
Anti-Microbial Activity of Phytocannabinoids and Endocannabinoids in the Light of Their Physiological and Pathophysiological Roles
Abstract
Antibiotic resistance has become an increasing challenge in the treatment of various infectious diseases, especially those associated with biofilm formation on biotic and abiotic materials. There is an urgent need for new treatment protocols that can also target biofilm-embedded bacteria. Many secondary metabolites of plants possess anti-bacterial activities, and especially the phytocannabinoids of the Cannabis sativa L. varieties have reached a renaissance and attracted much attention for their anti-microbial and anti-biofilm activities at concentrations below the cytotoxic threshold on normal mammalian cells. Accordingly, many synthetic cannabinoids have been designed with the intention to increase the specificity and selectivity of the compounds. The structurally unrelated endocannabinoids have also been found to have anti-microbial and anti-biofilm activities. Recent data suggest for a mutual communication between the endocannabinoid system and the gut microbiota. The present review focuses on the anti-microbial activities of phytocannabinoids and endocannabinoids integrated with some selected issues of their many physiological and pharmacological activities.
Article type: Review Article
Keywords: anti-microbial activity, anti-biofilm activity, endocannabinoids, gut microbiota, pathogens, phytocannabinoids
License: © 2022 by the authors. CC BY 4.0 Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Article links: DOI: 10.3390/biomedicines10030631 | PubMed: 35327432 | PMC: PMC8945038
Relevance: Moderate: mentioned 3+ times in text
Full text: PDF (2.8 MB)
1. Introduction
Plant medicine has often been used for the treatment of diverse diseases, including bacterial and fungal infections [ref. 1,ref. 2,ref. 3,ref. 4,ref. 5,ref. 6,ref. 7,ref. 8]. The plants produce a series of secondary metabolites, many of which have pharmacological as well as anti-microbial activities [ref. 4,ref. 5,ref. 6,ref. 9,ref. 10,ref. 11]. Evolutionarily, plants have developed various anti-microbial mechanisms to protect them from infectious diseases [ref. 11]. Usually, these include the production of compounds that have anti-biofilm and bacteriostatic activities rather than biocidal effect [ref. 11]. Compounds with anti-biofilm activities are believed not to induce resistance mechanisms in the microbes, since they target processes not essential for their survival. In contrast, compounds with bactericidal activity might lead to the development of resistance mechanisms in the microbe as part of the bacterial fitness adaptation process with increased probability of developing microbial plant infections.
Cannabis sativa L. subspecies are plants that contain a large variety of secondary metabolites, including phytocannabinoids, terpenoids and flavonoids, which have profound anti-microbial activities, in addition to possessing anti-inflammatory, anti-oxidative and neuromodulatory properties [ref. 12,ref. 13,ref. 14]. In mammalians, the phytocannabinoids interact with the same receptors (e.g., cannabinoid receptors CB1 and CB2) as the endocannabinoids [ref. 15], which are endogenous substances with anti-microbial, anti-inflammatory and neuromodulatory activities [ref. 16,ref. 17,ref. 18,ref. 19,ref. 20,ref. 21,ref. 22,ref. 23,ref. 24]. While much is known about the cannabinoid targets in mammalians, so far, little is known about the microbial targets of these compounds. It is likely that these compounds also interact with specific targets in the microbes. The present review focuses on the anti-microbial activities of phytocannabinoids and endocannabinoids interwoven with selected aspects of their many physiological and pathophysiological activities.
2. Cannabis sativa L.
The hemp plant (Cannabis sativa L.; L = Linnaeus) belonging to the family Cannabaceae, originates in central-northeast Asia where it has been cultivated for more than 5000 years [ref. 15,ref. 25,ref. 26]. The Han Chinese dynasty used Cannabis to treat inflammatory disorders and malaria [ref. 27,ref. 28]. The Chinese pharmacopoeia of the Emperor Shen Nung, who lived approximately around 2700 BCE and is considered “The Father of Chinese Medicine”, indicated Cannabis plant usage for the treatment of rheumatic pain, constipation, malaria, and gynecological disorders [ref. 26]. In modern times, this plant has been used for different medical conditions, including alleviating chronic pain (e.g., in cancer patients and in rheumatic diseases), muscle spasms (e.g., in multiple sclerosis), epileptic convulsion (e.g., Dravet syndrome and Lennox–Gastaut syndrome in children), nausea (e.g., following chemotherapy), intestinal inflammation (e.g., colitis, inflammatory bowel disease (IBD)), and for stimulating appetite (e.g., in devastating AIDS syndrome, anorexia, and cancer patients) [ref. 26,ref. 29,ref. 30]. It has also been used as a treatment remedy for cancer patients, since the phytocannabinoids can inhibit cell growth of certain tumor cells and enhance the efficacy of certain cancer therapeutics [ref. 31].
The phenotypes of Cannabis plants are highly variable and can be classified into three major subspecies: Cannabis sativa subsp. sativa, Cannabis sativa subsp. indica, and Cannabis sativa subsp. ruderalis [ref. 32]. The different subspecies have all been classified to the Cannabis sativa L. species [ref. 32]. There are also several chemovariants, chemotypes, or cultivars of this plant harboring different composition of chemical compounds [ref. 33,ref. 34,ref. 35,ref. 36]. Different Cannabis cultivars or chemotypes have been developed that contain various ratios of cannabidiol (CBD) and Δ9-tetrahydrocannabinol (Δ9-THC), and even those containing high CBD and low Δ9-THC content, which is favorable for avoiding the psychomimetic effects of Δ9-THC [ref. 33,ref. 37]. The cannabinoids are found in most parts of the plant, with the highest concentrations in glandular trichomes on the surfaces of leaves and flowers [ref. 38,ref. 39,ref. 40,ref. 41,ref. 42].
The chemical composition of Cannabis is affected by the ripeness and maturation state of the plant, growth conditions, the sowing and the harvest times, as well as the storage conditions [ref. 34,ref. 38,ref. 39,ref. 40,ref. 41,ref. 43]. The plant composition of phytocannabinoids is affected by light, temperature, water supply, nutrition, heavy metals, phytohormones, soil bacteria, insects and microbial pathogens, among others [ref. 44,ref. 45,ref. 46,ref. 47]. Cannabidiolic acid (CBDA), the precursor of cannabinols, predominates in the unripen plant, while it is converted to CBD, Δ9-THC and cannabinol (CBN) upon ripening of the resin [ref. 48]. In the intermediate ripening state, CBD is predominant, then Δ9-THC dominates in the ripened state, while CBN, the final conversion product, is the major compound in the overripened resin [ref. 48]. High anti-microbial activity was found especially in unripen Cannabis harvested from regions with unfavorable climate for this plant, whereas ripened Cannabis taken from tropical areas had a more hashish-active composition [ref. 48]. For the optimal production of essential oil, the recommended stage for harvest is one to three weeks before seed maturity [ref. 43].
The difference between industrial hemp and the high Δ9-THC hemp breed type marijuana is that the industrial hemp contains minute amounts of Δ9-THC (less than 0.2% (w/v)), while marijuana flowers and leaves may contain as much as 17–28% Δ9-THC [ref. 49]. Even concentrated THC products, such as oil, shatter, and dab, have been produced with a concentration of up to 95% Δ9-THC [ref. 49]. The use of marijuana is associated with hallucinations due to the high Δ9-THC content and may lead to addiction, lack of judgement, and reduced cognition, especially during adolescence when the brain is undergoing significant development [ref. 49]. Smoking hemp may lead to decreased immune function with a consequent increase in opportunistic infections [ref. 50,ref. 51,ref. 52,ref. 53]. Cannabis users have a higher probability to get fungal infections than non-Cannabis users, which might in part be due to fungal contamination of the Cannabis product [ref. 54].
2.1. Anti-Microbial Activity of Cannabis sativa L. Extracts
Z. Krejčí, in the 1950s, observed that Cannabis has antibiotic activity and introduced it to the clinics in Czechoslovakia [ref. 55], a practice that was discontinued in 1990 [ref. 33]. The first compound identified by Krejčí with antibiotic activity was named cannabidiolic acid (CBDA) [ref. 56,ref. 57]. From then on, several other Cannabis components with antibiotic activities have been isolated and characterized [ref. 48,ref. 58,ref. 59,ref. 60,ref. 61,ref. 62,ref. 63], which will be further discussed below. In 1956, L. Ferenczy published a paper documenting that plant seeds from various plant species, including those from Cannabis sativa, exhibited antibacterial activity, especially against Gram-positive bacteria [ref. 64]. Wasim et al. [ref. 65] documented that both ethanolic and petroleum ether extracts of Cannabis sativa leaves showed anti-microbial activity against Bacillus subtilis, Staphylococcus aureus, Micrococcus flavus, Bordetella bronchiseptica, Proteus vulgaris, Aspergillus niger, and Candida albicans. Ali et al. [ref. 66] observed that the oil of the seeds of Cannabis sativa exerted pronounced anti-bacterial activity against Bacillus subtilis and Staphylococcus aureus, with moderate activity against Escherichia coli and Pseudomonas aeruginosa, without any activity against Aspergillus niger and Candida albicans. The petroleum ether extract of the whole plant showed high anti-bacterial activity against Bacillus subtilis and Staphylococcus aureus, moderate activity against Escherichia coli, while no activity against Pseudomonas aeruginosa or the tested fungi [ref. 66]. Thus, the extraction method and the source affect the composition of the anti-microbial content and the spectrum of responding microbes.
2.2. Anti-Microbial Activity of Essential Oils from Cannabis sativa L.
Novak et al. [ref. 67] analyzed the anti-bacterial effect of essential oils prepared from five different cultivars of Cannabis sativa L. These essential oils contained, among others, α-pinene, myrcene, trans-β-ocimene, α-terpinolene, trans-caryophyllene, and α-humulene, but undetectable levels of Δ9-THC and very poor levels of other cannabinoids [ref. 67]. They observed differences in the anti-bacterial activity between the various cultivars. All five essential oils showed anti-bacterial activity against Acinetobacter calcoaceticus, Beneckea natriegens, Brochothrix thermosphacta and Staphylococcus aureus [ref. 67]. Only one of the five essential oils had an anti-bacterial effect on Escherichia coli, while none affected Enterobacter aerogenes, Klebsiella pneumoniae, Proteus vulgaris, Salmonella pullorum, Serratia marcescens, or Streptococcus faecalis [ref. 67].
Nissen et al. [ref. 34] observed that essential oils of Cannabis sativa L., prepared from 50–70% of seed maturity, showed anti-bacterial activity against the Gram-positive bacteria Enterococcus faecium and Streptococcus salivarius at less than 1% (v/v) but were unable to inhibit the growth of the yeast Saccharomyces cerevisiae. Zengin et al. [ref. 68] found that essential oils distilled from leaves, inflorescences, and thinner stems of the hemp plant showed anti-oxidative properties and had significant anti-bacterial activity against clinical Helicobacter pylori strains (MIC = 16–64 μg/mL), with lower activity against clinical Staphylococcus aureus isolates (MIC = 8 mg/mL) and no significant activity against Candida spp. and Malassezia spp. The minimum bacterial biofilm inhibitory concentration (MBIC) of the hemp essential oil against Helicobacter pyroli was similar to the MIC [ref. 68]. The hemp essential oil showed cytotoxicity against human breast cancer, cholangiocarcinoma, and colon carcinoma cell lines at 50–75 μg/mL, while 250 μg/mL was required to inhibit the cell proliferation of a nonmalignant cholangiocyte cell line [ref. 68]. The LD50 of hemp essential oil against larvae of Galleria mellonella was found to be 1.56 mg/mL, which is much higher than the anti-bacterial activity against Helicobacter pyroli, but lower than that found to be active against Staphylococcus aureus strains [ref. 68].
Pellegrini et al. [ref. 69] observed that essential oil prepared from Cannabis sativa L. cultivar Futura 75 inflorescences with low Δ9-THC content (<0.2%) cultivated in the Abruzzo territory showed anti-bacterial activity against Staphylococcus aureus and Listeria monocytogenes with a MIC of 1.25–5 µL/mL, while being ineffective against Salmonella enterica. They also showed that the essential oil possessed anti-oxidative properties [ref. 69]. The essential oils produced from the Cannabis sativa L. cultivar Futura 75 inflorescences was also found to have insecticidal activity with LD50 values of 65.8 μg/larva on Spodoptera littoralis, 122.1 μg/adult on Musca domestica, and LC50 of 124.5 μL/L on Culex quinquefasciatus larvae [ref. 70]. The insecticidal effect might in part be due to an inhibition of the enzyme acetylcholinesterase (AChE) [ref. 70]. Thomas et al. [ref. 71] found that essential oil of Cannabis sativa could induce 100% mortality in the mosquito larvae of Culex tritaeniorhynchus, Anopheles stephensi, Aedes aegypti, and Culex quinquefasciatus at concentrations of 0.06, 0.1, 0.12, and 0.2 μL/mL, respectively.
Palmieri et al. [ref. 72] studied the variability of Cannabis essential oils from various origins and observed that the time of distillation affected the chemical composition of terpenic components, sesquiterpenes, and CBD with consequent variations in the anti-microbial activities against Staphylococcus aureus, Listeria monocytogenes, and Enterococcus faecium. Zheljazkov et al. [ref. 73] compared the anti-microbial activity of nine wild hemp (Cannabis sativa spp. spontanea Vavilov) accessions sampled from agricultural fields in northeastern Serbia with 13 EU registered cultivars, eight breeding lines, and one cannabidiol (CBD) hemp strain, which showed variations in the secondary metabolites β-caryophyllene, α-humulene, caryophyllene oxide, and humulene epoxide. The CBD concentration in the essential oils of wild hemp varied from 6.9 to 52.4%, while the CBD content in the essential oils of the registered cultivars, breeding lines, and the CBD strain varied from 7.1 to 25%; 6.4 to 25%; and 7.4 to 8.8%, respectively [ref. 73]. The Δ9-THC concentration showed high variability between the different strains, with the highest concentration being 3.5% [ref. 73]. The essential oils of the wild hemp had greater anti-microbial activity compared with the essential oil of registered cultivars [ref. 73]. In general, with variations between the different essential oils, anti-microbial activity was observed toward Staphylococcus aureus, Enterococcus faecalis, Streptococcus pneumoniae, Pseudomonas aeruginosa, Yersenia enterocolitica, Salmonella enterica, Candida albicans, Candida krusei, and Candida tropicalis using the disc diffusion method [ref. 73]. Altogether, the data presented above show that there is high variability of the composition of hemp essential oils, which might explain the many contradictory publications of the anti-microbial activities toward the same microbial species. In general, a good anti-bacterial response is achieved on Gram-positive bacteria, with less or no effect on Gram-negative bacteria, and variable effect on fungi.
2.3. Anti-Microbial Activity of Terpenoids in Cannabis Essential Oils
Several terpenoids in the Cannabis essential oils have been demonstrated to have anti-microbial effect, which include the monoterpenes α-pinene, linalool, and limonene, and the bitter-tasting sesquiterpenes nerolidol, β-caryophyllene, and caryophyllene oxide [ref. 33,ref. 74,ref. 75,ref. 76]. α-Pinene inhibited the growth of both Gram-positive bacteria (e.g., various Clostridium species, Enterococcus faecium, Streptococcus salivarius, Staphylococcus aureus, Staphylococcus epidermidis, Streptococcus pyogenes, Streptococcus pneumoniae) and Gram-negative bacteria (e.g., various Pseudomonas species), as well as the fungus Candida albicans [ref. 34,ref. 77,ref. 78,ref. 79]. Myrcene, which is also found in tea tree oil, inhibited the growth of Staphylococcus aureus that was associated with the leakage of K+ ions from the bacterial cells and damage to the cell membrane [ref. 80]. Linalool, a monoterpenoid alcohol, and α-terpineol, a fragrant terpene, showed anti-bacterial activity against Propionibacterium acne and Staphylococcus epidermidis with a minimum inhibitory concentration (MIC) of 0.625–1.25 µg/mL [ref. 77]. Linalool is also effective against the yeast and hyphal forms of Candida albicans, where it alters the membrane integrity and induces cell cycle arrest [ref. 81]. Limonene showed anti-bacterial activity against Staphylococcus epidermidis [ref. 77] and Listeria monocytogenes [ref. 82], and exerted anti-biofilm activity against Streptococcus pyogenes, Streptococcus mutans, and Streptococcus mitis [ref. 83]. α-Humulene showed potent anti-fungal activity against Cryptococcus neoformans, Candida glabrata, and Candida krusei with MIC values of 5.0, 1.45, and 10.0 μg/mL, respectively, without any effect on methicillin-sensitive Staphylococcus aureus (MSSA) 29213, methicillin-resistant Staphylococcus aureus (MRSA) 33591, or Mycobacterium intracellulare [ref. 84]. Nerolidol is a sesquiterpene with sedative properties and inhibits the growth of Leishmania amazonensis, Leishmania braziliensis, and Leishmania chagasi promastigotes, and Leishmania amazonensis amastigotes [ref. 85], as well as the growth of Plasmodium falciparum at the trophozoite and schizont stages [ref. 86,ref. 87]. The anti-oxidative β-caryophyllene possesses anti-microbial activity against Staphylococcus aureus (MIC 2–4 µM), Bacillus subtilis (MIC 6–10 µM), Escherichia coli (MIC 7–11 µM), Pseudomonas aeruginosa (6–8 µM), Aspergillus niger (MIC 5–7 µM), and Trichoderma reesei (MIC 3–5 µM) without any significant cytotoxic effect on normal mammalian cell lines [ref. 88]. The anti-inflammatory oxygenated sesquiterpene caryophyllene oxide exhibited anti-fungal activities against the dermatophytes Trichophyton mentagrophytes var. mentagrophytes, Trichophyton mentagrophytes var. interdigitale, and Trichophyton rubrum [ref. 89].
3. Phytocannabinoids
The Cannabis sativa L. plants produce more than 560 chemicals, including at least 144 cannabinoids and 200 terpenoids, as well as flavonoids and polyunsaturated fatty acids [ref. 15,ref. 33,ref. 34,ref. 42,ref. 63,ref. 67,ref. 72,ref. 73,ref. 90,ref. 91,ref. 92,ref. 93,ref. 94,ref. 95,ref. 96,ref. 97,ref. 98,ref. 99,ref. 100,ref. 101,ref. 102,ref. 103,ref. 104,ref. 105,ref. 106,ref. 107]. The most common phytocannabinoids are Δ9-tetrahydrocannabinol (Δ9-THC) and cannabidiol (CBD), which are the neutral homologs of tetrahydrocannabinolic acid (THCA) and cannabidiol acid (CBDA), respectively [ref. 108]. The phytocannabinoids are terpenophenolic compounds containing a resorcinyl core with a para-positioned isoprenyl, alkyl, or aralkyl side chain [ref. 39,ref. 40] (Figure 1). The tetrahydrobenzochromen ring is quite unique to the genus Cannabis, although a related compound has been found in the liverwort Radula marginata [ref. 109], and cannabigerol (CBG) and its corresponding acid have been isolated from Helichrysum umbraculigerum [ref. 110].
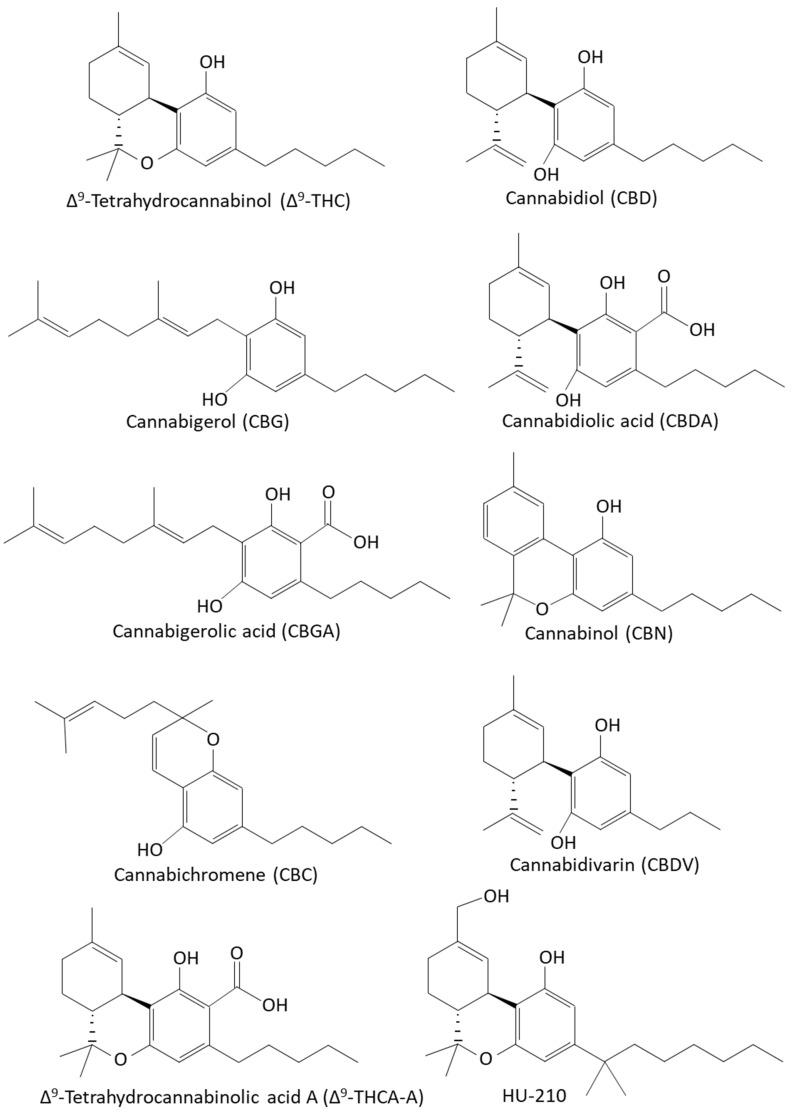
Apart from exerting anti-microbial activities, which will be discussed in more detail below (Section 3.3), phytocannabinoids modulate several physiological and pathophysiological processes in humans and other mammalians, making them potential therapeutic drugs in various settings [ref. 12,ref. 13,ref. 14,ref. 31,ref. 111,ref. 112,ref. 113,ref. 114,ref. 115]. Among others, these compounds have been shown to have anti-inflammatory, anti-oxidative, anti-nausea, anti-nociceptive, anti-convulsant, anti-neoplastic, anxiolytic, and neuroprotective properties [ref. 14,ref. 111,ref. 112,ref. 114,ref. 115,ref. 116,ref. 117]. Cannabinoids also affect cognition, such as learning and memory, consciousness, and emotion, including anxiety and depression [ref. 118,ref. 119].
Some cannabinoid-based drugs (e.g., Marinol, Syndros, Cesamet, Sativex, and Epidiolex) have been approved by the U.S. Food and Drug Administration (FDA) for the treatment of epilepsy, Dravet syndrome, Lennox–Gastaut syndrome, Parkinson’s disease, spasticity associated with multiple sclerosis, neuropathic pain, mental illnesses, chemotherapy-induced nausea, and AIDS wasting syndrome [ref. 117,ref. 120,ref. 121,ref. 122]. Marinol and Syndros contain the (-)-trans-Δ9-THC dronabinol; Cesamet contains the synthetic cannabinoid nabilone that shows structural similarities to Δ9-THC; and Epidiolex contains CBD. Sativex is produced from a Cannabis-derived extract that is composed of approximately equal quantities of Δ9-THC and CBD. A major concern is the production of many psychotropic synthetic cannabinoids distributed on the illicit market, which poses a potential health treat due to their high potency and toxicity [ref. 123].
3.1. Cannabinoid Receptors
The effects of phytocannabinoids on humans and other mammalians are partly mediated by the Gi/o protein-coupled CB1 (encoded by the CNR1 gene) and CB2 (encoded by the CNR2 gene) cannabinoid receptors that consist of seven transmembrane domains [ref. 124,ref. 125,ref. 126]. The stimulation of these receptors leads to the inhibition of adenylyl cyclase with consequent reduction in the intracellular cAMP levels, activation of potassium channels, activation of mitogen-activated protein kinases (MAPKs) such as the extracellular signal-regulated kinase (ERK) and c-Jun N-terminal kinase (JNK), as well as activation of the phosphoinositide-3 kinase (PI3K)/Akt signaling pathways and the mammalian target of rapamycin (mTOR) [ref. 126,ref. 127,ref. 128,ref. 129,ref. 130,ref. 131,ref. 132,ref. 133,ref. 134].
The CB1 and CB2 receptors also recognize the endogenous arachidonic acid-derived endocannabinoids, such as N-arachidonoylethanolamine (anandamide; AEA) and 2-arachidonoylglycerol (2-AG) [ref. 134,ref. 135,ref. 136]. Both CB1 and CB2 are expressed in various cells in the brain and in peripheral tissues [ref. 137]. CB1 is especially expressed at high levels in the neocortex, hippocampus, basal ganglia, cerebellum, and brainstem, but it is also found in peripheral nerve terminals and some tissues, such as the vascular endothelium, spleen, testis, and eye [ref. 137]. CB2 is predominantly found in cells of the immune system, and in the central nervous system, it is primarily localized to microglia and tissue macrophages [ref. 137].
The CB1 receptor regulates the balance between excitatory and inhibitory neuronal activity. The psychoactive effect is believed to be mediated through the CB1 receptor in the brain, whereas the immunomodulatory effects are anticipated to be mediated via the CB2 receptor expressed on immune cells [ref. 138,ref. 139]. In addition, CB1 signaling affects metabolism and is involved in maintaining whole body energy homeostasis by increasing appetite and stimulating feeding [ref. 140]. Many efforts have been made to develop CB2 specific agonists at an attempt to achieve anti-inflammatory actions without psychotropic adverse effects [ref. 13,ref. 141,ref. 142,ref. 143]. The sesquiterpene (E)-β-caryophyllene produced by Cannabis as well as other plants, including oregano (Origanum vulgare L.), cinnamon (Cinnamomum spp.), and black pepper (Piper nigrum L.), was found to bind selectively to the CB2 receptor and exert anti-inflammatory activities [ref. 144,ref. 145,ref. 146,ref. 147].
Other cannabinoid receptors include transient receptor potential vanilloid 1 (TRPV1), the G-protein-coupled receptors GPR18 and GPR55, and peroxisome proliferator activated receptors (PPARs) [ref. 126,ref. 134,ref. 148,ref. 149,ref. 150,ref. 151,ref. 152]. The anti-nociceptive effect of Cannabis sativa extracts was found to be mediated by the binding of CBD to TRPV1 [ref. 153]. A study by Ibrahim et al. [ref. 154] showed that activation of the CB2 receptor by its agonist AM1241 stimulated the release of beta-endorphin from keratinocytes, which, in turn, acted on neuronal μ-opoid receptors to inhibit nociception. The Cannabis sativa extract containing multiple cannabinoids, terpenes, and flavonoids had stronger anti-nociceptive effect than a single cannabinoid given alone [ref. 153], suggesting an “entourage” effect of the various Cannabis-containing compounds [ref. 74].
The CB1 and CB2 can form receptor heteromers [ref. 155]. The activity of the receptor heteromer is affected by the agonists and antagonists that bind to each of them. A CB1 antagonist can block the effect of a CB2 agonist and vice versa; a CB2 antagonist can block the effect of a CB1 receptor agonist [ref. 155]. CB1 has also been shown to form heteromers with dopamine and adenosine receptors [ref. 156,ref. 157,ref. 158], AT1 angiotensin receptor [ref. 159], μ1-opoid receptor [ref. 160,ref. 161], and OX1 orexin A receptor [ref. 162]. The many interacting partners put CB1 signaling under strict regulation.
3.2. Pharmacological Effects of Selected Phytocannabinoids
3.2.1. Δ9-Tetrahydrocannibinol (Δ9-THC)
Δ9-THC binds to CB1 and CB2 receptors at a more or less equal affinity [ref. 138,ref. 163,ref. 164]. It also acts on CB1-CB2 receptor heterodimers [ref. 165]. Δ9-THC is well known for its psychomimetic activities that are exerted by its binding to CB1 receptor in the brain, resulting in a calm and sedated mental state [ref. 49]. Besides euphoria, Δ9-THC is an appetite stimulator [ref. 166]. Oral Δ9-THC (Dronabinol, Marinol) and its synthetic nabilone (Cesamet) have been used for the treatment of nausea and appetite stimulation for people undergoing chemotherapy and for AIDS wasting syndrome [ref. 167,ref. 168]. The activation of CB1 by Δ9-THC is believed to mediate its anti-nausea and anti-emetic effects [ref. 169]. Sativex, which contains a combination of Δ9-THC and CBD, has been used for relief of neuropathic pain in multiple sclerosis [ref. 170].
