
The Inflammatory Signals Associated with Psychosis: Impact of Comorbid Drug Abuse
Abstract
Psychosis and substance use disorders are two diagnostic categories whose association has been studied for decades. In addition, both psychosis spectrum disorders and drug abuse have recently been linked to multiple pro-inflammatory changes in the central nervous system. We have carried out a narrative review of the literature through a holistic approach. We used PubMed as our search engine. We included in the review all relevant studies looking at pro-inflammatory changes in psychotic disorders and substance use disorders. We found that there are multiple studies that relate various pro-inflammatory lipids and proteins with psychosis and substance use disorders, with an overlap between the two. The main findings involve inflammatory mediators such as cytokines, chemokines, endocannabinoids, eicosanoids, lysophospholipds and/or bacterial products. Many of these findings are present in different phases of psychosis and in substance use disorders such as cannabis, cocaine, methamphetamines, alcohol and nicotine. Psychosis and substance use disorders may have a common origin in an abnormal neurodevelopment caused, among other factors, by a neuroinflammatory process. A possible convergent pathway is that which interrelates the transcriptional factors NFκB and PPARγ. This may have future clinical implications.
Article type: Review Article
Keywords: psychosis, addiction, neuroinflammation, NFκB, PPARγ
Affiliations: Unidad de Gestión Clínica de Salud Mental, Instituto de Investigación Biomédica de Málaga (IBIMA), Hospital Regional Universitario de Málaga, 29010 Málaga, Spain; Facultad de Medicina, Universidad de Málaga, Andalucía Tech, Campus de Teatinos s/n, 29071 Málaga, Spain; Departamento de Farmacología y Pediatría, Universidad de Málaga, Andalucía Tech, Campus de Teatinos s/n, 29071 Málaga, Spain; Facultad de Psicología, Universidad de Málaga, Andalucía Tech, Campus de Teatinos s/n, 29071 Málaga, Spain
License: © 2023 by the authors. CC BY 4.0 Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Article links: DOI: 10.3390/biomedicines11020454 | PubMed: 36830990 | PMC: PMC9953424
Relevance: Moderate: mentioned 3+ times in text
Full text: PDF (1.0 MB)
1. Introduction
Psychosis is generally defined as a mental phenomenon of a pathological nature in which the affected subject experiences a loss of contact with reality. In classical psychiatry, “psychosis” was conceptualized as a large syndrome opposite to “neurosis”, in the form of a characteristic structure that begins to form in childhood and hatches and develops at the beginning of adulthood. Today, psychosis is considered as a common and functional set of symptoms that are present in many psychiatric, neurological, medical and neurodevelopmental conditions [ref. 1].
Despite the advances made in the field of the study of the causes and factors involved in the pathogenesis of psychosis, there are still unresolved questions; thus, it remains an old aspiration of biological psychiatry to find biomarkers that can serve as diagnostic criteria for the different disorders of the psychotic spectrum. A new approach to the phenomenon is to “deconstruct” psychosis in its clinical dimensions to identify the pathogenic causes and mechanisms of each of its symptoms and psychopathological dimensions. Thus, we will be able to understand their interaction in individual cases [ref. 2].
Why does psychosis occur? This is one of the questions that researchers continue to ask themselves around the world. Although more and more light has been shed on the biological processes that lead a subject to lose touch with their surrounding reality, the truth is that there is still a long way to go. The classical hypothesis that explains the development of psychotic disorders, taking schizophrenia as a paradigmatic example, is the dopaminergic hypothesis. According to this theory, based largely on the efficacy of drugs that act on this pathway of the central nervous system, an aberrant neurotransmission in certain brain circuits would be ultimately responsible for the appearance of psychotic symptoms. Roughly speaking, we could talk about an “excess” of dopamine in the mesolimbic pathways—responsible for the positive symptoms—and a “defect” of dopamine in the mesocortical pathways—responsible for the negative symptoms [ref. 3]. However, how does this aberrant neurotransmission in the brain of an adolescent or young adult occur?
1.1. Pathogenic Factors of Psychosis
Classically, researchers have focused on studying a number of pathogenic factors that could explain the etiology of psychotic spectrum disorders. For example, gene markers have been identified that may be related to the development of psychosis. Therefore, large-scale studies on the genomics of psychiatric diseases are beginning to illuminate genetic–molecular contributions to major mental disorders. On the one hand, we find common variants with a small effect size and, on the other, rare mutations with a larger effect size [ref. 4,ref. 5,ref. 6,ref. 7].
Another aspect long studied is childhood trauma. The literature indicate that the proportion of subjects with psychotic symptoms (especially hallucinations) who have suffered abuse in childhood is much higher than in the general population, indicating as possible bridges between both induction of social defeat and reduced self-value, sensitization of the mesolimbic dopamine system, changes in the stress and immune system, and concomitant changes in stress-related brain structures, such as the hippocampus and the amygdala [ref. 8,ref. 9].
However, if there is a risk factor long studied and related to the development of psychosis, it is undoubtedly the consumption of drugs of abuse [ref. 10,ref. 11]. For example, cannabis abuse induces transient psychotic disorders and is a predictor for an increased risk of schizophrenia (especially in young adults) [ref. 12]. Furthermore, it is associated with an earlier onset of psychosis, as well as transitioning to it in high-risk subjects [ref. 13]. Cocaine use has traditionally been associated with various psychiatric problems [ref. 14] and the appearance of psychotic symptoms is one of the most common complications [ref. 15]. Since cocaine has an agonist effect on dopaminergic neurotransmission at the central level, it is quite common for transient psychotic symptoms to occur in the context of acute intoxication [ref. 16]. On the other hand, in patients with primary psychotic disorders, such as schizophrenia, cocaine use is higher than in the general population [ref. 17], and its consumption has been associated with an earlier onset of psychotic illness [ref. 18]. Another psychostimulant with psychotic–mimetic properties similar to cocaine is methamphetamine (MA) [ref. 19]. Alcohol consumption and tobacco smoking have also been strongly associated with the presence of psychosis [ref. 20,ref. 21].
In analyzing the relationship between psychosis and drug abuse, a diagnostic and prognostic difficulty lies in the distinction between primary and substance-induced psychoses. Notably, due to the increasing use of drugs that possess the potential to produce psychosis, substance-induced psychotic symptoms are becoming a relatively frequent phenomenon. It has been reported that the onset of psychosis can take place up to 2 years earlier in substance abusers (and 2.7 years earlier if only cannabis is considered) [ref. 22]. Also, the propensity to develop psychosis seems to be related to the severity of use and dependence [ref. 11].
1.2. Psychosis, Drug Abuse and Neuroinflammation
Recently, the study of immuno-inflammatory factors that may be involved in the genesis of psychosis has become more important, describing a “pro-inflammatory” state enhanced in these disorders [ref. 23]. In relation to the inflammatory processes that may be underlying these alterations of the immune system, different hypotheses have been formulated: from the macrophage-T lymphocyte theory of Smith and Maes [ref. 24,ref. 25] to the theory of Schwarz et al. [ref. 26], who proposed the Th2 hypothesis. In turn, Monji et al. [ref. 27] described the microglia hypothesis: that activated central nervous system microglia release pro-inflammatory cytokines and free radicals, which induce abnormal neurogenesis that contributes to the pathophysiology of schizophrenia.
The study of the question is complicated, since drug abuse, in addition to being able to act as an independent risk factor for psychosis, has also been linked to immuno-inflammatory factors. Evidence suggests glial cells have an essential role in the development and maintenance of drug abuse by influencing neuronal and synaptic functions in multifaceted ways. For example, microglia and astrocytes perform critical functions in synapse formation and refinement in the developing brain [ref. 28,ref. 29].
Could substance use disorders and psychosis have a common origin? Are there biomarkers that can differentiate primary psychoses from induced psychoses? Below, we will discuss a number of inflammatory markers that have been linked to the onset of schizophrenia and other related diseases, and drug abuse. Our aim was to test the hypothesis that there could be an overlap between the different biomarkers found in psychosis disorders and substance use disorders, which would be able to tell us about a common pathway.
Subsequently, we try to relate these findings and ask ourselves a series of questions that can have a clinical impact on the care of our patients.
2. Methods
We carried out a narrative review of the literature through a search of studies in PubMed. We entered as search terms “psychotic disorders”, “schizophrenia”, “substance use disorder”, “cytokines”, “chemokines”, “endocannabinoid system”, “eicosanoids”, “lysophoshpolipids” and “bacterial products”. We filtered those studies that, from a narrative and nonsystematic approach, seemed most relevant to meet the aims of our work. We focused on clinical studies with human participants, although sometimes we cite findings from preclinical studies, especially when data in patients are scarce. At the same time, we did not consider for review those studies that only provided redundant information (for example, original research that was contained in already selected reviews).
3. Results
We summarized the main features and aims of the studies reviewed in Table 1 and Table 2. We integrated the results of the studies and present these in the following subsections.
Table 1: Reviewed studies in psychotic disorders.
| Study | Type | Population | Aim |
|---|---|---|---|
| García-Bueno et al.[ref. 30] | Observational original research | 117 patients (mean age 23.91 ± 5.83 years; 69.2% male) during the first year after their first episode of psychosis (FEP) according to the DSM-IV criteria and 106 matched controls | To contrast the hypothesis that the physiological balance between interrelated pro-inflammatory/anti-inflammatory pathways may be disrupted in FEP |
| Delaney et al.[ref. 31] | Observational original research | 47 children, adolescents and young adults with psychosis (mean age 24.30 ± 5.91; 63.83% male), 17 individuals at clinical high risk for psychosis (mean age 22.76 ± 3.68; 64.71% male) and 33 unaffected comparison-matched controls | To compare the levels of vitamin D, C-reactive protein (CRP), antibodies to lipopolysaccharide (LPS) and IL-6 between the groups |
| Park et al.[ref. 32] | Systematic review | 8 studies of subjects at high-risk for psychosis (mean age range from 16.1 to 26.2; 31–100% male) versus matched controls (n range for cases from 14 to 76, and for controls from 39 to 115); 4 studies of high-risk converters versus non-converters (n range for converters from 14 to 56, and for non-converters from 60 to 129; mean age range from 16.1 to 21.9; 31–69% male) | To perform a meta-analysis of cytokine and C-reactive protein levels in high-risk psychosis |
| Dawidowski et al.[ref. 33] | Review | N/A | To present the most valuable evidence on cytokine dysregulation in schizophrenia, the links between cytokine levels and psychopathological presentation, as well as their alterations in response to antipsychotics |
| Lesh et al.[ref. 34] | Observational original research | 69 first-episode schizophrenia-spectrum patients (mean age 19.9 ± 3.5; 88% male), 16 first-episode bipolar patients with psychotic features (mean age 21.4 ± 3.4 years; 75% male) and 53 healthy matched controls | To investigate differences in cytokine levels in plasma between individuals with first-episode schizophrenia, first-episode bipolar disorder with psychotic features, and healthy controls |
| Boczek et al.[ref. 35] | Review | N/A | To provide a summary of GPCR (G protein-coupled receptor)-acting neurotransmitters and chemokines and their role in schizophrenia |
| Ellman et al.[ref. 36] | Observational original research | 17 cases diagnosed with schizophrenia (mean age 39.96 ± 1.78 years; 70.59% male) and 8 healthy matched controls | To determine the association between fetal exposure to IL-8 and structural brain changes among schizophrenia cases and controls |
| Brown et al.[ref. 37] | Review | N/A | To discuss the contribution of exposure to infection to the etiology of schizophrenia |
| Michino et al.[ref. 38] | Systematic review | 18 studies in subjects with schizophrenia-related illnesses (44.3–75% male) versus healthy matched controls (n range for cases from 10 to 162, and for controls from 11 to 94). | To conduct a systematic review and meta-analysis of the blood and cerebrospinal fluid (CSF) measures of the endocannabinoid system (ECS) in psychotic disorders |
| Leweke et al.[ref. 39] | Observational original research | 25 first-episode, antipsychotic-naïve schizophrenics with low frequency cannabis use (mean age 28.4 ± 8.8 years; 64% male); 19 first-episode, antipsychotic-naïve schizophrenics with high frequency of cannabis use (mean age 30.3; 84.21% male); and 81 healthy matched controls | To examine how cannabis use alters levels of anandamide in cerebrospinal fluid (CSF) in schizophrenic patients |
| Giuffrida et al.[ref. 40] | Observational original research | 47 antipsychotic-naïve first-episode paranoid schizophrenics (mean age 28.9 ± 9 years; 53.6% male), 13 dementia patients (mean age 77.8 ± 7.8 years; 46.2% male), 22 affective disorder patients (mean age 44.7 ± 15.8 years; 46.2% male), 71 schizophrenics patients treated with antipsychotics (mean age 29.1 ± 10.1 years; 80.3% male), and 84 volunteers with no family history of psychiatric disturbances (mean age 27.9 ± 2.8 years; 53.6% male) | To examine the role of endocannabinoid signaling in psychotic states by measuring levels of the endocannabinoid anandamide in cerebrospinal fluid (CSF) of acute paranoid-type schizophrenic patients |
| Chase et al.[ref. 41] | Observational original research | 35 participants with schizophrenia (mean age 36.14 ± 11.64 years; 51.43% male) and 35 healthy matched controls | To measure mRNA levels of cannabinoid receptors (CBRs) in human peripheral blood mononuclear cells (PBMCs) |
| Smesny et al.[ref. 42] | Observational original research | 48 patients with first episode of schizophrenia (mean age 19.41 ± 2.94 years; 72.92% male) and 22 healthy matched controls | To investigate if intracellular phospholipases A2 (inPLA2) activity is associated with symptoms severity and treatment response in first-episode schizophrenia (FES) |
| Noponen et al.[ref. 43] | Observational original research | 39 schizophrenics patients (mean age 44.8 ± 10.9 years; 100% male), 26 psychiatric non-schizophrenics patients (mean age 47.2 ± 12.6 years; 100% male) and 26 non-psychiatric and healthy volunteers (mean age 38.3 ± 11 years; 53.85% male) | To measure serum phospholipase A2 (PLA2) activity in all participants |
| Gattaz et al.[ref. 44] | Observational original research | 20 paranoid schizophrenic patients (mean age 32 ± 12 years; 45% male), 6 non-schizophrenic psychiatric patients (mean age 37 ± 11 years; 0% male), and 21 non-psychiatric and healthy volunteers (mean age 31 ± 7 years; 47.62% male) | To investigate the activity of phospholipase A2 in the plasma of schizophrenic patients and healthy controls, as well as in a small group of non-schizophrenic psychiatric patients |
| Pereira et al.[ref. 45] | Observational original research | 67 ultra-high-risk (UHR) for psychosis individuals (mean age 23.5 ± 3.4 years; 41.79% male) and 55 healthy matched controls | To investigate whether the study of the inflammatory COX-2 pathway through the quantification of the eicosanoid levels can be a useful approach for the characterization of ultra-high-risk (UHR) for psychosis individuals |
| Bowden et al.[ref. 46] | Observational original research | 14 individuals with schizophrenia (mean age 35.86 years; 78.57% male) and 14 non-psychiatric matched controls | To generate gene expression profiles from peripheral blood lymphocytes from participants |
| Omori et al.[ref. 47] | Observational original research | 27 patients with schizophrenia (mean age 40.1 ± 10.0 years; 51.9% male), 26 patients with major depressive disorder (mean age 40.4 ± 8.3 years; 50% male) and 27 healthy matched controls | To investigate the levels of lysophospholipid species, including LPA and related metabolic enzymes, in cerebrospinal fluid (CSF) of patients with major depressive disorder and schizophrenia |
| Gotoh et al.[ref. 48] | Observational original research | Study 1 (cerebrospinal fluid levels): 49 patients with schizophrenia (mean age 39.7 ± 10.5 years; 48.98% male) and 49 healthy matched controls;Study 2 (plasma levels): 42 patients with schizophrenia (mean age 40.7 ± 8.9 years; 54.76% male) and 44 healthy matched controls | To measure lysophosphatidic acid (LPA) levels by enzyme-linked immunosorbent assay in cerebrospinal fluid (CSF) (study 1) and plasma (study 2) samples |
| Madrid-Gambín et al.[ref. 49] | Observational original research | 48 patients with psychotic experiences at 18 years of age but who did not have psychotic disorder (45.83% male) and 67 controls (58.2% male) of the same birth cohort | To identify early biomarkers of psychotic experiences in plasma samples of the participants when they were 12 years old |
| Maes et al.[ref. 50] | Secondary data analysis | Subjects with first episode of psychosis and schizophrenia | To delineate a) the characteristics of the protein–protein interaction (PPI) network of antipsychotic-naïve first-episode psychosis (AN-FEP) and its transition to schizophrenia; and b) the biological functions, pathways and molecular patterns which are over-represented in FEP/schizophrenia |
| Maes et al.[ref. 51] | Observational original research | 21 subjects with first episode of schizophrenia (mean age 38.4 ± 12 years; 47.62% male), 58 subjects with multiple-episode schizophrenia (mean age 41.9 ± 10.7 years; 59.6% male), and 40 healthy matched controls | To delineate (a) the differences in several pathological and neuroimmune pathways between stable-phase, first- (FES) and multiple (MES)-episode schizophrenia and (b) the pathways that determine the behavioral–cognitive–physical–psychosocial (BCPS) deterioration in FES/MES |
| Maes et al.[ref. 52] | Observational original research | 80 schizophrenia patients and 38 healthy controls (overall mean age 39.95 years; 55.93% male) | To measure plasma IgA/IgM responses to five Gram-negative bacteria in association with IgM responses to malondialdehyde (MDA) and azelaic acid in participants |
Table 2: Reviewed studies in substance use disorders.
