomyces cerevisiae. Bioresour. Technol., 2021. [DOI | PubMed]
- S. Dusséaux, W.T. Wajn, Y. Liu, C. Ignea, S.C. Kampranis. Transforming yeast peroxisomes into microfactories for the efficient production of high-value isoprenoids. Proc. Natl. Acad. Sci. USA, 2020. [DOI | PubMed]
- J.L. Poulos, A.N. Farnia. Production of Cannabinoids in Yeast 2016. WO2016010827A1. Librede Inc, USA
- B.K. Szamecz, S. Varszegi, A. Nemeth, L. Szabo. Microorganisms and Methods for the Fermentation of Cannabinoids 2020. WO2019071000A1. INTREXON CORPORATION, USA
- R.T. Sayre, E. Carvalho-Gonçalves, T. Zidenga. Generation of Water-Soluble Cannabinoid Compounds in Yeast and Plant Cell Suspension Cultures and Compositions of Matter 2019. US20190078168A1. Trait Biosciences Inc., USA
- S. Mookerjee, A.J. Campbell, Z.D. Wiltshire, K.J. Chen. Method and Cell Line for Production of Phytocannabinoids and Phytocannabinoid Analogues in Yeast. WO2018148848A1. Hyasynth Biologicals Inc., Canada. 2020
- J.D. Keasling, L. D’espaux, J. Wong, X. Luo, M. Reiter, C. Denby, A. Lechner. Recombinant Microorganisms and Methods for Producing Cannabinoids and Cannabinoid Derivatives 2020. US10563211B2. The Regents of the University of California, USA
- O. Kayser, F.-O. Stehle. Biotechnological Production of Cannabinoids 2020. WO2020016287A1. Technische Universität Dortmund, Germany
- A. Horwitz, L. D’espaux, J. Wong, R. Bector, A.K. Hjelmeland, D. Platt, J. Ubersax. Optimized Expression Systems for Producing Cannabinoid Synthase Polypeptides, Cannabinoids, and Cannabinoid Derivatives 2020. WO2020069214A3. Demetrix Inc., USA
- J.U. Bowie, T.P. Korman, M. Valliere. Biosynthetic Platform for the Production of Cannabinoids and other Prenylated Compounds 2021. WO2020028722A1. The Regents of the University of California, USA
- T.A. Beardslee. Biosynthetic Cannabinoid Production in Engineered Microorganisms 2020. WO2020198679A1. Rynetech Bio Inc., USA
- J. Ma, Y. Gu, P. Xu. Biosynthesis of cannabinoid precursor olivetolic acid in genetically engineered Yarrowia lipolytica. Commun. Biol., 2022. [DOI | PubMed]
- I.C. Okorafor, M. Chen, Y. Tang. High-Titer Production of Olivetolic Acid and Analogs in Engineered Fungal Host Using a Nonplant Biosynthetic Pathway. ACS Synth. Biol., 2021. [DOI | PubMed]
- R.P. Elander. Industrial production of beta-lactam antibiotics. Appl. Microbiol. Biotechnol., 2003. [DOI | PubMed]
- J.M. Cantoral, S. Gutiérrez, F. Fierro, S. Gil-Espinosa, H. van Liempt, J.F. Martín. Biochemical characterization and molecular genetics of nine mutants of Penicillium chrysogenum impaired in penicillin biosynthesis. J. Biol. Chem., 1993. [DOI | PubMed]
- F. Fierro, E. Montenegro, S. Gutiérrez, J.F. Martín. Mutants blocked in penicillin biosynthesis show a deletion of the entire penicillin gene cluster at a specific site within a conserved hexanucleotide sequence. Appl. Microbiol. Biotechnol., 1996. [DOI | PubMed]
- F. Teufel, J.J. Almagro Armenteros, A.R. Johansen, M.H. Gíslason, S.I. Pihl, K.D. Tsirigos, O. Winther, S. Brunak, G. von Heijne, H. Nielsen. SignalP 6.0 predicts all five types of signal peptides using protein language models. Nat. Biotechnol., 2022. [DOI | PubMed]
- M.S. Jami, C. García-Estrada, C. Barreiro, A.A. Cuadrado, Z. Salehi-Najafabadi, J.F. Martín. The Penicillium chrysogenum extracellular proteome. Conversion from a food-rotting strain to a versatile cell factory for white biotechnology. Mol. Cell. Proteom., 2010. [DOI | PubMed]
- M.S. Jami, C. Barreiro, C. García-Estrada, J.F. Martín. Proteome analysis of the penicillin producer Penicillium chrysogenum: Characterization of protein changes during the industrial strain improvement. Mol. Cell. Proteom., 2010. [DOI]
- B. De Backer, B. Debrus, P. Lebrun, L. Theunis, N. Dubois, L. Decock, A. Verstraete, P. Hubert, C. Charlier. Innovative development and validation of an HPLC/DAD method for the qualitative and quantitative determination of major cannabinoids in cannabis plant material. J. Chromatogr. B, 2009. [DOI | PubMed]
- M.A. Valliere, T.P. Korman, M.A. Arbing, J.U. Bowie. A bio-inspired cell-free system for cannabinoid production from inexpensive inputs. Nat. Chem. Biol., 2020. [DOI | PubMed]