3.2.2. Cannabidiol (CBD)
The non-psychotropic cannabidiol (CBD) shows low affinity to the CB1 and CB2 receptors [ref. 135] and can exert antagonistic modulatory actions on these receptors [ref. 138,ref. 171]. CBD can also activate the TRPV1 channel, serotonin 1A (5-HT1A) receptors, and opioid receptors [ref. 24,ref. 172]. CBD has anti-inflammatory, anti-oxidative, anti-epileptic, analgesic, anti-neoplastic, sedative, neuroprotective, and anti-anxiety activities [ref. 173,ref. 174,ref. 175,ref. 176,ref. 177,ref. 178,ref. 179,ref. 180,ref. 181,ref. 182,ref. 183,ref. 184,ref. 185,ref. 186,ref. 187,ref. 188]. Moreover, CBD inhibits sebocyte lipogenesis by activating the TRPV4 ion channel that interferes with the pro-lipogenic ERK1/2 MAPK pathway [ref. 189].
The neuroprotective activity of CBD has been attributed in part to its anti-oxidative activity [ref. 190,ref. 191]. Based on its immunomodulatory activities, CBD has been implicated in the treatment of various autoimmune diseases [ref. 14,ref. 21], and its anti-nociceptive activity was found to be beneficial in relieving chronic pain [ref. 192]. In addition, CBD has potential uses in psychiatry due to its neuromodulatory activities in the brain that control recognition, emotional and behavioral responses [ref. 111,ref. 193,ref. 194]. CBD has especially been reported to have therapeutic effect for psychopathological conditions, such as substance use disorders, chronic psychosis, and anxiety [ref. 193]. CBD has been shown to be well tolerated in humans at concentrations as high as 3500–6000 mg/day [ref. 195,ref. 196,ref. 197], and the FDA-approved CBD (marketed as Epidiolex) is indicated for preventing epileptic seizures in Lennox–Gastaut syndrome and Dravet syndrome in children [ref. 198].
In experimental mice and rat models, CBD has been shown to have immunosuppressive activities [ref. 181], which are partly due to inhibition of TNFα production [ref. 199,ref. 200] and induction of myeloid-derived suppressor cells (MDSCs) [ref. 201]. CBD alleviated the symptoms of experimental autoimmune encephalomyelitis (EAE) and collagen-induced arthritis and prevented the onset of autoimmune diabetes in experimental murine models [ref. 199,ref. 200,ref. 202]. In mice, the anti-inflammatory activity of CBD was found to have a bell-shaped dose–response with an optimal dose of 5 mg/kg [ref. 203]. The use of a standardized extract from a CBD-rich, ∆9-THClow Cannabis indica cultivar overcame this bell-shaped dose–response, suggesting a synergistic effect among the different compounds of the Cannabis extract [ref. 199].
3.2.3. Cannabigerol (CBG)
CBG is another non-psychoactive Cannabis component that is produced at elevated levels in some industrial hemps [ref. 204,ref. 205,ref. 206]. It binds to both CB1 and CB2 receptors and modulates the signaling through these receptors, as well as the CB1-CB2 receptor heteromer, at concentrations as low as 0.1–1 μM [ref. 207]. CBG competes with the binding of [3H]-WIN-55,212-2 to CB2, but not to CB1 [ref. 207]. Further studies suggest that CBG is a partial agonist of CB1 and CB2 [ref. 207,ref. 208,ref. 209]. CBG activates TRPV1, TRPV2, TRPV3, TRPV4, TRPA1, 5-HT1a receptor, α2-adrenergic receptor, and PPARγ, while being a TRPM8 antagonist [ref. 210,ref. 211,ref. 212,ref. 213,ref. 214,ref. 215]. CBG has anti-inflammatory, anti-oxidative, and anti-nociceptive activities [ref. 117,ref. 209,ref. 213,ref. 216]. The anti-inflammatory property is thought to be achieved by modulating the CB2 receptor, TRP channels, and PPARγ, and by inhibiting cyclooxygenase 1 and 2 (COX-1/2) [ref. 210,ref. 211,ref. 217], while the analgesic effect of CBG is thought to be mediated through the α2-adrenergic receptor [ref. 211]. CBG has been shown to have potential beneficial effects in treating inflammatory bowel disease and neurological disorders, such as Huntington’s disease, Parkinson’s disease, and multiple sclerosis [ref. 213,ref. 215,ref. 216,ref. 218,ref. 219].
3.2.4. Cannabichromene (CBC)
CBC is a non-psychoactive phytocannabinoid that activates the CB1 and CB2 receptors, resulting in decreased intracellular levels of cAMP [ref. 209]. CBC also activates the TRPA1, TRPV3, and TRPV4 channels [ref. 210]. CBC has anti-inflammatory, anti-nociceptive, and neuroprotective activities [ref. 220,ref. 221,ref. 222,ref. 223,ref. 224,ref. 225]. CBC reduces the activity of both the ON and OFF neurons in the rostral ventromedial medulla (RVM) and elevates the endocannabinoid levels in the ventrolateral periaqueductal gray matter [ref. 221]. The anti-nociceptive activity of CBC is mediated by the adenosine A1 and TRPA1 receptors [ref. 221]. CBC increases the viability of neural stem progenitor cells through activation of the adenosine A1 receptor [ref. 224]. Moreover, it has been shown to suppress reactive astrocytes, thus offering a protective effect against neuro-inflammation and Alzheimer’s disease [ref. 225]. CBC had anti-convulsant properties in a mouse model of Dravet syndrome [ref. 226], and it exhibited cytotoxic activity against some carcinoma cells [ref. 227,ref. 228].
3.2.5. Cannabidiolic Acid (CBDA)
CBDA has low affinity for both CB1 and CB2 receptors, with moderate inhibition of adenylyl cyclase activity [ref. 209,ref. 229], and functions as an allosteric regulator on the 5-HT1A receptor, resulting in anti-emetic effects [ref. 230,ref. 231,ref. 232,ref. 233]. In addition, it activates PPARα and PPARγ [ref. 212]. CBDA shows anti-nociceptive and anti-inflammatory effects that are in part mediated by COX-2 inhibition and activation of the TRPV1 channel [ref. 217,ref. 234,ref. 235]. CBDA has anxiolytic and anti-convulsant effects in animal models [ref. 236,ref. 237,ref. 238].
3.2.6. Cannabigerolic Acid (CBGA)
CBGA displays low affinity for both CB1 and CB2 receptors but causes a similar decrease in intracellular cAMP levels as Δ9-THC [ref. 229]. Since CBGA can activate PPARs [ref. 212], it is expected to affect lipid metabolism [ref. 117]. A Cannabis sativa cultivar containing high levels of CBG and CBGA inhibited the activity of the aldose reductase enzyme, which catalyzes the reduction of glucose to sorbitol [ref. 239]. Since the aldose reductase level is increased at high glucose levels and has been implicated in the development of neuropathy, nephropathy, retinopathy, and cataract in diabetes, CBGA has been suggested as a potential drug in preventing diabetic complications [ref. 239]. In the Scn1a+/− mouse model of Dravet syndrome, CBGA was found to have an anti-convulsant effect that was mediated by its interaction with the GPR55, TRPV1, and GABAA receptors [ref. 240].
3.2.7. Cannabinol (CBN)
CBN is formed during the degradation of Δ9-THC and has a lower binding affinity to CB1 and CB2 receptors than Δ9-THC [ref. 117]. CBN is an agonist of the TRPV1, TRPV2, TRPV3, TRPV4, and TRPA1 cation channels [ref. 210]. CBN is a non-psychotropic phytocannabinoid with analgesic and anti-inflammatory properties and acts as an appetite stimulant [ref. 117]. CBN has neuroprotective activity that is associated with its anti-oxidative actions, trophic support, and elimination of intraneuronal β-amyloid in neuronal cells [ref. 241]. CBN preserves mitochondrial functions, such as redox regulation, calcium uptake, mitochondrial membrane potential, and bioenergetics [ref. 242]. CBN promotes endogenous antioxidant defense mechanisms and triggers AMP-activated protein kinase (AMPK) signaling pathways [ref. 242].
3.3. Anti-Microbial Effects of Phytocannabinoids
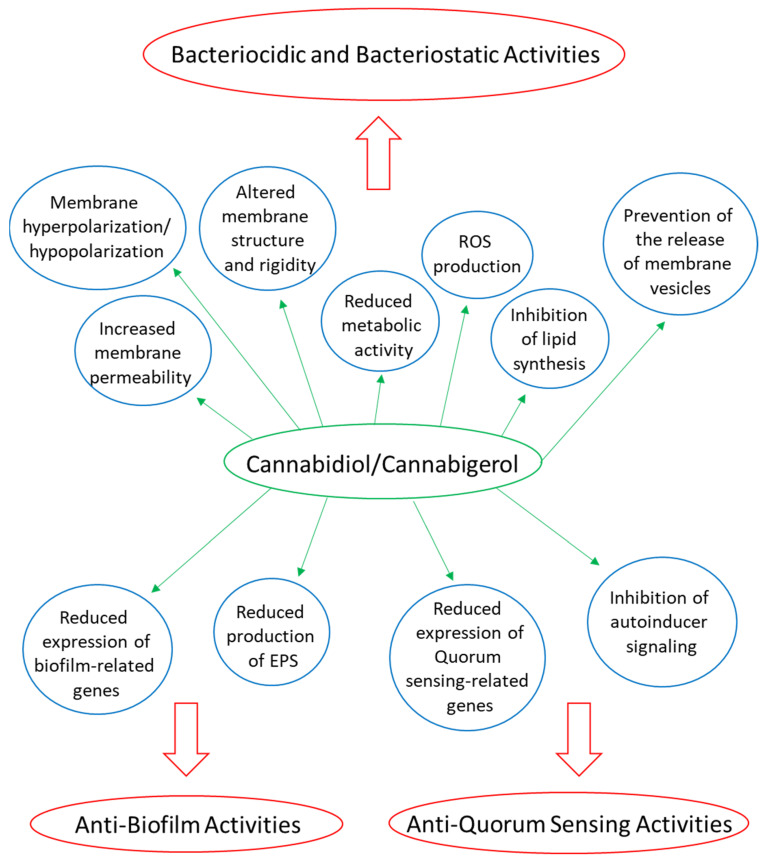
Several phytocannabinoids have been shown to have anti-bacterial activities, especially on Gram-positive bacteria, including various antibiotic-resistant strains [ref. 58,ref. 59,ref. 62,ref. 63,ref. 101,ref. 220,ref. 243,ref. 244,ref. 245,ref. 246,ref. 247] (Table 1). Phytocannabinoids have been shown to exert both bactericidal and bacteriostatic effects [ref. 61,ref. 62,ref. 244,ref. 247]. Most of the studies have analyzed the half maximal inhibitory concentration (IC50) or minimum inhibitory concentration (MIC) for each of the compounds against different bacterial species, fungi, and protozoa, while only a few studies have looked at the underlying mechanisms [ref. 61,ref. 243,ref. 244,ref. 247,ref. 248,ref. 249,ref. 250] (Figure 2).
Table 1: Examples of Cannabis sativa constituents that have been documented to possess anti-bacterial, anti-fungal, and/or anti-protozoal activities *.
| Phytocannabinoids | Anti-Microbial Activity | Reference |
|---|---|---|
| ∆9-Tetrahydrocannabinol (Δ9-THC) | MIC: 2–5 μg/mL against Staphylococcus aureus ATCC 6538MIC: 1 μg/mL against Staphylococcus aureus ATCC 25923MIC: 2 μg/mL against Staphylococcus aureus SA-1199B (NorA overexpression)MIC: 2 μg/mL against Staphylococcus aureus EMRSA-15MIC: 0.5 μg/mL against Staphylococcus aureus EMRSA-16MIC: 2 μg/mL against MRSA USA300MIC: 4–8 μg/mL against MRSA ATCC 43300MIC: 5 μg/mL against Streptococcus pyogenesMIC: 2 μg/mL against Streptococcus milleriMIC: 5 μg/mL against Streptococcus faecalisMIC: 4–8 μg/mL against Neisseria gonorrhoeae ATCC 19424IC50: 4.8 μM against Staphylococcus aureus ATCC 29213IC50: 6.9 μM against Bacillus cereus IIIM 25IC50: 2.8 μM against Lactococcus lactis MTCC 440IC50: 3.5 μM against Shigella boydii NC-09357IC50: 6.4 μM against Staphylococcus warneri MTCC 4436No effect against Escherichia coli, Salmonella typhi or Proteus vulgaris | [ref. 58,ref. 61,ref. 245,ref. 246,ref. 247] |
| Cannabidiol (CBD) | MIC: 1–5 μg/mL against S. aureus ATCC 6538MIC: 0.5–1 μg/mL against Staphylococcus aureus ATCC 25923MIC: 1 μg/mL against Staphylococcus aureus SA-1199B (NorA overexpression)MIC: 1 μg/mL against Staphylococcus aureus EMRSA-15MIC: 1 μg/mL against Staphylococcus aureus EMRSA-16MIC: 1–4 μg/mL against MRSA USA300MIC: 1–2 μg/mL against various Staphylococcus aureus isolates.MIC: 1–2 μg/mL against Staphylococcus epidermidis.MIC: 4 μg/mL against methicillin-resistant Staphylococcus epidermidis.MIC: 2 μg/mL against Streptococcus pyogenesMIC: 1 μg/mL against Streptococcus milleriMIC: 5 μg/mL against Streptococcus faecalisMIC: 1–4 μg/mL against various Streptococcus pneumoniae speciesMIC: 0.5–4 μg/mL against various Enterococcus faecalis speciesMIC: 4 μg/mL against Listereria monocytogenesMIC: 1–2 μg/mL against Cutibacterium (Propionibacterium) acnes ATCC 6919MIC: 2–4 μg/mL against Clostridioides (Clostridium) difficile M7404 human ribotype 027MIC: 1–2 μg/mL against various Neisseria gonorrhoeae isolates.MIC: 0.25 μg/mL against various Neisseria meningitidis ATCC 13090MIC: 1 μg/mL against Moraxella catarrhalis MMX 3782MIC: 1 μg/mL against Legionella pneumophila MMX 7515IC50: 3.8 μM against Staphylococcus aureus ATCC 29213IC50: 9.5–11.1 μM against Staphylococcus aureus ATCC 6538 IC50: 9.8 μM against Bacillus cereus IIIM 25IC50: 2.9 μM against Lactococcus lactis MTCC 440IC50: 4.3 μM against Shigella boydii NC-09357IC50: 4.1 μM against Pseudomonas fluorescens MTCC 103IC50: 5.7 μM against Staphylococcus warneri MTCC 4436Moderate effect against Mycobacterium smegmatis (MIC 16 μg/mL) and marginal activity against Mycobacterium tuberculosis H37Rv, Candida albicans, and Cryptococcus neoformans with a MIC > 64 μg/mL.No effect against Escherichia coli, Salmonella typhimurium, Shigella dysenteriae, Proteus vulgaris, Proteus mirabilis, Klebsiella pneumoniae, Pseudomonas aeruginosa, Acinetobacter baumannii, Serratia marcescens, Burkholderia cepacian, and Haemophilus influenzae.Anti-biofilm effect:MBEC: 1–4 μg/mL against MSSA and MRSA biofilms.BIC50: 12.5 μg/mL against Candida albicans SC5314MBIC: 100 μg/mL against Candida albicans SC5314 | [ref. 58,ref. 61,ref. 62,ref. 245,ref. 246,ref. 247,ref. 251,ref. 252,ref. 253] |
| Cannabigerol (CBG) | MIC: 0.5 μg/mL against Staphylococcus aureus ATCC 25923MIC: 1 μg/mL against Staphylococcus aureus SA-1199B (NorA overexpression)MIC: 2 μg/mL against Staphylococcus aureus EMRSA-15MIC: 1 μg/mL against Staphylococcus aureus EMRSA-16MIC: 2 μg/mL against MRSA USA300MIC: 2–4 μg/mL against various MRSA clinical isolates, with some requiring > 8 μg/mLMIC: 4–8 μg/mL against MRSA ATCC 43300MIC: 2.5 μg/mL against Streptococcus mutans UA159 ATCC 700610MIC: 1 μg/mL against Streptococcus sanguis ATCC 10556MIC: 5 μg/mL against Streptococcus sobrinus ATCC 27351MIC: 5 μg/mL against Streptococcus salivarius ATCC 25975MIC: 1–2 μg/mL against Neisseria gonorrhoeae ATCC 19424 IC50: 15 μg/mL against Mycobacterium intracellulareAnti-biofilm effect:MBIC: 2–4 μg/mL against biofilm formation by MRSA4 μg/mL eradicated preformed biofilms of MRSAMBIC: 2.5 μg/mL against biofilm formation by Streptococcus mutans UA159 ATCC 70061Anti-quorum sensing effect1 μg/mL CBG inhibited quorum sensing in Vibrio harveyi BB120. | [ref. 58,ref. 61,ref. 100,ref. 243,ref. 244,ref. 247,ref. 248] |
| Cannabidiolic acid (CBDA) | MIC: 1–2 μg/mL against Neisseria gonorrhoeae ATCC 19424MIC: 2 μg/mL against Staphylococcus aureus ATCC 25923MIC: 4 μg/mL against Staphylococcus aureus USA300MIC: 4 μg/mL against Staphylococcus epidermidis CA#71 and ATCC 51625MIC: 16–32 μg/mL against MRSA ATCC 43300No effect on Escherichia coli ATCC 25922 or Pseudomonas aeruginosa PA01 with a MIC > 64 μg/mL. | [ref. 62,ref. 247] |
| Cannabigerolic acid (CBGA) | IC50: 12 μg/mL against Leishmania donovaniMIC: 4 μg/mL against MRSA USA300MIC: 2–4 μg/mL against MRSA ATCC 43300MIC: 1–2 μg/mL against Neisseria gonorrhoeae ATCC 19424 | [ref. 61,ref. 100,ref. 247] |
| Cannabichromene (CBC) | MIC: 1.56 μg/mL against Staphylococcus aureus ATCC 6538MIC: 2 μg/mL against Staphylococcus aureus ATCC 25923MIC: 2 μg/mL against Staphylococcus aureus SA-1199B (NorA overexpression)MIC: 2 μg/mL against Staphylococcus aureus EMRSA-15MIC: 2 μg/mL against Staphylococcus aureus EMRSA-16MIC: 8 μg/mL against MRSA USA300MIC: 0.39 μg/mL against Bacillus subtilis ATCC 6633MIC 12.5 μg/mL against Mycobacterium smegmatis ATCC 607IC50: 5.9 μM against Staphylococcus aureus ATCC 29213IC50: 9.2 μM against Bacillus cereus IIIM 25IC50: 2.6 μM against Lactococcus lactis MTCC 440IC50: 3.4 μM against Shigella boydii NC-09357IC50: 5.6 μM against Staphylococcus warneri MTCC 4436 | [ref. 58,ref. 61,ref. 220,ref. 246] |
| Cannabichromenic acid (CBCA) | MIC: 2 μg/mL against MRSA USA300MIC: 7.8 μM against Staphylococcus aureus MSSA 34397MIC: 3.9 μM against a clinical MRSA isolateMIC: 7.8 μM against vancomycin-resistance Enterococcus faecalis (VRE) | [ref. 61,ref. 254] |
| Cannabinol (CBN) | MIC: 1 μg/mL against Staphylococcus aureus ATCC 25923MIC: 1 μg/mL against Staphylococcus aureus SA-1199B (NorA overexpression)MIC: 1 μg/mL against Staphylococcus aureus EMRSA-15MIC: 2 μg/mL against MRSA USA300IC50: 7.9 μM against Staphylococcus aureus ATCC 29213IC50: 3.2 μM against Bacillus cereus IIIM 25IC50: 5.8 μM against Lactococcus lactis MTCC 440IC50: 11.7 μM against Shigella boydii NC-09357IC50: 8.3 μM against Pseudomonas fluorescens MTCC 103IC50: 9.2 μM against Staphylococcus warneri MTCC 4436 | [ref. 58,ref. 61,ref. 246] |
| Cannabidivarin (CBDV) | MIC: 2–4 μg/mL against MRSA ATCC 43300MIC: 0.03–0.5 μg/mL against Neisseria gonorrhoeae ATCC 19424IC50: 7.8 μM against Staphylococcus aureus ATCC 29213IC50: 3.1 μM against Bacillus cereus IIIM 25IC50: 3.2 μM against Lactococcus lactis MTCC 440IC50: 10.4 μM against Shigella boydii NC-09357IC50: 5.9 μM against Pseudomonas fluorescens MTCC 103IC50: 7.9 μM against Staphylococcus warneri MTCC 4436IC50: 11.9 μM against Candida albicans MTCC 4748MIC: 8 μg/mL against MRSA USA300 | [ref. 61,ref. 246,ref. 247] |
| (-)Δ8-Tetrahydrocannabinol(Δ8-THC) | MIC: 2 μg/mL against MRSA USA300MIC: 4–8 μg/mL against MRSA ATCC 43300MIC: 2–4 μg/mL against Neisseria gonorrhoeae ATCC 19424 | [ref. 61,ref. 247] |
| Exo-tetrahydrocannabinol (exo-THC) | MIC: 2 μg/mL against MRSA USA300 | [ref. 61] |
| Δ9-Tetrahydrocannabinolicacid A (THCA-A) | MIC: 4 μg/mL against MRSA USA300 | [ref. 61] |
| Δ9-Tetrahydrocannabivarin (THCV) | MIC: 4 μg/mL against MRSA USA300MIC: 64 μg/mL against MRSA ATCC 43300MIC: 16 μg/mL against Neisseria gonorrhoeae ATCC 19424 | [ref. 61,ref. 247] |
| Δ1-Tetrahydrocannabidivarol | IC50: 6.9 μM against Staphylococcus aureus ATCC 29213IC50: 6.9 μM against Bacillus cereus IIIM 25IC50: 5.1 μM against Lactococcus lactis MTCC 440IC50: 3.9 μM against Shigella boydii NC-09357IC50: 7.8 μM against Pseudomonas fluorescens MTCC 103IC50: 7.6 μM against Staphylococcus warneri MTCC 4436 | [ref. 246] |
| (±)-4-Acetoxycannabichromene | IC50: 40.3 μM against Leishmania donovani IC50: 4–7.2 μM against Plasmodium falciparum | [ref. 63] |
| (±)-3″-Hydroxy-Δ(4″,5″) cannabichromene | IC50: 24.4 μM against MRSA ATCC 33591IC50: 29.6 μM against Staphylococcus aureus ATCC 29213IC50: 60.5 μM against Candida albicans ATCC 90028IC50: 60.5 μM against Candida krusei ATCC 6258IC50: 57.5 μM against Leishmania donovaniNot active against Escherichia coli, Mycobacterium intracellulare, or Plasmodium falciparum. | [ref. 63] |
| 5-Acetyl-4-hydroxycannabigerol | IC50: 53.4 μM against MRSA ATCC 33591IC50: 10.7 μM against Leishmania donovaniIC50: 6.7–7.2 μM against Plasmodium falciparumNot active against Staphylococcus aureus, Escherichia coli, Mycobacterium intracellulare, or Candida albicans. | [ref. 63] |
| 4-Acetoxy-2-geranyl-5-hydroxy-3-n-pentylphenol | IC50: 6.7 μM against MRSA ATCC 33591IC50: 12.2 μM against Staphylococcus aureus ATCC 29213IC50: 53.4 μM against Candida krusei ATCC 6258IC50: 42.7 μM against Leishmania donovaniNot active against Escherichia coli, Mycobacterium intracellulare, Candida albicans, or Plasmodium falciparum. | [ref. 63] |
| 8-Hydroxycannabinol | IC50: 4.6 μM against Candida albicans ATCC 90028IC50: 30.6 μM against Mycobacterium intracellulareNot active against Escherichia coli. | [ref. 63] |
| 8-Hydroxycannabinolic acid A | IC50: 54 μM against Candida krusei ATCC 6258IC50: 3.5 μM against Staphylococcus aureus ATCC 29213IC50: 54 μM against Escherichia coliNot active against Mycobacterium intracellulare. | [ref. 63] |
| Non-Cannabinoid constituents of Cannabis sativa L. | ||
| 5-Acetoxy-6-geranyl-3-n-pentyl-1,4-benzoquinone | IC50: 15 μg/mL against MRSA ATCC 43300IC50: 13 μg/mL against Leishmania donovaniIC50: 2.6–2.8 μg/mL against Plasmodium falciparum | [ref. 101] |
| Cannflavin A | IC50: 4.5 μg/mL against Leishmania donovani | [ref. 101] |
| Cannflavin B | IC50: 5 μg/mL against Leishmania donovani | [ref. 100] |
| Cannflavin C | IC50: 17 μg/mL against Leishmania donovani | [ref. 101] |
| 6-Prenylapigenin | IC50: 6.5 μg/mL against MRSA ATCC 43300IC50: 20 μg/mL against Candida albicansIC50: 2.0–2.8 μg/mL against Plasmodium falciparum | [ref. 101] |
| Prenylspirodinone | IC50: 49.6 μM against Bacillus thuringiensis MTCC 809 | [ref. 246] |
* BIC50 = The test concentration that prevents 50% biofilm formation compared to control cells. IC50 = The test concentration that causes 50% growth inhibition in comparison to control cells. MBEC = Minimum biofilm eradication concentration is the lowest concentration that completely eradicates preformed biofilm. MBIC = Minimum biofilm inhibitory concentration is the lowest concentration that is required to completely prevent any biofilm formation. MIC = Minimum inhibitory concentration is the lowest concentration that completely inhibits bacterial growth (when no turbidity is observed).
3.3.1. Bacterial Growth Inhibitory Effects of Phytocannabinoids
The minimum inhibitory concentration (MIC) of Δ9-THC and CBD on various Staphylococcus aureus strains, including MRSA and Streptococci species (e.g., Streptococcus pyogenes and Streptococcus. faecalis) was found to be in the range of 1–5 μg/mL [ref. 58,ref. 62,ref. 245,ref. 246]. There was no significant difference between the anti-bacterial effect of Δ9-THC and CBD [ref. 58,ref. 245,ref. 246]. The anti-microbial effect was attenuated by the presence of either serum or blood, suggesting that serum components can bind the compounds and prevent them from acting on the microorganisms [ref. 245]. CBG shows anti-bacterial activity against Gram-positive bacteria, including MSSA, MRSA, and the oral cariogenic Streptococcus mutans at low concentrations similar to CBD [ref. 58,ref. 61,ref. 244,ref. 247]. CBC and CBDA showed a MIC of 1–2 μg/mL against Staphylococcus aureus and Staphylococcus epidermidis [ref. 62,ref. 220]. In these studies, CBDA was less active than CBD [ref. 62]. Cannabichromenic acid (CBCA) caused a rapid reduction in the colony-forming units (CFUs) of a clinical MRSA isolate both during the exponential and stationary growth phase, suggesting a bactericidal activity that is independent of the metabolic state of the bacteria [ref. 254]. None of the phytocannabinoids had any significant anti-bacterial activity against Gram-negative bacteria, such as Escherichia coli, Salmonella typhi, Pseudomonas aeruginosa, and Proteus vulgaris [ref. 61,ref. 62,ref. 220,ref. 245,ref. 247]. This might be due to the inability of these compounds to penetrate the outer membrane of the Gram-negative bacteria [ref. 61], or the outer membrane protects the bacteria from cell death caused by damage to the inner membrane.
3.3.2. Outer Membrane Permeabilization of Gram-Negative Bacteria Sensitizes Them to Phytocannabinoids
Interestingly, CBD and CBG could act on some Gram-negative bacteria (e.g., Escherichia coli, Acinetobacter baumannii, Klebsiella pneumoniae, Pseudomonas aeruginosa) if the outer membrane was permeabilized with the LPS-binding antibiotic polymyxin B [ref. 61,ref. 247]. It was shown that an Escherichia coli ΔbamBΔtolC deletion strain that renders the bacteria hyperpermeable to many small molecules was sensitive to CBG with a MIC of 4 μg/mL, which is in contrast to the parental Escherichia coli wild-type strain that showed a MIC above 128 μg/mL [ref. 61]. Similarly, a lipo-oligosaccharide-deficient Acinetobacter baumannii strain became sensitive to CBG with a MIC of 0.5 μg/mL compared to the parental strain showing a MIC of 64 μg/mL [ref. 61].
3.3.3. Combined Treatment of Phytocannabinoids with Antibiotics
No synergistic or antagonistic effects of CBD were observed on MRSA strain USA300 when combined with different conventional antibiotics, such as clindamycin, ofloxacin, meropenem, tobramycin, methicillin, teicoplanin, and vancomycin [ref. 62]. These authors concluded that the membrane-perturbing effect of CBD was not sufficient to enhance the uptake of conventional antibiotics [ref. 62]. However, Wassmann et al. [ref. 251] observed that CBD could reduce the MIC value of bacitracin against several Gram-positive bacteria, including Staphylococcus species, Listeria monocytogenes, and Enterococcus faecalis. The simultaneous use of CBD and bacitracin on MRSA USA300 resulted in the formation of multiple septa during cell division, appearance of membrane irregularities, reduced autolysis, and decreased membrane potential [ref. 251]. The combined CBD/bacitracin treatment did not affect the growth of the Gram-negative bacteria Pseudomonas aeruginosa, Salmonella typhimurium, Klebsiella pneumoniae, and Escherichia coli [ref. 251].