| Study | Type | Population | Aim |
|---|---|---|---|
| Bayazit et al.[ref. 53] | Observational original research | 34 patients with cannabis use disorder (mean age of 26 ± 9 years; 100% male) and 34 healthy matched controls | To evaluate oxidant and antioxidant status and cytokine levels in individuals with cannabis use disorder |
| Pacifi et al.[ref. 54] | Observational original research | 37 polydrug consumers of 3,4-methylenedioxymethamphetamine (MDMA) and cannabis (mean age 23.6 ± 3.5 years; 51.4% male) compared to 23 cannabis users only (mean age 22 ± 1.9 years; 34.8% male) and 34 non-consumers (mean age 22 ± 2.6 years; 26.5% male) | To investigate cell-mediated immune function and the occurrence of mild infectious diseases in participants |
| Fernandez-Egea et al.[ref. 55] | Observational original research | 18 current cannabis users (mean age 22.3 ± 5.1 years; 66% male), 33 past cannabis users (mean age 23.6 ± 4.3 years; 66% male) and 36 subjects who never used cannabis (mean age 24.3 ± 4.7 years; 68% male) | To explore the possibility that cannabis use influenced CCL11 chemokine plasma levels |
| Ribeiro et al.[ref. 56] | Observational original research | 21 cannabis users (mean age 28.6 ± 8.54 years; 100% male), 12 cocaine users (mean age 37.8 ± 4.87 years; 100% male), 27 cannabis-plus-cocaine users (mean age 32.3 ± 7.91 years; 100% male) and 21 non-drug users (mean age 33.42 ± 9.73 years; 100% male) | To investigate the effects of illicit drugs on circulating lipopolysaccharide (LPS), systemic inflammation and oxidative stress markers in drug users |
| Keen et al.[ref. 57] | Observational original research | 77 lifetime non-drug users (32% male), 46 lifetime marijuana only users (67% male) and 45 lifetime marijuana and other drug users (60% male); overall median age = 47 years | To explore potential differential effects of lifetime marijuana use on interleukin-1 alpha (IL-1α) and tumor necrosis factor (TNF) in a community-based sample |
| Araos et al.[ref. 58] | Observational original research | 82 abstinent cocaine users who sought outpatient cocaine treatment (mean age 36.9 ± 7.8 years; 18.3% male) and 65 healthy matched controls | To examine the plasma pro-inflammatory cytokine and chemokine profile in participants |
| Moreira et al.[ref. 59] | Observational original research | 12 cocaine users (mean age 24.92 ± 4.80 years; 75% male) and 24 healthy matched controls | To investigate serum levels of pro and anti-inflammatory cytokines, IL-6 and IL-10, respectively, in cocaine users from a young population-based sample |
| Maza-Quiroga et al.[ref. 60] | Observational original research | 79 patients diagnosed with cocaine use disorder (CUD) in abstinence (34.87 ± 7.18 years) and 81 healthy matched controls | To test the hypothesis that patients with CUD in abstinence might have altered circulating levels of signaling proteins related to systemic inflammation |
| Halpern et al.[ref. 61] | Experimental original research | 30 healthy participants with a history of cocaine use (16 females (mean age 25.8 ± 1.0 years)) and 14 males (mean age 27.8 ± 1.5 years)) | To measure neuroendocrine and immunological responses to IV injection of 0.4 mg/kg cocaine or saline placebo |
| Pedraz et al.[ref. 62] | Observational original research | 55 abstinent cocaine-addicted subjects diagnosed with lifetime cocaine use disorders (40 men (mean age 37.1 ± 6.7 years) and 15 women (mean age 42.8 ± 6.2 years)) and 73 healthy matched controls | To evaluate the sex differences in psychiatric comorbidity and the concentrations of plasma mediators that have been reported to be affected by cocaine |
| Levandowski et al.[ref. 63] | Observational original research | 42 crack cocaine-dependent women (mean age 31.22 ± 7.83 years) and 52 healthy matched controls | To investigate the association between plasma interleukin 6 (IL-6) levels and executive function (EF) in crack cocaine-dependent women |
| Anchur et al.[ref. 64] | Review | N/A | To discuss cytokine biomarker candidates for alcohol abuse and alcoholism |
| Moura et al.[ref. 65] | Systematic review | 23 studies of subjects with alcohol use disorder (56–100% male) and healthy matched controls (n range for cases from 9 to 42, and for controls from 6 to 46) | To assess differences in blood inflammatory cytokines between people with alcohol use disorder (AUD) and healthy controls (HC) |
| Crews et al.[ref. 66] | Review | Alcohol users | To discuss the contribution of cytokines in alcohol use and alcoholic pathologies |
| García-Marchena et al.[ref. 67] | Observational original research | 85 abstinent subjects with alcohol use disorders (mean age 47.16 ± 7.27 years; 68.24% male) and 55 healthy matched controls | To explore possible associations in circulating plasma cytokine concentrations in abstinent patients diagnosed with alcohol use disorders |
| Fox et al.[ref. 68] | Experimental original research | 33 alcohol-dependent individuals (12 with low depressive symptoms (mean age 38.7 ± 2.6 years; 86% male) and 21 with high depressive symptoms (mean age 39.1 ± 2.4 years; 84% male)) and 37 social drinkers (21 with low depressive symptoms (mean age 31.9 ± 2.2 years; 62% male) and 16 with high depressive symptoms (mean age 35.7 ± 2.4 years; 38% male)) | To examine cytokine responses to stress in alcohol-dependent individuals and social drinkers, both with and without subclinical depression |
| Clark et al.[ref. 69] | Review | N/A | To review evidence that psychostimulants of abuse (cocaine, methamphetamine, ecstasy) are associated with activation of the innate immune response |
| Loftis et al.[ref. 70] | Observational original research (+preclinical experimental model) | 20 adults in remission from methamphetamine (MA) dependence (mean age 33.3 ± 1.84 years; 70% male) and 20 healthy matched controls (+preclinical model)Preclinical model: 32 male C57BL/6J mice that were administered MA (1 mg/kg) or saline subcutaneously for 7 consecutive days | To test the hypothesis that immune factors, such as cytokines, chemokines and cellular adhesion molecules, contribute to MA-induced immune dysfunction, neuronal injury and persistent cognitive impairments |
| Yamamoto et al.[ref. 71] | Review | N/A | To discuss long-term decreases in markers of biogenic amine neurotransmission in methamphetamine and MDMA users |
| Yamamoto et al.[ref. 72] | Review | N/A | To highlight experimental evidence that methamphetamine and MDMA increase oxidative stress, produce mitochondrial dysfunction and increase inflammation, all of which converge and culminate in long-term toxicity to dopaminergic and serotonergic neurons |
| Qiu et al.[ref. 73] | Review | N/A | To review the influence of smoking on major components of both innate and adaptive immune cells, and summarize cellular and molecular mechanisms underlying effects of cigarette smoking on the immune system |
| Lee et al.[ref. 74] | Review | N/A | To discuss specific mechanisms by which cigarette smoking affects host immunity |
| Walter et al.[ref. 75] | Experimental original research(+preclinical experimental model) | 15 healthy young men (mean age, 28.1 ± 3.1 years) and 15 women (mean age, 26.6 ± 2.4 years);preclinical model: 20 female Lewis rats (10 rats per treatment group) that were administered 3 mg/kg of the CB1/CB2-agonist WIN 55,212-2 dissolved in 1:1 dimethyl sulfoxide/phosphate buffer | To investigate effects in circulating concentrations of endocannabinoids after administration of a single oral dose of 20 mg delta9-tetrahydrocannabinol (THC) to 30 healthy volunteers and comparison with placebo |
| Manza et al.[ref. 76] | Observational original research | 89 individuals with cannabis dependence (mean age 28.6 ± 3.5 years; 71.9% male) and 89 healthy matched controls | To lend insight into biological processes that might link chronic cannabis use to brain structural abnormalities |
| Spindle et al.[ref. 77] | Experimental original research | 10 females with cannabis use disorder (mean age 23.2 ± 2.7 years), 10 female healthy controls (mean age 25.5 ± 5 years) and 7 male non-cannabis consumers (mean age 29.6 ± 6.9 years) | To explore the relation between acute cannabis effects and mood/craving/withdrawal and CB1 receptor availability |
| Ceccarini et al.[ref. 78] | Observational original research | 10 chronic cannabis users (mean age 26.0 ± 4.1 years; 80% male) and 10 healthy matched controls | To test that chronic cannabis use may alter specific regional CB1 receptor expression |
| Hirvonen et al.[ref. 79] | Observational original research | 30 male cannabis smokers (mean age 28 ± 8 years) and 28 healthy matched controls | To demonstrate downregulation of brain cannabinoid CB1 (cannabinoid receptor type 1) receptors after chronic exposure to cannabis in humans |
| Pavon et al.[ref. 80] | Observational original research | 88 abstinent cocaine addicts (mean age 36.9 ± 8.4 years; 88.6% male) and 46 matched healthy control | To evaluate circulating endocannabinoid-related lipids as biomarkers of cocaine use disorder |
| Voegel et al.[ref. 81] | Observational original research | 73 chronic cocaine users (mean age 31.3 ± 9.5 years; 67.12% male) and 67 healthy matched controls | To investigate alterations of hypothalamic–pituitary–adrenal (HPA) axis and endocannabinoid (eCB) system markers in individuals with chronic cocaine use disorder |
| Kunos et al.[ref. 82] | Review | N/A | To show evidence accumulated over the last two decades to indicate that both the addictive neural effects of ethanol and its organ toxic effects in the liver and elsewhere are mediated, to a large extent, by endocannabinoid signaling |
| García-Marchena et al.[ref. 83] | Observational original research | 79 abstinent alcohol-dependent subjects (49.13 ± 9.6 years; 65.8% male) and 79 healthy matched controls | To characterize the plasma acylethanolamides in alcohol dependence |
| Yang et al.[ref. 84] | Review | N/A | To discuss the role of cyclooxygenase-2 (COX-2) in synaptic signaling |
| Copeland et al.[ref. 85] | Experimental preclinical study | Preclinical model: 51 young, male rabbits that were randomly selected to receive subcutaneous injections of either cocaine or saline solution | To study cocaine-induced alterations in cerebrovascular function and metabolism |
| Mastrogiannis et al.[ref. 86] | Experimental in vitro model | Endothelial cells were isolated from human umbilical veins derived from uncomplicated pregnancies with a negative history of drug abuse | To investigate the possible effects of cocaine on prostacyclin and prostaglandin (PG) E2 production from endothelial cells derived from human umbilical cord |
| Anton et al.[ref. 87] | Review | N/A | To review the nature and role of prostaglandins in the central nervous system, and what is known about the effect of ethanol on prostaglandin production in brain |
| Murphy et al.[ref. 88] | Review | N/A | To discuss effects of ethanol in synthesis of prostaglandins and leukotrienes |
| García-Marchena et al.[ref. 89] | Observational original research | 55 abstinent alcohol use disorder (AUD) patients (mean age 47.7 ± 7.7 years; 81.8% male) and 34 healthy matched controls | To investigate whether the relevant species of LPA were associated with clinical features of alcohol addiction |
3.1. Inflammation and Psychosis
Below we discuss several immuno-inflammatory markers that have been linked to psychosis spectrum disorders. We have summarized the data in Table 3.
Table 3: Immuno-inflammatory markers in psychotic spectrum disorders.
| Type of Biomarker | Main Findings | References |
|---|---|---|
| Cytokines | ↑ Pro-inflammatory cytokines↓ Anti-inflammatory cytokines↑ IL-6 as a trait marker of schizophrenia (lower level of evidence for IL-1ß and TNF-α)↑ IFN-γ and TGF-β as state markers of schizophrenia (lower level of evidence for IL-4 and IL-10)Overlap between altered cytokine profiles in schizophrenia and bipolar disorder | [ref. 30,ref. 31,ref. 32,ref. 33,ref. 34] |
| Chemokines | Correlation between ↑ pro-inflammatory chemokines (e.g., CCL2, CCL4, CCL11, CCL17, CCL22, and CCL24) and schizophrenic symptomsRelationship between exposure to IL-8/CXCL8 and impaired neurodevelopmentMCP-1 (CCL2) as a trait marker of schizophrenia | [ref. 33,ref. 35,ref. 36,ref. 37] |
| Endocannabinoids | ↑ Anandamide in CSF and blood↑ Expression CB1R in peripheral immune cellsNegative correlation between positive and negative symptoms and anandamide levels in CSFPositive correlation between positive and negative symptoms of CB1R and CB2R in mononuclear cells | [ref. 38,ref. 39,ref. 40,ref. 41] |
| Eicosanoids | ↑ Baseline PLA2 activity in FEP and drug-free schizophrenic patientsPositive correlation between PLA2 activity and severity of schizophrenic symptoms↑ PGE2 and TxB2 in patients with criteria for ultra-high risk of psychosis | [ref. 42,ref. 43,ref. 44,ref. 45] |
| Lysophospholipids | Downward regulation of the LPA1R gene↓ LPA 22:6 in CSF Negative correlation between plasma levels of LPA and score on the PANSS scale↑ Lysophosphatidylcholines years before the onset of the disease | [ref. 46,ref. 47,ref. 48,ref. 49] |
| Bacterial products | Positive correlation between ↑ anti-LPS antibody and IL-6 Overexpression of genes in patients with first episode of psychosis involved in the response to LPS↑ Increased IgA/IgM responses to LPS in FEP and deficit schizophrenia and association between them and general cognitive impairment | [ref. 31,ref. 50,ref. 51,ref. 52] |
CSF: cerebrospinal fluid; FEP: first episode of psychosis; PANSS: positive and negative syndrome scale. ↑: Elevation of the biomarker. ↓: Descent of the biomarker.
3.1.1. Cytokines
Cytokines are key signaling molecules of the immune system that exert their effects on the periphery and in the brain. They are produced by immune and non-immune cells and exert their effects by binding to their specific receptors present on a variety of target cells [ref. 90].
Cytokines (and cytokine receptors or antagonists) are key regulators of inflammation, which includes the activation and recruitment of immune cells and increases blood flow and vascular permeability. They coordinate both the natural and adaptive type of the immune system. They can be classified in several ways, the most used method being through their biological functions [ref. 91]. We can distinguish pro-inflammatory, anti-inflammatory and hematopoietic cytokines. The first group would include molecules such as IL-1, IL-6 and TNF; the second, IL-4, IL-10 and IL-13; and the third, IL-3 and IL-5.
It has been shown that in individuals with a first episode of psychosis there is an imbalance in favor of cytokines and other pro-inflammatory mediators compared to other anti-inflammatory mediators, suggesting that immune system abnormalities could be an endophenotype of the disease [ref. 30]. These findings suggest the possibility that cytokines (and cytokine receptor) and other inflammatory mediators may be a biomarker of disease recurrence and/or response to adjuvant anti-inflammatory therapy for a subset of patients. The interest in the question has led to attempts to replicate these findings, with disparate results, in individuals in a clinical phase prior to the first episode of psychosis itself—the so-called high clinical risk of psychosis—so that an adequate early identification could be associated with an early approach and better results [ref. 31,ref. 32].
What is the neurobiological process underlying the inflammatory insult that we can observe through the presence of pro-inflammatory cytokines, and that can trigger psychosis? On the one hand, an overactivation of astrocytes and microglia has been described, as well as presynaptic stimulation of dopaminergic receptors in the midbrain [ref. 92]. In addition, these pro-inflammatory cytokines affect the regulation of the kynurenine pathway and alter glutamatergic transmission [ref. 93].
Given the accumulation of evidence that the alteration in the cytokine count correlates with the existence of psychotic disorders and with their own symptoms, the question that researchers are asking is whether a laboratory analysis could serve as a reliable biomarker for the diagnosis of these diseases [ref. 94]. In a recent review, Dawidowski et al. [ref. 33] draw the following conclusions in this regard:
- The elevation of the pro-inflammatory cytokine IL-6 is a finding that, with greater consistency between the different studies, could be considered a trait marker in schizophrenia; that is, one that is associated with hereditary and neurodevelopmental factors and, therefore, of susceptibility to the disease, remaining more or less stable during the different phases of it. Other potential trait markers would be elevations of the pro-inflammatory cytokines IL-1β and TNF-α, but there is insufficient evidence of their rise in high- or ultra-high-risk states of psychosis.
- Elevation of the pro-inflammatory cytokines IFN-γ and TGF-β could be considered state markers of schizophrenia, i.e., those that are associated with the disease itself and its symptoms, and may vary depending on the phase of the disease or antipsychotic treatment. Other potential state markers are IL-4 and IL-10, although in this case, only in chronic schizophrenia and not in the first episodes of psychosis.
Alterations in cytokine levels have also been studied for other psychotic spectrum disorders that share clinical features with schizophrenia, such as bipolar disorder [ref. 95,ref. 96]. An interesting fact is that an overlap of elevated levels of the same peripheral cytokines has been observed in patients with both the first episode of schizophrenia and bipolar disorder with psychotic characteristics [ref. 34].