3.3.4. Phytocannabinoids Also Act on Persister Cells and Do Not Induce Drug Resistance
CBG was found to be active against MRSA persister cells, which are dormant, non-dividing bacteria [ref. 61]. This trait is therapeutically important, since many antibiotics require cell division to be effective, and they are frequently unable to eradicate persister cells that usually recover after antibiotic withdrawal [ref. 255,ref. 256,ref. 257]. Another obstacle of antibiotic therapy is the development of drug resistance, a frequent reason for treatment failure [ref. 258]. Farha et al. [ref. 61] attempted to develop CBG-resistant bacteria in the hopes of finding the target molecules. Despite rechallenging the MRSA with 2x and 16x MIC concentration of CBG, they were unable to get any spontaneously CBG-resistant mutants [ref. 61]. Similarly, MRSA that had been daily exposed to sub-lethal concentration of CBD for 20 days were still sensitive to CBD [ref. 247]. The authors of these two studies [ref. 61,ref. 247] concluded that CBD and CBG do not induce drug resistance. However, it should be noted that following exposure to CBD or CBG, the surviving growth-arrested bacteria could regain growth after withdrawal of the drug.
3.3.5. Therapeutic Anti-Microbial Potential of Phytocannabinoids
The hemolytic activity of CBD and CBG was found to be 256 μg/mL and 32 μg/mL, respectively, which is far above the MIC of 1–4 μg/mL for MRSA [ref. 61,ref. 247]. Additionally, the hemolytic activity of CBDA was found to be above 32 μg/mL [ref. 62]. This makes phytocannabinoids potential drugs that can act within a reasonable therapeutic window.
Farha et al. [ref. 61] observed that treating MRSA-infected mice with a high dose of 100 mg/kg CBG could reduce the bacterial burden in the spleen by a 2.8 log10 of CFU. Blaskovich et al. [ref. 247] tried various CBD-containing ointment formulations that could reduce a 2–3 log10 of CFU of MRSA inoculated on porcine skin after 1 h and a reduction of more than 5 log10 of CFU after a 24 h incubation. CBD, however, failed to significantly reduce the bacterial load of MRSA ATCC 43300 in a thigh infection mouse model [ref. 247].
3.3.6. Anti-Biofilm Activities of Phytocannabinoids
Biofilms are communities of bacteria embedded in an extracellular matrix that have attached to a biotic surface (e.g., lung tissue, gastrointestinal tract, nasal mucosa, inner ear) or an abiotic surface (e.g., medical devices, such as catheters, heart valves, stents, prostheses) [ref. 259]. The majority of infectious diseases involve bacterial biofilms that are usually difficult to eradicate due to reduced antibiotic sensitivity [ref. 259,ref. 260]. Several studies show that CBD and CBG can prevent biofilm formation of various Gram-positive bacteria (e.g., MSSA, MRSA, Streptococcus mutans) [ref. 61,ref. 243,ref. 247]. The extent of anti-biofilm activity of CBD and CBG against these bacteria correlated with their anti-bacterial activity [ref. 61,ref. 243,ref. 244,ref. 247]. In most cases, a similar concentration of these compounds was required to achieve both effects, suggesting that some of the anti-biofilm effect is caused by the anti-bacterial activity [ref. 61,ref. 243,ref. 244]. Moreover, CBD was found to be able to eradicate preformed MSSA and MRSA biofilms with a minimum biofilm eradication concentration (MBEC) of 1–4 μg/mL, indicating that CBD can penetrate the biofilms and act on the biofilm-embedded bacteria [ref. 247]. Some cannabinoids (e.g., CBD, CBG, CBC, and CBN) were shown to reduce the bacterial content of dental plaques in an in vitro assay where dental plaques were spread on agar plates coated with the cannabinoids [ref. 261]. The anti-biofilm activity of the cannabinoids has significant clinical importance, since the bacteria-embedded bacteria frequently show antibiotic resistance, and some antibiotics are unable to penetrate through the extracellular matrix of the biofilms [ref. 259,ref. 262,ref. 263].
3.3.7. Anti-Fungal Biofilm Activities of Phytocannabinoids
CBD barely affects the viability of Candida albicans with a MIC above 50–100 µg/mL [ref. 247,ref. 253], but it reduces biofilm formation with a biofilm inhibitory concentration 50 (BIC50) at 12.5 µg/mL and a MBIC90 of 100 µg/mL [ref. 253]. CBD reduced the metabolic activity of preformed Candida albicans biofilms by 50–60% at 6.25 µg/mL with no further reduction at higher concentrations, even at 100 µg/mL [ref. 253]. The morphology of the Candida albicans biofilm becomes altered in the presence of CBD. While the hyphal form was predominant in control biofilms, the CBD (25 µg/mL)-treated biofilms appeared in clusters mostly in yeast and pseudohyphal forms [ref. 253]. CBD caused a dose-dependent reduction in the cell wall chitin content and the intracellular ATP level, while increasing the intracellular reactive oxygen species (ROS) levels [ref. 253]. Gene expression studies showed that after a 24 h incubation with 25 µg/mL CBD, there is a significant downregulation of: ADH5 (Alcohol dehydrogenase 5), involved in extracellular matrix production; BIG1, required for synthesis of the extracellular matrix component β-1,6-glucan; ECE1 (extent of cell elongation protein 1), involved in biofilm formation; EED1, involved in filamentous growth; CHT1 and CHT3 chitinases, involved in the remodeling of chitin in the fungal cell wall; and TRR1 (thioredoxin reductase) with anti-oxidant properties. On the other hand, a significant upregulation of YWP1 (yeast-form wall protein 1) which is expressed predominantly in the yeast form, was observed [ref. 253]. These changes in gene expression might explain, at least in part, the reduced biofilm mass of Candida albicans in the presence of CBD and the increase in oxidative stress [ref. 253].
3.3.8. Anti-Viral Activities of Phytocannabinoids
There are some lines of evidence for an anti-viral activity of phytocannabinoids [ref. 60,ref. 264]. Some phytocannabinoids, especially Δ9-THC and CBD, bind to the Mpro protease of SARS-CoV-2, which plays a role in viral replication [ref. 60,ref. 264]. CBGA and CBDA were found to be allosteric and orthosteric ligands for the spike protein of SARS-CoV-2 and prevented infection of human epithelial cells by a pseudovirus expressing the SARS-CoV-2 spike protein [ref. 265]. Phytocannabinoids might indirectly relieve the disease progress of COVID-19 patients through their anti-inflammatory properties [ref. 266]. However, CBD failed to alter the clinical disease development of COVID-19 when given at a daily dose of 300 mg for 14 days [ref. 267]. Additionally, caution should be taken into account due to the immunosuppressive activities of phytocannabinoids that can prevent proper anti-viral immune responses [ref. 268]. Notably, the use of Cannabis was increased in U.S. and Canada by 6–8% during the COVID-19 pandemic in comparison to the pre-pandemic period [ref. 269], with a special increase among people with mental health [ref. 270]. Vulnerability to COVID-19 was correlated with genetic liability to Cannabis use disorder (CUD) [ref. 271].
3.4. Some Mechanistic Insight into the Anti-Bacterial Activity of Phytocannabinoids
The ability of phytocannabinoids such as CBD and CBG, to kill MRSA, NorA-overexpressing Staphylococcus aureus, vancomycin-resistant Staphylococcus aureus (VRSA), vancomycin-resistant enterococci (VRE) to a similar extent as the respective antibiotic-sensitive strains [ref. 58,ref. 245,ref. 247], suggests that its action mechanism is not hindered by the common antibiotic-resistance mechanisms. Thus, phytocannabinoids can be used as an alternative drug or an antibiotic adjuvant for infectious diseases caused by drug-resistant Gram-positive bacteria.
3.4.1. CBD and CBG Target the Cytoplasmic Membrane, Increase Membrane Permeability, and Reduce Metabolic Activity
There is evidence that CBD and CBG act by targeting the cytoplasmic membrane of the Gram-positive bacteria [ref. 61,ref. 247]. Exposure of MSSA and MRSA to CBD or CBG caused a dose-dependent increase in the fluorescence of the potentiometric probe 3,3′-dipropylthiadicarbocyanine iodide [DiSC3(5)], suggesting a CBG-induced membrane depolarization [ref. 61,ref. 247]. CBD inhibited protein, DNA, RNA, and peptidoglycan synthesis in a Staphylococcus aureus strain when using concentrations close to the MIC [ref. 247]. At sub-MIC levels, CBD inhibited lipid synthesis [ref. 247]. CBG was found to inhibit the enzyme enoyl acyl carrier protein reductase (InhA) [ref. 272], which is involved in type II fatty acid biosynthesis in Mycobacterium tuberculosis. The rapid uptake of the SYTOX green dye into Staphylococcus aureus and Bacillus subtilis by CBD at MIC, suggests that CBD causes an increase in membrane permeability [ref. 247].
CBG prevents the growth of oral cariogenic Streptococcus mutans in a concentration and bacterial cell density manner [ref. 243]. At a MIC of 2.5 μg/mL, CBG exhibited a bacteriostatic effect on Streptococcus mutans, while at 2x MIC and 4x MIC, a bactericidal activity was observed [ref. 243]. CBG treatment was found to alter the morphology of Streptococcus mutans and cause intracellular accumulation of membrane-like structures [ref. 243]. CBG induced an immediate membrane hyperpolarization, followed by increased uptake of propidium iodide, suggesting increased membrane permeabilization [ref. 243]. At the same time, Laurdan incorporation into the membranes was reduced in a dose-dependent manner [ref. 243], indicative of a more rigid membrane structure. The metabolic activity was decreased in a dose-dependent manner, which might contribute to the growth inhibitory effect [ref. 243].
3.4.2. CBD Inhibits the Release of Membrane Vesicles from Escherichia coli
Kosgodage et al. [ref. 250] observed that CBD inhibits the release of membrane vesicles from the Gram-negative Escherichia coli VCS257, while having negligible effect on the membrane vesicle release from the Gram-positive Staphylococcus aureus subsp. aureus Rosenbach. Membrane vesicles participate in inter-bacterial communication by the transfer of cargo molecules and virulence factors [ref. 273]. CBD was found to enhance the anti-bacterial effect of erythromycin, rifampicin, and vancomycin against the tested Escherichia coli strain [ref. 250].
3.4.3. CBG Reduces the Expression of Biofilm and Quorum Sensing-Related Genes in Streptococcus mutans
CBG inhibited sucrose-induced biofilm formation by Streptococcus mutans with a minimum biofilm inhibitory concentration (MBIC) of 2.5 μg/mL [ref. 243]. Higher concentrations (10 μg/mL) of CBG were required to reduce the metabolic activity of preformed Streptococcus mutans biofilms [ref. 243]. CBG reduced the expression of various biofilm-related genes (e.g., gtfB, gtfC, gtfD, ftf, gbpA, gbpA, brpA, wapA) with concomitant reduction in the production of extracellular polymeric substances (EPS) [ref. 243]. The quorum sensing-related genes comE, comD, and luxS were downregulated by CBG, while no effect was observed on the gene expression of the stress-associated chaperones groEL and dnaK [ref. 243]. Moreover, CBG induced reactive oxygen species (ROS) production in Streptococcus mutans, which might be related to the reduced expression of the oxidative stress defense genes, sod and nox [ref. 243]. Thus, CBG has specific anti-biofilm activity unrelated to its membrane-acting effect. This conclusion is further supported by the study of Aqawi et al. [ref. 248] showing that CBG inhibited quorum sensing, bacterial motility, and biofilm formation of the marine Gram-negative Vibrio harveyi without affecting the planktonic growth.
3.4.4. CBG and HU-210 Inhibit Quorum Sensing in Vibrio harveyi
Quorum sensing is an inter-bacterial communication system mediated by secreted autoinducers that interact with their respective receptors, resulting in the activation of a signal transduction cascade that alters the gene expression repertoire in a cell-density-dependent manner [ref. 274]. CBG prevented the bioluminescence induced by the master quorum sensing regulator LuxR of Vibrio harveyi at a concentration of 1 µg/mL [ref. 248]. Using a ΔluxM, ΔlusS Vibrio harveyi mutant that does not produce autoinducers AI-1 and AI-2, CBG was found to prevent the signals delivered by exogenously added autoinducers, with a more profound inhibitory effect on the AI-2-induced than on the AI-1-induced bioluminescence [ref. 248]. Further studies show that CBG prevented the expression of several quorum sensing genes in Vibrio harveyi, including luxU, luxO, qrr1–5, and luxR, which can explain the inhibitory effect of CBG on LuxR-mediated bioluminescence [ref. 248]. Altogether, these data demonstrate that CBG can interfere with bacterial quorum sensing.
The synthetic cannabinoid HU-210, which is a dimethylheptyl analog of Δ8-THC (Figure 1) and acts as a high-affinity CB1 and CB2 agonist [ref. 275,ref. 276], has been shown to inhibit quorum sensing in the Vibrio harveyi AI-1−, AI-2+ BB152 mutant, but it had barely any effect on the wild-type bacteria or the AI-1+, AI-2− MM30 mutant [ref. 249]. This suggests that HU-210 specifically antagonizes the AI-2 pathway [ref. 249]. The concentration of HU-210 required to achieve the anti-quorum sensing activity was relatively high (20–200 µg/mL) [ref. 249], which is 2–3 magnitudes higher than that of CBG [ref. 248]. HU-210 prevented biofilm formation of the AI-1−, AI-2+ BB152 mutant with a BIC50 of 2 µg/mL and MBIC90 of 200 µg/mL, while no significant effect was seen on biofilm formation by the wild-type bacteria or the AI-1+, AI-2− MM30 mutant [ref. 249]. However, the motility of Vibrio harveyi was reduced in all three strains at both 20 and 200 µg/mL HU-210 [ref. 249]. Gene expression studies showed that HU-210 at a concentration of 2 µg/mL reduced the expression of the master regulator luxR in both wild-type and AI-1−, AI-2+ BB152 strain, while it had no effect on the AI-1+, AI-2− MM30 Vibrio harveyi mutant strain [ref. 249]. The luxM gene that encodes for AI-1 was upregulated by HU-210 [ref. 249].
4. Endocannabinoids
The endocannabinoid system (ECS) modulates many physiological processes, including the cardiovascular, gastrointestinal and immune systems, pain, learning, memory, perception, mood, appetite, metabolism, emotions, and sleep [ref. 22,ref. 112,ref. 113,ref. 277,ref. 278,ref. 279,ref. 280,ref. 281,ref. 282,ref. 283,ref. 284,ref. 285]. The bioactive endocannabinoid lipid mediators have potent anti-inflammatory activities [ref. 286,ref. 287,ref. 288,ref. 289,ref. 290,ref. 291]. In addition, they promote neural progenitor cell proliferation and differentiation, and have neuroprotective effects [ref. 20,ref. 292,ref. 293,ref. 294]. The effect on neural cell proliferation is mediated by both the CB1 and CB2 receptors [ref. 293,ref. 295,ref. 296].
4.1. The Endocannabinoid System
The endocannabinoid system is composed of: (1) the lipid active endogenous ligands N-arachidonoylethanolamine (anandamide; AEA) and 2-arachidonoylglycerol (2-AG); (2) their biosynthetic enzymes (e.g., diacylglycerol lipases (DAGL), N-acyl-phosphatidylethanolamine phospholipase D-like esterase (NAPE-PLD), and Ca2+-dependent and Ca2+-independent N-acetyltransferases); (3) their degradative enzymes (e.g., fatty acyl amide hydrolase (FAAH) and monoacylglycerol lipase (MAGL)); and (4) the CB1 and CB2 cannabinoid receptors [ref. 15,ref. 297,ref. 298]. The precursors of endocannabinoids (e.g., N-acyl-phosphatidylethanolamine (NAPE) and phosphatidylinositol-4,5-bisphosphate (PIP2)) are present in the lipid membranes, and the endocannabinoids are produced upon demand, usually after activation of certain G-protein-coupled receptors (GPCRs) and in response to an increase in the intracellular calcium levels [ref. 299,ref. 300,ref. 301,ref. 302].
4.2. The Production of AEA and 2-AG
The production of endocannabinoids requires one or two enzymatic steps, followed by their release into the extracellular space. AEA is usually produced from N-arachidonoyl-phosphatidylethanolamine phospholipid, and 2-AG is produced primarily from membrane phospholipid 1-stearoyl-2-arachidonoyl-sn-glycerol [ref. 297]. The synthesis of 2-AG involves two steps: first, the hydrolysis of its precursor phospholipid by a phospholipase (PLCβ, PLCγ2, or PLCε), followed by further cleavage by diacylglycerol lipase (DAGL) [ref. 303,ref. 304,ref. 305]. The biosynthesis of these endocannabinoids occurs in areas of the brain functionally related to cognitive processes, motivation, and movement control [ref. 306,ref. 307]. 2-AG was found to be present at 170 times higher concentrations than AEA in brain lysate [ref. 308]. While AEA was initially detected in the brain [ref. 135] and 2-AG in the canine gut [ref. 309], today it is known that these host-derived endocannabinoid lipid hormones are found in various peripheral tissues (e.g., the intestine) and in the serum, and produced by certain immune cells [ref. 23,ref. 290,ref. 309,ref. 310,ref. 311,ref. 312,ref. 313,ref. 314,ref. 315,ref. 316,ref. 317]. For instance, lipopolysaccharides induced the production of AEA in adipose tissue macrophages [ref. 318]. T and B cells produce elevated levels of 2-AG upon activation [ref. 290]. Astrocytes were found to produce AEA, as well as homo-γ-linolenoylethanolamine (HEA), docosatetraenoylethanolamine (DEA), oleoylethanolamine (OAE), and palmitoylethanolamine (PEA) [ref. 319].
4.3. The Circulating Levels of AEA and 2-AG
The circulating endocannabinoid levels are affected by various factors, and under physiological conditions, the AEA serum level was found to be between 1 to 5 nM, and the 2-AG serum level between 10–500 nM [ref. 316,ref. 320]. Physical exercise mobilizes endocannabinoids, which could contribute to the analgesic and mood-elevating effects of exercise [ref. 316]. The circulating levels of 2-AG show a circadian rhythm that gets altered when sleep is disrupted [ref. 316,ref. 320]. CBD inhibits the degradation of AEA and 2-AG, which is associated with the anti-inflammatory and anti-oxidative activities [ref. 321].
4.4. Endogenous Receptors for AEA and 2-AG
AEA and 2-AG act as agonists of the CB1 and CB2 receptors [ref. 135,ref. 322,ref. 323,ref. 324,ref. 325]. While 2-AG binds with high affinity to CB1 and CB2 cannabinoid receptors, AEA binds with low affinity to these receptors [ref. 323,ref. 324]. Although phytocannabinoids and endocannabinoids bind to the same CB1 and CB2 receptors, their chemical structure is quite different [ref. 297] (Figure 1 and Figure 3). Both AEA and 2-AG have an alkyl-amide (alkamide) chemical structure, while cannabinoids are terpenophenolic compounds.
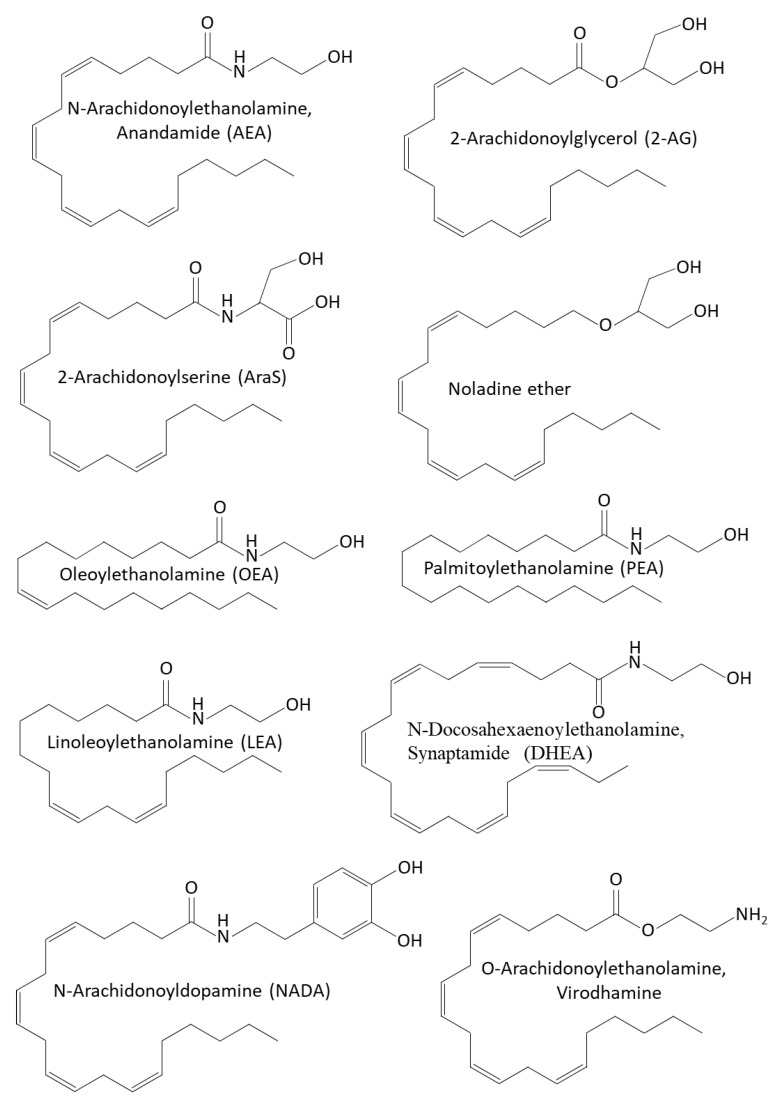
In addition to acting on CB1 and CB2, AEA activates the ionotropic TRPV1 channel, resulting in the opening of the ion channel and Ca2+ influx [ref. 312,ref. 326,ref. 327,ref. 328,ref. 329,ref. 330,ref. 331], the G-protein-coupled receptor GPR55 [ref. 332,ref. 333], and the cation channel TRPA1 [ref. 334], while it inhibits the TRPM8 channel [ref. 334]. In addition, AEA activates PPARγ, and 2-AG activates PPARα [ref. 335]. The vasodilation action of AEA was found to be mediated via activation of TRPV1 [ref. 336]. Endocannabinoids activating TRPV1 have been included in the endovanilloid system [ref. 337,ref. 338,ref. 339]. Recent studies suggest that potassium channels are also the targets of endocannabinoids [ref. 340].
In the brain, endocannabinoids serve as retrograde synaptic messengers [ref. 299,ref. 341]. They are released from postsynaptic neurons and inhibit the release of presynaptic neurotransmitters, such as glutamate and gamma-aminobutyric acid (GABA) by binding to the CB1 receptor and TRPV1 expressed in the presynaptic terminals [ref. 299,ref. 342,ref. 343]. This has led to the hypothesis that endocannabinoids regulate over-excitability and promote synaptic homeostasis [ref. 344]. Endocannabinoids differ from the classical neurotransmitters in that they are not stored in vesicles but are released immediately after their production.
The solubility of endocannabinoids is low in water, raising the question of how AEA diffuses through the synaptic cleft [ref. 345]. There is evidence that AEA can interact with cholesterol and ceramide, which are required for their insertion into and transport through the membrane [ref. 345,ref. 346,ref. 347]. In the brain, the lipid-binding protein α-synuclein is involved in the transport of arachidonic acid [ref. 348]. Fatty acid binding proteins have been shown to be intracellular carriers of AEA [ref. 349].
Another communication system that exists between neurons is the release of lipid-based transport systems such as exosomes from neurons following a synaptic response, that are taken up by neighboring cells [ref. 350,ref. 351]. Gabrielli et al. [ref. 352] observed that endocannabinoids are secreted on extracellular membrane vesicles. In this study, extracellular vesicles secreted by microglial cells were found to carry AEA on their surface that was able to stimulate the CB1 receptor expressed on neurons and inhibit presynaptic transmission [ref. 352]. Microglial cells release endocannabinoids at much higher levels than neurons and astrocytes [ref. 319,ref. 353,ref. 354] and are thought to play a role in regulating the synaptic activity by a process termed gliotransmission, which functions to bridge the non-synaptic inter-neuronal communication [ref. 355].
4.5. Other Endocannabinoids and Endocannabinoid-like Compounds
Other endocannabinoids include the oleoyl- and palmitoyl-ethanolamines (OEA and PEA) that affect intestinal permeability by acting on TRPV-1 and PPARα [ref. 356,ref. 357], and 2-AG-ether and O-arachidonoylethanolamine (virodhamine) [ref. 22,ref. 358] (Figure 3). PEA is produced by neurons, microglia, and astrocytes in the central nervous system [ref. 359,ref. 360] where it plays an important role in neuroprotection [ref. 361,ref. 362]. Moreover, it was shown to have both anti-nociceptive and anti-inflammatory activities [ref. 363,ref. 364,ref. 365,ref. 366]. Immune cells release PEA that activates the CB2 receptor, resulting in downregulation of the inflammatory processes [ref. 367,ref. 368]. PEA, which is synthesized along with AEA, potentiates the action of AEA by increasing receptor affinity or reducing the degradation of AEA by FAAH [ref. 357,ref. 369,ref. 370,ref. 371]. The study of Lo Verme et al. [ref. 372] showed that PPARα was required for the anti-inflammatory effect of PEA. Borrelli et al. [ref. 365] observed that PEA alleviates the inflammation in a murine colitis model through acting on CB2, GPR55, and PPARα. OEA acts on PPARα and is secreted in the proximal intestine where it controls appetite, exhibits anti-inflammatory properties, and stimulates lipolysis and fatty acid oxidation [ref. 373,ref. 374,ref. 375,ref. 376].
The endocannabinoid noladin ether acts on CB2 and inhibits the intracellular effector adenylyl cyclase [ref. 377]. The endocannabinoid virodhamine, which is composed of arachidonic acid and ethanolamine joined by an ester linkage, is a partial agonist with an antagonist activity on CB1, while being a full agonist on CB2 [ref. 378]. At low concentrations, virodhamine activates GPR55, while at high concentrations it acts as an antagonist [ref. 379]. The endocannabinoid N-arachidonoyl-dopamine (NADA), which is highly expressed in the striatum, hippocampus, and cerebellum, activates TRPV1, induces the release of substance P and calcitonin gene-related peptide from dorsal spinal cord slices, and enhances hippocampal paired-pulse depression [ref. 380]. NADA and its epoxide metabolites also act as an agonist for the CB1 and CB2 receptors and show anti-inflammatory activities [ref. 337,ref. 381,ref. 382,ref. 383]. Other dopamine-related endocannabinoids include N-oleoyldopamine (OLDA), N-palmitoyldopamine (PALDA), and N-stearoyldopamine (STEARDA) [ref. 384]. OLDA is only a weak ligand of CB1, but it induced calcium influx, reduced the latency of paw withdrawal from a radiant heat source, and produced nocifensive behavior [ref. 384].
N-Arachidonoyl-L-serine (AraS) is an endogenous bioactive lipid found both in the central nervous system (CNS) and in the periphery, with a similar structure and physiological functions as AEA [ref. 385,ref. 386] (Figure 3). It possesses vasoactive, pro-angiogenic, pro-neurogenic, and neuroprotective properties [ref. 386,ref. 387,ref. 388]. Since AraS binds weakly to CB1 and CB2, it is not classified as an endocannabinoid, but rather has been coined as an “endocannabinoid-like” substance [ref. 386]. The pro-angiogenic activity of AraS is achieved by activation of GPR55 [ref. 387]. Moreover, AraS stimulates phosphorylation of MAPK and Akt protein kinases [ref. 385].
4.6. Anti-Microbial Activities of Endocannabinoids and Endocannabinoid-like Compounds
The anti-microbial effect of endocannabinoids depends on the strain studied and the endocannabinoid used [ref. 16,ref. 17,ref. 18,ref. 389,ref. 390] (Table 2). Among the tested organisms, Streptococcus salivarius, Bacteroides fragilis, and Enterococcus faecalis were the most susceptible bacteria to AEA and N-Linoleoylethanolamine (LEA) [ref. 390]. MSSA and MDRSA become immediately growth arrested by AEA, an effect that was transient and relieved upon time [ref. 16]. On the other hand, the growth of Lactobacillus gasseri species becomes enhanced by LEA and OEA [ref. 390].
Table 2: Anti-microbial activities of endocannabinoids and endocannabinoid-like compounds.
| Endocannabinoids | Anti-Microbial Activity | References |
|---|---|---|
| Anandamide (AEA) | [ref. 16,ref. 17,ref. 18,ref. 389,ref. 390] | |
| N-Arachidonoyl-L-serine (AraS) | [ref. 17,ref. 18,ref. 389] | |
| 2-Arachidonoylglycerol (2-AG) | [ref. 389] | |
| N-Linoleoylethanolamine (LEA) | [ref. 390] | |
| Oleoylethanolamine (OEA) | [ref. 390] | |
| Palmitoylethanolamine (PEA) | [ref. 390] |
4.6.1. AEA and AraS Exert Bacteriostatic Activity on Both Drug-Sensitive and Drug-Resistant Staphylococcus aureus
Feldman et al. [ref. 18] observed that the MIC of AEA toward three MRSA species (MRSA ATCC 33592, MRSA ATCC 43300, and a MRSA clinical isolate) was above 256 µg/mL. AraS had a MIC of 16 and 128 µg/mL on MRSA ATCC 33592 and MRSA ATCC 43300, respectively, and a MIC above 256 µg/mL for the clinical MRSA isolate [ref. 18]. A kinetic study of AEA on a multidrug-resistant Staphylococcus aureus (MDRSA) clinical isolate and the MSSA ATCC 25923 strain showed that AEA caused a transient bacteriostatic effect that was overcome with time [ref. 16]. The bacteriostatic effect of AEA was independent of the drug-resistant phenotype [ref. 16]. Further analysis showed that AEA inhibited cell division just prior to daughter cell separation [ref. 16]. Gene expression studies showed that AEA reduced the expression of some autolysin genes, which might in part contribute to the growth arrest [ref. 16]. AEA altered the membrane structure of the MDRSA and caused an immediate membrane depolarization that recovered with time [ref. 16]. Both AEA and AraS reduced the hydrophobicity index of MRSA at a concentration of 16 µg/mL [ref. 18].
4.6.2. AEA and AraS Sensitize Drug-Resistant Staphylococcus aureus to Antibiotics
Importantly, it was observed that AEA and AraS sensitize MRSA and MDRSA strains to various antibiotics, including β-lactam antibiotics (ampicillin and methicillin), gentamicin, tetracycline, and norfloxacin [ref. 16,ref. 17]. For instance, the MIC of ampicillin against MRSA ATCC 33592 and ATCC 43300 was 128 and 256 µg/mL, respectively, but in the presence of 8–16 µg/mL AEA, it was reduced to 8 µg/mL [ref. 17]. The MIC of gentamicin against MRSA ATCC 33592 was 128 µg/mL, but in the presence of 8 µg/mL AEA, it was reduced to 4 µg/mL [ref. 17]. Treating a MDRSA clinical isolate with 50 µg/mL AEA reduced the MIC of methicillin from above 500 µg/mL to 50 µg/mL [ref. 16]. AEA was found to prevent drug efflux, resulting in intracellular drug accumulation, which might explain, at least in part, the sensitization of the bacteria to antibiotics [ref. 16]. Gene expression analysis shows that AEA reduces the expression of some efflux pump genes, including norB, norC, mepA, kdpA, and opp1C in MDRSA [ref. 16], but it is likely that the alterations in the membrane structure caused by AEA also contribute to intracellular drug retention.
It is notable that the sensitization of MRSA to methicillin takes place even when bacterial growth is inhibited by AEA [ref. 16], suggesting that the anti-bacterial effect of methicillin and other β-lactams does not require cell division as previously documented when used as a single agent [ref. 391,ref. 392]. Indeed, FtsZ inhibitors that arrest bacterial cell growth, also sensitize drug-resistant Staphylococcus aureus to β-lactam antibiotics, which was related to membrane relocalization of penicillin-binding proteins (PBPs) [ref. 393]. Further studies are required to fully understand the antibiotic-sensitization mechanisms of AEA and AraS.
4.6.3. AEA and AraS Exhibit Anti-Biofilm Activity against Drug-Sensitive and Drug-Resistant Staphylococcus aureus
AEA and AraS prevent biofilm formation of MRSA and MDRSA with a maximum effect at 12.5–35 µg/mL [ref. 16,ref. 18]. AEA and AraS had a rather weak effect on preformed biofilm of MRSA and MDRSA, where concentrations as high as 64 µg/mL were required to eradicate 50% of the biofilms after a 24 h incubation [ref. 16,ref. 18]. The simultaneous treatment of the MRSA and MDRSA strains with endocannabinoids and antibiotics significantly lowered the effective dose of the two compounds [ref. 16,ref. 17]. For instance, the MBICs of AEA and ampicillin on MRSA ATCC 33592 were, respectively, 33.8 and 128 µg/mL, while in combination, 8 µg/mL of each compound was required for inhibiting biofilm formation [ref. 17]. When combining the sub-MBIC concentration 3.125 µg/mL of AEA with 50 µg/mL norfloxacin, which, as a single agent, had no anti-biofilm effect, an 80% reduction in biofilm formation by MDRSA was observed [ref. 16]. A 90% reduction in preformed MDRSA biofilm was observed when 50 µg/mL of AEA was combined with 50 µg/mL methicillin, which is the synergistic condition required for killing the bacteria [ref. 16]. The latter observation shows that the combination of AEA with antibiotics is also effective against biofilm-embedded bacteria.
Gene expression studies showed that AEA reduced the expression of the regulatory RNAIII and the virulence gene α-helical phenol-soluble modulin (psmα) in MDRSA [ref. 16]. Additionally, the genes fnbB, hla, and hld encoding for the virulence factors fibronectin binding protein, α-hemolysin and δ-hemolysin (δ-toxin), respectively, were downregulated by AEA [ref. 16]. As Psmα plays a central role in Staphylococcus aureus biofilm formation by stabilizing the biofilms through amyloid formation [ref. 394,ref. 395,ref. 396], the inhibition of its expression might be one mechanism for the anti-biofilm effect of AEA (Figure 4).
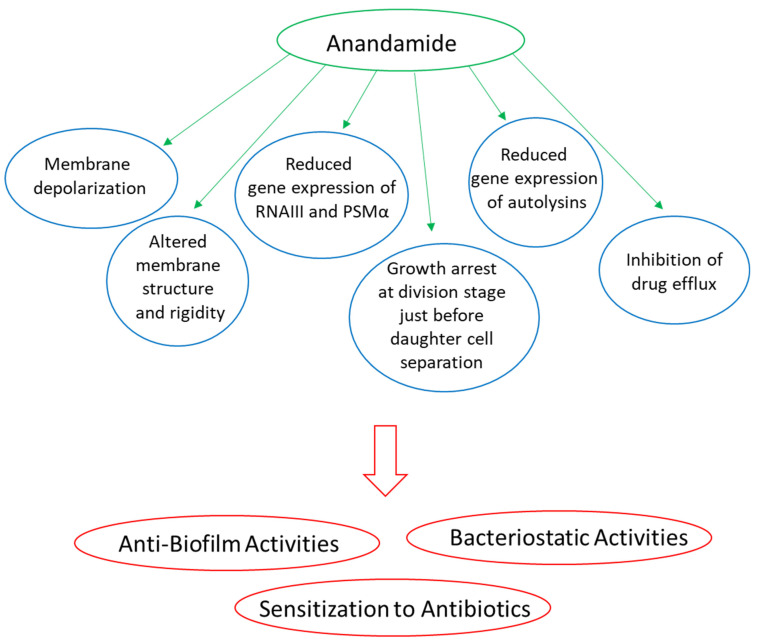
4.6.4. AEA and AraS Inhibit Yeast-Hypha Transition of Candida albicans and Prevent Adhesion of Candida albicans Hyphae to Epithelial Cells
AEA and AraS were found to inhibit yeast-hypha transition of Candida albicans at 125 and 250 µg/mL [ref. 389]. At 50 µg/mL, there was only a partial inhibition on the yeast-hypha transition, but this concentration was sufficient to prevent hyphal extension [ref. 389]. Importantly, Candida albicans hyphae that have been exposed to AEA at 50 µg/mL and higher concentrations showed strong reduction in their ability to adhere to the HeLa cervical epithelial carcinoma cells [ref. 389]. AraS-treated Candida albicans hyphae showed deficient adherence to HeLa cervical carcinoma cells similar to AEA-treated fungi, while 2-AG treatment had only a minor effect at the concentrations analyzed (up to 250 µg/mL) [ref. 389]. None of the endocannabinoids affected the adherence of the Candida albicans hyphae to polystyrene tissue culture plates within the first hour of incubation, while AraS and 2-AG, but not AEA, reduced the biofilm mass formed on the polystyrene tissue culture plates after a 24 h incubation [ref. 389].
Gene expression studies showed that AEA increased the expression of NRG1, which is a transcriptional repressor of filamentous growth, but reduced the expression of the hyphal cell wall protein 1 (HWP1), the Agglutinin-like protein 3 (ALS3), the Hypha-specific G1 cyclin-related protein 1 (HGC1), the Ras-like protein 1 (RAS1), the enhanced filamentous growth protein 1 (EFG1), the cell surface hydrophobicity-associated protein CSH1, and the extent of cell elongation protein 1 (ECE1). The combined effect of AEA on the expression of these genes might cumulate in the observed effects of AEA on Candida albicans adherence and hyphal growth (Figure 5).
4.7. Dialog between the Gut Microbiota and the Endocannabinoid System
An intercommunication system has been found to exist between the gut microbiota and the endocannabinoid system [ref. 285,ref. 298,ref. 397,ref. 398,ref. 399,ref. 400,ref. 401,ref. 402]. The gut microbiota, representing more than 100 trillion microorganisms, including at least 1000 distinct species, lives in symbiosis with the host and assists in controlling the metabolic health of the host by degrading nutrients that the host is unable to digest and by providing a whole battery of small signaling molecules, metabolites, and nutrients beneficial for the host physiology [ref. 298,ref. 403]. The gut microbiome differs from individual to individual, and the composition of the microbiota is believed to affect various metabolic disorders, such as obesity, hyperglycemia, and dyslipidemia, which are risk factors for type 2 diabetes, hepatosteatosis, and arteriosclerosis [ref. 404]. Reduced diversity of gut microbiota has been linked with various pathophysiological conditions, such as depression, schizophrenia, neurological disorders, and chronic fatigue [ref. 405,ref. 406,ref. 407,ref. 408,ref. 409]. Microbiota can affect the endocannabinoid system and the nervous system, and vice versa; the nervous system and the endocannabinoids can influence the enteric microbiota composition [ref. 112,ref. 285,ref. 390,ref. 397,ref. 410,ref. 411].
4.7.1. The Relationship between Gut Microbiota, the Endocannabinoid System, and Depression
The effect of the gut microbiota on depressive-like behaviors in mice was found to be mediated by the endocannabinoid system [ref. 411]. These authors showed that the transfer of microbiota from stress-induced depressive mice to naïve unstressed hosts induced a depressive-like state in the recipients. This was accompanied by a reduced adult hippocampal neurogenesis that was related to decreased hippocampal 2-AG levels and deficient CB1-mediated activation of the mTOR signaling pathways [ref. 411]. The detrimental effects on hippocampal neurogenesis could be restored by a MAGL inhibitor that prevents 2-AG degradation, addition of the 2-AG precursor arachidonic acid to the diet, or by complementation with Lactobacillus plantarumWJL [ref. 411]. The microbiota from the stress-induced depressive mice showed an increase in Ruminococcaceae and Porphyromonodaceae species, with a decrease in Lactobacillaceae [ref. 411]. The complementation with Lactobacillus plantarumWJL restored hippocampal 2-AG to normal levels, as well as increased the levels of AEA, n-3, and n-6 polyunsaturated fatty acids (PUFAs) [ref. 411]. Lactobacillus species can regulate fatty acid metabolism, absorption, and fatty acid composition of the host [ref. 412,ref. 413], which in turn affects the endocannabinoid system [ref. 411]. Rousseaux et al. [ref. 414] observed that oral administration of Lactobacillus acidophilus induced the expression of both the µ1 opioid receptor and CB2 receptor on colon epithelial cells, resulting in reduced abdominal pain in a rat colorectal distension model.
4.7.2. Association between Gut Microbiota, PEA, and Anhedonia/Amotivation
PEA was shown to mediate the association between gut microbial diversity and anhedonia/amotivation [ref. 410]. Increased serum levels of PEA were associated with anti-depressive effects [ref. 357,ref. 415], while increased stool levels of PEA, indicative of increased excretion of PEA, were associated with alterations in synaptic plasticity, learning, and emotional responses [ref. 410,ref. 416,ref. 417]. The stool PEA levels were associated with gut microbial diversity, with implications on host mental health [ref. 285,ref. 357,ref. 410,ref. 418]. The relative abundance of microbes of the Blautia and Dorea taxa was particularly associated with fecal PEA and anhedonia/amotivation [ref. 410]. Reduced microbial diversity corresponded with increased excretion of PEA and more severe anhedonia/amotivation [ref. 410]. PEA was also found to counteract autistic-like behaviors in BTBR T+ tf/J mice by dampening inflammation, reducing oxidative stress, reducing gut permeability, and altering the gut microbiota, besides its neuroprotection through induction of PPARα [ref. 419]. These authors found that PEA treatment increased the ratio of Firmicutes/Bacteroidetes, which was due to an increase in Firmicutes (e.g., Clostridials) and a decrease in Bacteroidetes [ref. 419].
4.7.3. The Relationship between Gut Microbiota, AEA, and Acute Respiratory Distress Syndrome
AEA was found to attenuate acute respiratory distress syndrome through modulating the gut microbiota [ref. 420]. In this study, the researchers investigated the effect of AEA on staphylococcal enterotoxin B (SEB)-mediated acute respiratory distress syndrome. SEB caused an increase in pathogenic bacteria in both the lungs and the gut [ref. 420]. AEA-treated mice showed increased level of anti-microbial peptides in the lung epithelial cells and prevented the increase in pathogenic bacteria induced by SEB [ref. 420]. AEA increased the level of several bacterial species (e.g., Lachnospiraceae and Clostridia) that produce elevated levels of SCFAs, such as butyrate and valerate, important for stabilizing the gut–lung microbial axis and suppressing inflammation [ref. 420]. In addition, AEA treatment increased the abundance of Muribaculaceae and reduced the abundance of Pseudomonas and Enterobacteriaceae [ref. 420].
4.7.4. The Relationship between Gut Microbiota, the Endocannabinoid System, and Obesity
Obesity is often characterized by low-grade inflammation, with increased levels of endocannabinoids in the plasma and adipose tissues and altered expression of CB1 [ref. 421]. Activation of CB1 and CB2 receptors reduces motility, limits secretion, and decreases hypersensitivity in the gut [ref. 422,ref. 423,ref. 424]. Impaired CB1 signaling protected against the development of obesity and steatosis [ref. 425,ref. 426,ref. 427]. Lipopolysaccharides from Gram-negative bacteria induce the production of endocannabinoids under inflammatory conditions that dampen the inflammatory response [ref. 318,ref. 428,ref. 429,ref. 430]. Vice versa, the activation of CB1 in mice increases circulating levels of lipopolysaccharides due to reduced expression of the tight junction proteins occludin and zonula occludens-1 (ZO-1), resulting in increased gut epithelial permeability [ref. 421,ref. 431].
The gut microbiota was found to modulate colon CB1 receptor expression in both normal and obese mice [ref. 421]. Obese mice fed with the prebiotic oligofructose showed reduced CB1 expression, lower AEA content, and increased expression of FAAH [ref. 421]. Obese mice treated with the CB1 antagonist SR141716A (Rimonabant) improved the gut barrier function and reduced body weight gain [ref. 421]. Mehrpouya-Bahrami et al. [ref. 432] observed that SR141716A attenuated diet-induced obesity and inflammation that was correlated with increased relative abundance of Akkermansia muciniphila and decreased abundance of Lanchnospiraceae and Erysipelotrichaceae in the gut. Interestingly, SR141716A prevented the intracellular replication of macrophage-phagocytosed Brucella suis by activating the macrophages, which was related to the inhibition of CB1 [ref. 433]. It would be interesting to study whether SR141716A also has a direct anti-microbial effect. In this context, it is worth mentioning that SR141716A could potentiate the anti-fungal activity of amphotericin B against Candida albicans and Cryptococcus neoformans by increasing cellular oxidative stress and cell membrane permeability [ref. 434].
Mice fed on high-fat, high-glucose diet showed altered microbiome with concomitant increase in AEA and 2-AG in the plasma [ref. 398]. The relative abundances of Adlercreutzia, Barnesiella, Coprobacillus, Eubacterium, and Parasutterella in the ileum were negatively associated with AEA levels [ref. 398]. The level of the AEA congener N-docosahexaenoylethanolamine (synaptamide, DHEA), which is required for normal brain development [ref. 435], was negatively associated with Barnesiella, Enterococcus, Eubacterium, Flavonifractor, and Intestinimonas in the ileum [ref. 398]. These authors also found a negative correlation between the Delftia genus and N-linoleoylethanolamine (LEA), while the Lactobacillus genus was associated with increased 2-docosahexaenoyl-glycerol (2-DHG) levels [ref. 398].
Repeated administration of OAE to mice fed on normal chow pellet diet for 11 days led to alteration in fecal microbial composition with an increase in Bacteroidetes (e.g., Bacteroides genus) and a decrease in Firmicutes (Lactobacillus), which is considered a “lean-like” phenotype [ref. 436]. OAE also reduced intestinal cytokines expression by immune cells isolated from Peyer’s patches [ref. 436].
4.7.5. The Relationship between Gut Microbiota, the Endocannabinoid System, and Inflammatory Bowel Diseases
Both AEA and PEA have been observed to reduce inflammation in murine models of colitis and inflammatory bowel disease [ref. 437,ref. 438,ref. 439]. Elevating the levels of the endocannabinoids by inhibiting FAAH could relieve colitis and inflammatory bowel disease [ref. 440,ref. 441]. Butyrate that is produced by gut microbiota (e.g., bacteria of the Ruminococcaceae and Lachnospiraceae family) [ref. 442] reduces inflammation and pain in colitis animal models, which can in part be mediated through the endocannabinoid system [ref. 443]. Vijay et al. [ref. 443] studied the association of the endocannabinoids AEA, 2-AG, OEA, and PEA with gut microbiome composition upon exercise. Under resting condition, AEA and OEA were positively associated with alpha diversity and with SCFA producing bacteria such as Bifidobacterium, Coprococcus 3, and Faecalibacterium, while being negatively associated with Collinsella [ref. 443]. AEA, OEA, and PEA increased with exercise, and changes in AEA correlated with bacterial butyrate production [ref. 443]. The increases in AEA and PEA correlated with decreased expression of the inflammatory mediators TNFα and IL-6 and increased expression of the anti-inflammatory cytokine IL-10 [ref. 443].
PEA was found to increase the phagocytosis and intracellular killing of encapsulated Escherichia coli K1 by activated microglial cells and macrophages [ref. 444,ref. 445]. Pre-treatment with PEA significantly increased the survival of mice challenged with Escherichia coli K1 [ref. 445]. Similarly, Heide et al. [ref. 446] observed that prophylactic PEA attenuated inflammation and increased the survival of mice challenged with intracerebral Escherichia coli K1 infection. Lower bacterial loads were observed in the spleen, liver, and blood of the PEA pretreated animals [ref. 446]. This was related to the anti-inflammatory effect, since PEA at 1 μg/mL had no effect on Escherichia coli growth in vitro [ref. 446].
The gut microbiota of IBD patients differs from healthy individuals, with a decrease in butyrate- and indole-producing bacteria, decrease in bile salt-sensitive bacteria, while an enrichment in bile acid-metabolizing bacteria [ref. 390,ref. 447,ref. 448,ref. 449]. Among others, Escherichia coli, Lactobacillus gasseri, Ruminococcus gnavus, and Blautia producta, were more abundant in IBD, while Bacteroides cellulosilyticus, Bacteroides fragilis, and Streptococcus salivarius were depleted [ref. 390]. Fornelos et al. [ref. 390] observed that certain N-acylethanolamines (NAEs), such as LEA, PEA, OEA and AEA, are elevated in the stool of IBD, Crohn’s disease, and/or ulcerative colitis patients, and stimulate the growth of bacterial species overrepresented in IBD while inhibiting bacterial species lacking in IBD. N-acylethanolamine levels were highest in samples with most differences in the microbiome, suggesting a connection between N-acylethanolamines and altered microbiota in IBD [ref. 390]. These researchers observed that LEA inhibited the growth of Bacteroides fragilis, Bacteroides cellulosilyticus, and Enterococcus faecalis, while slightly enhanced the growth of Escherichia coli, Ruminococcus gnavus, and Blautia producta [ref. 390]. AEA also transiently inhibited the growth of Bacteroides fragilis and Enterococcus faecalis that recovered with time [ref. 390]. The growth of Lactobacillus gasseri, which is enriched in IBD, was enhanced by OEA and LEA, and to a lesser extent by AEA [ref. 390]. LEA and AEA partly inhibited the growth of Alistipes shahii and Ruminococcus lactaris that are underrepresented in IBD subjects [ref. 390]. The growth of Streptococcus salivarius was completely prevented at a concentration of 50 μM LEA, AEA, or OEA, but only slightly inhibited by PEA [ref. 390].
4.7.6. Effect of N-acylethanolamines on the Microbial Composition of Stool Chemostats
Fornelos et al. [ref. 390] also studied the effect of N-acylethanolamines, including endocannabinoids, on the composition of two different stool chemostats. In the control chemostat A, the Enterobacteriaceae, Clostridiaceae, and Veillonellaceae taxa dominated, while in the presence of AEA, the bacterial community was almost entirely overtaken by Enterobacteriaceae [ref. 390]. LEA-treated chemostat A was dominated by Enterococcaceae, Veillonellaceae and Enterobacteriaceae at the expense of Streptococcaceae, Erysipelotrichaceae, Porphyromonadacea, Bacteroidaceae, and Rikenellaceae, while OEA treatment did not impact Enterobacteriaceae abundance but increased the relative abundances of Enterococcaceae and Streptococcaceae and decreased those of Bacteroidaceae and Rikenellaceae [ref. 390]. AEA and LEA also reduced the abundance of Barnesiella intestinihominis, Alistipes, and Bacteroides species, while they increased the abundance of Escherichia species [ref. 390]. In Chemostat B, the abundance of Blautia producta, Clostridium clostridioforme, Klebsiella pneumoniae, and Proteus mirabilis was increased in the presence of AEA or LEA [ref. 390]. These data indicate that LEA, AEA, and other N-acylethanolamines can shift the microbiome of a healthy individual into an IBD profile [ref. 390].
Transcriptional analysis showed that AEA upregulates both the anaerobic, reductive, and oxidative branches of the citrate cycle concomitant with increased energy metabolism and increased respiratory electron transport chain activity, especially in Enterobacteriaceae [ref. 390]. Metabolic changes occurring upon exposure to AEA, LEA, and other NAEs include the activation of bacterial processes involved in NAE metabolism [ref. 390].
Searching for an action mechanism of LEA and AEA on Bacteroides fragilis, the researchers found that the most upregulated genes are those encoding for membrane-associated efflux transport proteins, and the most downregulated gene was the long chain fatty acid (LCFA) importer fadL [ref. 390]. They further showed that two other genes involved in fatty acid metabolic processing were repressed: a fadD homolog that catalyzes esterification of incoming fatty acids into CoA thioesters and a fadE homolog involved in downstream fatty acid breakdown [ref. 390]. These data indicate that bacteria have developed mechanisms that can respond to endocannabinoids.
5. Conclusions
Both phytocannabinoids and endocannabinoids have diverse physiological activities that are, in part, mediated by common receptors in mammalians, each compound with its specificity and affinity, being agonists, partly agonists, inverse agonists, or antagonists. While these mechanisms have been widely investigated, the mechanisms leading to their anti-microbial effects are less understood. Despite the quite different structures of phytocannabinoids and endocannabinoids (Figure 1 and Figure 3), there are some common dominators that characterize their anti-bacterial activities (Figure 2 and Figure 4). For instance, they exert bacteriostatic activity, alter the membrane structure, induce either membrane hyperpolarization or depolarization, modulate gene expression including those involved in metabolism, affect virulence factors, and prevent biofilm formation (Figure 2 and Figure 4). Additionally, the AEA-mediated inhibition of yeast-hypha transition of Candida albicans and the hyphal adherence to epithelial cells seem to be mediated by alterations in gene expression (Figure 5). The multiple actions of phytocannabinoids and endocannabinoids suggest that the compounds do much more than just affecting membrane permeability as previously thought.
The emergence of antibiotic-resistant microbes is a clinical problem worldwide, and novel treatment strategies are urged. The important observation that some of the phytocannabinoids and endocannabinoids act on both drug-sensitive and drug-resistant Staphylococcus aureus makes them potential antibiotic adjuvants in treating drug-resistant infections, for instance for topical infectious wound treatment. Especially important is the ability of AEA and AraS to sensitize drug-resistant Staphylococcus aureus to antibiotics in virtue of their ability to prevent drug efflux and induce growth arrest. The addition of these endocannabinoids to the treatment cocktail will revive the use of already existing antibiotics. The ability of both CBG and the synthetic cannabinoid HU-210 to antagonize quorum sensing may have implications in the new era where quorum sensing inhibitors or quorum quenchers have attracted attention for alternative antibiotic drugs for antibiotic-resistant bacteria [ref. 450]. To reach this goal, further studies should be performed to clarify the spectrum of bacteria whose quorum sensing pathways are affected by cannabinoids. The increasing recognition of the complex interplay between the gut microbiota and the endocannabinoid system with the accompanying implications for various physiological and pathophysiological conditions, places the therapeutic uses of cannabinoids into a new spotlight.