3.1.2. Chemokines
Chemokines are small chemoattractant proteins (8–14 kDa) that have a role in mobilizing white blood cells to the site of inflammation at the peripheral level. In turn, they are involved in the development, maturation, survival and regeneration of nerve cells (glia and neurons) in the CNS. Chemokines have been classified into four subfamilies based on the number of N-terminal cysteine present: CXC, CC, XC and CX3C. However, functionally, the classic distinction between homeostatic and inflammatory chemokines is not as clear and many of the chemokines show a dual profile depending on the cell type and/or the inflammatory process triggered [ref. 97].
Chemokines play an important role in our biological systems and have been implicated in various diseases, including cancer, HIV/AIDS and atherosclerosis [ref. 98]. In general, chemokines trigger their functional activities by binding to G-protein-coupled receptors on the cell surface.
Thus, chemokines and their receptors play an important role in the nervous system, acting as trophic and protective factors that increase neuronal survival, regulate neuronal migration and synaptic transmission [ref. 35]. They are constantly secreted and are responsible for proper cell migration, for example, during the growth of the body. In the brain, the level of chemokines increases due to their secretion by many different cells: microglia, astrocytes, oligodendrocytes, and endothelial cells from blood vessels [ref. 99]. The blood–brain barrier plays a particular role, and its most important function is the precise exchange of chemical compounds between the CNS and the circulatory system. The integrity of the structure of the blood–brain barrier supports brain homeostasis and allows many neurological functions to be performed. Chemokines play a special role mainly in some CNS diseases, when damage to the blood–brain barrier and the blood–spine/brain fluid barrier causes leukocyte infiltration, triggering inflammatory processes [ref. 100].
Recently, chemokines have sparked the interest of numerous researchers regarding the role they may play in the genesis, development and natural history of psychotic disorders, especially schizophrenia. However, despite being considered a special type of cytokines, chemokines have so far not received the same attention as their older sisters, with the possible exception of IL-8 (CXCL8) [ref. 101,ref. 102,ref. 103,ref. 104]. However, accumulated evidence suggests that an increase in the serum level of pro-inflammatory chemokines (e.g., CCL2, CCL4, CCL11, CCL17, CCL22 and CCL24) strongly correlates with schizophrenic symptoms, including deficits in attention, working memory, episodic and semantic memory, and executive functions [ref. 35]. It has also been observed that fetal exposure to IL-8/CXCL8 may alter the early stages of neurodevelopment [ref. 36,ref. 37].
In the review we have discussed above, Dawidowski et al. [ref. 33] identify MCP-1 (CCL2) as another potential trait marker in schizophrenia, although there is a lack of solid data to support it in the population of first episode drug-naïve patients [ref. 105]. MCP-1 and IL-8 have also been linked to other psychiatric illnesses such as major depression or bipolar disorder [ref. 106,ref. 107,ref. 108], as well as postpartum psychosis [ref. 109].
3.1.3. Endocannabinoids
The endocannabinoid system is defined as the set of two G protein-coupled receptors and seven transmembrane domains for Δ-9-tetrahydrocannabinol (THC): cannabinoid receptor type 1 (CB1R) and cannabinoid receptor type 2 (CB2R) [ref. 110]. CB1R is the prominent subtype in the central nervous system (CNS) and has attracted great attention as a possible therapeutic pathway in several pathological conditions, including neuropsychological disorders and neurodegenerative diseases. In addition, cannabinoids also modulate signal transduction pathways and exert profound effects on peripheral sites [ref. 111].
In 1992, the first endogenous ligand of the cannabinoid receptor type 1, a derivative of arachidonic acid that was called “arachidonylethanolamide” or “anandaminde”, was discovered. The fact that anandamide (AEA) cannot fully reproduce the effects induced by THC led to the discovery of another important endocannabinoid, 2-arachidonoylglycerol (2-AG) [ref. 112]. Endocannabinoids are prominently involved in suppressing synaptic transmission through multiple mechanisms, with microglial cells and astrocytes being shown to be capable of producing their own 2-AG or AEA [ref. 113].
Endocannabinoid-like compounds comprise several molecules that are structurally related to endocannabinoids although they do not bind to the same receptors. These endocannabinoid-like compounds include N-acylethanolamines and 2-monoacylglycerols. Their structural resemblance to the endocannabinoids makes them players in the endocannabinoid system, where they can interfere with the actions of the true endocannabinoids, because they, in several cases, engage the same synthesizing and degrading enzymes [ref. 114]. Although we could speak of an “endocannabinoidome”, for the purposes of review, we will simplify and include these compounds in the endocannabinoid system.
Since the endogenous cannabinoid system has been involved in different neuropsychiatric disorders, Minichino et al. [ref. 38] have wondered, in a recent review, whether such a system could be abnormal in people with psychosis. The main findings were increased levels of anandamide in CSF and blood and increased expression of CB1R in peripheral immune cells of people with psychotic disease, compared to healthy controls. In addition, the studies reviewed report that the severity of positive and negative symptoms was associated with a decrease in anandamide levels in the CSF [ref. 39,ref. 40], and with increased expression of CB1 and CB2 receptors in mononuclear cells in peripheral blood [ref. 41].
In summary, a higher endocannabinoid system tone is evidenced at an early stage of the disease and in individuals free of antipsychotics (naïve), as well as an inverse association with the severity of symptoms. Such an increase in tone was normalized after successful treatment. Since mechanically, the release of anandamide in the brain can provide retrograde inhibition of the mesolimbic hyperdopaminergic state [ref. 115] (and therefore a reduction of positive symptoms), a feedback mechanism has been proposed in which the CNS of a subject with psychosis “brakes” it through its endocannabinoid system, which would end up acting as a natural protector against the disease. Thus, successful treatments would decrease this need for negative feedback [ref. 116,ref. 117].
3.1.4. Eicosanoids
Eicosanoids are oxidized derivatives of 20-carbon polyunsaturated fatty acids (PUFAs) formed by the cyclooxygenase (COX), lipoxygenase (LOX) and cytochrome P450 (cytP450) pathways. Arachidonic acid (ARA) is the usual substrate for eicosanoid synthesis. The COX pathways form prostaglandins (PGs) and thromboxanes (TXs), the LOX pathways form leukotrienes (LTs) and lipoxins (LXs), and the cytP450 pathways form various epoxy, hydroxy and dihydroxy derivatives [ref. 118].
These substances are highly bioactive, acting on many cell types through the G-protein-coupled receptors of the cell membrane, or as nuclear receptor ligands; and have been implicated in the pathophysiology of various diseases, such as cardiovascular disorders [ref. 119], gastrointestinal disorders, [ref. 120], cancer [ref. 121], Alzheimer’s disease [ref. 122] or even COVID-19 [ref. 123]. Many eicosanoids are known to have multiple effects, sometimes pleiotropic, on inflammation and immunity [ref. 124], and they control important cellular processes, including cell proliferation, apoptosis, metabolism and migration [ref. 125].
Eicosanoid biosynthesis is usually initiated by the activation of membrane phospholipase A2 (PLA2) in mammalian cells, which releases arachidonic acid, the main substrate of this process [ref. 126]. It has been observed that the activation of PLA2 modulates various neurotransmission systems that are involved in the pathophysiology of schizophrenia, such as the dopaminergic, serotonergic or glutamatergic systems [ref. 127]. A study by Smesny et al. [ref. 42] found an association between increased baseline PLA2 activity and a first episode of psychosis, as well as severity of schizophrenic symptoms. Previously, an increased cytoplasmic PLA2 activity in the serum of drug-free schizophrenic patients had already been reported in several studies [ref. 43,ref. 44,ref. 128].
A holistic explanation of the role that eicosanoid metabolism plays in the pathophysiology of psychosis would include increased activity of enzymatic pathways leading from lipid precursors in cell membranes to the synthesis of inflammatory products such as thromboxane A2 (TxA2), thromboxane B2 (TxB2) and prostaglandin E2 (PGE2) [ref. 129]. In a recent study, Pereira et al. [ref. 45] found an association between the elevation of the latter two and the existence of criteria for ultra-high risk of psychosis. These pathways could be slowed down by both antipsychotic drugs and drugs with direct anti-inflammatory action [ref. 129].
3.1.5. Lysophospholipids
The second product of the enzymatic action of PLA2 are lysophospholipids which, in turn, are converted into platelet-activating factors. These lipid mediators play critical roles in the initiation, maintenance and modulation of neuroinflammation and oxidative stress [ref. 130]. The two main categories of lysophospholipids, characterized according to their backbone, are lysoglycerophospholipids and lysosphingolipids. In cells, these lysophospholipids are intermediate precursors for the biosynthesis of other lipids in cells. Their intracellular concentrations are lower than those found in extracellular fluids [ref. 131].
In recent years, some of these lysophospholipids were identified as signaling molecules [ref. 132]. To date, in vivo and in vitro studies have indicated that signaling by lysophosphatidic acid (LPA) and sphingosine 1-phosphate (S1P) (two of the best-studied lysophospholipids) may play multiple roles in the relevance of CNS disorders, based on model systems analyses for psychiatric disorders including schizophrenia, anxiety, memory impairment, and neurological disorders such as Alzheimer’s disease (AD) [ref. 133]. Thus, for example, the lysophosphatidic acid subtype LPA1 has been involved with neurochemical and biochemical changes and other schizophrenia-related alterations in null mutant rodents [ref. 134,ref. 135,ref. 136,ref. 137] and the downward regulation of the LPA1R gene in patients has been pointed out [ref. 46].
Autotaxin (ATX) is a secreted enzyme that produces LPA from lysophosphatidylcholine (LPC), a type of lysoglycerolphospholipid. A recent study in Japanese patients has revealed significantly lower levels of one of the species of LPA, LPA 22:6 (LPA—docosahexaenoic acid), in CSF of patients with schizophrenia and major depressive disorder compared to healthy controls [ref. 47]. In addition, it appears that there may be a negative correlation between plasma levels of LPA and score on the PANSS scale, which could be useful for symptomatic assessment in schizophrenia [ref. 48]. For their part, Madrid-Gambín et al. identified early markers of psychosis through a longitudinal study in which they analyzed the blood of 115 children (12 years of age) who were identified for the first time as patients with psychotic experiences at 18 years of age, evidencing an increase in the levels of 4 lysophosphatidylcholines [ref. 49].
All this informs us of an alteration in the metabolism of the LPC-ATX-LPA pathway that may already be present in stages much earlier than the clinical development of the disease, and that would play an important role in its genesis and subsequent development. However, the results are still very preliminary, and further study of these biochemical species is needed.
3.1.6. Bacterial Products
For decades, the relationship that can exist between different infectious agents and the appearance of schizophrenia or schizophrenic spectrum disorders has been studied. For example, this relationship has been revealed in multiple ecological studies that relate the birth of subjects who will later develop psychosis in adulthood with influenza epidemics [ref. 138,ref. 139,ref. 140,ref. 141,ref. 142,ref. 143]. This relationship has also been studied for depressive disorder [ref. 144].
Sorensen et al. crossed data from the Copenhagen Perinatal Cohort and the Danish National Psychiatric Registry to test the hypothesis that a bacterial infection during pregnancy would confer an increased risk to offspring of developing schizophrenia in adulthood [ref. 145]. Exposure in the first trimester conferred an elevated risk of schizophrenia, which decreased from the second trimester. A relationship between the risk of schizophrenia and intrauterine exposure to rubella [ref. 146] and toxoplasma [ref. 147] has also been reported.
In recent years, these studies have shifted from eco-designs, which determine epidemic-based infection in populations, to research that has focused on reliable biomarkers in individual pregnancies [ref. 148]. On the other hand, there is a growing body of research that consistently involves the gut microbiome and the axis it forms with the CNS in the pathophysiology of various mental illnesses, including psychotic spectrum disorders [ref. 149].
Associations have been found between various bacterial products and psychosis. Lipopolysaccharides (LPS) are endotoxins found in the outer leaflet of the outer membrane of most Gram-negative bacteria [ref. 150]. Anti-LPS antibodies may serve as an indirect measure of LPS leakage into the circulation, which in turn would reflect a defect in intestinal barrier function [ref. 151]. Although, in a recent study, Delaney et al. [ref. 31] failed to find statistically significant differences in anti-LPS antibody levels between children, adolescents and young adults with psychosis, a population at ultra-high risk of psychosis, and healthy controls, they did find—for the psychosis group—a significant correlation between the increase in these antibodies and another well-established biomarker of psychosis, IL-6. This correlation could be due to the biological action produced by the binding of endotoxin to toll-like receptor 4 (TLR4), releasing the pro-inflammatory cytokine [ref. 152].
Maes et al. [ref. 50] have identified an overexpression of genes in patients with a first episode of psychosis involved in the response to LPS. These findings are in line with other studies reporting increased IgA/IgM responses to LPS from Gram-negative bacteria in this group of patients and deficit schizophrenia, and a significant association between these bacterial load indicators and general cognitive impairment and psychosis [ref. 51,ref. 52].
3.2. Inflammation and Drug Abuse
In Table 4 we have summarized the data presented on immuno-inflammatory markers in substance abuse disorders.
Table 4: Immuno-inflammatory markers in substance use disorders.
| Type of Biomarker | Main Findings | References | |
|---|---|---|---|
| Inflammatory proteins | Cannabis | ↑ IL-1ß, IL-8, TGF-ß, CCL11↓ IL-2Conflicts results for IL-6, TNFα | [ref. 53,ref. 54,ref. 55,ref. 56,ref. 57] |
| Cocaine | ↓ TNF-α, TGF-α, IL-1β, CXCL12, CCL2, CCL3 and CX3CL1 in substance-dependent humans during periods of withdrawal (but some of them elevated during acute cocaine exposure in mice)↓ TGF-α as a potential biomarker of dual diagnosisPositive correlation between plasma levels of IL-1β, CXCL12 and CX3CL1, and criteria of abuse and dependence↓ IL-10 in active users↓ IL-17 in abstinence usersConflicts results for IL-6 | [ref. 58,ref. 59,ref. 60,ref. 61,ref. 62,ref. 63] | |
| Alcohol | Dysregulation of TNFα, IL-1α, IL-1ß, IL-6, IL-8, IL-12 and MCP-1↑IL-1ß, IL-6 and TNFα in abstinent users↓IL- 4, IL-17A and IFN- γ in abstinent usersTransition from Th1 to Th2 cytokinesCorrelation in changes in IL-6 and IL-17A with liver and pancreatic diseasesStress-related suppression of peripheral cytokines may predict future relapse in alcohol-dependent individuals | [ref. 64,ref. 65,ref. 66,ref. 67,ref. 68] | |
| MA | ↑ TNF α, IL-1ß, IL-6 and CCL2 | [ref. 69,ref. 70,ref. 71,ref. 72] | |
| Tobacco | Dysregulation in production of pro/anti-inflammatory cytokines | [ref. 73,ref. 74] | |
| Inflammatory lipids | Cannabis | Altered levels of anandamide, 2-AG, PEA and OEA↓ Density CB1R in brain | [ref. 75,ref. 76,ref. 77,ref. 78,ref. 79] |
| Cocaine | ↑ NAEs in abstinent cocaine addicts↓ 2-AG in abstinent cocaine addicts↓ Hair concentrations of OEA and PEA in DCU compared with RCU and controlsAltered plasma levels of PGI2, PDE2 and TXA2 | [ref. 80,ref. 81,ref. 84,ref. 85,ref. 86] | |
| Alcohol | ↑ Anandamide, 2-AG and NAEsAlterations in the production of prostaglandins, thromboxane and leukotrienes↓ Total LPA | [ref. 82,ref. 83,ref. 87,ref. 88,ref. 89] |
MA: methamphetamines; 2-AG: 2-acylglycerols; PEA: palmitoylethanolamide; OEA: oleoylethanolamide; NAEs: N-acyl-ethanolamines; DCU: dependent cocaine users; RCU: recreational cocaine users. ↑: Elevation of the biomarker. ↓: Descent of the biomarker.
3.2.1. Inflammatory Proteins and Drug Abuse
Cytokines and chemokines have emerged as a new study biomarker in substance use disorders, along with other immuno-inflammatory mediators [ref. 29,ref. 153]. This provides us a different framework for understanding the functional and behavioral changes that contribute to drug addiction [ref. 154].
Studies in consumer patients, as well as in animal models of addiction, also show an imbalance in favor of pro-inflammatory factors. On the one hand, it has been proposed that a pro-inflammatory state could alter the neuromodulation of circuits involved in the pathophysiology of addiction [ref. 154] and, on the other hand, that exposure to substances of abuse can modify the secretion of cytokines and chemokines [ref. 153].
The involvement of cytokines and chemokines in different substance use disorders related to psychosis has been reported. For example, serum/plasma and other biological samples elevations of IL-1ß [ref. 53], IL-8 [ref. 53], TGF-ß [ref. 54] and CCL11 [ref. 55], and a decrease in IL-2 levels [ref. 54], have been described for cannabis use. Conflicting results have been reported for IL-6 [ref. 53,ref. 56] and TNFα [ref. 53,ref. 56,ref. 57]. Other studies link lung inflammation to smoking marijuana use [ref. 155,ref. 156] and gene expression analyses performed on the epithelium of the airways of cannabis users show an upward regulation of several toll-like receptors involved in the initiation of the inflammatory cascade [ref. 157].
However, the relationship of cannabinoids to inflammation remains controversial and complex. There are reports that take into account the anti-inflammatory properties of these compounds through multiple mechanisms related to the immune system [ref. 158,ref. 159,ref. 160]. Overall, the accumulated evidence suggests that cannabis extracts could modulate such a biological system [ref. 153].