References
- A. Sofowora, E. Ogunbodede, A. Onayade. The role and place of medicinal plants in the strategies for disease prevention. Afr. J. Tradit. Complement. Altern. Med., 2013. [DOI | PubMed]
- W.R. Bidlack, S.T. Omaye, M.S. Meskin, D.K. Topham. Phytochemicals as Bioactive Agents, 2000
- I.A. Ross. Medicinal Plants of the World, Volumes 3: Chemical Constituents, Traditional and Modern Medicinal Uses, 2007
- C.L. Gorlenko, H.Y. Kiselev, E.V. Budanova, A.A. Zamyatnin, L.N. Ikryannikova. Plant secondary metabolites in the battle of drugs and drug-resistant bacteria: New heroes or worse clones of antibiotics?. Antibiotics, 2020. [DOI | PubMed]
- N.S. Radulović, P.D. Blagojević, Z.Z. Stojanović-Radić, N.M. Stojanović. Antimicrobial plant metabolites: Structural diversity and mechanism of action. Curr. Med. Chem., 2013. [PubMed]
- N. Vaou, E. Stavropoulou, C. Voidarou, C. Tsigalou, E. Bezirtzoglou. Towards advances in medicinal plant antimicrobial activity: A review study on challenges and future perspectives. Microorganisms, 2021. [DOI | PubMed]
- D. Lahiri, S. Dash, R. Dutta, M. Nag. Elucidating the effect of anti-biofilm activity of bioactive compounds extracted from plants. J. Biosci., 2019. [DOI | PubMed]
- M. Ekor. The growing use of herbal medicines: Issues relating to adverse reactions and challenges in monitoring safety. Front. Pharmacol., 2014. [DOI | PubMed]
- R.A. Hussein, A.A. El-Anssary. Chapter 2: Plants secondary metabolites: The key drivers of the pharmacological actions of medicinal plants. Herb. Med., 2019. [DOI]
- C.D.R. Nunes, M. Barreto Arantes, S. Menezes de Faria Pereira, L. Leandro da Cruz, M. de Souza Passos, L. Pereira de Moraes, I.J.C. Vieira, D. Barros de Oliveira. Plants as sources of anti-inflammatory agents. Molecules, 2020. [DOI]
- A. Borges, A.C. Abreu, C. Dias, M.J. Saavedra, F. Borges, M. Simões. New perspectives on the use of phytochemicals as an emergent strategy to control bacterial infections including biofilms. Molecules, 2016. [DOI]
- S. Kaur, N. Sharma, A. Roy. Role of cannabinoids in various diseases: A review. Curr. Pharm. Biotechnol., 2021. [DOI]
- N.M. Kogan, R. Mechoulam. Cannabinoids in health and disease. Dialogues Clin. Neurosci., 2007. [DOI | PubMed]
- X.M. Rodríguez Mesa, A.F. Moreno Vergara, L.A. Contreras Bolaños, N. Guevara Moriones, A.L. Mejía Piñeros, S.P. Santander González. Therapeutic prospects of cannabinoids in the immunomodulation of prevalent autoimmune diseases. Cannabis Cannabinoid Res., 2021. [DOI | PubMed]
- R. Mechoulam, L.O. Hanuš, R. Pertwee, A.C. Howlett. Early phytocannabinoid chemistry to endocannabinoids and beyond. Nat. Rev. Neurosci., 2014. [DOI | PubMed]
- S. Banerjee, R.V. Sionov, M. Feldman, R. Smoum, R. Mechoulam, D. Steinberg. Anandamide alters the membrane properties, halts the cell division and prevents drug efflux in multidrug resistant Staphylococcus aureus. Sci. Rep., 2021. [DOI | PubMed]
- M. Feldman, R. Smoum, R. Mechoulam, D. Steinberg. Potential combinations of endocannabinoid/endocannabinoid-like compounds and antibiotics against methicillin-resistant Staphylococcus aureus. PLoS ONE, 2020. [DOI | PubMed]
- M. Feldman, R. Smoum, R. Mechoulam, D. Steinberg. Antimicrobial potential of endocannabinoid and endocannabinoid-like compounds against methicillin-resistant Staphylococcus aureus. Sci. Rep., 2018. [DOI | PubMed]
- M. Feldman, R. Sionov, R. Smoum, R. Mechoulam, I. Ginsburg, D. Steinberg. Comparative evaluation of combinatory interaction between endocannabinoid system compounds and Poly-L-lysine against Streptococcus mutans growth and biofilm formation. Biomed. Res. Int., 2020. [DOI | PubMed]
- A. Bilkei-Gorzo. The endocannabinoid system in normal and pathological brain ageing. Philos. Trans. R. Soc. B Biol. Sci., 2012. [DOI]
- G.A. Cabral, T.J. Rogers, A.H. Lichtman. Turning Over a New Leaf: Cannabinoid and endocannabinoid modulation of immune function. J. Neuroimmune Pharmacol., 2015. [DOI | PubMed]
- V. Chiurchiù, L. Battistini, M. Maccarrone. Endocannabinoid signalling in innate and adaptive immunity. Immunology, 2015. [DOI | PubMed]
- M. Maccarrone, I. Bab, T. Bíró, G.A. Cabral, S.K. Dey, V. Di Marzo, J.C. Konje, G. Kunos, R. Mechoulam, P. Pacher. Endocannabinoid signaling at the periphery: 50 years after THC. Trends Pharmacol. Sci., 2015. [DOI | PubMed]
- P. Pacher, N.M. Kogan, R. Mechoulam. Beyond THC and endocannabinoids. Annu. Rev. Pharmacol. Toxicol., 2020. [DOI | PubMed]
- H.L. Li. The origin and use of Cannabis in eastern Asia linguistic-cultural implications. J. Econ. Bot., 1973. [DOI]
- S. Pisanti, M. Bifulco. Medical Cannabis: A plurimillennial history of an evergreen. J. Cell. Physiol., 2019. [DOI | PubMed]
- H.E. Jiang, X. Li, Y.X. Zhao, D.K. Ferguson, F. Hueber, S. Bera, Y.F. Wang, L.C. Zhao, C.J. Liu, C.S. Li. A new insight into Cannabis sativa (Cannabaceae) utilization from 2500-year-old Yanghai Tombs, Xinjiang, China. J. Ethnopharmacol., 2006. [DOI | PubMed]
- M. Touw. The religious and medicinal uses of Cannabis in China, India and Tibet. J. Psychoact. Drugs, 1981. [DOI | PubMed]
- E.P. Baron, P. Lucas, J. Eades, O. Hogue. Patterns of medicinal cannabis use, strain analysis, and substitution effect among patients with migraine, headache, arthritis, and chronic pain in a medicinal cannabis cohort. J. Headache Pain, 2018. [DOI | PubMed]
- T. Kantonen, T. Karjalainen, L. Pekkarinen, J. Isojärvi, K. Kalliokoski, V. Kaasinen, J. Hirvonen, P. Nuutila, L. Nummenmaa. Cerebral μ-opioid and CB1 receptor systems have distinct roles in human feeding behavior. Transl. Psychiatry, 2021. [DOI | PubMed]
- S. Lal, A. Shekher, Puneet, A.S. Narula, H. Abrahamse, S.C. Gupta. Cannabis and its constituents for cancer: History, biogenesis, chemistry and pharmacological activities. Pharmacol. Res., 2021. [DOI | PubMed]
- A. Pollio. The Name of Cannabis: A short guide for nonbotanists. Cannabis Cannabinoid Res., 2016. [DOI | PubMed]
- L.O. Hanuš, Y. Hod. Terpenes/terpenoids in Cannabis: Are They important?. Med. Cannabis Cannabinoids, 2020. [DOI | PubMed]
- L. Nissen, A. Zatta, I. Stefanini, S. Grandi, B. Sgorbati, B. Biavati, A. Monti. Characterization and antimicrobial activity of essential oils of industrial hemp varieties (Cannabis sativa L.). Fitoterapia, 2010. [DOI | PubMed]
- D. Vergara, R. Gaudino, T. Blank, B. Keegan. Modeling cannabinoids from a large-scale sample of Cannabis sativa chemotypes. PLoS ONE, 2020. [DOI | PubMed]
- E. Borroto Fernandez, V. Peterseil, G. Hackl, S. Menges, E. de Meijer, C. Staginnus. Distribution of chemical phenotypes (chemotypes) in European agricultural hemp (Cannabis sativa L.) cultivars. J. Forensic Sci., 2020. [DOI | PubMed]
- C. Casajuana Köguel, H. López-Pelayo, M.M. Balcells-Olivero, J. Colom, A. Gual. Psychoactive constituents of Cannabis and their clinical implications: A systematic review. Adicciones, 2018. [DOI | PubMed]
- S. Farag, O. Kayser. Cultivation and breeding of Cannabis sativa L. for preparation of standardized extracts for medicinal purposes. Medicinal and Aromatic Plants of the World: Scientific, Production, Commercial and Utilization Aspects, 2015. [DOI]
- L.O. Hanuš, S.M. Meyer, E. Muñoz, O. Taglialatela-Scafati, G. Appendino. Phytocannabinoids: A unified critical inventory. Nat. Prod. Rep., 2016. [DOI | PubMed]
- T. Gülck, B.L. Møller. Phytocannabinoids: Origins and biosynthesis. Trends Plant Sci., 2020. [DOI | PubMed]
- S.J. Livingston, T.D. Quilichini, J.K. Booth, D.C.J. Wong, K.H. Rensing, J. Laflamme-Yonkman, S.D. Castellarin, J. Bohlmann, J.E. Page, A.L. Samuels. Cannabis glandular trichomes alter morphology and metabolite content during flower maturation. Plant J., 2020. [DOI | PubMed]
- C.M. Andre, J.F. Hausman, G. Guerriero. Cannabis sativa: The plant of the thousand and one molecules. Front. Plant Sci., 2016. [DOI | PubMed]
- V. Mediavilla, M. Jonquera, I. Schmid-Slembrouck, A. Soldati. Decimal code for growth stages of hemp (Cannabis sativa L.). J. Int. Hemp. Assoc., 1998
- S. Eichhorn Bilodeau, B.S. Wu, A.S. Rufyikiri, S. MacPherson, M. Lefsrud. An update on plant photobiology and implications for Cannabis production. Front. Plant Sci., 2019. [DOI | PubMed]
- G. Magagnini, G. Grassi, S. Kotiranta. The effect of light spectrum on the morphology and cannabinoid content of Cannabis sativa L.. Med. Cannabis Cannabinoids, 2018. [DOI | PubMed]
- N. Bernstein, J. Gorelick, R. Zerahia, S. Koch. Impact of N, P, K, and humic acid supplementation on the chemical profile of medical Cannabis (Cannabis sativa L.). Front. Plant Sci., 2019. [DOI | PubMed]
- L. Burgel, J. Hartung, D. Schibano, S. Graeff-Hönninger. Impact of different phytohormones on morphology, yield and cannabinoid content of Cannabis sativa L.. Plants, 2020. [DOI]
- A. Radoševič, M. Kupinič, L. Grlič. Antibiotic activity of various types of Cannabis resin. Nature, 1962. [DOI | PubMed]
- E. Stuyt. The problem with the current high potency THC marijuana from the perspective of an addiction psychiatrist. Mo. Med., 2018. [PubMed]
- W.T. Caiaffa, D. Vlahov, N.M. Graham, J. Astemborski, L. Solomon, K.E. Nelson, A. Muñoz. Drug smoking, Pneumocystis carinii pneumonia, and immunosuppression increase risk of bacterial pneumonia in human immunodeficiency virus-seropositive injection drug users. Am. J. Respir. Crit. Care Med., 1994. [DOI | PubMed]
- M.D. Roth, K. Whittaker, K. Salehi, D.P. Tashkin, G.C. Baldwin. Mechanisms for impaired effector function in alveolar macrophages from marijuana and cocaine smokers. J. Neuroimmunol., 2004. [DOI | PubMed]
- G.A. Cabral, D.A. Dove Pettit. Drugs and immunity: Cannabinoids and their role in decreased resistance to infectious disease. J. Neuroimmunol., 1998. [DOI | PubMed]
- S.B. Maggirwar, J.H. Khalsa. The link between Cannabis use, immune system, and viral infections. Viruses, 2021. [DOI | PubMed]
- K. Benedict, G.R. Thompson, B.R. Jackson. Cannabis use and fungal infections in a commercially insured population, United States, 2016. Emerg. Infect. Dis., 2020. [DOI | PubMed]
- Z. Krejčí. Antibacterial action of Cannabis indica. Lek. Listy, 1952. [PubMed]
- Z. Krejčí, M. Horak, F. Santavy. Hemp (Cannabis sativa)-an antibiotic drug. 3. Isolation and constitution of two acids from Cannabis sativa. Die Pharmazie, 1959. [PubMed]
- Z. Krejčí, F. Šantavý. Isolace dalších látek z listí indického konopí Cannabis sativa L.. Acta Univ. Palacki. Olomuc., 1955
- G. Appendino, S. Gibbons, A. Giana, A. Pagani, G. Grassi, M. Stavri, E. Smith, M.M. Rahman. Antibacterial cannabinoids from Cannabis sativa: A structure-activity study. J. Nat. Prod., 2008. [DOI | PubMed]
- R. Mechoulam, Y. Gaoni. Hashish, IV. The isolation and structure of cannabinolic cannabidiolic and cannabigerolic acids. Tetrahedron, 1965. [DOI | PubMed]
- M.S. Mahmud, M.S. Hossain, A. Ahmed, M.Z. Islam, M.E. Sarker, M.R. Islam. Antimicrobial and antiviral (SARS-CoV-2) potential of cannabinoids and Cannabis sativa: A comprehensive review. Molecules, 2021. [DOI | PubMed]
- M.A. Farha, O.M. El-Halfawy, R.T. Gale, C.R. MacNair, L.A. Carfrae, X. Zhang, N.G. Jentsch, J. Magolan, E.D. Brown. Uncovering the hidden antibiotic potential of Cannabis. ACS Infect. Dis., 2020. [DOI | PubMed]
- L.D. Martinenghi, R. Jønsson, T. Lund, H. Jenssen. Isolation, purification, and antimicrobial characterization of cannabidiolic acid and cannabidiol from Cannabis sativa L.. Biomolecules, 2020. [DOI]
- M.M. Radwan, M.A. Elsohly, D. Slade, S.A. Ahmed, I.A. Khan, S.A. Ross. Biologically active cannabinoids from high-potency Cannabis sativa. J. Nat. Prod., 2009. [DOI | PubMed]
- L. Ferenczy. Antibacterial substances in seeds. Nature, 1956. [DOI | PubMed]
- K. Wasim, I. Haq, M. Ashraf. Antimicrobial studies of the leaf of Cannabis sativa L.. Pak. J. Pharm. Sci., 1995. [PubMed]
- E. Ali, A. Almagboul, S. Khogali, U. Gergeir. Antimicrobial activity of Cannabis sativa L.. Chin. Med., 2012. [DOI]
- J. Novak, K. Zitterl-Eglseer, S.G. Deans, C.M. Franz. Essential oils of different cultivars of Cannabis sativa L. and their antimicrobial activity. Flavour Fragance J., 2001. [DOI]
- G. Zengin, L. Menghini, A. Di Sotto, R. Mancinelli, F. Sisto, S. Carradori, S. Cesa, C. Fraschetti, A. Filippi, L. Angiolella. Chromatographic analyses, In vitro biological activities, and cytotoxicity of Cannabis sativa L. essential oil: A multidisciplinary study. Molecules, 2018. [DOI]
- M. Pellegrini, S. Palmieri, A. Ricci, A. Serio, A. Paparella, C. Lo Sterzo. In vitro antioxidant and antimicrobial activity of Cannabis sativa L. cv ‘Futura 75’ essential oil. Nat. Prod. Res., 2021. [DOI | PubMed]
- G. Benelli, R. Pavela, G. Lupidi, M. Nabissi, R. Petrelli, S.L. Ngahang Kamte, L. Cappellacci, D. Fiorini, S. Sut, S. Dall’Acqua. The crop-residue of fiber hemp cv. Futura 75: From a waste product to a source of botanical insecticides. Environ. Sci. Pollut. Res. Int., 2018. [DOI | PubMed]
- T.G. Thomas, S.K. Sharma, A. Prakash, B.R. Sharma. Insecticidal properties of essential oil of Cannabis sativa Linn. against mosquito larvae. Entomon, 2000
- S. Palmieri, F. Maggio, M. Pellegrini, A. Ricci, A. Serio, A. Paparella, C. Lo Sterzo. Effect of the distillation time on the chemical composition, antioxidant potential and antimicrobial activity of essential oils from different Cannabis sativa L. cultivars. Molecules, 2021. [DOI | PubMed]
- V.D. Zheljazkov, V. Sikora, I. Dincheva, M. Kačániová, T. Astatkie, I.B. Semerdjieva, D. Latkovic. Industrial, CBD, and wild hemp: How different are their essential oil profile and antimicrobial activity?. Molecules, 2020. [DOI]
- E.B. Russo. Taming THC: Potential cannabis synergy and phytocannabinoid-terpenoid entourage effects. Br. J. Pharmacol., 2011. [DOI | PubMed]
- L. Schofs, M.D. Sparo, S.F. Sánchez Bruni. The antimicrobial effect behind Cannabis sativa. Pharmacol. Res. Perspect., 2021. [DOI | PubMed]
- J.A. Karas, L.J.M. Wong, O.K.A. Paulin, A.C. Mazeh, M.H. Hussein, J. Li, T. Velkov. The antimicrobial activity of cannabinoids. Antibiotics, 2020. [DOI]
- S.S. Kim, J.S. Baik, T.H. Oh, W.J. Yoon, N.H. Lee, C.G. Hyun. Biological activities of Korean Citrus obovoides and Citrus natsudaidai essential oils against acne-inducing bacteria. Biosci. Biotechnol. Biochem., 2008. [DOI | PubMed]
- A.C. Rivas da Silva, P.M. Lopes, M.M. Barros de Azevedo, D.C. Costa, C.S. Alviano, D.S. Alviano. Biological activities of α-pinene and β-pinene enantiomers. Molecules, 2012. [DOI | PubMed]
- A.M. Leite, E.O. Lima, E.L. Souza, M.F.F.M. Diniz, V.N. Trajano, I.A. Medeiros. Inhibitory effect of β-pinene, α-pinene and eugenol on the growth of potential infectious endocarditis causing Gram-positive bacteria. Rev. Bras. De Ciências Farm., 2007. [DOI]
- Y. Inoue, A. Shiraishi, T. Hada, H. Hamashima, J. Shimada. The antibacterial effects of myrcene on Staphylococcus aureus and its role in the essential oil of the tea tree (Melaleuca alternifolia). Nat. Med., 2004
- G.B. Zore, A.D. Thakre, S. Jadhav, S.M. Karuppayil. Terpenoids inhibit Candida albicans growth by affecting membrane integrity and arrest of cell cycle. Phytomedicine, 2011. [DOI | PubMed]
- Y. Han, Z. Sun, W. Chen. Antimicrobial susceptibility and antibacterial mechanism of limonene against Listeria monocytogenes. Molecules, 2019. [DOI]
- G.A. Subramenium, K. Vijayakumar, S.K. Pandian. Limonene inhibits streptococcal biofilm formation by targeting surface-associated virulence factors. J. Med. Microbiol., 2015. [DOI | PubMed]
- A.S. Wanas, M.M. Radwan, Z. Mehmedic, M. Jacob, I.A. Khan, M.A. Elsohly. Antifungal activity of the volatiles of high potency Cannabis sativa L. against Cryptococcus neoformans. Rec. Nat. Prod., 2016
- D.C. Arruda, F.L. D’Alexandri, A.M. Katzin, S.R. Uliana. Antileishmanial activity of the terpene nerolidol. Antimicrob. Agents Chemother., 2005. [DOI | PubMed]
- N.P. Lopes, M.J. Kato, E.H. Andrade, J.G. Maia, M. Yoshida, A.R. Planchart, A.M. Katzin. Antimalarial use of volatile oil from leaves of Virola surinamensis (Rol.) Warb. by Waiãpi Amazon Indians. J. Ethnopharmacol., 1999. [DOI | PubMed]
- H. Rodrigues Goulart, E.A. Kimura, V.J. Peres, A.S. Couto, F.A. Aquino Duarte, A.M. Katzin. Terpenes arrest parasite development and inhibit biosynthesis of isoprenoids in Plasmodium falciparum. Antimicrob. Agents Chemother., 2004. [DOI | PubMed]
- S.S. Dahham, Y.M. Tabana, M.A. Iqbal, M.B. Ahamed, M.O. Ezzat, A.S. Majid, A.M. Majid. The anticancer, antioxidant and antimicrobial properties of the sesquiterpene β-caryophyllene from the essential oil of Aquilaria crassna. Molecules, 2015. [DOI | PubMed]
- D. Yang, L. Michel, J.P. Chaumont, J. Millet-Clerc. Use of caryophyllene oxide as an antifungal agent in an in vitro experimental model of onychomycosis. Mycopathologia, 1999. [DOI | PubMed]
- M.A. ElSohly, M.M. Radwan, W. Gul, S. Chandra, A. Galal. Phytochemistry of Cannabis sativa L.. Prog. Chem. Org. Nat. Prod., 2017. [DOI | PubMed]
- M. Pourseyed Lazarjani, S. Torres, T. Hooker, C. Fowlie, O. Young, A. Seyfoddin. Methods for quantification of cannabinoids: A narrative review. J. Cannabis Res., 2020. [DOI | PubMed]
- A. Hazekamp, Y.H. Choi, R. Verpoorte. Quantitative analysis of cannabinoids from Cannabis sativa using 1H-NMR. Chem. Pharm. Bull., 2004. [DOI]
- J. Fischedick, F. Van Der Kooy, R. Verpoorte. Cannabinoid receptor 1 binding activity and quantitative analysis of Cannabis sativa L. smoke and vapor. Chem. Pharm. Bull., 2010. [DOI]
- E.A. Ibrahim, W. Gul, S.W. Gul, B.J. Stamper, G.M. Hadad, R.A. Abdel Salam, A.K. Ibrahim, S.A. Ahmed, S. Chandra, H. Lata. Determination of acid and neutral cannabinoids in extracts of different strains of Cannabis sativa using GC-FID. Planta Med., 2018. [DOI | PubMed]
- E.B. Russo, J. Marcu. Cannabis Pharmacology: The usual suspects and a few promising leads. Adv. Pharmacol., 2017. [DOI | PubMed]
- F. Pellati, V. Brighenti, J. Sperlea, L. Marchetti, D. Bertelli, S. Benvenuti. New methods for the comprehensive analysis of bioactive compounds in Cannabis sativa L. (hemp). Molecules, 2018. [DOI]
- R. Iseppi, V. Brighenti, M. Licata, A. Lambertini, C. Sabia, P. Messi, F. Pellati, S. Benvenuti. Chemical Characterization and evaluation of the antibacterial activity of essential oils from fibre-type Cannabis sativa L. (Hemp). Molecules, 2019. [DOI]
- R. Mechoulam, Y. Gaoni. Recent advances in the chemistry of hashish. Fortschr. Chem. Org. Nat., 1967. [DOI]
- M.A. Elsohly, D. Slade. Chemical constituents of marijuana: The complex mixture of natural cannabinoids. Life Sci., 2005. [DOI | PubMed]
- M.M. Radwan, S.A. Ross, D. Slade, S.A. Ahmed, F. Zulfiqar, M.A. Elsohly. Isolation and characterization of new Cannabis constituents from a high potency variety. Planta Med., 2008. [DOI | PubMed]
- M.M. Radwan, M.A. Elsohly, D. Slade, S.A. Ahmed, L. Wilson, A.T. El-Alfy, I.A. Khan, S.A. Ross. Non-cannabinoid constituents from a high potency Cannabis sativa variety. Phytochemistry, 2008. [DOI | PubMed]
- C. Leizer, D. Ribnicky, A. Poulev, S. Dushenkov, I. Raskin. The composition of Hemp seed oil and its potential as an important source of nutrition. J. Nutraceuticals Funct. Med. Foods, 2000. [DOI]
- M.M. Radwan, S. Chandra, S. Gul, M.A. ElSohly. Cannabinoids, phenolics, terpenes and alkaloids of Cannabis. Molecules, 2021. [DOI | PubMed]
- C.E. Turner, M.A. Elsohly, E.G. Boeren. Constituents of Cannabis sativa L. XVII. A review of the natural constituents. J. Nat. Prod., 1980. [DOI | PubMed]
- G. Appendino, G. Chianese, O. Taglialatela-Scafati. Cannabinoids: Occurrence and medicinal chemistry. Curr. Med. Chem., 2011. [DOI | PubMed]
- Y. Yang, R. Vyawahare, M. Lewis-Bakker, H.A. Clarke, A.H.C. Wong, L.P. Kotra. Bioactive chemical composition of Cannabis extracts and cannabinoid receptors. Molecules, 2020. [DOI]
- E.M. Rock, L.A. Parker. Constituents of Cannabis sativa. Adv. Exp. Med. Biol., 2021. [DOI | PubMed]
- O. Aizpurua-Olaizola, U. Soydaner, E. Öztürk, D. Schibano, Y. Simsir, P. Navarro, N. Etxebarria, A. Usobiaga. Evolution of the cannabinoid and terpene content during the growth of Cannabis sativa plants from different chemotypes. J. Nat. Prod., 2016. [DOI | PubMed]
- M. Toyota, T. Shimamura, H. Ishii, M. Renner, J. Braggins, Y. Asakawa. New bibenzyl cannabinoid from the New Zealand liverwort Radula marginata. Chem. Pharm. Bull., 2002. [DOI]
- F. Bohlmann, E. Hoffmann. Cannabigerol-ähnliche verbindungen aus Helichrysum umbraculigerum. Phytochemistry, 1979. [DOI]
- A.E. Kirkland, M.C. Fadus, S.A. Gruber, K.M. Gray, T.E. Wilens, L.M. Squeglia. A scoping review of the use of cannabidiol in psychiatric disorders. Psychiatry Res., 2021. [DOI | PubMed]
- D.P. Finn, S. Haroutounian, A.G. Hohmann, E. Krane, N. Soliman, A.S.C. Rice. Cannabinoids, the endocannabinoid system, and pain: A review of preclinical studies. Pain, 2021. [DOI | PubMed]
- Y. Lu, H.D. Anderson. Cannabinoid signaling in health and disease. Can. J. Physiol. Pharmacol., 2017. [DOI | PubMed]
- D.E. Slawek, S.A. Curtis, J.H. Arnsten, C.O. Cunningham. Clinical approaches to Cannabis: A narrative review. Med. Clin. N. Am., 2022. [DOI | PubMed]
- A. Ligresti, L. De Petrocellis, V. Di Marzo. From phytocannabinoids to cannabinoid receptors and endocannabinoids: Pleiotropic physiological and pathological roles through complex pharmacology. Physiol. Rev., 2016. [DOI | PubMed]
- S. Atalay, I. Jarocka-Karpowicz, E. Skrzydlewska. Antioxidative and anti-inflammatory properties of cannabidiol. Antioxidants, 2019. [DOI]
- K.B. Walsh, A.E. McKinney, A.E. Holmes. Minor cannabinoids: Biosynthesis, molecular pharmacology and potential therapeutic uses. Front. Pharmacol., 2021. [DOI | PubMed]
- C. Zanettini, L.V. Panlilio, M. Alicki, S.R. Goldberg, J. Haller, S. Yasar. Effects of endocannabinoid system modulation on cognitive and emotional behavior. Front. Behav. Neurosci., 2011. [DOI | PubMed]
- E. Murillo-Rodriguez, J.C. Pastrana-Trejo, M. Salas-Crisóstomo, M. de-la-Cruz. The endocannabinoid system modulating levels of consciousness, emotions and likely dream contents. CNS Neurol. Disord. Drug Targets, 2017. [DOI | PubMed]
- C.S. Pauli, M. Conroy, B.D. Vanden Heuvel, S.H. Park. Cannabidiol drugs clinical trial outcomes and adverse effects. Front. Pharmacol., 2020. [DOI | PubMed]
- A.I. Fraguas-Sánchez, A.I. Torres-Suárez. Medical use of cannabinoids. Drugs, 2018. [DOI | PubMed]
- E.A. Romero-Sandoval, J.E. Fincham, A.L. Kolano, B.N. Sharpe, P.A. Alvarado-Vázquez. Cannabis for chronic pain: Challenges and considerations. Pharmacotherapy, 2018. [DOI | PubMed]
- S. Malaca, F.P. Busardò, G. Nittari, A. Sirignano, G. Ricci. Fourth generation of synthetic cannabinoid receptor agonists: A review on the latest insights. Curr. Pharm. Des., 2021. [DOI]
- L.A. Matsuda, S.J. Lolait, M.J. Brownstein, A.C. Young, T.I. Bonner. Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature, 1990. [DOI | PubMed]
- S. Munro, K.L. Thomas, M. Abu-Shaar. Molecular characterization of a peripheral receptor for cannabinoids. Nature, 1993. [DOI | PubMed]
- R.G. Pertwee, A.C. Howlett, M.E. Abood, S.P. Alexander, V. Di Marzo, M.R. Elphick, P.J. Greasley, H.S. Hansen, G. Kunos, K. Mackie. International Union of Basic and Clinical Pharmacology. LXXIX. Cannabinoid receptors and their ligands: Beyond CB₁ and CB₂. Pharmacol. Rev., 2010. [DOI | PubMed]