Araos et al. [ref. 58] have found plasma levels of TNF-α, IL-1β, CXCL12, CCL2 and CX3CL1 to be elevated during acute cocaine exposure in mice, but reduced in substance-dependent humans during periods of withdrawal. In addition, plasma levels of IL-1β, CXCL12 and CX3CL1 were positively associated with criteria of abuse and dependence.
Two clinical studies in cocaine users showed a decrease in IL-10 (anti-inflammatory) and IL-17 (pro-inflammatory) at the peripheral level in active and abstinence users, respectively [ref. 59,ref. 60]. Again, contradictory results have been reported for IL-6 [ref. 59,ref. 61,ref. 62,ref. 63]. A peripheral decrease in TNF-α, CCL2, CCL3 and TGF-α has also been reported in abstinent cocaine users, and a decrease in TGF-α levels has been proposed as a potential biomarker of dual diagnosis [ref. 60,ref. 62]. All these integrated findings would suggest an unbalanced immune system towards the pro-inflammatory pole during active consumption that would be inhibited in periods of withdrawal.
One chemokine receptor system involved in the pathophysiology of cocaine addiction is CCR5 [ref. 161], whose ligands are CCL3 and CCL5. It has also been noted that dysregulation of cytokine and chemokine signaling during cocaine exposure would perpetuate cocaine use disorder [ref. 162,ref. 163,ref. 164] and would play a prominent role in the psychiatric comorbidity present in chronic cocaine users [ref. 165].
Alcohol consumption has been linked to an altered neuroimmune response that includes the production and release of pro-inflammatory cytokines [ref. 166]. Cytokines and chemokines such as TNFα, IL-1α, IL-1ß, IL-6, IL-8, IL-12 and MCP-1, can be regulated by alcohol and alcohol-induced tissue damage [ref. 64]. A recent meta-analysis showed consistent evidence of IL-6 elevation in subjects diagnosed with alcohol use disorder compared to healthy subjects [ref. 65]. Recent studies further suggest that the transition from Th1 to Th2 cytokines contributes to liver fibrosis and cirrhosis, so ethanol disruption of cytokines and inflammation may contribute in multiple ways to a diversity of alcoholic pathologies [ref. 66].
García-Marchena et al. [ref. 67] found that plasma concentrations of IL-1ß, IL-6 and TNFα were increased, whereas plasma concentrations of IL-4, IL-17A and IFN-γ were decreased, in abstinent alcohol use disorder patients as compared with control subjects. Moreover, they found that changes in IL-6 and IL-17A plasma concentrations in alcohol use disorder patients were associated with the presence of liver and pancreatic diseases. It has also been reported that stress-related suppression of peripheral cytokines may predict future relapse in alcohol-dependent individuals [ref. 68].
Methamphetamine-mediated neuroinflammation has also been the subject of recent study, with stimulation in the secretion of cytokines and chemokines such as TNFα, IL-1ß, IL-6 and CCL2 having been described [ref. 69,ref. 70,ref. 71,ref. 72]. Tobacco smoking has been implicated in the production of many immune or inflammatory mediators, including both pro-inflammatory and anti-inflammatory cytokines [ref. 73]. However, the effects of tobacco on inflammation are complex, since it could produce, at the same time, a pro-inflammatory effect in certain places and anti-inflammatory in others, generating a transition from immune response Th1 to Th2 [ref. 74]. The use of other substances generally less involved in the pathophysiology of psychosis, such as opioids, has also been linked to immuno-inflammatory changes [ref. 167].
3.2.2. Inflammatory Lipids and Drug Abuse
Small molecular weight lipids function as neuromodulators in the brain and, as such, play a role in the synaptic plasticity that occurs after exposure to drugs of abuse [ref. 168]. Recently, the important role played by the endocannabinoid system (ECS) in regulating the reinforcing and motivational properties of these substances has been described [ref. 169].
Exogenous administration of THC has been shown, for example, to influence endogenous cannabinoids circulating in humans, with altered levels of anandamide, 2-AG, palmitoylethanolamide and oleoylethanolamide [ref. 75]. In a recent study conducted on post-mortem brain tissue samples, Manza et al. [ref. 76] have reported that those subjects in whom physiologically restricted endogenous cannabinoid signaling may exist may be more vulnerable to the effects of chronic cannabis use on cortical thickness. Neuroimaging studies also show a decrease in the density of CB1 receptors in patients who use cannabis [ref. 77,ref. 78,ref. 79]. Genetic links to different critical elements of the ECS have also been proposed as a risk factor for cannabis dependence [ref. 169].
Two major human clinical studies analyzed concentrations of various substances in the endocannabinoid system in cocaine-using patients. Pavón et al. [ref. 80] carried out a study in which they characterized both free N-acyl-ethanolamines (NAEs) and 2-acyl-glycerols in abstinent cocaine addicts from outpatient treatment programs who were diagnosed with cocaine use disorder, and healthy control volunteers. While NAEs were found to increase, 2-acyl-glyceroles decreased in subjects with CUD compared to controls. Voegel et al. [ref. 81] compared hair concentrations of 2-arachidonylglycerol, anandamide (AEA), oleoylethanolamide (OEA) and palmitoylethanolamide (PEA) between recreational cocaine users (RCU), dependent cocaine users (DCU) and stimulant-naïve controls. Hair concentrations of OEA and PEA were significantly lower in DCU compared with RCU and controls. On the other hand, the interactions between the cannabinoid system and alcohol are well established. An increase in anandamide and 2-AG production has been reported in response to ethanol consumption [ref. 82]. García-Marchena et al. [ref. 83] have found that acylethanolamides were significantly increased in alcohol-dependent patients compared with control subjects.
Prostaglandin receptors are found in the cerebral cortex, hippocampus and midbrain [ref. 170]. The restoration of drug abuse is mainly mediated by the synergistic effect of the cannabinoid system and the products of arachidonic acid [ref. 171]. From this point of view, prostaglandins have an important role in drug addiction [ref. 172]. The influence of cocaine on the endothelial or plasma levels of PGI2, PDE2 or TXA2 has been reported [ref. 84,ref. 85,ref. 86]. In a review, Orio et al. [ref. 173] propose that, however, it is unknown whether the peripheral effects of cocaine on PG levels may influence central structures related to the addictive process. They relate, in different ways, the eicosanoids metabolism with cocaine addiction. Alterations in the production of prostaglandins, thromboxane and leukotrienes have also been reported in relation to alcohol consumption [ref. 87,ref. 88]. Some of these molecules have been linked to physical opioid dependence [ref. 174].
Because of its role in the control of both neural plasticity events and neurogenesis and its interrelation with the endocannabinoid system, LPA and its receptors might be involved in drug addiction-associated neuroadaptations [ref. 173]. Chronic cocaine administration induced conditioned locomotion deficits in LPA-1-receptor-null mice [ref. 175]. García-Marchena et al. have found a decrease in total LPA concentrations in patients with alcohol use disorder and healthy controls [ref. 89].
4. Discussion
Our review provides a panoramic—though not exhaustive—view of the main neuroinflammatory mechanisms that have been linked to psychotic disorders (especially schizophrenia) and substance use disorders. Although some prospective cohorts have been studied to analyze the risk of schizophrenia in adulthood of subjects who may have been exposed to toxic agents during development, most findings come from retrospective and observational studies. This should make us cautious with conclusions. However, some preclinical or in vitro data seem to support the findings in human participants [ref. 70,ref. 75,ref. 85,ref. 86]. Furthermore, data about inflammatory biomarkers in patients with dual diagnosis (psychosis and addiction) are reported in scientific literature and TGFα has been proposed as a potential biomarker of this double condition [ref. 60,ref. 62,ref. 80].
4.1. Drug Abuse and Psychosis: An Inflammatory Convergent Origin?
In this paper, we have reviewed, from a narrative approach, how both psychosis and drug abuse have been related to immuno-inflammatory changes and mechanisms. These would include a series of cellular and molecular systems in which an imbalance in favor of pro-inflammatory factors has been described, and there would also be an overlap between many signals found in both disorders. Considering that the reinforcing effects of drugs depend mainly on dopamine signaling in the nucleus accumbens [ref. 176], the main neuroanatomical substrate of many cardinal symptoms of psychosis, this overlap should not surprise us, and should make us wonder if both conditions could not ultimately have a common or convergent origin.
This new “inflammatory hypothesis” of mental disorders would not replace, but would complement, the classical hypotheses based on alterations in the metabolism and signaling of brain neurotransmitters. Many immune molecules interact with neurotransmitter systems and play an essential role in modulating synaptic function [ref. 154]. For example, TNFα modulates the traffic of AMPA-type glutamate receptors and GABA receptors [ref. 177,ref. 178,ref. 179], IL-1β modulates long-term potentiation [ref. 180,ref. 181], CCL2 and CXCL-12 regulate the release of glutamate, GABA and dopamine [ref. 182,ref. 183]. We further know that many of these neuroimmune molecules are also involved in all stages of neurodevelopment, play a key role in neurogenesis and gliogenesis, and interfere with neuronal migration and neurocircuit formation [ref. 184,ref. 185,ref. 186].
The role that chronic stress can have in psychosis spectrum disorders has long been discussed in the scientific literature [ref. 187]. In a recent review, Vargas et al. [ref. 188] list several converging mechanisms that would be involved in the development of psychosis. These would include hypothalamic–pituitary–adrenal axis activation, dopamine and glutamate dysregulation… and chronic inflammation. Therefore, peripheral inflammation resulting from stress exposure could lead to neuroinflammatory responses impacting cortico-amygdala function, as well as prefrontal cortex structure, function and development [ref. 189].
Considering all these findings, Millan et al. [ref. 190] have recently described an anomalous neurodevelopmental model for schizophrenia that we find especially interesting. Thus, in a developing individual, a first wave of biological and psychological insults would act (which would include factors such as prenatal infection, childhood trauma, malnutrition, etc.). Later, in a second wave, in the period comprising adolescence and early youth, drug use or social isolation would appear. All this would configure an abnormally developed brain and CNS that would make psychosis hatch in its positive dimension once the damage is produced.
In all this, it is worth asking: why do patients with psychosis have such a high rate of substance abuse? In general, the association between a substance use disorder and another mental illness is known as “dual diagnosis” [ref. 191]. Different etiological hypotheses have been proposed to explain this phenomenon [ref. 192]: that addiction is a consequence of primary mental illness (as a mechanism, for example, of “self-medication” in the face of psychological distress that these subjects would experience), that mental illness is a direct consequence of the addictive substance (as in induced psychoses) or, finally, that both disorders are independent and share common etiological factors. Within these, neuroinflammation could have a prominent role.
The search for that common inflammatory origin for psychosis and substance use disorders has led us to explore a neuroinflammatory and neurotoxic pathway that has its onset in the activation of the transcriptional factor NFκB. This is a ubiquitous transcription factor well known for its role in the innate immune response whose involvement in drug abuse has been the subject of a recent review [ref. 193]. Under normal conditions, NFκB is bound in the cytosol to its inhibitory complex, IκB. This prevents it from being transferred to the core and activating a pro-inflammatory cascade. However, certain stimuli, such as the activation of cytokine receptors or toll-like receptors (associated with bacterial infections), can cause the activation of an intracellular signaling pathway that leads to the phosphorylation of IκB, which would decouple from NFκB and allow the translocation of the latter to the cell nucleus [ref. 194]. There, they bind to specific DNA sequences in the promoter region of a wide range of genes encoding a number of inflammatory mediators [ref. 195,ref. 196]. All of this would generate a positive feedback loop [ref. 193].
García-Bueno et al. [ref. 30] have found increased NFκB activity in nuclear extracts of peripheral blood mononuclear cells (PBMC) from FEP patients versus healthy controls. They have proposed an integrative inflammatory hypothesis of schizophrenia that starts from the degradation of the inhibitory complex IκB and the activation by NFκB activity of pro-inflammatory enzymatic pathways through inducible nitric oxide synthase (iNOS) and isoform 2 of the enzyme cyclooxygenase-2 (COX-2). This would result in an accumulation of oxidative and nitrosative mediators, such as nitric oxide, peroxynitrite anion and PGE2, that would cause the depletion of endogenous antioxidant defenses and the attack on membrane phospholipids, causing cell damage through a lipid peroxidation process [ref. 197].
It has been shown, on the other hand, that NFκB activity can increase through diverse environmental stimuli that would include substances of abuse such as cocaine [ref. 198,ref. 199], opiates [ref. 200] or alcohol [ref. 201,ref. 202,ref. 203]. In addition, NFκB could play an important role in the functional regulation of mechanisms associated with the development of substance dependence. Thus, for example, in preclinical models it has been possible to reduce the sensitivity to the rewarding properties of cocaine by eliminating NFκB activity [ref. 199]. Finally, NFκB could influence drug seeking through a predominant biological role in the response to emotional stress or in physiological behavioral processes involved in memory formation [ref. 193].
NFκB would have an interesting counterpart that would act as a transcription factor dependent on ligands, and that would generate effects contrary to the receptor activated by the peroxisome proliferator, especially in its gamma isoform (PPARγ) [ref. 204]. Once PPARγ activated, it would translocate to the nucleus and modify the cell’s gene expression by inhibiting the inflammatory response. Interestingly, a known ligand of PPARγ is 15d-prostaglandin J2 (15d PGJ2), which, in turn, is a product of the COX pathway. Therefore, this pathway would be activated as a compensatory mechanism after the cascade initiated by NFκB, and the balance or imbalance of both would condition, respectively, the physiological or pathological result of neuroinflammatory processes. Recently, PPAR have been connected to the endocannabinoid system. These nuclear receptors, indeed, mediate the effects of anandamide and endocannabinoid-like substances such as OEA and PEA [ref. 205].
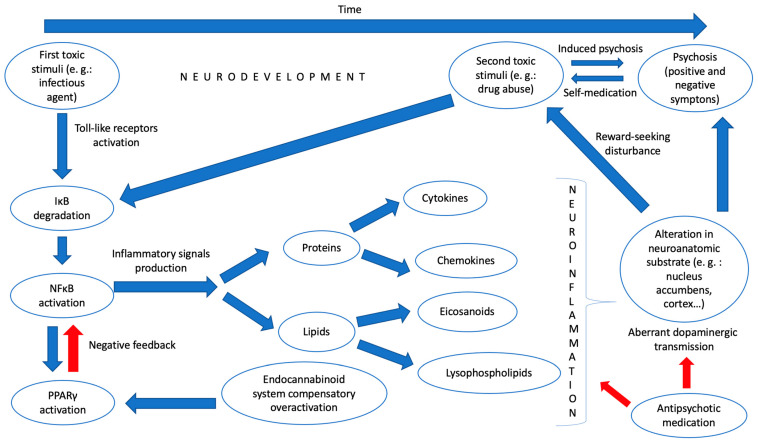
Given all these findings, we dare to hypothesize a model of neuroinflammation that can somehow link psychosis with substance abuse and that could be detected through various biological “signals” in blood and other tissue samples. Thus, in the first place, at a very early stage of development, a toxic environmental stimulus (for example, a bacterial infection) would provoke, through the stimulation of toll-like receptors and the participation of other inflammatory mediators, an activation of the master regulator of inflammation NFκB by degrading its inhibitory complex. This would activate the production of inflammatory mediators, among which would be lipids, such as eicosanoids or lysophospholipids, and proteins, such as cytokines and chemokines, which, in turn, would generate, by activating microglia, a “pro-inflammatory state” in the CNS of the young individual. It would trigger, concomitantly, and through the PPARγ pathway, an overactivation of natural anti-inflammatory systems, such as the endocannabinoid system, that, despite everything, could not counteract the previous effect. This pro-inflammatory state would influence the genesis of neurocircuits that would be pathologically formed, with alterations in critical pathways related to the search for pleasure or reward in various neuroanatomical substrates such as the nucleus accumbens. All this would generate a vulnerability to the development of psychosis or a substance use disorder. However, a second environmental insult is still necessary, in line with what was proposed by Millan et al., which would occur in adolescence with the effective consumption of a drug or another type of biological aggression to this vulnerable CNS. Finally, these pathological neurocircuits create an aberrant dopaminergic transmission in various parts of the brain, which would produce the known positive and negative symptoms of psychosis (and condition the development of dependence on the drug consumed).
In Figure 1, we have summarized our neuroinflammatory hypothesis about the common origin of psychosis and drug abuse.
4.2. Future Perspectives
If we look at the neurodevelopmental model of psychosis (which, as we have seen, we can also apply to substance use disorders to a large extent), we are faced with a bleak clinical picture: we are acting only at the end of the disease process, when brain damage is already established. In addition, we are acting not only at the end of the process at the chronological level, but at the tip of the iceberg of the biological processes underlying the etiopathogenesis of these disorders, which would be the alteration in the transmission of dopamine. This is the classic known mechanism of action of antipsychotic drugs. Notably, these drugs, with respect to which we have few reliable biomarkers to assess effectiveness or adherence (beyond the measurement of their plasma levels), are usually used indiscriminately and empirically without attending to the possible endophenotypes of disease that we may find. The new explanatory approaches to psychosis (whether primary, dual or toxic) should also help us to adopt novel clinical perspectives which change the way we understand its nature and, therefore, that suppose a change in the therapeutic approach to our patients.
A first question that could be addressed in light of these new findings is patient stratification. Recently, more and more authors dare to doubt that some diseases that we thought well categorized, such as schizophrenia, are not actually a construct whose validity is questionable [ref. 206]. Endophenotypes—measurable components not seen with the naked eye along the pathway between the disease and the distal genotype [ref. 207]—could serve as biological markers in schizophrenia. Some of these, related to microvascular abnormality [ref. 208] or impaired prostaglandin signaling [ref. 209], have been proposed. We propose, like other authors, to conceptualize psychosis as a “fever” of serious mental illnesses, a serious but non-specific indicator [ref. 210]. Having reliable biomarkers, such as inflammatory signals that we have analyzed in this review, can make us change the classic nomenclature still in force in international classifications and better distinguish some patients from others.
When we also introduce into this equation the variable associated with substance use, things get complicated. It is common that in many Western countries there is a separate network of care for mental health and drug addiction, which generates difficulties in the care of patients with dual diagnosis, and a deficit of professionals who are experts in treating both types of pathologies [ref. 211]. Despite the very high co-prevalence between substance use disorders and serious mental illnesses, clinical guidelines for both tend to have limited considerations for coexisting disorders in diagnosis, treatment and management, as evidenced by a recent review [ref. 212]. This, in our opinion, ends up generating a vicious circle in the circuits of attention to dual patients: they are continually referred from one care service to another without being treated properly. This is what has come to be called the “wrong door syndrome” [ref. 213].
In this review, we believe we have shown that psychosis and drug abuse are not only related from the epidemiological point of view, but also share a common vulnerability model and convergent etiopathogenic pathways. Therefore, their treatment must be integrated. Much importance has generally been granted to the distinction between “primary” and “induced” psychoses [ref. 214], despite the fact that a high rate of patients originally diagnosed with induced psychosis end up developing schizophrenia or bipolar disorder [ref. 215].
Another important perspective to consider in our review is response to treatment. There is still a high percentage of patients who are refractory to usual treatment, estimated at around 20–30% of the total [ref. 216]. However, if we consider as a therapeutic objective not only the remission of positive and/or negative symptoms but the functional recovery, then only one in seven patients will end up achieving it [ref. 217]. However, why do some patients respond well (or acceptably well) to conventional antipsychotics and others do not? A recent systematic review looking at multiple patient characteristics found that only a higher positive symptom score at baseline predicted a higher response rate [ref. 218]. We can say that, in a certain sense, we are quite blind, both when we start an antipsychotic treatment in a patient with psychosis and when we predict the response to second-line treatments.
However, something may be changing in recent times. There is already talk of a “precision” or “personalized” psychiatry that would consist of considering mental illness as a dysregulation, unique to each patient, of the molecular pathways that govern the development and functioning of the brain [ref. 219]. Within this approach, we think that an adequate phenotypic characterization of the inflammatory mechanisms involved in the genesis of psychosis can have important prognostic implications and, to some extent, predict the response to treatment.
In addition to predicting response to treatment, another important aspect of antipsychotic therapy is monitoring it. In our review, we have identified some immuno-inflammatory signals that could act as endogenous markers of response to psychotropic drugs. At least, we should consider the alterations in the concentrations of the pro-inflammatory cytokines IFN-γ and TGF-β as well as the variations we find in the lipid signaling of the molecules that are part of the endocannabinoid system, whose level of overactivation would also be mediated by the treatment. The final issue related to the treatment of people with psychosis that we want to discuss is the growing evidence of the effectiveness of some anti-inflammatory drugs, such as aspirin, estrogens, minocycline and NAC, for this condition [ref. 220].
Last but not least, we would like to point out some aspects that seem key to the prevention of psychosis. It may be the case that acting on adolescents in a clinical state of high risk for psychosis (which, until now, has been considered the priority objective of prevention programs) is already too late: a recently published review of Catalán et al. [ref. 221] concluded that there was insufficient evidence to recommend a specific treatment over others (including control conditions) to prevent the transition to psychosis. However, some interventions with potentially beneficial effects in patients at high risk for psychosis include omega-3 fatty acid supplementation (EPA and DHA), whose role in developmental psychopathology has been highlighted by Agostoni et al., and which appear to play a key role in resolving inflammation [ref. 222]. Furthermore, in this population, some neuroinflammatory signals could also act as early diagnostic or prognostic factors. Thus, a reliable trait marker that we could measure in this population and that seems to rise consistently during the prodromal phase of the disease is the cytokine IL-6. Eicosanoid products, such as thromboxane B2 (TxB2) and prostaglandin E2 (PGE2), could also be elevated at this phase. Finally, overactivation of the endocannabinoid system, or an alteration in the metabolism of the LPC-ATX-LPA pathway, may also be an early marker of disease, although further studies are required.
The fact that the care of adolescents in a high-risk mental state has not provided all the results we expected has led authors to propose that we move to a scenario of universal primary prevention [ref. 223]. Considering how brain inflammation, from the early stages of development, can condition the subsequent development of psychosis should lead us to improve pre- and perinatal care (especially in mothers with schizophrenia) [ref. 224,ref. 225], decrease prenatal infections and nutritional deficiencies [ref. 226], act in children at risk of psychosis by virtue of an increased familial or genetic risk [ref. 227], etc. Special mention deserves the action on drug abuse. Especially striking is the case of cannabis, having been calculated that, if high-potency cannabis were not available in Europe, 12 to 50% of the first episodes of psychosis could be prevented, depending on the geographical area [ref. 228]. We believe that, just as the fight against lung cancer has justified the proliferation of preventive campaigns to prevent nicotine abuse, a clear, forceful and clear response from the authorities cannot be delayed any longer: the use of cannabis considerably increases the risk of psychosis.
4.3. Limitations and Strengths
Of course, the review we have carried out has some limitations. First, the lack of a systematic search for results. However, it is known that sometimes narrative reviews like ours can offer some advantages and allow a more flexible and holistic approach to the question analyzed [ref. 229]. Second, we have not reviewed in-depth preclinical studies that could support the findings discussed in human participants. Finally, we must not forget an aspect that is not insignificant: to explore whether the association between inflammation and psychosis (and dual diagnosis) represents causality or rather a confounded epiphenomenon. Köhler-Forsberg et al. posed this question in a recent editorial [ref. 230]. In this sense, other mental disorders of high prevalence, such as major depression, have also been linked to neuroinflammatory mechanisms, including NFκB signaling [ref. 231]. However, the recurrent genetic overlap between different mental illnesses is already known, which should lead us to consider the sometimes-arbitrary limits between the diagnostic categories currently established in psychiatry [ref. 232]. Therefore, perhaps the common origin proposed in our review for psychosis and drug addiction is only a part of the convergent pathological pathways between numerous psychiatric disorders.
5. Conclusions
A significant number of neuroinflammatory signals that would include cytokines, chemokines, endocananbinoids, eicosanoids, lysophospholipids and bacterial products have been linked to psychotic disorders. Many of these markers also appear in substance use disorders, with an overlap between the two diagnostic categories.
Psychosis and substance use disorders, whose relationship has been studied for decades, may share a common origin. This would be an abnormal neurodevelopment where a first toxic stimulus would trigger an inflammatory imbalance in the body, with a production of pro-inflammatory lipids and proteins and an inability of endogenous anti-inflammatory systems, such as the endocannabinoid system, to counteract it. A possible convergent pathway is that which interrelates the transcriptional factors NFκB and PPARγ. Subsequently, a second toxic stimulus would precipitate the appearance of symptoms.
This finding may have implications for the prevention, diagnosis and treatment of our patients. However, we must be cautious with the interpretation of these findings, and more studies are needed in the future to delve into many of these issues.
References
- D.B. Arciniegas. Psychosis. Contin. Minneap. Minn., 2015. [DOI]
- W. Gaebel, J. Zielasek. Focus on Psychosis. Dialogues Clin. Neurosci., 2015. [DOI | PubMed]
- S.M. Stahl. Psicosis y Esquizofrenia. Psicofarmacología Esencial de Stahl, 2013
- D.H. Geschwind, J. Flint. Genetics and Genomics of Psychiatric Disease HHS Public Access. Science, 2015. [DOI | PubMed]
- H. Stefansson, D. Rujescu, S. Cichon, O.P.H. Pietiläinen, A. Ingason, S. Steinberg, R. Fossdal, E. Sigurdsson, T. Sigmundsson, J.E. Buizer-Voskamp. Large Recurrent Microdeletions Associated with Schizophrenia. Nature, 2008. [DOI | PubMed]
- M. Burmeister, M.G. McInnis, S. Zöllner. Heritability Psychiatric Genetics: Progress amid Controversy. Nat. Rev. Genet., 2008. [DOI | PubMed]
- J. Sanders, M. Gill. Unravelling the Genome: A Review of Molecular Genetic Research in Schizophrenia. Ir. J. Med. Sci., 2007. [DOI | PubMed]
- J. Read, J. van Os, A.P. Morrison, C.A. Ross. Childhood Trauma, Psychosis and Schizophrenia: A Literature Review with Theoretical and Clinical Implications. Acta Psychiatr. Scand., 2005. [DOI | PubMed]
- R. van Winkel, M. van Nierop, I. Myin-Germeys, J. van Os. Childhood Trauma as a Cause of Psychosis: Linking Genes, Psychology, and Biology. Can. J. Psychiatry, 2013. [DOI]
- A. Gururajan, E.E. Manning, M. Klug, M. van den Buuse. Drugs of Abuse and Increased Risk of Psychosis Development. Aust. N. Z. J. Psychiatry, 2012. [DOI | PubMed]
- A. Fiorentini, L.S. Volonteri, F. Dragogna, C. Rovera, M. Maffini, M.C. Mauri, C.A. Altamura. Substance-Induced Psychoses: A Critical Review of the Literature. Curr. Drug Abus. Rev., 2011. [DOI]
- A. Damjanoviü, M. Pantoviü, A. Damjanoviü, B. Dunjiü-Kostiü, M. Ivkoviü, S. Milovanoviü, M. Lackoviü, I. Dimitrijeviü. Cannabis and psychosis revisited. Psychiatr. Danub., 2015. [PubMed]
- C. Andrade. Cannabis and Neuropsychiatry, 2: The Longitudinal Risk of Psychosis as an Adverse Outcome. J. Clin. Psychiatry, 2016. [DOI | PubMed]
- M.J. Herrero, A. Domingo-Salvany, M. Torrens, M.T. Brugal. ITINERE Investigators Psychiatric Comorbidity in Young Cocaine Users: Induced versus Independent Disorders. Addict. Abingdon Engl., 2008. [DOI]
- J.J. Mahoney, A.D. Kalechstein, R. de La Garza, T.F. Newton. Presence and Persistence of Psychotic Symptoms in Cocaine- versus Methamphetamine-Dependent Participants. Am. J. Addict., 2008. [DOI | PubMed]
- C. Roncero, M. Comín, C. Daigre, L. Grau-López, N. Martínez-Luna, F.J. Eiroa-Orosa, C. Barral, M. Torrens, M. Casas. Clinical Differences between Cocaine-Induced Psychotic Disorder and Psychotic Symptoms in Cocaine-Dependent Patients. Psychiatry Res., 2014. [DOI | PubMed]
- E. Vergara-Moragues, J.I. Mestre-Pintó, P. Araos Gómez, F. Rodríguez-Fonseca, M. Torrens, F. González-Saiz. Can Symptoms Help in Differential Diagnosis between Substance-Induced vs. Independent Psychosis in Adults with a Lifetime Diagnosis of Cocaine Use Disorder?. Psychiatry Res., 2016. [DOI | PubMed]
- A.I. Green, R.E. Drake, M.F. Brunette, D.L. Noordsy. Treatment in Psychiatry Schizophrenia and Co-Occurring Substance Use Disorder. Am. J. Psychiatry, 2007. [DOI | PubMed]
- J.E. Zweben, J.B. Cohen, D. Christian, G.P. Galloway, M. Salinardi, D. Parent, M. Iguchi. Methamphetamine Treatment Project Psychiatric Symptoms in Methamphetamine Users. Am. J. Addict. Am. Acad. Psychiatr. Alcohol Addict., 2004. [DOI]
- L. Archibald, M.F. Brunette, D.J. Wallin, A.I. Green. Alcohol Use Disorder and Schizophrenia or Schizoaffective Disorder. Alcohol Res., 2019. [DOI]
- H. Quigley, J.H. MacCabe. The Relationship between Nicotine and Psychosis. Adv. Psychopharmacol., 2019. [DOI]
- M. Large, S. Sharma, M.T. Compton, T. Slade, O. Nielssen. Cannabis Use and Earlier Onset of Psychosis: A Systematic Meta-Analysis. Arch. Gen. Psychiatry, 2011. [DOI | PubMed]
- J. Suvisaari, O. Mantere. Inflammation Theories in Psychotic Disorders: A Critical Review. Infect. Disord. Drug Targets, 2013. [DOI | PubMed]
- R.S. Smith. A Comprehensive Macrophage-T-Lymphocyte Theory of Schizophrenia. Med. Hypotheses, 1992. [DOI | PubMed]