- T. Gómez del Pulgar, G. Velasco, M. Guzmán. The CB1 cannabinoid receptor is coupled to the activation of protein kinase B/Akt. Biochem. J., 2000. [DOI | PubMed]
- D. Rueda, I. Galve-Roperh, A. Haro, M. Guzmán. The CB1 cannabinoid receptor is coupled to the activation of c-Jun N-terminal kinase. Mol. Pharmacol., 2000. [DOI | PubMed]
- I. Galve-Roperh, D. Rueda, T. Gómez del Pulgar, G. Velasco, M. Guzmán. Mechanism of extracellular signal-regulated kinase activation by the CB1 cannabinoid receptor. Mol. Pharmacol., 2002. [DOI | PubMed]
- C. Blázquez, A. Chiarlone, L. Bellocchio, E. Resel, P. Pruunsild, D. García-Rincón, M. Sendtner, T. Timmusk, B. Lutz, I. Galve-Roperh. The CB₁ cannabinoid receptor signals striatal neuroprotection via a PI3K/Akt/mTORC1/BDNF pathway. Cell Death Differ., 2015. [DOI | PubMed]
- E. Molina-Holgado, J.M. Vela, A. Arévalo-Martín, G. Almazán, F. Molina-Holgado, J. Borrell, C. Guaza. Cannabinoids promote oligodendrocyte progenitor survival: Involvement of cannabinoid receptors and phosphatidylinositol-3 kinase/Akt signaling. J. Neurosci., 2002. [DOI | PubMed]
- O. Gomez, A. Sanchez-Rodriguez, M. Le, C. Sanchez-Caro, F. Molina-Holgado, E. Molina-Holgado. Cannabinoid receptor agonists modulate oligodendrocyte differentiation by activating PI3K/Akt and the mammalian target of rapamycin (mTOR) pathways. Br. J. Pharmacol., 2011. [DOI | PubMed]
- M.H. Rhee, Z. Vogel, J. Barg, M. Bayewitch, R. Levy, L. Hanus, A. Breuer, R. Mechoulam. Cannabinol derivatives: Binding to cannabinoid receptors and inhibition of adenylylcyclase. J. Med. Chem., 1997. [DOI | PubMed]
- A.C. Howlett, M.E. Abood. CB1 and CB2 receptor pharmacology. Adv. Pharmacol., 2017. [DOI | PubMed]
- W.A. Devane, L. Hanus, A. Breuer, R.G. Pertwee, L.A. Stevenson, G. Griffin, D. Gibson, A. Mandelbaum, A. Etinger, R. Mechoulam. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science, 1992. [DOI | PubMed]
- T. Sugiura, K. Waku. 2-Arachidonoylglycerol and the cannabinoid receptors. Chem. Phys. Lipids, 2000. [DOI | PubMed]
- D.A. Kendall, G.A. Yudowski. Cannabinoid receptors in the central nervous system: Their signaling and roles in disease. Front. Cell. Neurosci., 2016. [DOI | PubMed]
- R.G. Pertwee. The diverse CB1 and CB2 receptor pharmacology of three plant cannabinoids: Delta9-tetrahydrocannabinol, cannabidiol and delta9-tetrahydrocannabivarin. Br. J. Pharmacol., 2008. [DOI | PubMed]
- I. Svízenská, P. Dubový, A. Sulcová. Cannabinoid receptors 1 and 2 (CB1 and CB2), their distribution, ligands and functional involvement in nervous system structures—A short review. Pharmacol. Biochem. Behav., 2008. [DOI | PubMed]
- M. Koch. Cannabinoid receptor signaling in central regulation of feeding behavior: A mini-review. Front. Neurosci., 2017. [DOI | PubMed]
- J.C. Ashton, J.L. Wright, J.M. McPartland, J.D. Tyndall. Cannabinoid CB1 and CB2 receptor ligand specificity and the development of CB2-selective agonists. Curr. Med. Chem., 2008. [DOI | PubMed]
- C. Wei, L. Huang, Y. Zheng, X. Cai. Selective activation of cannabinoid receptor 2 regulates Treg/Th17 balance to ameliorate neutrophilic asthma in mice. Ann. Transl. Med., 2021. [DOI | PubMed]
- S.L. Saldaña-Shumaker, A.J. Grenning, C.W. Cunningham. Modern approaches to the development of synthetic cannabinoid receptor probes. Pharmacol. Biochem. Behav., 2021. [DOI | PubMed]
- J. Gertsch, M. Leonti, S. Raduner, I. Racz, J.Z. Chen, X.Q. Xie, K.H. Altmann, M. Karsak, A. Zimmer. Beta-caryophyllene is a dietary cannabinoid. Proc. Natl. Acad. Sci. USA, 2008. [DOI | PubMed]
- V.S. Kumawat, G. Kaur. Cannabinoid 2 receptor agonist and L-arginine combination attenuates diabetic cardiomyopathy in rats via NF-ĸβ inhibition. Can. J. Physiol. Pharmacol., 2021. [DOI | PubMed]
- J. Mlost, P. Kac, M. Kędziora, K. Starowicz. Antinociceptive and chondroprotective effects of prolonged β-caryophyllene treatment in the animal model of osteoarthritis: Focus on tolerance development. Neuropharmacology, 2022. [DOI | PubMed]
- S. Katsuyama, H. Mizoguchi, H. Kuwahata, T. Komatsu, K. Nagaoka, H. Nakamura, G. Bagetta, T. Sakurada, S. Sakurada. Involvement of peripheral cannabinoid and opioid receptors in β-caryophyllene-induced antinociception. Eur. J. Pain, 2013. [DOI | PubMed]
- T. Lowin, R.H. Straub. Cannabinoid-based drugs targeting CB1 and TRPV1, the sympathetic nervous system, and arthritis. Arthritis Res. Ther., 2015. [DOI | PubMed]
- R.A. Ross. Anandamide and vanilloid TRPV1 receptors. Br. J. Pharmacol., 2003. [DOI | PubMed]
- E. Ryberg, N. Larsson, S. Sjögren, S. Hjorth, N.O. Hermansson, J. Leonova, T. Elebring, K. Nilsson, T. Drmota, P.J. Greasley. The orphan receptor GPR55 is a novel cannabinoid receptor. Br. J. Pharmacol., 2007. [DOI | PubMed]
- D. McHugh, J. Page, E. Dunn, H.B. Bradshaw. Δ(9)-Tetrahydrocannabinol and N-arachidonyl glycine are full agonists at GPR18 receptors and induce migration in human endometrial HEC-1B cells. Br. J. Pharmacol., 2012. [DOI | PubMed]
- S.E. O’Sullivan. Cannabinoids go nuclear: Evidence for activation of peroxisome proliferator-activated receptors. Br. J. Pharmacol., 2007. [DOI | PubMed]
- F. Comelli, G. Giagnoni, I. Bettoni, M. Colleoni, B. Costa. Antihyperalgesic effect of a Cannabis sativa extract in a rat model of neuropathic pain: Mechanisms involved. Phytother. Res., 2008. [DOI | PubMed]
- M.M. Ibrahim, F. Porreca, J. Lai, P.J. Albrecht, F.L. Rice, A. Khodorova, G. Davar, A. Makriyannis, T.W. Vanderah, H.P. Mata. CB2 cannabinoid receptor activation produces antinociception by stimulating peripheral release of endogenous opioids. Proc. Natl. Acad. Sci. USA, 2005. [DOI | PubMed]
- L. Callén, E. Moreno, P. Barroso-Chinea, D. Moreno-Delgado, A. Cortés, J. Mallol, V. Casadó, J.L. Lanciego, R. Franco, C. Lluis. Cannabinoid receptors CB1 and CB2 form functional heteromers in brain. J. Biol. Chem., 2012. [DOI | PubMed]
- P. Carriba, O. Ortiz, K. Patkar, Z. Justinova, J. Stroik, A. Themann, C. Müller, A.S. Woods, B.T. Hope, F. Ciruela. Striatal adenosine A2A and cannabinoid CB1 receptors form functional heteromeric complexes that mediate the motor effects of cannabinoids. Neuropsychopharmacology, 2007. [DOI | PubMed]
- G. Navarro, P. Carriba, J. Gandía, F. Ciruela, V. Casadó, A. Cortés, J. Mallol, E.I. Canela, C. Lluis, R. Franco. Detection of heteromers formed by cannabinoid CB1, dopamine D2, and adenosine A2A G-protein-coupled receptors by combining bimolecular fluorescence complementation and bioluminescence energy transfer. Sci. World J., 2008. [DOI | PubMed]
- M.T. Tebano, A. Martire, V. Chiodi, R. Pepponi, A. Ferrante, M.R. Domenici, C. Frank, J.F. Chen, C. Ledent, P. Popoli. Adenosine A2A receptors enable the synaptic effects of cannabinoid CB1 receptors in the rodent striatum. J. Neurochem., 2009. [DOI | PubMed]
- R. Rozenfeld, A. Gupta, K. Gagnidze, M.P. Lim, I. Gomes, D. Lee-Ramos, N. Nieto, L.A. Devi. AT1R-CB₁R heteromerization reveals a new mechanism for the pathogenic properties of angiotensin II. EMBO J., 2011. [DOI | PubMed]
- M. Hojo, Y. Sudo, Y. Ando, K. Minami, M. Takada, T. Matsubara, M. Kanaide, K. Taniyama, K. Sumikawa, Y. Uezono. Mu-Opioid receptor forms a functional heterodimer with cannabinoid CB1 receptor: Electrophysiological and FRET assay analysis. J. Pharmacol. Sci., 2008. [DOI | PubMed]
- J.A. López-Moreno, A. López-Jiménez, M.A. Gorriti, F.R. de Fonseca. Functional interactions between endogenous cannabinoid and opioid systems: Focus on alcohol, genetics and drug-addicted behaviors. Curr. Drug Targets, 2010. [DOI | PubMed]
- R.J. Ward, J.D. Pediani, G. Milligan. Heteromultimerization of cannabinoid CB1 receptor and orexin OX1 receptor generates a unique complex in which both protomers are regulated by orexin A. J. Biol. Chem., 2011. [DOI | PubMed]
- S. Zou, U. Kumar. Cannabinoid Receptors and the endocannabinoid system: Signaling and function in the central nervous system. Int. J. Mol. Sci., 2018. [DOI]
- F. Shahbazi, V. Grandi, A. Banerjee, J.F. Trant. Cannabinoids and cannabinoid receptors: The story so far. iScience, 2020. [DOI | PubMed]
- I. Raïch, R. Rivas-Santisteban, A. Lillo, J. Lillo, I. Reyes-Resina, X. Nadal, C. Ferreiro-Vera, V.S. de Medina, M. Majellaro, E. Sotelo. Similarities and differences upon binding of naturally occurring Δ9-tetrahydrocannabinol-derivatives to cannabinoid CB1 and CB2 receptors. Pharmacol. Res., 2021. [DOI | PubMed]
- N.D. Volkow, R.D. Baler, W.M. Compton, S.R. Weiss. Adverse health effects of marijuana use. N. Engl. J. Med., 2014. [DOI | PubMed]
- M.E. Badowski. A review of oral cannabinoids and medical marijuana for the treatment of chemotherapy-induced nausea and vomiting: A focus on pharmacokinetic variability and pharmacodynamics. Cancer Chemother. Pharmacol., 2017. [DOI | PubMed]
- T. Ng, V. Gupta. Tetrahydrocannabinol (THC). StatPearls, 2021
- P. Pacher, G. Kunos. Modulating the endocannabinoid system in human health and disease—Successes and failures. FEBS J., 2013. [DOI | PubMed]
- A. Conte, C. Vila Silván. Review of available data for the efficacy and effectiveness of nabiximols oromucosal spray (Sativex®) in multiple sclerosis patients with moderate to severe spasticity. Neurodegener. Dis., 2021. [DOI]
- R.B. Laprairie, A.M. Bagher, M.E. Kelly, E.M. Denovan-Wright. Cannabidiol is a negative allosteric modulator of the cannabinoid CB1 receptor. Br. J. Pharmacol., 2015. [DOI | PubMed]
- A. de la Harpe, N. Beukes, C.L. Frost. CBD activation of TRPV1 induces oxidative signaling and subsequent ER stress in breast cancer cell lines. Biotechnol. Appl. Biochem., 2021. [DOI]
- R. Mechoulam, M. Peters, E. Murillo-Rodriguez, L.O. Hanus. Cannabidiol—Recent advances. Chem. Biodivers., 2007. [DOI | PubMed]
- V.P. Soares, A.C. Campos. Evidences for the anti-panic actions of cannabidiol. Curr. Neuropharmacol., 2017. [DOI | PubMed]
- A.W. Zuardi, J.A. Crippa, J.E. Hallak, F.A. Moreira, F.S. Guimarães. Cannabidiol, a Cannabis sativa constituent, as an antipsychotic drug. Braz. J. Med. Biol. Res., 2006. [DOI | PubMed]
- J. Moltke, C. Hindocha. Reasons for cannabidiol use: A cross-sectional study of CBD users, focusing on self-perceived stress, anxiety, and sleep problems. J. Cannabis Res., 2021. [DOI | PubMed]
- N.A. Jones, A.J. Hill, I. Smith, S.A. Bevan, C.M. Williams, B.J. Whalley, G.J. Stephens. Cannabidiol displays antiepileptiform and antiseizure properties in vitro and in vivo. J. Pharmacol. Exp. Ther., 2010. [DOI | PubMed]
- E.J. Carrier, J.A. Auchampach, C.J. Hillard. Inhibition of an equilibrative nucleoside transporter by cannabidiol: A mechanism of cannabinoid immunosuppression. Proc. Natl. Acad. Sci. USA, 2006. [DOI | PubMed]
- O. Devinsky, M.R. Cilio, H. Cross, J. Fernandez-Ruiz, J. French, C. Hill, R. Katz, V. Di Marzo, D. Jutras-Aswad, W.G. Notcutt. Cannabidiol: Pharmacology and potential therapeutic role in epilepsy and other neuropsychiatric disorders. Epilepsia, 2014. [DOI | PubMed]
- S. Burstein. Cannabidiol (CBD) and its analogs: A review of their effects on inflammation. Bioorg. Med. Chem., 2015. [DOI | PubMed]
- B. Costa, M. Colleoni, S. Conti, D. Parolaro, C. Franke, A.E. Trovato, G. Giagnoni. Oral anti-inflammatory activity of cannabidiol, a non-psychoactive constituent of cannabis, in acute carrageenan-induced inflammation in the rat paw. Naunyn Schmiedebergs Arch. Pharm., 2004. [DOI | PubMed]
- B. Costa, A.E. Trovato, F. Comelli, G. Giagnoni, M. Colleoni. The non-psychoactive cannabis constituent cannabidiol is an orally effective therapeutic agent in rat chronic inflammatory and neuropathic pain. Eur. J. Pharmacol., 2007. [DOI | PubMed]
- S.R. Pereira, B. Hackett, D.N. O’Driscoll, M.C. Sun, E.J. Downer. Cannabidiol modulation of oxidative stress and signalling. Neuronal Signal., 2021. [DOI]
- M. Graczyk, A.A. Lewandowska, T. Dzierżanowski. The therapeutic potential of Cannabis in counteracting oxidative stress and inflammation. Molecules, 2021. [DOI | PubMed]
- S. Jeong, M.J. Jo, H.K. Yun, D.Y. Kim, B.R. Kim, J.L. Kim, S.H. Park, Y.J. Na, Y.A. Jeong, B.G. Kim. Cannabidiol promotes apoptosis via regulation of XIAP/Smac in gastric cancer. Cell Death Dis., 2019. [DOI | PubMed]
- H. Hamad, B.B. Olsen. Cannabidiol induces cell death in human lung cancer cells and cancer stem cells. Pharmaceuticals, 2021. [DOI | PubMed]
- A. Shrivastava, P.M. Kuzontkoski, J.E. Groopman, A. Prasad. Cannabidiol induces programmed cell death in breast cancer cells by coordinating the cross-talk between apoptosis and autophagy. Mol. Cancer Ther., 2011. [DOI | PubMed]
- O. Kovalchuk, I. Kovalchuk. Cannabinoids as anticancer therapeutic agents. Cell Cycle, 2020. [DOI | PubMed]
- A. Oláh, B.I. Tóth, I. Borbíró, K. Sugawara, A.G. Szöllõsi, G. Czifra, B. Pál, L. Ambrus, J. Kloepper, E. Camera. Cannabidiol exerts sebostatic and antiinflammatory effects on human sebocytes. J. Clin. Investig., 2014. [DOI | PubMed]
- A.J. Hampson, M. Grimaldi, M. Lolic, D. Wink, R. Rosenthal, J. Axelrod. Neuroprotective antioxidants from marijuana. Ann. N. Y. Acad. Sci., 2000. [DOI | PubMed]
- V. Di Giacomo, A. Chiavaroli, L. Recinella, G. Orlando, A. Cataldi, M. Rapino, V. Di Valerio, M. Ronci, S. Leone, L. Brunetti. Antioxidant and neuroprotective effects induced by cannabidiol and cannabigerol in rat CTX-TNA2 astrocytes and isolated cortexes. Int. J. Mol. Sci., 2020. [DOI | PubMed]
- S. Boyaji, J. Merkow, R.N.M. Elman, A.D. Kaye, R.J. Yong, R.D. Urman. The role of cannabidiol (CBD) in chronic pain management: An Assessment of current evidence. Curr. Pain Headache Rep., 2020. [DOI | PubMed]
- S. Bonaccorso, A. Ricciardi, C. Zangani, S. Chiappini, F. Schifano. Cannabidiol (CBD) use in psychiatric disorders: A systematic review. Neurotoxicology, 2019. [DOI | PubMed]
- M. Graczyk, M. Łukowicz, T. Dzierzanowski. Prospects for the use of cannabinoids in psychiatric disorders. Front. Psychiatry, 2021. [DOI | PubMed]
- M.M. Bergamaschi, R.H. Queiroz, A.W. Zuardi, J.A. Crippa. Safety and side effects of cannabidiol, a Cannabis sativa constituent. Curr. Drug Saf., 2011. [DOI | PubMed]
- O. Devinsky, J.H. Cross, L. Laux, E. Marsh, I. Miller, R. Nabbout, I.E. Scheffer, E.A. Thiele, S. Wright. Trial of cannabidiol for drug-resistant seizures in the Dravet Syndrome. N. Engl. J. Med., 2017. [DOI | PubMed]
- L. Taylor, B. Gidal, G. Blakey, B. Tayo, G. Morrison. A Phase I, Randomized, double-blind, placebo-controlled, single ascending dose, multiple dose, and food effect trial of the safety, tolerability and pharmacokinetics of highly purified cannabidiol in healthy subjects. CNS Drugs, 2018. [DOI | PubMed]
- K.B. Walsh, H.K. Andersen. Molecular pharmacology of synthetic cannabinoids: Delineating CB1 receptor-mediated cell signaling. Int. J. Mol. Sci., 2020. [DOI | PubMed]
- R. Gallily, Z. Yekhtin. Avidekel Cannabis extracts and cannabidiol are as efficient as Copaxone in suppressing EAE in SJL/J mice. Inflammopharmacology, 2019. [DOI | PubMed]
- A.M. Malfait, R. Gallily, P.F. Sumariwalla, A.S. Malik, E. Andreakos, R. Mechoulam, M. Feldmann. The nonpsychoactive cannabis constituent cannabidiol is an oral anti-arthritic therapeutic in murine collagen-induced arthritis. Proc. Natl. Acad. Sci. USA, 2000. [DOI | PubMed]
- D.M. Elliott, N. Singh, M. Nagarkatti, P.S. Nagarkatti. Cannabidiol attenuates experimental autoimmune encephalomyelitis model of multiple sclerosis through induction of myeloid-derived suppressor cells. Front. Immunol., 2018. [DOI | PubMed]
- L. Weiss, M. Zeira, S. Reich, S. Slavin, I. Raz, R. Mechoulam, R. Gallily. Cannabidiol arrests onset of autoimmune diabetes in NOD mice. Neuropharmacology, 2008. [DOI | PubMed]
- R. Gallily, Z. Yekhtin, L.O. Hanuš. Overcoming the bell-shaped dose-response of cannabidiol by using Cannabis extract enriched in cannabidiol. Pharmacol. Pharm., 2015. [DOI]
- Y. Gaoni, R. Mechoulam. The isolation and structure of delta-1-tetrahydrocannabinol and other neutral cannabinoids from hashish. J. Am. Chem. Soc., 1971. [DOI | PubMed]
- G. Fournier, C. Richez-Dumanois, J. Duvezin, J.P. Mathieu, M. Paris. Identification of a new chemotype in Cannabis sativa: Cannabigerol-dominant plants, biogenetic and agronomic prospects. Planta Med., 1987. [DOI | PubMed]
- K. Ioannidis, E. Dadiotis, V. Mitsis, E. Melliou, P. Magiatis. Biotechnological approaches on two high CBD and CBG Cannabis sativa L. (Cannabaceae) varieties: In vitro regeneration and phytochemical consistency evaluation of micropropagated plants using quantitative 1H-NMR. Molecules, 2020. [DOI]
- G. Navarro, K. Varani, I. Reyes-Resina, V. Sánchez de Medina, R. Rivas-Santisteban, C. Sánchez-Carnerero Callado, F. Vincenzi, S. Casano, C. Ferreiro-Vera, E.I. Canela. Cannabigerol action at cannabinoid CB1 and CB2 receptors and at CB1-CB2 heteroreceptor complexes. Front. Pharmacol., 2018. [DOI | PubMed]
- A.S. Husni, C.R. McCurdy, M.M. Radwan, S.A. Ahmed, D. Slade, S.A. Ross, M.A. ElSohly, S.J. Cutler. Evaluation of phytocannabinoids from high potency Cannabis sativa using in vitro bioassays to determine structure-activity relationships for cannabinoid receptor 1 and cannabinoid receptor 2. Med. Chem. Res., 2014. [DOI | PubMed]
- A. Zagzoog, K.A. Mohamed, H.J.J. Kim, E.D. Kim, C.S. Frank, T. Black, P.D. Jadhav, L.A. Holbrook, R.B. Laprairie. In vitro and in vivo pharmacological activity of minor cannabinoids isolated from Cannabis sativa. Sci. Rep., 2020. [DOI | PubMed]
- L. De Petrocellis, A. Ligresti, A.S. Moriello, M. Allarà, T. Bisogno, S. Petrosino, C.G. Stott, V. Di Marzo. Effects of cannabinoids and cannabinoid-enriched Cannabis extracts on TRP channels and endocannabinoid metabolic enzymes. Br. J. Pharmacol., 2011. [DOI | PubMed]
- M.G. Cascio, L.A. Gauson, L.A. Stevenson, R.A. Ross, R.G. Pertwee. Evidence that the plant cannabinoid cannabigerol is a highly potent alpha2-adrenoceptor agonist and moderately potent 5HT1A receptor antagonist. Br. J. Pharmacol., 2010. [DOI | PubMed]
- E. D’Aniello, T. Fellous, F.A. Iannotti, A. Gentile, M. Allarà, F. Balestrieri, R. Gray, P. Amodeo, R.M. Vitale, V. Di Marzo. Identification and characterization of phytocannabinoids as novel dual PPARα/γ agonists by a computational and in vitro experimental approach. Biochim. Biophys. Acta Gen. Subj., 2019. [DOI | PubMed]
- F. Borrelli, E. Pagano, B. Romano, S. Panzera, F. Maiello, D. Coppola, L. De Petrocellis, L. Buono, P. Orlando, A.A. Izzo. Colon carcinogenesis is inhibited by the TRPM8 antagonist cannabigerol, a Cannabis-derived non-psychotropic cannabinoid. Carcinogenesis, 2014. [DOI | PubMed]
- L. De Petrocellis, V. Vellani, A. Schiano-Moriello, P. Marini, P.C. Magherini, P. Orlando, V. Di Marzo. Plant-derived cannabinoids modulate the activity of transient receptor potential channels of ankyrin type-1 and melastatin type-8. J. Pharmacol. Exp. Ther., 2008. [DOI | PubMed]
- R. Nachnani, W.M. Raup-Konsavage, K.E. Vrana. The pharmacological case for cannabigerol. J. Pharmacol. Exp. Ther., 2021. [DOI | PubMed]
- E. Pagano, F.A. Iannotti, F. Piscitelli, B. Romano, G. Lucariello, T. Venneri, V. Di Marzo, A.A. Izzo, F. Borrelli. Efficacy of combined therapy with fish oil and phytocannabinoids in murine intestinal inflammation. Phytother. Res., 2021. [DOI | PubMed]
- L.R. Ruhaak, J. Felth, P.C. Karlsson, J.J. Rafter, R. Verpoorte, L. Bohlin. Evaluation of the cyclooxygenase inhibiting effects of six major cannabinoids isolated from Cannabis sativa. Biol. Pharm. Bull., 2011. [DOI | PubMed]
- A.J. Hill, C.M. Williams, B.J. Whalley, G.J. Stephens. Phytocannabinoids as novel therapeutic agents in CNS disorders. Pharmacol. Ther., 2012. [DOI | PubMed]
- F. Borrelli, I. Fasolino, B. Romano, R. Capasso, F. Maiello, D. Coppola, P. Orlando, G. Battista, E. Pagano, V. Di Marzo. Beneficial effect of the non-psychotropic plant cannabinoid cannabigerol on experimental inflammatory bowel disease. Biochem. Pharmacol., 2013. [DOI | PubMed]
- C.E. Turner, M.A. Elsohly. Biological activity of cannabichromene, its homologs and isomers. J. Clin. Pharmacol., 1981. [DOI | PubMed]
- S. Maione, F. Piscitelli, L. Gatta, D. Vita, L. De Petrocellis, E. Palazzo, V. de Novellis, V. Di Marzo. Non-psychoactive cannabinoids modulate the descending pathway of antinociception in anaesthetized rats through several mechanisms of action. Br. J. Pharmacol., 2011. [DOI | PubMed]
- B. Romano, F. Borrelli, I. Fasolino, R. Capasso, F. Piscitelli, M. Cascio, R. Pertwee, D. Coppola, L. Vassallo, P. Orlando. The cannabinoid TRPA1 agonist cannabichromene inhibits nitric oxide production in macrophages and ameliorates murine colitis. Br. J. Pharmacol., 2013. [DOI | PubMed]
- A.A. Izzo, R. Capasso, G. Aviello, F. Borrelli, B. Romano, F. Piscitelli, L. Gallo, F. Capasso, P. Orlando, V. Di Marzo. Inhibitory effect of cannabichromene, a major non-psychotropic cannabinoid extracted from Cannabis sativa, on inflammation-induced hypermotility in mice. Br. J. Pharmacol., 2012. [DOI | PubMed]
- N. Shinjyo, V. Di Marzo. The effect of cannabichromene on adult neural stem/progenitor cells. Neurochem. Int., 2013. [DOI | PubMed]
- A. Covelo, A. Eraso-Pichot, I. Fernández-Moncada, R. Serrat, G. Marsicano. CB1R-dependent regulation of astrocyte physiology and astrocyte-neuron interactions. Neuropharmacology, 2021. [DOI | PubMed]
- L.L. Anderson, A. Ametovski, J. Lin Luo, D. Everett-Morgan, I.S. McGregor, S.D. Banister, J.C. Arnold. Cannabichromene, related phytocannabinoids, and 5-fluoro-cannabichromene have anticonvulsant properties in a mouse model of Dravet Syndrome. ACS Chem. Neurosci., 2021. [DOI | PubMed]
- O. Anis, A.C. Vinayaka, N. Shalev, D. Namdar, S. Nadarajan, S.M. Anil, O. Cohen, E. Belausov, J. Ramon, E. Mayzlish Gati. Cannabis-derived compounds cannabichromene and Δ9-tetrahydrocannabinol interact and exhibit cytotoxic activity against urothelial cell carcinoma correlated with inhibition of cell migration and cytoskeleton organization. Molecules, 2021. [DOI | PubMed]
- L. De Petrocellis, A. Ligresti, A. Schiano Moriello, M. Iappelli, R. Verde, C.G. Stott, L. Cristino, P. Orlando, V. Di Marzo. Non-THC cannabinoids inhibit prostate carcinoma growth in vitro and in vivo: Pro-apoptotic effects and underlying mechanisms. Br. J. Pharmacol., 2013. [DOI | PubMed]
- G. Navarro, K. Varani, A. Lillo, F. Vincenzi, R. Rivas-Santisteban, I. Raïch, I. Reyes-Resina, C. Ferreiro-Vera, P.A. Borea, V. Sánchez de Medina. Pharmacological data of cannabidiol- and cannabigerol-type phytocannabinoids acting on cannabinoid CB1, CB2 and CB1/CB2 heteromer receptors. Pharmacol. Res., 2020. [DOI | PubMed]
- D. Bolognini, E.M. Rock, N.L. Cluny, M.G. Cascio, C.L. Limebeer, M. Duncan, C.G. Stott, F.A. Javid, L.A. Parker, R.G. Pertwee. Cannabidiolic acid prevents vomiting in Suncus murinus and nausea-induced behaviour in rats by enhancing 5-HT1A receptor activation. Br. J. Pharmacol., 2013. [DOI | PubMed]
- E.M. Rock, M.T. Sullivan, S.A. Collins, H. Goodman, C.L. Limebeer, R. Mechoulam, L.A. Parker. Evaluation of repeated or acute treatment with cannabidiol (CBD), cannabidiolic acid (CBDA) or CBDA methyl ester (HU-580) on nausea and/or vomiting in rats and shrews. Psychopharmacology, 2020. [DOI | PubMed]
- E.M. Rock, C.L. Limebeer, R.G. Pertwee, R. Mechoulam, L.A. Parker. Therapeutic potential of cannabidiol, cannabidiolic acid, and cannabidiolic acid methyl ester as treatments for nausea and vomiting. Cannabis Cannabinoid Res., 2021. [DOI | PubMed]
- R.G. Pertwee, E.M. Rock, K. Guenther, C.L. Limebeer, L.A. Stevenson, C. Haj, R. Smoum, L.A. Parker, R. Mechoulam. Cannabidiolic acid methyl ester, a stable synthetic analogue of cannabidiolic acid, can produce 5-HT1A receptor-mediated suppression of nausea and anxiety in rats. Br. J. Pharmacol., 2018. [DOI | PubMed]
- S. Takeda, K. Misawa, I. Yamamoto, K. Watanabe. Cannabidiolic acid as a selective cyclooxygenase-2 inhibitory component in cannabis. Drug Metab. Dispos., 2008. [DOI | PubMed]
- E.M. Rock, C.L. Limebeer, L.A. Parker. Effect of cannabidiolic acid and ∆9-tetrahydrocannabinol on carrageenan-induced hyperalgesia and edema in a rodent model of inflammatory pain. Psychopharmacology, 2018. [DOI | PubMed]
- L.L. Anderson, I.K. Low, S.D. Banister, I.S. McGregor, J.C. Arnold. Pharmacokinetics of phytocannabinoid acids and anticonvulsant effect of cannabidiolic acid in a mouse model of Dravet Syndrome. J. Nat. Prod., 2019. [DOI | PubMed]
- N. Assareh, A. Gururajan, C. Zhou, J.L. Luo, R.C. Kevin, J.C. Arnold. Cannabidiol disrupts conditioned fear expression and cannabidiolic acid reduces trauma-induced anxiety-related behaviour in mice. Behav. Pharmacol., 2020. [DOI | PubMed]
- E.M. Rock, C.L. Limebeer, G.N. Petrie, L.A. Williams, R. Mechoulam, L.A. Parker. Effect of prior foot shock stress and Δ9-tetrahydrocannabinol, cannabidiolic acid, and cannabidiol on anxiety-like responding in the light-dark emergence test in rats. Psychopharmacology, 2017. [DOI | PubMed]
- A. Smeriglio, S.V. Giofrè, E.M. Galati, M.T. Monforte, N. Cicero, V. D’Angelo, G. Grassi, C. Circosta. Inhibition of aldose reductase activity by Cannabis sativa chemotypes extracts with high content of cannabidiol or cannabigerol. Fitoterapia, 2018. [DOI | PubMed]
- L.L. Anderson, M. Heblinski, N.L. Absalom, N.A. Hawkins, M.T. Bowen, M.J. Benson, F. Zhang, D. Bahceci, P.T. Doohan, M. Chebib. Cannabigerolic acid, a major biosynthetic precursor molecule in cannabis, exhibits divergent effects on seizures in mouse models of epilepsy. Br. J. Pharmacol., 2021. [DOI | PubMed]
- D. Schubert, D. Kepchia, Z. Liang, R. Dargusch, J. Goldberg, P. Maher. Efficacy of cannabinoids in a pre-clinical drug-screening platform for Alzheimer’s disease. Mol. Neurobiol., 2019. [DOI | PubMed]
- Z. Liang, D. Soriano-Castell, D. Kepchia, B.M. Duggan, A. Currais, D. Schubert, P. Maher. Cannabinol inhibits oxytosis/ferroptosis by directly targeting mitochondria independently of cannabinoid receptors. Free Radic. Biol. Med., 2022. [DOI | PubMed]
- M. Aqawi, R.V. Sionov, R. Gallily, M. Friedman, D. Steinberg. Anti-biofilm activity of cannabigerol against Streptococcus mutans. Microorganisms, 2021. [DOI | PubMed]
- M. Aqawi, R.V. Sionov, R. Gallily, M. Friedman, D. Steinberg. Anti-bacterial properties of cannabigerol toward Streptococcus mutans. Front. Microbiol., 2021. [DOI | PubMed]
- B. Van Klingeren, M. Ten Ham. Antibacterial activity of delta9-tetrahydrocannabinol and cannabidiol. Antonie Van Leeuwenhoek, 1976. [DOI | PubMed]
- Y. Nalli, P. Arora, S. Riyaz-Ul-Hassan, A. Ali. Chemical investigation of Cannabis sativa leading to the discovery of a prenylspirodinone with anti-microbial potential. Tetrahedron Lett., 2018. [DOI]
- M.A.T. Blaskovich, A.M. Kavanagh, A.G. Elliott, B. Zhang, S. Ramu, M. Amado, G.J. Lowe, A.O. Hinton, D.M.T. Pham, J. Zuegg. The antimicrobial potential of cannabidiol. Commun. Biol., 2021. [DOI | PubMed]
- M. Aqawi, R. Gallily, R.V. Sionov, B. Zaks, M. Friedman, D. Steinberg. Cannabigerol prevents quorum sensing and biofilm formation of Vibrio harveyi. Front. Microbiol., 2020. [DOI | PubMed]
- D. Soni, R. Smoum, A. Breuer, R. Mechoulam, D. Steinberg. Effect of the synthetic cannabinoid HU-210 on quorum sensing and on the production of quorum sensing-mediated virulence factors by Vibrio harveyi. BMC Microbiol., 2015. [DOI | PubMed]
- U.S. Kosgodage, P. Matewele, B. Awamaria, I. Kraev, P. Warde, G. Mastroianni, A.V. Nunn, G.W. Guy, J.D. Bell, J.M. Inal. Cannabidiol is a novel modulator of bacterial membrane vesicles. Front. Cell. Infect. Microbiol., 2019. [DOI | PubMed]
- C.S. Wassmann, P. Højrup, J.K. Klitgaard. Cannabidiol is an effective helper compound in combination with bacitracin to kill Gram-positive bacteria. Sci. Rep., 2020. [DOI | PubMed]
- C. Russo, M. Lavorgna, R. Nugnes, E. Orlo, M. Isidori. Comparative assessment of antimicrobial, antiradical and cytotoxic activities of cannabidiol and its propyl analogue cannabidivarin. Sci. Rep., 2021. [DOI | PubMed]
- M. Feldman, R.V. Sionov, R. Mechoulam, D. Steinberg. Anti-biofilm activity of cannabidiol against Candida albicans. Microorganisms, 2021. [DOI | PubMed]
- M. Galletta, T.A. Reekie, G. Nagalingam, A.L. Bottomley, E.J. Harry, M. Kassiou, J.A. Triccas. Rapid Antibacterial activity of cannabichromenic acid against methicillin-resistant Staphylococcus aureus. Antibiotics, 2020. [DOI]
- T.K. Wood, S.J. Knabel, B.W. Kwan. Bacterial persister cell formation and dormancy. Appl. Environ. Microbiol., 2013. [DOI | PubMed]
- D.I. Andersson, D. Hughes. Persistence of antibiotic resistance in bacterial populations. FEMS Microbiol. Rev., 2011. [DOI | PubMed]
- J. Yan, B.L. Bassler. Surviving as a community: Antibiotic tolerance and persistence in bacterial biofilms. Cell Host Microbe, 2019. [DOI | PubMed]
- B. Aslam, W. Wang, M.I. Arshad, M. Khurshid, S. Muzammil, M.H. Rasool, M.A. Nisar, R.F. Alvi, M.A. Aslam, M.U. Qamar. Antibiotic resistance: A rundown of a global crisis. Infect. Drug Resist., 2018. [DOI | PubMed]
- D. Sharma, L. Misba, A.U. Khan. Antibiotics versus biofilm: An emerging battleground in microbial communities. Antimicrob. Resist. Infect. Control, 2019. [DOI | PubMed]
- O. Ciofu, C. Moser, P. Jensen, N. Høiby. Tolerance and resistance of microbial biofilms. Nat. Rev. Microbiol., 2022. [DOI]
- V. Stahl, K. Vasudevan. Comparison of efficacy of cannabinoids versus commercial oral care products in reducing bacterial content from dental plaque: A preliminary observation. Cureus, 2020. [DOI | PubMed]
- P. Bowler, C. Murphy, R. Wolcott. Biofilm exacerbates antibiotic resistance: Is this a current oversight in antimicrobial stewardship?. Antimicrob. Resist. Infect. Control, 2020. [DOI | PubMed]
- S. Singh, S. Datta, K.B. Narayanan, K.N. Rajnish. Bacterial exo-polysaccharides in biofilms: Role in antimicrobial resistance and treatments. J. Genet. Eng. Biotechnol., 2021. [DOI | PubMed]
- V. Raj, J.G. Park, K.H. Cho, P. Choi, T. Kim, J. Ham, J. Lee. Assessment of antiviral potencies of cannabinoids against SARS-CoV-2 using computational and in vitro approaches. Int. J. Biol. Macromol., 2021. [DOI | PubMed]
- R.B. van Breemen, R.N. Muchiri, T.A. Bates, J.B. Weinstein, H.C. Leier, S. Farley, F.G. Tafesse. Cannabinoids block cellular entry of SARS-CoV-2 and the emerging variants. J. Nat. Prod., 2022. [DOI | PubMed]
- N. Paland, A. Pechkovsky, M. Aswad, H. Hamza, T. Popov, E. Shahar, I. Louria-Hayon. The immunopathology of COVID-19 and the Cannabis paradigm. Front. Immunol., 2021. [DOI | PubMed]
- J.A.S. Crippa, J.C. Pacheco, A.W. Zuardi, F.S. Guimarães, A.C. Campos, F.L. Osório, S.R. Loureiro, R.G. Dos Santos, J.D.S. Souza, J.M. Ushirohira. Cannabidiol for COVID-19 patients with mild to moderate symptoms (CANDIDATE Study): A randomized, double-blind, placebo-controlled clinical trial. Cannabis Cannabinoid Res., 2021. [DOI]
- C.S. Reiss. Cannabinoids and viral infections. Pharmaceuticals, 2010. [DOI | PubMed]
- S. Zaami, A. Sirignano, Ó. García-Algar, E. Marinelli. COVID-19 pandemic, substance use disorders and body image issues, a worrisome correlation. Eur. Rev. Med. Pharmacol. Sci., 2022. [DOI | PubMed]
- D.C. Vidot, J.Y. Islam, C.-R. Marlene, M.B. Harrell, D.R. Rao, J.V. Chavez, G.O. Lucas, W.M. Hlaing, M. Weiner, S.E. Messiah. The COVID-19 cannabis health study: Results from an epidemiologic assessment of adults who use cannabis for medicinal reasons in the United States. J. Addict. Dis., 2021. [DOI | PubMed]
- A.S. Hatoum, C.L. Morrison, S.M.C. Colbert, E.A. Winiger, E.C. Johnson, A. Agrawal, R. Bogdan. Genetic liability to Cannabis use disorder and COVID-19 hospitalization. Biol. Psychiatry Glob. Open Sci., 2021. [DOI | PubMed]
- L. Pinzi, C. Lherbet, M. Baltas, F. Pellati, G. Rastelli. In silico repositioning of cannabigerol as a novel inhibitor of the enoyl acyl carrier protein (ACP) reductase (InhA). Molecules, 2019. [DOI | PubMed]
- M. Toyofuku, N. Nomura, L. Eberl. Types and origins of bacterial membrane vesicles. Nat. Rev. Microbiol., 2019. [DOI | PubMed]
- S. Mukherjee, B.L. Bassler. Bacterial quorum sensing in complex and dynamically changing environments. Nat. Rev. Microbiol., 2019. [DOI | PubMed]
- T.H. Burkey, R.M. Quock, P. Consroe, F.J. Ehlert, Y. Hosohata, W.R. Roeske, H.I. Yamamura. Relative efficacies of cannabinoid CB1 receptor agonists in the mouse brain. Eur. J. Pharmacol., 1997. [DOI | PubMed]
- A.C. Howlett, F. Barth, T.I. Bonner, G. Cabral, P. Casellas, W.A. Devane, C.C. Felder, M. Herkenham, K. Mackie, B.R. Martin. International Union of Pharmacology. XXVII. Classification of cannabinoid receptors. Pharmacol. Rev., 2002. [DOI | PubMed]
- T.W. Klein, C. Newton, K. Larsen, L. Lu, I. Perkins, L. Nong, H. Friedman. The cannabinoid system and immune modulation. J. Leukoc. Biol., 2003. [DOI | PubMed]
- K. Mackie, N. Stella. Cannabinoid receptors and endocannabinoids: Evidence for new players. AAPS J., 2006. [DOI | PubMed]
- E. Murillo-Rodríguez, H. Budde, A.B. Veras, N.B. Rocha, D. Telles-Correia, D. Monteiro, L. Cid, T. Yamamoto, S. Machado, P. Torterolo. The endocannabinoid system may modulate sleep disorders in aging. Curr. Neuropharmacol., 2020. [DOI | PubMed]
- C.H. Ashton, P.B. Moore. Endocannabinoid system dysfunction in mood and related disorders. Acta Psychiatr. Scand., 2011. [DOI | PubMed]
- I. Gallego-Landin, A. García-Baos, A. Castro-Zavala, O. Valverde. Reviewing the role of the endocannabinoid system in the pathophysiology of depression. Front. Pharmacol., 2021. [DOI | PubMed]
- M. Braile, S. Marcella, G. Marone, M.R. Galdiero, G. Varricchi, S. Loffredo. The interplay between the immune and the endocannabinoid systems in cancer. Cells, 2021. [DOI | PubMed]
- S.M.K. Rahman, T. Uyama, Z. Hussain, N. Ueda. Roles of endocannabinoids and endocannabinoid-like molecules in energy homeostasis and metabolic regulation: A nutritional perspective. Annu. Rev. Nutr., 2021. [DOI | PubMed]
- G. Marsicano, B. Lutz. Neuromodulatory functions of the endocannabinoid system. J. Endocrinol. Investig., 2006. [PubMed]
- P.D. Cani, H. Plovier, M. Van Hul, L. Geurts, N.M. Delzenne, C. Druart, A. Everard. Endocannabinoids—At the crossroads between the gut microbiota and host metabolism. Nat. Rev. Endocrinol., 2016. [DOI | PubMed]
- I. Ahmed, S.U. Rehman, S. Shahmohamadnejad, M.A. Zia, M. Ahmad, M.M. Saeed, Z. Akram, H.M.N. Iqbal, Q. Liu. Therapeutic attributes of endocannabinoid system against neuro-inflammatory autoimmune disorders. Molecules, 2021. [DOI | PubMed]
- A.R. Jackson, V.L. Hegde, P.S. Nagarkatti, M. Nagarkatti. Characterization of endocannabinoid-mediated induction of myeloid-derived suppressor cells involving mast cells and MCP-1. J. Leukoc. Biol., 2014. [DOI | PubMed]
- N. Osafo, O.K. Yeboah, A.O. Antwi. Endocannabinoid system and its modulation of brain, gut, joint and skin inflammation. Mol. Biol. Rep., 2021. [DOI | PubMed]
- R. Pandey, K. Mousawy, M. Nagarkatti, P. Nagarkatti. Endocannabinoids and immune regulation. Pharmacol. Res., 2009. [DOI | PubMed]
- J.M. Sido, P.S. Nagarkatti, M. Nagarkatti. Production of endocannabinoids by activated T cells and B cells modulates inflammation associated with delayed-type hypersensitivity. Eur. J. Immunol., 2016. [DOI | PubMed]
- O. Rahaman, D. Ganguly. Endocannabinoids in immune regulation and immunopathologies. Immunology, 2021. [DOI | PubMed]
- T. Aguado, K. Monory, J. Palazuelos, N. Stella, B. Cravatt, B. Lutz, G. Marsicano, Z. Kokaia, M. Guzmán, I. Galve-Roperh. The endocannabinoid system drives neural progenitor proliferation. FASEB J., 2005. [DOI | PubMed]
- J. Paraíso-Luna, J. Aguareles, R. Martín, A.C. Ayo-Martín, S. Simón-Sánchez, D. García-Rincón, C. Costas-Insua, E. García-Taboada, A. de Salas-Quiroga, J. Díaz-Alonso. Endocannabinoid signalling in stem cells and cerebral organoids drives differentiation to deep layer projection neurons via CB1 receptors. Development, 2020. [DOI | PubMed]
- W.B. Veldhuis, M. van Der Stelt, M.W. Wadman, G. van Zadelhoff, M. Maccarrone, F. Fezza, G.A. Veldink, J.F. Vliegenthart, P.R. Bär, K. Nicolay. Neuroprotection by the endogenous cannabinoid anandamide and arvanil against in vivo excitotoxicity in the rat: Role of vanilloid receptors and lipoxygenases. J. Neurosci., 2003. [DOI | PubMed]
- J. Palazuelos, T. Aguado, A. Egia, R. Mechoulam, M. Guzmán, I. Galve-Roperh. Non-psychoactive CB2 cannabinoid agonists stimulate neural progenitor proliferation. FASEB J., 2006. [DOI | PubMed]
- K. Jin, L. Xie, S.H. Kim, S. Parmentier-Batteur, Y. Sun, X.O. Mao, J. Childs, D.A. Greenberg. Defective adult neurogenesis in CB1 cannabinoid receptor knockout mice. Mol. Pharmacol., 2004. [DOI | PubMed]
- H.C. Lu, K. Mackie. An Introduction to the endogenous cannabinoid system. Biol. Psychiatry, 2016. [DOI | PubMed]
- F.A. Iannotti, V. Di Marzo. The gut microbiome, endocannabinoids and metabolic disorders. J. Endocrinol., 2021. [DOI | PubMed]
- A.C. Kreitzer. Neurotransmission: Emerging roles of endocannabinoids. Curr. Biol., 2005. [DOI | PubMed]
- V. Chevaleyre, K.A. Takahashi, P.E. Castillo. Endocannabinoid-mediated synaptic plasticity in the CNS. Annu. Rev. Neurosci., 2006. [DOI | PubMed]
- K. Mackie. Mechanisms of CB1 receptor signaling: Endocannabinoid modulation of synaptic strength. Int. J. Obes., 2006. [DOI | PubMed]
- B.D. Heifets, P.E. Castillo. Endocannabinoid signaling and long-term synaptic plasticity. Annu. Rev. Physiol., 2009. [DOI | PubMed]
- K.M. Jung, G. Astarita, C. Zhu, M. Wallace, K. Mackie, D. Piomelli. A key role for diacylglycerol lipase-alpha in metabotropic glutamate receptor-dependent endocannabinoid mobilization. Mol. Pharmacol., 2007. [DOI | PubMed]
- J. Tong, X. Liu, C. Vickstrom, Y. Li, L. Yu, Y. Lu, A.V. Smrcka, Q.S. Liu. The Epac-phospholipase Cε pathway regulates endocannabinoid signaling and cocaine-induced disinhibition of ventral tegmental area dopamine neurons. J. Neurosci., 2017. [DOI | PubMed]
- H. Jing, A. Reed, O.A. Ulanovskaya, J.S. Grigoleit, D.M. Herbst, C.L. Henry, H. Li, S. Barbas, J. Germain, K. Masuda. Phospholipase Cγ2 regulates endocannabinoid and eicosanoid networks in innate immune cells. Proc. Natl. Acad. Sci. USA, 2021. [DOI | PubMed]
- Y. Okamoto, J. Wang, J. Morishita, N. Ueda. Biosynthetic pathways of the endocannabinoid anandamide. Chem. Biodivers., 2007. [DOI | PubMed]
- E. Fride. Endocannabinoids in the central nervous system—An overview. Prostaglandins Leukot. Essent. Fat. Acids, 2002. [DOI]
- N. Stella, P. Schweitzer, D. Piomelli. A second endogenous cannabinoid that modulates long-term potentiation. Nature, 1997. [DOI | PubMed]
- R. Mechoulam, S. Ben-Shabat, L. Hanus, M. Ligumsky, N.E. Kaminski, A.R. Schatz, A. Gopher, S. Almog, B.R. Martin, D.R. Compton. Identification of an endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochem. Pharmacol., 1995. [DOI | PubMed]
- H.Y. Yang, F. Karoum, C. Felder, H. Badger, T.C. Wang, S.P. Markey. GC/MS analysis of anandamide and quantification of N-arachidonoylphosphatidylethanolamides in various brain regions, spinal cord, testis, and spleen of the rat. J. Neurochem., 1999. [DOI | PubMed]
- R.I. Staiano, S. Loffredo, F. Borriello, F.A. Iannotti, F. Piscitelli, P. Orlando, A. Secondo, F. Granata, M.T. Lepore, A. Fiorelli. Human lung-resident macrophages express CB1 and CB2 receptors whose activation inhibits the release of angiogenic and lymphangiogenic factors. J. Leukoc. Biol., 2016. [DOI | PubMed]
- V. Di Marzo. New approaches and challenges to targeting the endocannabinoid system. Nat. Rev. Drug Discov., 2018. [DOI | PubMed]
- V.K. Pestonjamasp, S.H. Burstein. Anandamide synthesis is induced by arachidonate mobilizing agonists in cells of the immune system. Biochim. Biophys. Acta, 1998. [DOI | PubMed]
- M. Mecha, A. Feliú, F.J. Carrillo-Salinas, A. Rueda-Zubiaurre, S. Ortega-Gutiérrez, R.G. de Sola, C. Guaza. Endocannabinoids drive the acquisition of an alternative phenotype in microglia. Brain Behav. Immun., 2015. [DOI | PubMed]
- P.M. Lam, T.H. Marczylo, J.C. Konje. Simultaneous measurement of three N-acylethanolamides in human bio-matrices using ultra performance liquid chromatography-tandem mass spectrometry. Anal. Bioanal. Chem., 2010. [DOI | PubMed]
- C.J. Hillard. Circulating endocannabinoids: From whence do they come and where are they going?. Neuropsychopharmacology, 2018. [DOI | PubMed]
- C.A. Opitz, N. Rimmerman, Y. Zhang, L.E. Mead, M.C. Yoder, D.A. Ingram, J.M. Walker, J. Rehman. Production of the endocannabinoids anandamide and 2-arachidonoylglycerol by endothelial progenitor cells. FEBS Lett., 2007. [DOI | PubMed]
- J. Liu, S. Batkai, P. Pacher, J. Harvey-White, J.A. Wagner, B.F. Cravatt, B. Gao, G. Kunos. Lipopolysaccharide induces anandamide synthesis in macrophages via CD14/MAPK/phosphoinositide 3-kinase/NF-kappaB independently of platelet-activating factor. J. Biol. Chem., 2003. [DOI | PubMed]
- L. Walter, A. Franklin, A. Witting, T. Moller, N. Stella. Astrocytes in culture produce anandamide and other acylethanolamides. J. Biol. Chem., 2002. [DOI | PubMed]
- C.J. Hillard, K.M. Weinlander, K.L. Stuhr. Contributions of endocannabinoid signaling to psychiatric disorders in humans: Genetic and biochemical evidence. Neuroscience, 2012. [DOI | PubMed]
- F.M. Leweke, D. Piomelli, F. Pahlisch, D. Muhl, C.W. Gerth, C. Hoyer, J. Klosterkötter, M. Hellmich, D. Koethe. Cannabidiol enhances anandamide signaling and alleviates psychotic symptoms of schizophrenia. Transl. Psychiatry, 2012. [DOI | PubMed]
- C.C. Felder, E.M. Briley, J. Axelrod, J.T. Simpson, K. Mackie, W.A. Devane. Anandamide, an endogenous cannabimimetic eicosanoid, binds to the cloned human cannabinoid receptor and stimulates receptor-mediated signal transduction. Proc. Natl. Acad. Sci. USA, 1993. [DOI | PubMed]
- W. Gonsiorek, C. Lunn, X. Fan, S. Narula, D. Lundell, R.W. Hipkin. Endocannabinoid 2-arachidonyl glycerol is a full agonist through human type 2 cannabinoid receptor: Antagonism by anandamide. Mol. Pharmacol., 2000. [PubMed]
- T. Luk, W. Jin, A. Zvonok, D. Lu, X.Z. Lin, C. Chavkin, A. Makriyannis, K. Mackie. Identification of a potent and highly efficacious, yet slowly desensitizing CB1 cannabinoid receptor agonist. Br. J. Pharmacol., 2004. [DOI | PubMed]
- T. Sugiura, S. Kondo, A. Sukagawa, S. Nakane, A. Shinoda, K. Itoh, A. Yamashita, K. Waku. 2-Arachidonoylglycerol: A possible endogenous cannabinoid receptor ligand in brain. Biochem. Biophys. Res. Commun., 1995. [DOI | PubMed]
- C. Muller, D.L. Lynch, D.P. Hurst, P.H. Reggio. TRPV1 activation by anandamide via a unique lipid pathway. J. Chem. Inf. Model., 2021. [DOI | PubMed]
- C. Muller, D.L. Lynch, D.P. Hurst, P.H. Reggio. A closer look at anandamide interaction with TRPV1. Front. Mol. Biosci., 2020. [DOI | PubMed]
- Y. Li, X. Chen, Y. Nie, Y. Tian, X. Xiao, F. Yang. Endocannabinoid activation of the TRPV1 ion channel is distinct from activation by capsaicin. J. Biol. Chem., 2021. [DOI | PubMed]
- V. Di Marzo, L. De Petrocellis. Endocannabinoids as regulators of transient receptor potential (TRP) channels: A further opportunity to develop new endocannabinoid-based therapeutic drugs. Curr. Med. Chem., 2010. [DOI | PubMed]
- A. Al-Hayani, K.N. Wease, R.A. Ross, R.G. Pertwee, S.N. Davies. The endogenous cannabinoid anandamide activates vanilloid receptors in the rat hippocampal slice. Neuropharmacology, 2001. [DOI | PubMed]
- D. Smart, M.J. Gunthorpe, J.C. Jerman, S. Nasir, J. Gray, A.I. Muir, J.K. Chambers, A.D. Randall, J.B. Davis. The endogenous lipid anandamide is a full agonist at the human vanilloid receptor (hVR1). Br. J. Pharmacol., 2000. [DOI | PubMed]
- J.E. Lauckner, J.B. Jensen, H.Y. Chen, H.C. Lu, B. Hille, K. Mackie. GPR55 is a cannabinoid receptor that increases intracellular calcium and inhibits M current. Proc. Natl. Acad. Sci. USA, 2008. [DOI | PubMed]
- E.C. Gootjes, L. Bakkerus, A.J. Ten Tije, P.O. Witteveen, T.E. Buffart, J.A. Bridgewater, J.N. Primrose, C. Verhoef, H.M.W. Verheul. The value of tumour debulking for patients with extensive multi-organ metastatic colorectal cancer. Eur. J. Cancer, 2018. [DOI | PubMed]
- L. De Petrocellis, A. Schiano Moriello, R. Imperatore, L. Cristino, K. Starowicz, V. Di Marzo. A re-evaluation of 9-HODE activity at TRPV1 channels in comparison with anandamide: Enantioselectivity and effects at other TRP channels and in sensory neurons. Br. J. Pharmacol., 2012. [DOI | PubMed]
- L. Cristino, T. Bisogno, V. Di Marzo. Cannabinoids and the expanded endocannabinoid system in neurological disorders. Nat. Rev. Neurol., 2020. [DOI | PubMed]
- P.M. Zygmunt, J. Petersson, D.A. Andersson, H. Chuang, M. Sørgård, V. Di Marzo, D. Julius, E.D. Högestätt. Vanilloid receptors on sensory nerves mediate the vasodilator action of anandamide. Nature, 1999. [DOI | PubMed]
- S.K. Lawton, F. Xu, A. Tran, E. Wong, A. Prakash, M. Schumacher, J. Hellman, K. Wilhelmsen. N-arachidonoyl dopamine modulates acute systemic inflammation via nonhematopoietic TRPV1. J. Immunol., 2017. [DOI | PubMed]
- I. Smaga, B. Bystrowska, D. Gawliński, E. Przegaliński, M. Filip. The endocannabinoid/endovanilloid system and depression. Curr. Neuropharmacol., 2014. [DOI | PubMed]
- L.A. Kasatkina, S. Rittchen, E.M. Sturm. Neuroprotective and immunomodulatory action of the endocannabinoid system under neuroinflammation. Int. J. Mol. Sci., 2021. [DOI | PubMed]
- Y.F. Lin. Potassium channels as molecular targets of endocannabinoids. Channels, 2021. [DOI | PubMed]
- B.E. Alger. Endocannabinoids: Getting the message across. Proc. Natl. Acad. Sci. USA, 2004. [DOI | PubMed]
- M.A. Diana, A. Marty. Endocannabinoid-mediated short-term synaptic plasticity: Depolarization-induced suppression of inhibition (DSI) and depolarization-induced suppression of excitation (DSE). Br. J. Pharmacol., 2004. [DOI | PubMed]
- A. Araque, P.E. Castillo, O.J. Manzoni, R. Tonini. Synaptic functions of endocannabinoid signaling in health and disease. Neuropharmacology, 2017. [DOI | PubMed]
- M. Kano, T. Ohno-Shosaku, Y. Hashimotodani, M. Uchigashima, M. Watanabe. Endocannabinoid-mediated control of synaptic transmission. Physiol. Rev., 2009. [DOI | PubMed]
- C. Di Scala, J. Fantini, N. Yahi, F.J. Barrantes, H. Chahinian. Anandamide Revisited: How cholesterol and ceramides control receptor-dependent and receptor-independent signal transmission pathways of a lipid neurotransmitter. Biomolecules, 2018. [DOI]
- E. Di Pasquale, H. Chahinian, P. Sanchez, J. Fantini. The insertion and transport of anandamide in synthetic lipid membranes are both cholesterol-dependent. PLoS ONE, 2009. [DOI | PubMed]