- R.S. Smith, M. Maes. The Macrophage-T-Lymphocyte Theory of Schizophrenia: Additional Evidence. Med. Hypotheses, 1995. [DOI | PubMed]
- M.J. Schwarz, N. Müller, M. Riedel, M. Ackenheil. The Th2-Hypothesis of Schizophrenia: A Strategy to Identify a Subgroup of Schizophrenia Caused by Immune Mechanisms. Med. Hypotheses, 2001. [DOI | PubMed]
- A. Monji, T. Kato, S. Kanba. Cytokines and Schizophrenia: Microglia Hypothesis of Schizophrenia. Psychiatry Clin. Neurosci., 2009. [DOI | PubMed]
- M.J. Lacagnina, P.D. Rivera, S.D. Bilbo. Glial and Neuroimmune Mechanisms as Critical Modulators of Drug Use and Abuse. Neuropsychopharmacology, 2017. [DOI | PubMed]
- O.C. Ahearn, M.N. Watson, S.M. Rawls. Chemokines, Cytokines and Substance Use Disorders. Drug Alcohol Depend., 2021. [DOI]
- B. García-Bueno, M. Bioque, K.S. Mac-Dowell, M.F. Barcones, M. Martínez-Cengotitabengoa, L. Pina-Camacho, R. Rodríguez-Jiménez, P.A. Sáiz, C. Castro, A. Lafuente. Pro-/Anti-Inflammatory Dysregulation in Patients with First Episode of Psychosis: Toward an Integrative Inflammatory Hypothesis of Schizophrenia. Schizophr. Bull., 2014. [DOI | PubMed]
- S. Delaney, B. Fallon, A. Alaedini, R. Yolken, A. Indart, T. Feng, Y. Wang, D. Javitt. Inflammatory Biomarkers in Psychosis and Clinical High Risk Populations. Schizophr. Res., 2019. [DOI | PubMed]
- S. Park, B.J. Miller. Meta-Analysis of Cytokine and C-Reactive Protein Levels in High-Risk Psychosis. Schizophr. Res., 2020. [DOI | PubMed]
- B. Dawidowski, A. Górniak, P. Podwalski, Z. Lebiecka, B. Misiak, J. Samochowiec. The Role of Cytokines in the Pathogenesis of Schizophrenia. J. Clin. Med., 2021. [DOI | PubMed]
- T.A. Lesh, M. Careaga, D.R. Rose, A.K. McAllister, J. van de Water, C.S. Carter, P. Ashwood. Cytokine Alterations in First-Episode Schizophrenia and Bipolar Disorder: Relationships to Brain Structure and Symptoms. J. Neuroinflammation, 2018. [DOI | PubMed]
- T. Boczek, J. Mackiewicz, M. Sobolczyk, J. Wawrzyniak, M. Lisek, B. Ferenc, F. Guo, L. Zylinska. The Role of g Protein-Coupled Receptors (Gpcrs) and Calcium Signaling in Schizophrenia. Focus on Gpcrs Activated by Neurotransmitters and Chemokines. Cells, 2021. [DOI | PubMed]
- L.M. Ellman, R.F. Deicken, S. Vinogradov, W.S. Kremen, J.H. Poole, D.M. Kern, W.Y. Tsai, C.A. Schaefer, A.S. Brown. Structural Brain Alterations in Schizophrenia Following Fetal Exposure to the Inflammatory Cytokine Interleukin-8. Schizophr. Res., 2010. [DOI | PubMed]
- A.S. Brown. Prenatal Infection as a Risk Factor for Schizophrenia. Schizophr. Bull., 2006. [DOI | PubMed]
- A. Minichino, M. Senior, N. Brondino, S.H. Zhang, B.R. Godwlewska, P.W.J. Burnet, A. Cipriani, B.R. Lennox. Measuring Disturbance of the Endocannabinoid System in Psychosis: A Systematic Review and Meta-Analysis. JAMA Psychiatry, 2019. [DOI | PubMed]
- F.M. Leweke, A. Giuffrida, D. Koethe, D. Schreiber, B.M. Nolden, L. Kranaster, M.A. Neatby, M. Schneider, C.W. Gerth, M. Hellmich. Anandamide Levels in Cerebrospinal Fluid of First-Episode Schizophrenic Patients: Impact of Cannabis Use. Schizophr. Res., 2007. [DOI | PubMed]
- A. Giuffrida, F.M. Leweke, C.W. Gerth, D. Schreiber, D. Koethe, J. Faulhaber, J. Klosterkötter, D. Piomelli. Cerebrospinal Anandamide Levels Are Elevated in Acute Schizophrenia and Are Inversely Correlated with Psychotic Symptoms. Neuropsychopharmacology, 2004. [DOI | PubMed]
- K.A. Chase, B. Feiner, C. Rosen, D.P. Gavin, R.P. Sharma. Characterization of Peripheral Cannabinoid Receptor Expression and Clinical Correlates in Schizophrenia. Psychiatry Res., 2016. [DOI | PubMed]
- S. Smesny, C. Kunstmann, S. Kunstmann, I. Willhardt, J. Lasch, R.A. Yotter, T.M. Proffitt, M. Kerr, C. Marculev, B. Milleit. Phospholipase A2 Activity in First Episode Schizophrenia: Associations with Symptom Severity and Outcome at Week 12. World J. Biol. Psychiatry, 2011. [DOI | PubMed]
- M. Noponen, M. Sanfilipo, K. Samanich, H. Ryer, G. Ko, B. Angrist, A. Wolkin, E. Duncan, J. Rotrosen. Elevated PLA2 Activity in Schizophrenics and Other Psychiatric Patients. Biol. Psychiatry, 1993. [DOI | PubMed]
- W.F. Gattaz, M. Köllisch, T. Thuren, J.A. Virtanen, P.K.J. Kinnunen. Increased Plasma Phospholipase-A2 Activity in Schizophrenic Patients: Reduction after Neuroleptic Therapy. Biol. Psychiatry, 1987. [DOI | PubMed]
- C.A.C. Pereira, A.C. Costa, H.P.G. Joaquim, L.L. Talib, M.T. van de Bilt, A.A. Loch, W.F. Gattaz. COX-2 Pathway Is Upregulated in Ultra-High Risk Individuals for Psychosis. World J. Biol. Psychiatry, 2021. [DOI | PubMed]
- N.A. Bowden, J. Weidenhofer, R.J. Scott, U. Schall, J. Todd, P.T. Michie, P.A. Tooney. Preliminary Investigation of Gene Expression Profiles in Peripheral Blood Lymphocytes in Schizophrenia. Schizophr. Res., 2006. [DOI | PubMed]
- W. Omori, K. Kano, K. Hattori, N. Kajitani, M. Okada-Tsuchioka, S. Boku, H. Kunugi, J. Aoki, M. Takebayashi. Reduced Cerebrospinal Fluid Levels of Lysophosphatidic Acid Docosahexaenoic Acid in Patients With Major Depressive Disorder and Schizophrenia. Int. J. Neuropsychopharmacol., 2021. [DOI | PubMed]
- L. Gotoh, M. Yamada, K. Hattori, D. Sasayama, T. Noda, S. Yoshida, H. Kunugi, M. Yamada. Levels of Lysophosphatidic Acid in Cerebrospinal Fluid and Plasma of Patients with Schizophrenia. Psychiatry Res., 2019. [DOI | PubMed]
- F. Madrid-Gambin, M. Föcking, S. Sabherwal, M. Heurich, J.A. English, A. O’Gorman, T. Suvitaival, L. Ahonen, M. Cannon, G. Lewis. Integrated Lipidomics and Proteomics Point to Early Blood-Based Changes in Childhood Preceding Later Development of Psychotic Experiences: Evidence From the Avon Longitudinal Study of Parents and Children. Biol. Psychiatry, 2019. [DOI | PubMed]
- M. Maes, K. Plaimas, A. Suratanee, C. Noto, B. Kanchanatawan. First Episode Psychosis and Schizophrenia Are Systemic Neuro-Immune Disorders Triggered by a Biotic Stimulus in Individuals with Reduced Immune Regulation and Neuroprotection. Cells, 2021. [DOI | PubMed]
- M. Maes, A. Vojdani, S. Sirivichayakul, D.S. Barbosa, B. Kanchanatawan. Inflammatory and Oxidative Pathways Are New Drug Targets in Multiple Episode Schizophrenia and Leaky Gut, Klebsiella Pneumoniae, and C1q Immune Complexes Are Additional Drug Targets in First Episode Schizophrenia. Mol. Neurobiol., 2021. [DOI | PubMed]
- M. Maes, B. Kanchanatawan, S. Sirivichayakul, A.F. Carvalho. In Schizophrenia, Increased Plasma IgM/IgA Responses to Gut Commensal Bacteria are Associated with Negative Symptoms, Neurocognitive Impairments, and the Deficit Phenotype. Neurotox. Res., 2019. [DOI | PubMed]
- H. Bayazit, S. Selek, I.F. Karababa, E. Cicek, N. Aksoy. Evaluation of Oxidant/Antioxidant Status and Cytokine Levels in Patients with Cannabis Use Disorder. Clin. Psychopharmacol. Neurosci., 2017. [DOI | PubMed]
- R. Pacifici, P. Zuccaro, M. Farré, S. Poudevida, S. Abanades, S. Pichini, K. Langohr, J. Segura, R. de La Torre. Combined Immunomodulating Properties of 3,4-Methylenedioxymethamphetamine (MDMA) and Cannabis in Humans. Addict. Abingdon Engl., 2007. [DOI]
- E. Fernandez-Egea, L. Scoriels, S. Theegala, M. Giro, S.E. Ozanne, K. Burling, P.B. Jones. Cannabis Use is Associated with Increased CCL11 Plasma Levels in Young Healthy Volunteers. Prog. Neuropsychopharmacol. Biol. Psychiatry, 2013. [DOI | PubMed]
- C.B. Ribeiro, F.D.O.F. de Castro, G.P. Dorneles, J.B. de Sousa Barros, J.M. Silva, C. Tavares, H.R. Carvalho, L. Carlos da Cunha, P. Nagib, C. Hoffmann. The Concomitant Use of Cannabis and Cocaine Coexists with Increased LPS Levels and Systemic Inflammation in Male Drug Users. Cytokine, 2021. [DOI | PubMed]
- L. Keen, A.D. Turner. Differential Effects of Self-Reported Lifetime Marijuana Use on Interleukin-1 Alpha and Tumor Necrosis Factor in African American Adults. J. Behav. Med., 2015. [DOI | PubMed]
- P. Araos, M. Pedraz, A. Serrano, M. Lucena, V. Barrios, N. García-Marchena, R. Campos-Cloute, J.J. Ruiz, P. Romero, J. Suárez. Plasma Profile of Pro-Inflammatory Cytokines and Chemokines in Cocaine Users under Outpatient Treatment: Influence of Cocaine Symptom Severity and Psychiatric Co-Morbidity. Addict. Biol., 2015. [DOI | PubMed]
- F.P. Moreira, J.R.C. Medeiros, A.C. Lhullier, L.D.M. de Souza, K. Jansen, L.V. Portela, D.R. Lara, R.A. da Silva, C.D. Wiener, J.P. Oses. Cocaine Abuse and Effects in the Serum Levels of Cytokines IL-6 and IL-10. Drug Alcohol Depend., 2016. [DOI | PubMed]
- R. Maza-Quiroga, N. García-Marchena, P. Romero-Sanchiz, V. Barrios, M. Pedraz, A. Serrano, R. Nogueira-Arjona, J.J. Ruiz, M. Soria, R. Campos. Evaluation of Plasma Cytokines in Patients with Cocaine Use Disorders in Abstinence Identifies Transforming Growth Factor Alpha (Tgfα) as a Potential Biomarker of Consumption and Dual Diagnosis. PeerJ, 2017. [DOI]
- J.H. Halpern, M.B. Sholar, J. Glowacki, N.K. Mello, J.H. Mendelson, A.J. Siegel. Diminished Interleukin-6 Response to Proinflammatory Challenge in Men and Women after Intravenous Cocaine Administration. J. Clin. Endocrinol. Metab., 2003. [DOI | PubMed]
- M. Pedraz, P. Araos, N. García-Marchena, A. Serrano, P. Romero-Sanchiz, J. Suárez, E. Castilla-Ortega, F. Mayoral-Cleries, J.J. Ruiz, A. Pastor. Sex Differences in Psychiatric Comorbidity and Plasma Biomarkers for Cocaine Addiction in Abstinent Cocaine-Addicted Subjects in Outpatient Settings. Front. Psychiatry, 2015. [DOI | PubMed]
- M.L. Levandowski, A.R.B. Hess, R. Grassi-Oliveira, R.M.M. de Almeida. Plasma Interleukin-6 and Executive Function in Crack Cocaine-Dependent Women. Neurosci. Lett., 2016. [DOI | PubMed]
- R.N. Achur, W.M. Freeman, K.E. Vrana. Circulating Cytokines as Biomarkers of Alcohol Abuse and Alcoholism. J. Neuroimmune Pharm., 2010. [DOI | PubMed]
- H.F. Moura, F. Hansen, F. Galland, D. Silvelo, F.P. Rebelatto, F. Ornell, R. Massuda, J.N. Scherer, F. Schuch, F.H. Kessler. Inflammatory Cytokines and Alcohol Use Disorder: Systematic Review and Meta-Analysis. Braz. J. Psychiatry, 2022. [DOI | PubMed]
- F.T. Crews, R. Bechara, L.A. Brown, D.M. Guidot, P. Mandrekar, S. Oak, L. Qin, G. Szabo, M. Wheeler, J. Zou. Cytokines and Alcohol. Alcohol Clin. Exp. Res., 2006. [DOI | PubMed]
- N. García-Marchena, R. Maza-Quiroga, A. Serrano, V. Barrios, N. Requena-Ocaña, J. Suárez, J.A. Chowen, J. Argente, G. Rubio, M. Torrens. Abstinent Patients with Alcohol Use Disorders Show an Altered Plasma Cytokine Profile: Identification of Both Interleukin 6 and Interleukin 17A as Potential Biomarkers of Consumption and Comorbid Liver and Pancreatic Diseases. J. Psychopharmacol., 2020. [DOI | PubMed]
- H.C. Fox, V. Milivojevic, A. MacDougall, H. LaVallee, C. Simpson, G.A. Angarita, R. Sinha. Stress-Related Suppression of Peripheral Cytokines Predicts Future Relapse in Alcohol-Dependent Individuals with and without Subclinical Depression. Addict. Biol., 2020. [DOI]
- K.H. Clark, C.A. Wiley, C.W. Bradberry. Psychostimulant Abuse and Neuroinflammation: Emerging Evidence of Their Interconnection. Neurotox. Res., 2013. [DOI | PubMed]
- J.M. Loftis, D. Choi, W. Hoffman, M.S. Huckans. Methamphetamine Causes Persistent Immune Dysregulation: A Cross-Species, Translational Report. Neurotox. Res., 2011. [DOI | PubMed]
- B.K. Yamamoto, A. Moszczynska, G.A. Gudelsky. Amphetamine Toxicities Classical and Emerging Mechanisms. Ann. N. Y. Acad. Sci., 2010. [DOI | PubMed]
- B.K. Yamamoto, J. Raudensky. The Role of Oxidative Stress, Metabolic Compromise, and Inflammation in Neuronal Injury Produced by Amphetamine-Related Drugs of Abuse. J. Neuroimmune Pharm., 2008. [DOI | PubMed]
- F. Qiu, C.L. Liang, H. Liu, Y.Q. Zeng, S. Hou, S. Huang, X. Lai, Z. Dai. Impacts of Cigarette Smoking on Immune Responsiveness: Up and down or Upside Down?. Oncotarget, 2017. [DOI | PubMed]
- J. Lee, V. Taneja, R. Vassallo. Cigarette Smoking and Inflammation: Cellular and Molecular Mechanisms. J. Dent. Res., 2012. [DOI | PubMed]
- C. Walter, N. Ferreirós, P. Bishay, G. Geisslinger, I. Tegeder, J. Lötsch. Exogenous Delta9-Tetrahydrocannabinol Influences Circulating Endogenous Cannabinoids in Humans. J. Clin. Psychopharmacol., 2013. [DOI | PubMed]
- P. Manza, K. Yuan, E. Shokri-Kojori, D. Tomasi, N.D. Volkow. Brain Structural Changes in Cannabis Dependence: Association with MAGL. Mol. Psychiatry, 2020. [DOI | PubMed]
- T.R. Spindle, H. Kuwabara, A. Eversole, A. Nandi, R. Vandrey, D.G. Antoine, A. Umbricht, A.S. Guarda, D.F. Wong, E.M. Weerts. Brain Imaging of Cannabinoid Type I (CB 1) Receptors in Women with Cannabis Use Disorder and Male and Female Healthy Controls. Addict. Biol., 2021. [DOI | PubMed]
- J. Ceccarini, R. Kuepper, D. Kemels, J. van Os, C. Henquet, K. van Laere. [18F]MK-9470 PET Measurement of Cannabinoid CB1 Receptor Availability in Chronic Cannabis Users. Addict. Biol., 2015. [DOI | PubMed]
- J. Hirvonen, R.S. Goodwin, C.T. Li, G.E. Terry, S.S. Zoghbi, C. Morse, V.W. Pike, N.D. Volkow, M.A. Huestis, R.B. Innis. Reversible and Regionally Selective Downregulation of Brain Cannabinoid CB1 Receptors in Chronic Daily Cannabis Smokers. Mol. Psychiatry, 2012. [DOI | PubMed]
- F.J. Pavõn, P. Araos, A. Pastor, M. Calado, M. Pedraz, R. Campos-Cloute, J.J. Ruiz, A. Serrano, E. Blanco, P. Rivera. Evaluation of Plasma-Free Endocannabinoids and Their Congeners in Abstinent Cocaine Addicts Seeking Outpatient Treatment: Impact of Psychiatric Co-Morbidity. Addict. Biol., 2013. [DOI | PubMed]
- C.D. Voegel, S.L. Kroll, M.W. Schmid, A.-K. Kexel, M.R. Baumgartner, T. Kraemer, T.M. Binz, B.B. Quednow. Alterations of Stress-Related Glucocorticoids and Endocannabinoids in Hair of Chronic Cocaine Users. Int. J. Neuropsychopharmacol., 2022. [DOI | PubMed]
- G. Kunos. Interactions Between Alcohol and the Endocannabinoid System. Alcohol Clin. Exp. Res., 2020. [DOI | PubMed]
- N. Garcia-Marchena, F.J. Pavon, A. Pastor, P. Araos, M. Pedraz, P. Romero-Sanchiz, M. Calado, J. Suarez, E. Castilla-Ortega, L. Orio. Plasma Concentrations of Oleoylethanolamide and Other Acylethanolamides Are Altered in Alcohol-Dependent Patients: Effect of Length of Abstinence. Addict. Biol., 2017. [DOI | PubMed]
- H. Yang, C. Chen. Cyclooxygenase-2 in Synaptic Signaling. Curr. Pharm. Des., 2008. [DOI | PubMed]
- J.R. Copeland, K.A. Willoughby, R.J. Police, E.F. Ellis. Repeated Cocaine Administration Reduces Bradykinin-Induced Dilation of Pial Arterioles. Am. J. Physiol., 1996. [DOI]
- D.S. Mastrogiannis, W.F. O’Brien. Cocaine Affects Prostaglandin Production in Human Umbilical Cord Cell Cultures. J. Matern. Fetal. Neonatal. Med., 2003. [DOI | PubMed]
- R.F. Anton, C.L. Randall. Central Nervous System Prostaglandins and Ethanol. Alcohol. Clin. Exp. Res., 1987. [DOI | PubMed]
- R.C. Murphy, J.Y. Westcott. Synthesis of Prostaglandins and Leukotrienes. Effects of Ethanol. Recent Dev. Alcohol., 1985. [DOI | PubMed]
- N. García-Marchena, N. Pizarro, F.J. Pavón, M. Martínez-Huélamo, M. Flores-López, N. Requena-Ocaña, P. Araos, D. Silva-Peña, J. Suárez, L.J. Santín. Potential Association of Plasma Lysophosphatidic Acid (LPA) Species with Cognitive Impairment in Abstinent Alcohol Use Disorders Outpatients. Sci. Rep., 2020. [DOI | PubMed]
- H. Holtmann, K. Resch. Cytokines. Naturwissenschaften, 1995. [DOI | PubMed]
- C. Liu, D. Chu, K. Kalantar-Zadeh, J. George, H.A. Young, G. Liu. Cytokines: From Clinical Significance to Quantification. Adv. Sci., 2021. [DOI]
- T.D. Purves-Tyson, U. Weber-Stadlbauer, J. Richetto, D.A. Rothmond, M.A. Labouesse, M. Polesel, K. Robinson, C. Shannon Weickert, U. Meyer. Increased Levels of Midbrain Immune-Related Transcripts in Schizophrenia and in Murine Offspring after Maternal Immune Activation. Mol. Psychiatry, 2021. [DOI | PubMed]
- B. Pedraz-Petrozzi, O. Elyamany, C. Rummel, C. Mulert. Effects of Inflammation on the Kynurenine Pathway in Schizophrenia—A Systematic Review. J. Neuroinflammation, 2020. [DOI | PubMed]
- S. Momtazmanesh, A. Zare-Shahabadi, N. Rezaei. Cytokine Alterations in Schizophrenia: An Updated Review. Front. Psychiatry, 2019. [DOI | PubMed]
- J.D. Rosenblat, R.S. McIntyre. Bipolar Disorder and Inflammation. Psychiatr. Clin. N. Am., 2016. [DOI]
- K. Munkholm, M. Vinberg, L. Vedel Kessing. Cytokines in Bipolar Disorder: A Systematic Review and Meta-Analysis. J. Affect. Disord., 2013. [DOI | PubMed]
- M.C. Miller, K.H. Mayo. Chemokines from a Structural Perspective. Int. J. Mol. Sci., 2017. [DOI | PubMed]
- D. Raman, T. Sobolik-Delmaire, A. Richmond. Chemokines in Health and Disease. Exp. Cell Res., 2011. [DOI | PubMed]
- W. Rostène, M.A. Dansereau, D. Godefroy, J. van Steenwinckel, A.R. Goazigo, S. Mélik-Parsadaniantz, E. Apartis, S. Hunot, N. Beaudet, P. Sarret. Neurochemokines: A Menage a Trois Providing New Insights on the Functions of Chemokines in the Central Nervous System. J. Neurochem., 2011. [DOI | PubMed]
- S. Najjar, S. Pahlajani, V. de Sanctis, J.N.H. Stern, A. Najjar, D. Chong. Neurovascular Unit Dysfunction and Blood–Brain Barrier Hyperpermeability Contribute to Schizophrenia Neurobiology: A Theoretical Integration of Clinical and Experimental Evidence. Front. Psychiatry, 2017. [DOI | PubMed]
- B.J. Miller, P. Buckley, W. Seabolt, A. Mellor, B. Kirkpatrick. Meta-Analysis of Cytokine Alterations in Schizophrenia: Clinical Status and Antipsychotic Effects. Biol. Psychiatry, 2011. [DOI | PubMed]
- D.R. Goldsmith, M.H. Rapaport, B.J. Miller. A Meta-Analysis of Blood Cytokine Network Alterations in Psychiatric Patients: Comparisons between Schizophrenia, Bipolar Disorder and Depression. Mol. Psychiatry, 2016. [DOI | PubMed]
- A.K. Wang, B.J. Miller. Meta-Analysis of Cerebrospinal Fluid Cytokine and Tryptophan Catabolite Alterations in Psychiatric Patients: Comparisons between Schizophrenia, Bipolar Disorder, and Depression. Schizophr. Bull., 2018. [DOI | PubMed]
- N. Çakici, A.L. Sutterland, B.W.J.H. Penninx, V.A. Dalm, L. de Haan, N.J.M. van Beveren. Altered Peripheral Blood Compounds in Drug-Naïve First-Episode Patients with Either Schizophrenia or Major Depressive Disorder: A Meta-Analysis. Brain Behav. Immun., 2020. [DOI | PubMed]
- D. Frydecka, M. Krzystek-Korpacka, A. Lubeiro, F. Stramecki, B. Stańczykiewicz, J.A. Beszłej, P. Piotrowski, K. Kotowicz, M. Szewczuk-Bogusławska, E. Pawlak-Adamska. Profiling Inflammatory Signatures of Schizophrenia: A Cross-Sectional and Meta-Analysis Study. Brain Behav. Immun., 2018. [DOI | PubMed]
- A. Isgren, C. Sellgren, C.J. Ekman, J. Holmén-Larsson, K. Blennow, H. Zetterberg, J. Jakobsson, M. Landén. Markers of Neuroinflammation and Neuronal Injury in Bipolar Disorder: Relation to Prospective Clinical Outcomes. Brain Behav. Immun., 2017. [DOI | PubMed]
- V. Johansson, S. Erhardt, G. Engberg, M. Kegel, M. Bhat, L. Schwieler, K. Blennow, H. Zetterberg, T.D. Cannon, L. Wetterberg. Twin Study Shows Association between Monocyte Chemoattractant Protein-1 and Kynurenic Acid in Cerebrospinal Fluid. Eur. Arch. Psychiatry Clin. Neurosci., 2020. [DOI | PubMed]
- B.I. Perry, R. Upthegrove, N. Kappelmann, P.B. Jones, S. Burgess, G.M. Khandaker. Associations of Immunological Proteins/Traits with Schizophrenia, Major Depression and Bipolar Disorder: A Bi-Directional Two-Sample Mendelian Randomization Study. Brain Behav. Immun., 2021. [DOI | PubMed]
- G. Sathyanarayanan, H. Thippeswamy, R. Mani, M. Venkataswamy, M. Kumar, M. Philip, P.S. Chandra. Cytokine Alterations in First-Onset Postpartum Psychosis-Clues for Underlying Immune Dysregulation. Asian J. Psychiatr., 2019. [DOI | PubMed]
- V. di Marzo, F. Piscitelli. The Endocannabinoid System and Its Modulation by Phytocannabinoids. Neurotherapeutics, 2015. [DOI | PubMed]
- S. Zou, U. Kumar. Cannabinoid Receptors and the Endocannabinoid System: Signaling and Function in the Central Nervous System. Int. J. Mol. Sci., 2018. [DOI | PubMed]
- T. Sugiura, S. Kondo, A. Sukagawa, S. Nakane, A. Shinoda, K. Itoh, A. Yamashita, K. Waku. 2-Arachidonoylglycerol: A Possible Endogenous Cannabinoid Receptor Ligand in Brain. Biochem. Biophys. Res. Commun., 1995. [DOI | PubMed]
- N. Stella. Endocannabinoid Signaling in Microglial Cells. Neuropharmacology, 2009. [DOI | PubMed]
- K. Kleberg, H.A. Hassing, H.S. Hansen. Classical Endocannabinoid-like Compounds and Their Regulation by Nutrients. Biofactors, 2014. [DOI | PubMed]
- B.E. Alger. Getting High on the Endocannabinoid System. Cerebrum, 2013. [PubMed]
- N. De Marchi, L. de Petrocellis, P. Orlando, F. Daniele, F. Fezza, V. di Marzo. di Endocannabinoid Signalling in the Blood of Patients with Schizophrenia. Lipids Health Dis., 2003. [DOI | PubMed]
- F.M. Leweke, D. Piomelli, F. Pahlisch, D. Muhl, C.W. Gerth, C. Hoyer, J. Klosterkötter, M. Hellmich, D. Koethe. Cannabidiol Enhances Anandamide Signaling and Alleviates Psychotic Symptoms of Schizophrenia. Transl. Psychiatry, 2012. [DOI | PubMed]
- P.C. Calde. Eicosanoids. Essays Biochem., 2020. [DOI | PubMed]
- J.A. Mitchell, N.S. Kirkby. Eicosanoids, Prostacyclin and Cyclooxygenase in the Cardiovascular System. Br. J. Pharm., 2019. [DOI]
- J.L. Wallace. Eicosanoids in the Gastrointestinal Tract. Br. J. Pharm., 2019. [DOI]
- D. Wang, R.N. Dubois. Eicosanoids and Cancer. Nat. Rev. Cancer, 2010. [DOI | PubMed]
- R.G. Biringer. The Role of Eicosanoids in Alzheimer’s Disease. Int. J. Env. Res. Public Health, 2019. [DOI | PubMed]
- B.D. Hammock, W. Wang, M.M. Gilligan, D. Panigrahy. Eicosanoids: The Overlooked Storm in Coronavirus Disease 2019 (COVID-19)?. Am. J. Pathol., 2020. [DOI | PubMed]
- E.A. Dennis, P.C. Norris. Eicosanoid Storm in Infection and Inflammation. Nat. Rev. Immunol., 2015. [DOI | PubMed]
- M.P. Wymann, R. Schneiter. Lipid Signalling in Disease. Nat. Rev. Mol. Cell Biol., 2008. [DOI | PubMed]
- H. Harizi, J.B. Corcuff, N. Gualde. Arachidonic-Acid-Derived Eicosanoids: Roles in Biology and Immunopathology. Trends Mol. Med., 2008. [DOI | PubMed]
- M.H. Law, R.G.H. Cotton, G.E. Berger. The Role of Phospholipases A2 in Schizophrenia. Mol. Psychiatry, 2006. [DOI | PubMed]
- W.F. Gattaz. Phospholipase A2 in Schizophrenia. Biol. Psychiatry, 1992. [DOI | PubMed]
- L. Schmidt, U. Ceglarek, L. Kortz, M. Hoop, K. Kirkby, J. Thiery, H. Himmerich. Mechanisms of Involvement of Eicosanoids and Their Precursors in the Pathophysiology and Treatment of Schizophrenia. Med. Chem., 2013. [DOI | PubMed]
- W.Y. Ong, T. Farooqui, G. Kokotos, A.A. Farooqui. Synthetic and Natural Inhibitors of Phospholipases A2: Their Importance for Understanding and Treatment of Neurological Disorders. ACS Chem. Neurosci., 2015. [DOI | PubMed]
- S.T. Tan, T. Ramesh, X.R. Toh, L.N. Nguyen. Emerging Roles of Lysophospholipids in Health and Disease. Prog. Lipid Res., 2020. [DOI | PubMed]
- Y. Xu, Y.J. Xiao, L.M. Baudhuin, B.M. Schwartz. The Role and Clinical Applications of Bioactive Lysolipids in Ovarian Cancer. J. Soc. Gynecol. Investig., 2001. [DOI | PubMed]
- J.W. Choi, J. Chun. Lysophospholipids and Their Receptors in the Central Nervous System. Biochim. Biophys. Acta, 2013. [DOI | PubMed]
- M.O. Cunningham, J. Hunt, S. Middleton, F.E.N. LeBeau, M.G. Gillies, C.H. Davies, P.R. Maycox, M.A. Whittington, C. Racca. Region-Specific Reduction in Entorhinal Gamma Oscillations and Parvalbumin-Immunoreactive Neurons in Animal Models of Psychiatric Illness. J. Neurosci., 2006. [DOI | PubMed]
- L. Musazzi, E. di Daniel, P. Maycox, G. Racagni, M. Popoli. Abnormalities in α/β-CaMKII and Related Mechanisms Suggest Synaptic Dysfunction in Hippocampus of LPA1 Receptor Knockout Mice. Int. J. Neuropsychopharmacol., 2011. [DOI | PubMed]
- C. Roberts, P. Winter, C.S. Shilliam, Z.A. Hughes, C. Langmead, P.R. Maycox, L.A. Dawson. Neurochemical Changes in LPA1 Receptor Deficient Mice—A Putative Model of Schizophrenia. Neurochem. Res., 2005. [DOI | PubMed]
- S.M. Harrison, C. Reavill, G. Brown, J.T. Brown, J.E. Cluderay, B. Crook, C.H. Davies, L.A. Dawson, E. Grau, C. Heidbreder. LPA1 Receptor-Deficient Mice Have Phenotypic Changes Observed in Psychiatric Disease. Mol. Cell Neurosci., 2003. [DOI | PubMed]
- L.M. Ellman, R.H. Yolken, S.L. Buka, E.F. Torrey, T.D. Cannon. Cognitive Functioning Prior to the Onset of Psychosis: The Role of Fetal Exposure to Serologically Determined Influenza Infection. Biol. Psychiatry, 2009. [DOI | PubMed]
- P.C. Sham, E. O’Callaghan, N. Takei, G.K. Murray, E.H. Hare, R.M. Murray. Schizophrenia Following Pre-Natal Exposure to Influenza Epidemics between 1939 and 1960. Br. J. Psychiatry, 1992. [DOI | PubMed]
- E. O’Callaghan, T. Gibson, H.A. Colohan, D. Walshe, P. Buckley, C. Larkin, J.L. Waddington. Season of Birth in Schizophrenia. Evidence for Confinement of an Excess of Winter Births to Patients without a Family History of Mental Disorder. Br. J. Psychiatry, 1991. [DOI | PubMed]
- C.E. Barr, S.A. Mednick, P. Munk-Jorgensen. Exposure to Influenza Epidemics during Gestation and Adult Schizophrenia. A 40-Year Study. Arch. Gen. Psychiatry, 1990. [DOI | PubMed]
- R.E. Kendell, I.W. Kemp. Maternal Influenza in the Etiology of Schizophrenia. Arch. Gen. Psychiatry, 1989. [DOI | PubMed]
- S.A. Mednick, R.A. Machon, M.O. Huttunen, D. Bonett. Adult Schizophrenia Following Prenatal Exposure to an Influenza Epidemic. Arch. Gen. Psychiatry, 1988. [DOI | PubMed]
- M. Cannon, D. Cotter, V.P. Coffey, P.C. Sham, N. Takei, C. Larkin, R.M. Murray, E. O’Callaghan. Prenatal Exposure to the 1957 Influenza Epidemic and Adult Schizophrenia: A Follow-up Study. Br. J. Psychiatry, 1996. [DOI | PubMed]
- H.J. Sorensen, E.L. Mortensen, J.M. Reinisch, S.A. Mednick. Association Between Prenatal Exposure to Bacterial Infection and Risk of Schizophrenia. Schizophr. Bull., 2009. [DOI | PubMed]
- A.S. Brown, P. Cohen, S. Greenwald, E. Susser. Nonaffective Psychosis after Prenatal Exposure to Rubella. Am. J. Psychiatry, 2000. [DOI | PubMed]
- P.B. Mortensen, B. Nørgaard-Pedersen, B.L. Waltoft, T.L. Sørensen, D. Hougaard, E.F. Torrey, R.H. Yolken. Toxoplasma Gondii as a Risk Factor for Early-Onset Schizophrenia: Analysis of Filter Paper Blood Samples Obtained at Birth. Biol. Psychiatry, 2007. [DOI | PubMed]
- A.S. Brown, E.J. Derkits. Prenatal Infection and Schizophrenia: A Review of Epidemiologic and Translational Studies. Am. J. Psychiatry, 2013. [DOI]
- K.V. Sandhu, E. Sherwin, H. Schellekens, C. Stanton, T.G. Dinan, J.F. Cryan. Feeding the Microbiota-Gut-Brain Axis: Diet, Microbiome, and Neuropsychiatry. Transl. Res., 2017. [DOI | PubMed]
- X. Wang, P.J. Quinn. Lipopolysaccharide: Biosynthetic Pathway and Structure Modification. Prog. Lipid Res., 2010. [DOI | PubMed]
- G.R. Barclay. Endogenous Endotoxin-Core Antibody (EndoCAb) as a Marker of Endotoxin Exposure and a Prognostic Indicator: A Review. Prog. Clin. Biol. Res., 1995. [PubMed]
- B. Cai, M. Wang, X. Zhu, J. Xu, W. Zheng, Y. Zhang, F. Zheng, Z. Feng, J. Zhu. The Fab Fragment of a Humanized Anti-Toll Like Receptor 4 (TLR4) Monoclonal Antibody Reduces the Lipopolysaccharide Response via TLR4 in Mouse Macrophages. Int. J. Mol. Sci, 2015. [DOI | PubMed]
- A. Morcuende, F. Navarrete, E. Nieto, J. Manzanares, T. Femenía. Inflammatory Biomarkers in Addictive Disorders. Biomolecules, 2021. [DOI | PubMed]
- C. Cui, D. Shurtleff, R.A. Harris. Neuroimmune Mechanisms of Alcohol and Drug Addiction. Int. Rev. Neurobiol., 2014. [DOI | PubMed]
- G.C. Baldwin, D.P. Tashkin, D.M. Buckley, A.N. Park, S.M. Dubinett, M.D. Roth. Marijuana and Cocaine Impair Alveolar Macrophage Function and Cytokine Production. Am. J. Respir. Crit. Care Med., 1997. [DOI | PubMed]
- M.D. Roth, A. Arora, S.H. Barsky, E.C. Kleerup, M. Simmons, D.P. Tashkin. Airway Inflammation in Young Marijuana and Tobacco Smokers. Am. J. Respir. Crit. Care Med., 1998. [DOI | PubMed]
- K.L. Bailey, T.A. Wyatt, D.M. Katafiasz, K.W. Taylor, A.J. Heires, J.H. Sisson, D.J. Romberger, E.L. Burnham. Alcohol and Cannabis Use Alter Pulmonary Innate Immunity. Alcohol, 2019. [DOI | PubMed]
- T.W. Klein, B. Lane, C.A. Newton, H. Friedman. The Cannabinoid System and Cytokine Network. Proc. Soc. Exp. Biol. Med., 2000. [DOI | PubMed]