- C. Di Scala, M. Mazzarino, N. Yahi, K. Varini, N. Garmy, J. Fantini, H. Chahinian. Ceramide binding to anandamide increases its half-life and potentiates its cytotoxicity in human neuroblastoma cells. Chem. Phys. Lipids, 2017. [DOI | PubMed]
- P.I. Castagnet, M.Y. Golovko, G.C. Barceló-Coblijn, R.L. Nussbaum, E.J. Murphy. Fatty acid incorporation is decreased in astrocytes cultured from alpha-synuclein gene-ablated mice. J. Neurochem., 2005. [DOI | PubMed]
- M. Kaczocha, S.T. Glaser, D.G. Deutsch. Identification of intracellular carriers for the endocannabinoid anandamide. Proc. Natl. Acad. Sci. USA, 2009. [DOI | PubMed]
- M. Chivet, F. Hemming, K. Pernet-Gallay, S. Fraboulet, R. Sadoul. Emerging role of neuronal exosomes in the central nervous system. Front. Physiol., 2012. [DOI | PubMed]
- L. Huo, X. Du, X. Li, S. Liu, Y. Xu. The emerging role of neural cell-derived exosomes in intercellular communication in health and neurodegenerative diseases. Front. Neurosci., 2021. [DOI | PubMed]
- M. Gabrielli, N. Battista, L. Riganti, I. Prada, F. Antonucci, L. Cantone, M. Matteoli, M. Maccarrone, C. Verderio. Active endocannabinoids are secreted on extracellular membrane vesicles. EMBO Rep., 2015. [DOI | PubMed]
- E.J. Carrier, C.S. Kearn, A.J. Barkmeier, N.M. Breese, W. Yang, K. Nithipatikom, S.L. Pfister, W.B. Campbell, C.J. Hillard. Cultured rat microglial cells synthesize the endocannabinoid 2-arachidonylglycerol, which increases proliferation via a CB2 receptor-dependent mechanism. Mol. Pharmacol., 2004. [DOI | PubMed]
- N. Stella. Endocannabinoid signaling in microglial cells. Neuropharmacology, 2009. [DOI | PubMed]
- M. Navarrete, A. Araque. Endocannabinoids mediate neuron-astrocyte communication. Neuron, 2008. [DOI | PubMed]
- M.A. Karwad, T. Macpherson, B. Wang, E. Theophilidou, S. Sarmad, D.A. Barrett, M. Larvin, K.L. Wright, J.N. Lund, S.E. O’Sullivan. Oleoylethanolamine and palmitoylethanolamine modulate intestinal permeability in vitro via TRPV1 and PPARα. FASEB J., 2017. [DOI | PubMed]
- D.G. Couch, H. Cook, C. Ortori, D. Barrett, J.N. Lund, S.E. O’Sullivan. Palmitoylethanolamide and cannabidiol prevent inflammation-induced hyperpermeability of the human gut in vitro and in vivo—A randomized, placebo-controlled, double-blind controlled trial. Inflamm. Bowel Dis., 2019. [DOI | PubMed]
- F. Fezza, M. Bari, R. Florio, E. Talamonti, M. Feole, M. Maccarrone. Endocannabinoids, related compounds and their metabolic routes. Molecules, 2014. [DOI | PubMed]
- G.G. Muccioli, N. Stella. Microglia produce and hydrolyze palmitoylethanolamide. Neuropharmacology, 2008. [DOI | PubMed]
- N. Stella, D. Piomelli. Receptor-dependent formation of endogenous cannabinoids in cortical neurons. Eur. J. Pharmacol., 2001. [DOI | PubMed]
- M. Koch, S. Kreutz, C. Böttger, A. Benz, E. Maronde, C. Ghadban, H.W. Korf, F. Dehghani. Palmitoylethanolamide protects dentate gyrus granule cells via peroxisome proliferator-activated receptor-α. Neurotox. Res., 2011. [DOI | PubMed]
- S.D. Skaper, L. Facci, P. Giusti. Glia and mast cells as targets for palmitoylethanolamide, an anti-inflammatory and neuroprotective lipid mediator. Mol. Neurobiol., 2013. [DOI | PubMed]
- P. Clayton, S. Subah, R. Venkatesh, M. Hill, N. Bogoda. Palmitoylethanolamide: A potential alternative to cannabidiol. J. Diet. Suppl., 2021. [DOI]
- M. Vaia, S. Petrosino, D. De Filippis, L. Negro, A. Guarino, R. Carnuccio, V. Di Marzo, T. Iuvone. Palmitoylethanolamide reduces inflammation and itch in a mouse model of contact allergic dermatitis. Eur. J. Pharmacol., 2016. [DOI | PubMed]
- F. Borrelli, B. Romano, S. Petrosino, E. Pagano, R. Capasso, D. Coppola, G. Battista, P. Orlando, V. Di Marzo, A.A. Izzo. Palmitoylethanolamide, a naturally occurring lipid, is an orally effective intestinal anti-inflammatory agent. Br. J. Pharmacol., 2015. [DOI | PubMed]
- J.M. Keppel Hesselink, D.J. Kopsky. Palmitoylethanolamide, a neutraceutical, in nerve compression syndromes: Efficacy and safety in sciatic pain and carpal tunnel syndrome. J. Pain Res., 2015. [DOI | PubMed]
- A. Calignano, G. La Rana, A. Giuffrida, D. Piomelli. Control of pain initiation by endogenous cannabinoids. Nature, 1998. [DOI | PubMed]
- L. Facci, R. Dal Toso, S. Romanello, A. Buriani, S.D. Skaper, A. Leon. Mast cells express a peripheral cannabinoid receptor with differential sensitivity to anandamide and palmitoylethanolamide. Proc. Natl. Acad. Sci. USA, 1995. [DOI | PubMed]
- N. Ueda, K. Yamanaka, S. Yamamoto. Purification and characterization of an acid amidase selective for N-palmitoylethanolamine, a putative endogenous anti-inflammatory substance. J. Biol. Chem., 2001. [DOI | PubMed]
- V. Di Marzo, D. Melck, P. Orlando, T. Bisogno, O. Zagoory, M. Bifulco, Z. Vogel, L. De Petrocellis. Palmitoylethanolamide inhibits the expression of fatty acid amide hydrolase and enhances the anti-proliferative effect of anandamide in human breast cancer cells. Biochem. J., 2001. [DOI | PubMed]
- F. Guida, L. Luongo, S. Boccella, M.E. Giordano, R. Romano, G. Bellini, I. Manzo, A. Furiano, A. Rizzo, R. Imperatore. Palmitoylethanolamide induces microglia changes associated with increased migration and phagocytic activity: Involvement of the CB2 receptor. Sci. Rep., 2017. [DOI | PubMed]
- J. Lo Verme, J. Fu, G. Astarita, G. La Rana, R. Russo, A. Calignano, D. Piomelli. The nuclear receptor peroxisome proliferator-activated receptor-alpha mediates the anti-inflammatory actions of palmitoylethanolamide. Mol. Pharmacol., 2005. [DOI | PubMed]
- D. Piomelli. A fatty gut feeling. Trends Endocrinol. Metab., 2013. [DOI | PubMed]
- G.J. Schwartz, J. Fu, G. Astarita, X. Li, S. Gaetani, P. Campolongo, V. Cuomo, D. Piomelli. The lipid messenger OEA links dietary fat intake to satiety. Cell Metab., 2008. [DOI | PubMed]
- J. Fu, F. Oveisi, S. Gaetani, E. Lin, D. Piomelli. Oleoylethanolamide, an endogenous PPAR-alpha agonist, lowers body weight and hyperlipidemia in obese rats. Neuropharmacology, 2005. [DOI | PubMed]
- J. Sihag, P.J.H. Jones. Oleoylethanolamide: The role of a bioactive lipid amide in modulating eating behaviour. Obes. Rev., 2018. [DOI | PubMed]
- J.L. Shoemaker, B.K. Joseph, M.B. Ruckle, P.R. Mayeux, P.L. Prather. The endocannabinoid noladin ether acts as a full agonist at human CB2 cannabinoid receptors. J. Pharmacol. Exp. Ther., 2005. [DOI | PubMed]
- A.C. Porter, J.M. Sauer, M.D. Knierman, G.W. Becker, M.J. Berna, J. Bao, G.G. Nomikos, P. Carter, F.P. Bymaster, A.B. Leese. Characterization of a novel endocannabinoid, virodhamine, with antagonist activity at the CB1 receptor. J. Pharmacol. Exp. Ther., 2002. [DOI | PubMed]
- H. Sharir, L. Console-Bram, C. Mundy, S.N. Popoff, A. Kapur, M.E. Abood. The endocannabinoids anandamide and virodhamine modulate the activity of the candidate cannabinoid receptor GPR55. J. Neuroimmune Pharmacol., 2012. [DOI | PubMed]
- S.M. Huang, T. Bisogno, M. Trevisani, A. Al-Hayani, L. De Petrocellis, F. Fezza, M. Tognetto, T.J. Petros, J.F. Krey, C.J. Chu. An endogenous capsaicin-like substance with high potency at recombinant and native vanilloid VR1 receptors. Proc. Natl. Acad. Sci. USA, 2002. [DOI | PubMed]
- T. Bisogno, D. Melck, M. Bobrov, N.M. Gretskaya, V.V. Bezuglov, L. De Petrocellis, V. Di Marzo. N-acyl-dopamines: Novel synthetic CB(1) cannabinoid-receptor ligands and inhibitors of anandamide inactivation with cannabimimetic activity in vitro and in vivo. Biochem. J., 2000. [DOI | PubMed]
- W.R. Arnold, L.N. Carnevale, Z. Xie, J.L. Baylon, E. Tajkhorshid, H. Hu, A. Das. Anti-inflammatory dopamine- and serotonin-based endocannabinoid epoxides reciprocally regulate cannabinoid receptors and the TRPV1 channel. Nat. Commun., 2021. [DOI | PubMed]
- U. Grabiec, F. Dehghani. N-Arachidonoyl Dopamine: A novel endocannabinoid and endovanilloid with widespread physiological and pharmacological activities. Cannabis Cannabinoid Res., 2017. [DOI | PubMed]
- C.J. Chu, S.M. Huang, L. De Petrocellis, T. Bisogno, S.A. Ewing, J.D. Miller, R.E. Zipkin, N. Daddario, G. Appendino, V. Di Marzo. N-oleoyldopamine, a novel endogenous capsaicin-like lipid that produces hyperalgesia. J. Biol. Chem., 2003. [DOI | PubMed]
- T. Kino, T. Tomori, R. Abutarboush, P. Castri, Y. Chen, F.A. Lenz, R.M. McCarron, M. Spatz. Effect of N-arachidonoyl-L-serine on human cerebromicrovascular endothelium. Biochem. Biophys. Rep., 2016. [DOI | PubMed]
- G. Milman, Y. Maor, S. Abu-Lafi, M. Horowitz, R. Gallily, S. Batkai, F.M. Mo, L. Offertaler, P. Pacher, G. Kunos. N-arachidonoyl L-serine, an endocannabinoid-like brain constituent with vasodilatory properties. Proc. Natl. Acad. Sci. USA, 2006. [DOI | PubMed]
- X. Zhang, Y. Maor, J.F. Wang, G. Kunos, J.E. Groopman. Endocannabinoid-like N-arachidonoyl serine is a novel pro-angiogenic mediator. Br. J. Pharmacol., 2010. [DOI | PubMed]
- A. Cohen-Yeshurun, D. Willner, V. Trembovler, A. Alexandrovich, R. Mechoulam, E. Shohami, R.R. Leker. N-arachidonoyl-L-serine (AraS) possesses proneurogenic properties in vitro and in vivo after traumatic brain injury. J. Cereb. Blood Flow Metab., 2013. [DOI | PubMed]
- R.V. Sionov, M. Feldman, R. Smoum, R. Mechoulam, D. Steinberg. Anandamide prevents the adhesion of filamentous Candida albicans to cervical epithelial cells. Sci. Rep., 2020. [DOI | PubMed]
- N. Fornelos, E.A. Franzosa, J. Bishai, J.W. Annand, A. Oka, J. Lloyd-Price, T.D. Arthur, A. Garner, J. Avila-Pacheco, H.J. Haiser. Growth effects of N-acylethanolamines on gut bacteria reflect altered bacterial abundances in inflammatory bowel disease. Nat. Microbiol., 2020. [DOI | PubMed]
- N.Q. Balaban, J. Merrin, R. Chait, L. Kowalik, S. Leibler. Bacterial persistence as a phenotypic switch. Science, 2004. [DOI | PubMed]
- A.J. Lee, S. Wang, H.R. Meredith, B. Zhuang, Z. Dai, L. You. Robust, linear correlations between growth rates and β-lactam-mediated lysis rates. Proc. Natl. Acad. Sci. USA, 2018. [DOI | PubMed]
- E. Ferrer-González, H. Huh, H.M. Al-Tameemi, J.M. Boyd, S.H. Lee, D.S. Pilch. Impact of FtsZ inhibition on the localization of the penicillin binding proteins in methicillin-resistant Staphylococcus aureus. J. Bacteriol., 2021. [DOI | PubMed]
- S. Periasamy, H.S. Joo, A.C. Duong, T.H. Bach, V.Y. Tan, S.S. Chatterjee, G.Y. Cheung, M. Otto. How Staphylococcus aureus biofilms develop their characteristic structure. Proc. Natl. Acad. Sci. USA, 2012. [DOI | PubMed]
- K. Schwartz, A.K. Syed, R.E. Stephenson, A.H. Rickard, B.R. Boles. Functional amyloids composed of phenol soluble modulins stabilize Staphylococcus aureus biofilms. PLoS Pathog., 2012. [DOI | PubMed]
- K. Schwartz, M. Ganesan, D.E. Payne, M.J. Solomon, B.R. Boles. Extracellular DNA facilitates the formation of functional amyloids in Staphylococcus aureus biofilms. Mol. Microbiol., 2016. [DOI | PubMed]
- R. Russo, C. Cristiano, C. Avagliano, C. De Caro, G. La Rana, G.M. Raso, R.B. Canani, R. Meli, A. Calignano. Gut-brain axis: Role of lipids in the regulation of inflammation, pain and CNS diseases. Curr. Med. Chem., 2018. [DOI | PubMed]
- S. Lacroix, F. Pechereau, N. Leblanc, B. Boubertakh, A. Houde, C. Martin, N. Flamand, C. Silvestri, F. Raymond, V. Di Marzo. Rapid and concomitant gut microbiota and endocannabinoidome response to diet-induced obesity in mice. MSystems, 2019. [DOI | PubMed]
- J. Sihag, V. Di Marzo. (Wh)olistic (E)ndocannabinoidome-Microbiome-Axis modulation through (N)utrition (WHEN) to curb obesity and related disorders. Lipids Health Dis., 2022. [DOI | PubMed]
- J. Lian, I. Casari, M. Falasca. Modulatory role of the endocannabinoidome in the pathophysiology of the gastrointestinal tract. Pharmacol. Res., 2022. [DOI | PubMed]
- T. Bisogno, A. Lauritano, F. Piscitelli. The endocannabinoid system: A bridge between Alzheimer’s disease and gut microbiota. Life, 2021. [DOI | PubMed]
- R.N. Khan, K. Maner-Smith, J. AOwens, M.E. Barbian, R.M. Jones, C. RNaudin. At the heart of microbial conversations: Endocannabinoids and the microbiome in cardiometabolic risk. Gut Microbes, 2021. [DOI | PubMed]
- G. Berg, D. Rybakova, D. Fischer, T. Cernava, M.C. Vergès, T. Charles, X. Chen, L. Cocolin, K. Eversole, G.H. Corral. Microbiome definition re-visited: Old concepts and new challenges. Microbiome, 2020. [DOI | PubMed]
- K.H. Allin, V. Tremaroli, R. Caesar, B.A.H. Jensen, M.T.F. Damgaard, M.I. Bahl, T.R. Licht, T.H. Hansen, T. Nielsen, T.M. Dantoft. Aberrant intestinal microbiota in individuals with prediabetes. Diabetologia, 2018. [DOI | PubMed]
- M. Valles-Colomer, G. Falony, Y. Darzi, E.F. Tigchelaar, J. Wang, R.Y. Tito, C. Schiweck, A. Kurilshikov, M. Joossens, C. Wijmenga. The neuroactive potential of the human gut microbiota in quality of life and depression. Nat. Microbiol., 2019. [DOI | PubMed]
- S.G. Sorboni, H.S. Moghaddam, R. Jafarzadeh-Esfehani, S. Soleimanpour. A comprehensive review on the role of the gut microbiome in human neurological disorders. Clin. Microbiol. Rev., 2022. [DOI | PubMed]
- L.M.T. Dicks, D. Hurn, D. Hermanus. Gut bacteria and neuropsychiatric disorders. Microorganisms, 2021. [DOI | PubMed]
- J.R. Kelly, C. Minuto, J.F. Cryan, G. Clarke, T.G. Dinan. The role of the gut microbiome in the development of schizophrenia. Schizophr. Res., 2021. [DOI | PubMed]
- L. Giloteaux, J.K. Goodrich, W.A. Walters, S.M. Levine, R.E. Ley, M.R. Hanson. Reduced diversity and altered composition of the gut microbiome in individuals with myalgic encephalomyelitis/chronic fatigue syndrome. Microbiome, 2016. [DOI | PubMed]
- A. Minichino, M.A. Jackson, M. Francesconi, C.J. Steves, C. Menni, P.W.J. Burnet, B.R. Lennox. Endocannabinoid system mediates the association between gut-microbial diversity and anhedonia/amotivation in a general population cohort. Mol. Psychiatry, 2021. [DOI | PubMed]
- G. Chevalier, E. Siopi, L. Guenin-Macé, M. Pascal, T. Laval, A. Rifflet, I.G. Boneca, C. Demangel, B. Colsch, A. Pruvost. Effect of gut microbiota on depressive-like behaviors in mice is mediated by the endocannabinoid system. Nat. Commun., 2020. [DOI | PubMed]
- H. Li, F. Liu, J. Lu, J. Shi, J. Guan, F. Yan, B. Li, G. Huo. Probiotic mixture of Lactobacillus plantarum strains improves lipid metabolism and gut microbiota structure in high fat diet-fed mice. Front. Microbiol., 2020. [DOI | PubMed]
- N. Xie, Y. Cui, Y.N. Yin, X. Zhao, J.W. Yang, Z.G. Wang, N. Fu, Y. Tang, X.H. Wang, X.W. Liu. Effects of two Lactobacillus strains on lipid metabolism and intestinal microflora in rats fed a high-cholesterol diet. BMC Complement. Altern. Med., 2011. [DOI | PubMed]
- C. Rousseaux, X. Thuru, A. Gelot, N. Barnich, C. Neut, L. Dubuquoy, C. Dubuquoy, E. Merour, K. Geboes, M. Chamaillard. Lactobacillus acidophilus modulates intestinal pain and induces opioid and cannabinoid receptors. Nat. Med., 2007. [DOI | PubMed]
- M. Alhouayek, G.G. Muccioli. Harnessing the anti-inflammatory potential of palmitoylethanolamide. Drug Discov. Today, 2014. [DOI | PubMed]
- M. Coppola, R. Mondola. Is there a role for palmitoylethanolamide in the treatment of depression?. Med. Hypotheses, 2014. [DOI | PubMed]
- T. Zimmermann, J.C. Bartsch, A. Beer, E. Lomazzo, S. Guggenhuber, M.D. Lange, L. Bindila, H.C. Pape, B. Lutz. Impaired anandamide/palmitoylethanolamide signaling in hippocampal glutamatergic neurons alters synaptic plasticity, learning, and emotional responses. Neuropsychopharmacology, 2019. [DOI | PubMed]
- F. Guida, S. Boccella, C. Belardo, M. Iannotta, F. Piscitelli, F. De Filippis, S. Paino, F. Ricciardi, D. Siniscalco, I. Marabese. Altered gut microbiota and endocannabinoid system tone in vitamin D deficiency-mediated chronic pain. Brain Behav. Immun., 2020. [DOI | PubMed]
- C. Cristiano, C. Pirozzi, L. Coretti, G. Cavaliere, A. Lama, R. Russo, F. Lembo, M.P. Mollica, R. Meli, A. Calignano. Palmitoylethanolamide counteracts autistic-like behaviours in BTBR T+tf/J mice: Contribution of central and peripheral mechanisms. Brain Behav. Immun., 2018. [DOI | PubMed]
- M. Sultan, K. Wilson, O.A. Abdulla, P.B. Busbee, A. Hall, T. Carter, N. Singh, S. Chatterjee, P. Nagarkatti, M. Nagarkatti. Endocannabinoid anandamide attenuates acute respiratory distress syndrome through modulation of microbiome in the gut-lung axis. Cells, 2021. [DOI | PubMed]
- G.G. Muccioli, D. Naslain, F. Bäckhed, C.S. Reigstad, D.M. Lambert, N.M. Delzenne, P.D. Cani. The endocannabinoid system links gut microbiota to adipogenesis. Mol. Syst. Biol., 2010. [DOI | PubMed]
- M.A. Storr, B. Yüce, C.N. Andrews, K.A. Sharkey. The role of the endocannabinoid system in the pathophysiology and treatment of irritable bowel syndrome. Neurogastroenterol. Motil., 2008. [DOI | PubMed]
- M. Camilleri. Cannabinoids and gastrointestinal motility: Pharmacology, clinical effects, and potential therapeutics in humans. Neurogastroenterol. Motil., 2018. [DOI | PubMed]
- R. Abalo, G. Vera, A.E. López-Pérez, M. Martínez-Villaluenga, M.I. Martín-Fontelles. The gastrointestinal pharmacology of cannabinoids: Focus on motility. Pharmacology, 2012. [DOI | PubMed]
- D. Osei-Hyiaman, M. DePetrillo, P. Pacher, J. Liu, S. Radaeva, S. Bátkai, J. Harvey-White, K. Mackie, L. Offertáler, L. Wang. Endocannabinoid activation at hepatic CB1 receptors stimulates fatty acid synthesis and contributes to diet-induced obesity. J. Clin. Investig., 2005. [DOI | PubMed]
- D. Osei-Hyiaman, J. Liu, L. Zhou, G. Godlewski, J. Harvey-White, W.I. Jeong, S. Bátkai, G. Marsicano, B. Lutz, C. Buettner. Hepatic CB1 receptor is required for development of diet-induced steatosis, dyslipidemia, and insulin and leptin resistance in mice. J. Clin. Investig., 2008. [DOI | PubMed]
- C. Ravinet Trillou, C. Delgorge, C. Menet, M. Arnone, P. Soubrié. CB1 cannabinoid receptor knockout in mice leads to leanness, resistance to diet-induced obesity and enhanced leptin sensitivity. Int. J. Obes. Relat. Metab. Disord., 2004. [DOI | PubMed]
- L. Hoareau, M. Buyse, F. Festy, P. Ravanan, M.P. Gonthier, I. Matias, S. Petrosino, F. Tallet, C.L. D’Hellencourt, M. Cesari. Anti-inflammatory effect of palmitoylethanolamide on human adipocytes. Obesity, 2009. [DOI | PubMed]
- V. Di Marzo, T. Bisogno, L. De Petrocellis, D. Melck, P. Orlando, J.A. Wagner, G. Kunos. Biosynthesis and inactivation of the endocannabinoid 2-arachidonoylglycerol in circulating and tumoral macrophages. Eur. J. Biochem., 1999. [DOI | PubMed]
- M. Maccarrone, L. De Petrocellis, M. Bari, F. Fezza, S. Salvati, V. Di Marzo, A. Finazzi-Agrò. Lipopolysaccharide downregulates fatty acid amide hydrolase expression and increases anandamide levels in human peripheral lymphocytes. Arch. Biochem. Biophys., 2001. [DOI | PubMed]
- N.V. Di Patrizio. Endocannabinoids in the gut. Cannabis Cannabinoid Res., 2016. [DOI | PubMed]
- P. Mehrpouya-Bahrami, K.N. Chitrala, M.S. Ganewatta, C. Tang, E.A. Murphy, R.T. Enos, K.T. Velazquez, J. McCellan, M. Nagarkatti, P. Nagarkatti. Blockade of CB1 cannabinoid receptor alters gut microbiota and attenuates inflammation and diet-induced obesity. Sci. Rep., 2017. [DOI | PubMed]
- A. Gross, A. Terraza, J. Marchant, M. Bouaboula, S. Ouahrani-Bettache, J.P. Liautard, P. Casellas, J. Dornand. A beneficial aspect of a CB1 cannabinoid receptor antagonist: SR141716A is a potent inhibitor of macrophage infection by the intracellular pathogen Brucella suis. J. Leukoc. Biol., 2000. [DOI | PubMed]
- M. Zhang, J. Lu, X. Duan, J. Chen, X. Jin, Z. Lin, Y. Pang, X. Wang, H. Lou, W. Chang. Rimonabant potentiates the antifungal activity of amphotericin B by increasing cellular oxidative stress and cell membrane permeability. FEMS Yeast Res., 2021. [DOI | PubMed]
- H.Y. Kim, A.A. Spector. N-Docosahexaenoylethanolamine: A neurotrophic and neuroprotective metabolite of docosahexaenoic acid. Mol. Asp. Med., 2018. [DOI]
- M. Di Paola, E. Bonechi, G. Provensi, A. Costa, G. Clarke, C. Ballerini, C. De Filippo, M.B. Passani. Oleoylethanolamide treatment affects gut microbiota composition and the expression of intestinal cytokines in Peyer’s patches of mice. Sci. Rep., 2018. [DOI | PubMed]
- S. Hryhorowicz, M. Kaczmarek-Ryś, A. Zielińska, R.J. Scott, R. Słomski, A. Pławski. Endocannabinoid system as a promising therapeutic target in inflammatory bowel disease—A systematic review. Front. Immunol., 2021. [DOI | PubMed]
- G. Esposito, E. Capoccia, F. Turco, I. Palumbo, J. Lu, A. Steardo, R. Cuomo, G. Sarnelli, L. Steardo. Palmitoylethanolamide improves colon inflammation through an enteric glia/toll like receptor 4-dependent PPAR-α activation. Gut, 2014. [DOI | PubMed]
- M.A. Engel, C.A. Kellermann, T. Rau, G. Burnat, E.G. Hahn, P.C. Konturek. Ulcerative colitis in AKR mice is attenuated by intraperitoneally administered anandamide. J. Physiol. Pharmacol., 2008. [PubMed]
- G. D’Argenio, M. Valenti, G. Scaglione, V. Cosenza, I. Sorrentini, V. Di Marzo. Up-regulation of anandamide levels as an endogenous mechanism and a pharmacological strategy to limit colon inflammation. FASEB J., 2006. [DOI | PubMed]
- M.A. Storr, C.M. Keenan, D. Emmerdinger, H. Zhang, B. Yüce, A. Sibaev, F. Massa, N.E. Buckley, B. Lutz, B. Göke. Targeting endocannabinoid degradation protects against experimental colitis in mice: Involvement of CB1 and CB2 receptors. J. Mol. Med., 2008. [DOI | PubMed]
- P. Louis, G.L. Hold, H.J. Flint. The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol., 2014. [DOI | PubMed]
- A. Vijay, A. Kouraki, S. Gohir, J. Turnbull, A. Kelly, V. Chapman, D.A. Barrett, W.J. Bulsiewicz, A.M. Valdes. The anti-inflammatory effect of bacterial short chain fatty acids is partially mediated by endocannabinoids. Gut Microbes, 2021. [DOI | PubMed]
- S. Redlich, S. Ribes, S. Schütze, D. Czesnik, R. Nau. Palmitoylethanolamide stimulates phagocytosis of Escherichia coli K1 and Streptococcus pneumoniae R6 by microglial cells. J. Neuroimmunol., 2012. [DOI | PubMed]
- S. Redlich, S. Ribes, S. Schütze, R. Nau. Palmitoylethanolamide stimulates phagocytosis of Escherichia coli K1 by macrophages and increases the resistance of mice against infections. J. Neuroinflammation, 2014. [DOI | PubMed]
- E.C. Heide, L. Bindila, J.M. Post, D. Malzahn, B. Lutz, J. Seele, R. Nau, S. Ribes. Prophylactic palmitoylethanolamide prolongs survival and decreases detrimental inflammation in aged mice with bacterial meningitis. Front. Immunol., 2018. [DOI | PubMed]
- J. Lloyd-Price, C. Arze, A.N. Ananthakrishnan, M. Schirmer, J. Avila-Pacheco, T.W. Poon, E. Andrews, N.J. Ajami, K.S. Bonham, C.J. Brislawn. Multi-omics of the gut microbial ecosystem in inflammatory bowel diseases. Nature, 2019. [DOI | PubMed]
- E.A. Franzosa, A. Sirota-Madi, J. Avila-Pacheco, N. Fornelos, H.J. Haiser, S. Reinker, T. Vatanen, A.B. Hall, H. Mallick, L.J. McIver. Gut microbiome structure and metabolic activity in inflammatory bowel disease. Nat. Microbiol., 2019. [DOI | PubMed]
- A. Wahlström, S.I. Sayin, H.U. Marschall, F. Bäckhed. Intestinal crosstalk between bile acids and microbiota and its impact on host metabolism. Cell Metab., 2016. [DOI | PubMed]
- B. Rémy, S. Mion, L. Plener, M. Elias, E. Chabrière, D. Daudé. Interference in bacterial quorum sensing: A biopharmaceutical perspective. Front. Pharmacol., 2018. [DOI | PubMed]