- K.C.M. Verhoeckx, H.A.A.J. Korthout, A.P. van Meeteren-Kreikamp, K.A. Ehlert, M. Wang, J. van der Greef, R.J.T. Rodenburg, R.F. Witkamp. Unheated Cannabis Sativa Extracts and Its Major Compound THC-Acid Have Potential Immuno-Modulating Properties Not Mediated by CB1 and CB2 Receptor Coupled Pathways. Int. Immunopharmacol., 2006. [DOI | PubMed]
- T.W. Klein. Cannabinoid-Based Drugs as Anti-Inflammatory Therapeutics. Nat. Rev. Immunol., 2005. [DOI | PubMed]
- S.U. Nayak, S. Cicalese, C. Tallarida, C.F. Oliver, S.M. Rawls. Chemokine CCR5 and Cocaine Interactions in the Brain: Cocaine Enhances Mesolimbic CCR5 MRNA Levels and Produces Place Preference and Locomotor Activation That Are Reduced by a CCR5 Antagonist. Brain Behav. Immun., 2020. [DOI | PubMed]
- A. Guyon, D. Skrzydelski, I. de Giry, C. Rovère, G. Conductier, J.M. Trocello, V. Daugé, P. Kitabgi, W. Rostène, J.L. Nahon. Long Term Exposure to the Chemokine CCL2 Activates the Nigrostriatal Dopamine System: A Novel Mechanism for the Control of Dopamine Release. Neuroscience, 2009. [DOI | PubMed]
- J. Trecki, E.M. Unterwald. Modulation of Cocaine-Induced Activity by Intracerebral Administration of CXCL12. Neuroscience, 2009. [DOI | PubMed]
- J.M. Trocello, W. Rostene, S. Melik-Parsadaniantz, D. Godefroy, E. Roze, P. Kitabgi, W.A. Kuziel, S. Chalon, J. Caboche, E. Apartis. Implication of CCR2 Chemokine Receptor in Cocaine-Induced Sensitization. J. Mol. Neurosci., 2011. [DOI | PubMed]
- F.T. Crews, J. Zou, L. Qin. Induction of Innate Immune Genes in Brain Create the Neurobiology of Addiction. Brain Behav. Immun., 2011. [DOI | PubMed]
- C. Adams, J.H. Conigrave, J. Lewohl, P. Haber, K.C. Morley. Alcohol Use Disorder and Circulating Cytokines: A Systematic Review and Meta-Analysis. Brain Behav. Immun., 2020. [DOI | PubMed]
- R.S. Hofford, S.J. Russo, D.D. Kiraly. Neuroimmune Mechanisms of Psychostimulant and Opioid Use Disorders. Eur. J. Neurosci., 2019. [DOI | PubMed]
- C.J. Hillard. Lipids and Drugs of Abuse. Life Sci., 2005. [DOI | PubMed]
- F. Navarrete, M.S. García-Gutiérrez, A. Gasparyan, D. Navarro, F. López-Picón, Á. Morcuende, T. Femenía, J. Manzanares. Biomarkers of the Endocannabinoid System in Substance Use Disorders. Biomolecules, 2022. [DOI | PubMed]
- Y. Akaneya. The Remarkable Mechanism of Prostaglandin E2 on Synaptic Plasticity. Gene Regul. Syst. Biol., 2007
- T. Yamamoto, K. Anggadiredja, T. Hiranita. New Perspectives in the Studies on Endocannabinoid and Cannabis: A Role for the Endocannabinoid-Arachidonic Acid Pathway in Drug Reward and Long-Lasting Relapse to Drug Taking. J. Pharm. Sci., 2004. [DOI]
- H. Famitafreshi, M. Karimian. Prostaglandins as the Agents That Modulate the Course of Brain Disorders. Degener. Neurol. Neuromuscul. Dis., 2020. [DOI | PubMed]
- L. Orio, F.J. Pavón, E. Blanco, A. Serrano, P. Araos, M. Pedraz, P. Rivera, M. Calado, J. Suárez, F. Rodríguez De Fonseca. Lipid Transmitter Signaling as a New Target for Treatment of Cocaine Addiction: New Roles for Acylethanolamides and Lysophosphatidic Acid. Curr. Pharm. Des, 2013. [DOI | PubMed]
- T. Trang, M. Sutak, R. Quirion, K. Jhamandas. The Role of Spinal Neuropeptides and Prostaglandins in Opioid Physical Dependence. Br. J. Pharm., 2002. [DOI | PubMed]
- A.R. Mohn, W.D. Yao, M.G. Caron. Genetic and Genomic Approaches to Reward and Addiction. Neuropharmacology, 2004. [DOI | PubMed]
- N.D. Volkow, M. Michaelides, R. Baler. The Neuroscience of Drug Reward and Addiction. Physiol. Rev., 2019. [DOI | PubMed]
- H. Pribiag, D. Stellwagen. TNF-α Downregulates Inhibitory Neurotransmission through Protein Phosphatase 1-Dependent Trafficking of GABAA Receptors. J. Neurosci., 2013. [DOI | PubMed]
- H. Pribiag, D. Stellwagen. Neuroimmune Regulation of Homeostatic Synaptic Plasticity. Neuropharmacology, 2014. [DOI | PubMed]
- D. Stellwagen, R.C. Malenka. Synaptic Scaling Mediated by Glial TNF-Alpha. Nature, 2006. [DOI | PubMed]
- A. Avital, I. Goshen, A. Kamsler, M. Segal, K. Iverfeldt, G. Richter-Levin, R. Yirmiya. Impaired Interleukin-1 Signaling is Associated with Deficits in Hippocampal Memory Processes and Neural Plasticity. Hippocampus, 2003. [DOI | PubMed]
- F. Mori, R. Nisticò, G. Mandolesi, S. Piccinin, D. Mango, H. Kusayanagi, N. Berretta, A. Bergami, A. Gentile, A. Musella. Interleukin-1β Promotes Long-Term Potentiation in Patients with Multiple Sclerosis. Neuromolecular. Med., 2014. [DOI | PubMed]
- S. Heinisch, L.G. Kirby. SDF-1alpha/CXCL12 Enhances GABA and Glutamate Synaptic Activity at Serotonin Neurons in the Rat Dorsal Raphe Nucleus. Neuropharmacology, 2010. [DOI | PubMed]
- W. Rostène, P. Kitabgi, S.M. Parsadaniantz. Chemokines: A New Class of Neuromodulator?. Nat. Rev. Neurosci., 2007. [DOI | PubMed]
- A. Guyon, J.L. Nahon. Multiple Actions of the Chemokine Stromal Cell-Derived Factor-1alpha on Neuronal Activity. J. Mol. Endocrinol., 2007. [DOI | PubMed]
- R.C. Paolicelli, G. Bolasco, F. Pagani, L. Maggi, M. Scianni, P. Panzanelli, M. Giustetto, T.A. Ferreira, E. Guiducci, L. Dumas. Synaptic Pruning by Microglia is Necessary for Normal Brain Development. Science, 2011. [DOI | PubMed]
- D.P. Schafer, E.K. Lehrman, A.G. Kautzman, R. Koyama, A.R. Mardinly, R. Yamasaki, R.M. Ransohoff, M.E. Greenberg, B.A. Barres, B. Stevens. Microglia Sculpt Postnatal Neural Circuits in an Activity and Complement-Dependent Manner. Neuron, 2012. [DOI | PubMed]
- M. Pruessner, A.E. Cullen, M. Aas, E.F. Walker. The Neural Diathesis-Stress Model of Schizophrenia Revisited: An Update on Recent Findings Considering Illness Stage and Neurobiological and Methodological Complexities. Neurosci. Biobehav. Rev., 2017. [DOI | PubMed]
- T. Vargas, R.E. Conley, V.A. Mittal. Chronic Stress, Structural Exposures and Neurobiological Mechanisms: A Stimulation, Discrepancy and Deprivation Model of Psychosis. Int. Rev. Neurobiol., 2020. [DOI | PubMed]
- R. Nusslock, G.E. Miller. Early-Life Adversity and Physical and Emotional Health across the Lifespan: A Neuro-Immune Network Hypothesis. Biol. Psychiatry, 2016. [DOI | PubMed]
- M.J. Millan, A. Andrieux, G. Bartzokis, K. Cadenhead, P. Dazzan, P. Fusar-Poli, J. Gallinat, J. Giedd, D.R. Grayson, M. Heinrichs. Altering the Course of Schizophrenia: Progress and Perspectives. Nat. Rev. Drug Discov., 2016. [DOI | PubMed]
- M.F. Sheehan. Dual Diagnosis. Psychiatr. Q., 1993. [DOI | PubMed]
- K.T. Mueser, R.E. Drake, M.A. Wallach. Dual Diagnosis: A Review of Etiological Theories. Addict. Behav., 1998. [DOI | PubMed]
- S.E. Nennig, J.R. Schank. The Role of NFkB in Drug Addiction: Beyond Inflammation. Alcohol, 2017. [DOI | PubMed]
- S. Gerondakis, T.S. Fulford, N.L. Messina, R.J. Grumont. NF-KB Control of T Cell Development. Nat. Immunol., 2014. [DOI | PubMed]
- M.S. Hayden, S. Ghosh. Signaling to NF-KappaB. Genes Dev., 2004. [DOI | PubMed]
- A. Oeckinghaus, S. Ghosh. The NF-KB Family of Transcription Factors and Its Regulation. Cold Spring Harb. Perspect. Biol., 2009. [DOI]
- B. García-Bueno, J.R. Caso, J.C. Leza. Stress as a Neuroinflammatory Condition in Brain: Damaging and Protective Mechanisms. Neurosci. Biobehav. Rev., 2008. [DOI | PubMed]
- E. Ang, J. Chen, P. Zagouras, H. Magna, J. Holland, E. Schaeffer, E.J. Nestler. Induction of Nuclear Factor-KappaB in Nucleus Accumbens by Chronic Cocaine Administration. J. Neurochem., 2001. [DOI | PubMed]
- S.J. Russo, M.B. Wilkinson, M.S. Mazei-Robison, D.M. Dietz, I. Maze, V. Krishnan, W. Renthal, A. Graham, S.G. Birnbaum, T.A. Green. Nuclear Factor KB Signaling Regulates Neuronal Morphology and Cocaine Reward. J. Neurosci., 2009. [DOI | PubMed]
- X. Zhang, Y. Cui, J. Jing, Y. Cui, W. Xin, X. Liu. Involvement of P38/NF-KB Signaling Pathway in the Nucleus Accumbens in the Rewarding Effects of Morphine in Rats. Behav. Brain Res., 2011. [DOI | PubMed]
- F. Crews, K. Nixon, D. Kim, J. Joseph, B. Shukitt-Hale, L. Qin, J. Zou. BHT Blocks NF-KappaB Activation and Ethanol-Induced Brain Damage. Alcohol. Clin. Exp. Res., 2006. [DOI | PubMed]
- L. Qin, J. He, R.N. Hanes, O. Pluzarev, J.S. Hong, F.T. Crews. Increased Systemic and Brain Cytokine Production and Neuroinflammation by Endotoxin Following Ethanol Treatment. J. Neuroinflammation, 2008. [DOI | PubMed]
- J. Zou, F. Crews. Induction of Innate Immune Gene Expression Cascades in Brain Slice Cultures by Ethanol: Key Role of NF-KB and Proinflammatory Cytokines. Alcohol. Clin. Exp. Res., 2010. [DOI | PubMed]
- B. García-Bueno, J.L.M. Madrigal, I. Lizasoain, M.A. Moro, P. Lorenzo, J.C. Leza. Peroxisome Proliferator-Activated Receptor Gamma Activation Decreases Neuroinflammation in Brain after Stress in Rats. Biol. Psychiatry, 2005. [DOI | PubMed]
- L. Brunetti, A. Carrieri, L. Piemontese, P. Tortorella, F. Loiodice, A. Laghezza. Beyond the Canonical Endocannabinoid System. A Screening of PPAR Ligands as FAAH Inhibitors. Int J. Mol. Sci., 2020. [DOI | PubMed]
- A. Jablensky. The Diagnostic Concept of Schizophrenia: Its History, Evolution, and Future Prospects. Dialogues Clin. Neurosci., 2010. [DOI | PubMed]
- I.I. Gottesman, T.D. Gould. The Endophenotype Concept in Psychiatry: Etymology and Strategic Intentions. Am. J. Psychiatry, 2003. [DOI | PubMed]
- L. Hosak, K. Hakeem, M. Raad, J. Studnicka. Is Microvascular Abnormality a New Endophenotype in Schizophrenia?. Psychiatr. Danub., 2015. [PubMed]
- S. Smesny, S. Klemm, M. Stockebrand, S. Grunwald, U.J. Gerhard, T. Rosburg, H. Sauer, B. Blanz. Endophenotype Properties of Niacin Sensitivity as Marker of Impaired Prostaglandin Signalling in Schizophrenia. Prostaglandins Leukot Essent Fat. Acids, 2007. [DOI | PubMed]
- M.T. Tsuang, W.S. Stone, S.V. Faraone. Toward Reformulating the Diagnosis of Schizophrenia. Am. J. Psychiatry, 2000. [DOI | PubMed]
- M. Torrens, J.I. Mestre-Pintó, L. Montanari, J. Vicente, A. Domingo-Salvany. Dual Diagnosis: An European Perspective. Adicciones, 2017. [DOI | PubMed]
- R. Alsuhaibani, D.C. Smith, R. Lowrie, S. Aljhani, V. Paudyal. Scope, Quality and Inclusivity of International Clinical Guidelines on Mental Health and Substance Abuse in Relation to Dual Diagnosis, Social and Community Outcomes: A Systematic Review. BMC Psychiatry, 2021. [DOI | PubMed]
- M.N. Szerman. Patología Dual: Integrando La Salud Mental. Rev. Int. Investig. Adicciones, 2017. [DOI]
- M.S. Keshavan, Y. Kaneko. Secondary Psychoses: An Update. World Psychiatry, 2013. [DOI | PubMed]
- M.S.K. Starzer, M. Nordentoft, C. Hjorthøj. Rates and Predictors of Conversion to Schizophrenia or Bipolar Disorder Following Substance-Induced Psychosis. Am. J. Psychiatry, 2018. [DOI | PubMed]
- C.U. Correll, O.D. Howes. Treatment-Resistant Schizophrenia: Definition, Predictors, and Therapy Options. J. Clin. Psychiatry, 2021. [DOI]
- M.A. Silva, D. Restrepo. Functional Recovery in Schizophrenia. Rev. Colomb. Psiquiatr. Engl. Ed., 2019. [DOI | PubMed]
- A. Seppälä, J. Pylvänäinen, H. Lehtiniemi, N. Hirvonen, I. Corripio, H. Koponen, J. Seppälä, A. Ahmed, M. Isohanni, J. Miettunen. Predictors of Response to Pharmacological Treatments in Treatment-Resistant Schizophrenia—A Systematic Review and Meta-Analysis. Schizophr. Res., 2021. [DOI | PubMed]
- A. Levchenko, T. Nurgaliev, A. Kanapin, A. Samsonova, R.R. Gainetdinov. Current Challenges and Possible Future Developments in Personalized Psychiatry with an Emphasis on Psychotic Disorders. Heliyon, 2020. [DOI | PubMed]
- N. Çakici, N.J.M. van Beveren, G. Judge-Hundal, M.M. Koola, I.E.C. Sommer. An Update on the Efficacy of Anti-Inflammatory Agents for Patients with Schizophrenia: A Meta-Analysis. Psychol. Med., 2019. [DOI | PubMed]
- A. Catalan, G. Salazar de Pablo, J. Vaquerizo Serrano, P. Mosillo, H. Baldwin, A. Fernández-Rivas, C. Moreno, C. Arango, C.U. Correll, I. Bonoldi. Annual Research Review: Prevention of Psychosis in Adolescents–Systematic Review and Meta-Analysis of Advances in Detection, Prognosis and Intervention. J. Child Psychol. Psychiatry, 2021. [DOI | PubMed]
- C. Agostoni, M. Nobile, V. Ciappolino, G. Delvecchio, A. Tesei, S. Turolo, A. Crippa, A. Mazzocchi, C.A. Altamura, P. Brambilla. The Role of Omega-3 Fatty Acids in Developmental Psychopathology: A Systematic Review on Early Psychosis, Autism, and ADHD. Int. J. Mol. Sci., 2017. [DOI | PubMed]
- R.M. Murray, A.S. David, O. Ajnakina. Prevention of Psychosis: Moving on from the at-Risk Mental State to Universal Primary Prevention. Psychol. Med., 2021. [DOI | PubMed]
- J. Mcgrath. Universal Interventions for the Primary Prevention of Schizophrenia. Aust. N. Z. J. Psychiatry, 2000. [DOI | PubMed]
- R. Warner. The Prevention of Schizophrenia: What Interventions Are Safe and Effective?. Schizophr. Bull., 2001. [DOI | PubMed]
- A.S. Brown, J.J. McGrath. The Prevention of Schizophrenia. Schizophr. Bull., 2011. [DOI | PubMed]
- I.E. Sommer, C.E. Bearden, E. van Dellen, E.J. Breetvelt, S.N. Duijff, K. Maijer, T. van Amelsvoort, L. de Haan, R.E. Gur, C. Arango. Early Interventions in Risk Groups for Schizophrenia: What Are We Waiting For?. NPJ Schizophr., 2016. [DOI | PubMed]
- M. di Forti, D. Quattrone, T.P. Freeman, G. Tripoli, C. Gayer-Anderson, H. Quigley, V. Rodriguez, H.E. Jongsma, L. Ferraro, C. la Cascia. The Contribution of Cannabis Use to Variation in the Incidence of Psychotic Disorder across Europe (EU-GEI): A Multicentre Case-Control Study. Lancet Psychiatry, 2019. [DOI | PubMed]
- J.A. Collins, B.C.J.M. Fauser. Balancing the Strengths of Systematic and Narrative Reviews. Hum. Reprod. Update, 2005. [DOI | PubMed]
- O. Köhler-Forsberg, N. Müller, B.R. Lennox. Editorial: The Role of Inflammation in the Etiology and Treatment of Schizophrenia. Front. Psychiatry, 2020. [DOI | PubMed]
- A. Caviedes, C. Lafourcade, C. Soto, U. Wyneken. BDNF/NF-KB Signaling in the Neurobiology of Depression. Curr. Pharm. Des., 2017. [DOI | PubMed]
- V. Anttila, B. Bulik-Sullivan, H.K. Finucane, R.K. Walters, J. Bras, L. Duncan, V. Escott-Price, G.J. Falcone, P. Gormley, R. Malik. Analysis of Shared Heritability in Common Disorders of the Brain. Science, 2018. [DOI]