
Pharmacological Insights into Cannabidiol for Wound Healing and Bone Regeneration
Abstract
Cannabidiol (CBD), a major nonpsychoactive phytocannabinoid derived from Cannabis sativa L., has recently gained prominence for its broad pharmacological profile and emerging applications in regenerative medicine. Beyond its well-established neuroprotective, antiepileptic, anxiolytic, antipsychotic, anti-inflammatory, analgesic, and anticancer effects, CBD has demonstrated the capacity to modulate key biological processes involved in tissue repair. Increasing evidence indicates that CBD promotes wound healing by regulating inflammatory responses, cellular proliferation, and extracellular matrix remodeling through interactions with cannabinoid and noncannabinoid receptors expressed in neural, immune, and epithelial cells. Notably, these receptors are also present in osteogenic and progenitor cells, suggesting that CBD may influence bone metabolism and regeneration. Recent preclinical studies have reported that CBD enhances osteoblastic differentiation, angiogenesis, and matrix mineralization, highlighting its potential as a bioactive molecule for bone tissue engineering. Within the dental field, such properties open new perspectives for the development of CBD-based biomaterials aimed at improving osseointegration, soft tissue healing, and the overall biological performance of implantable devices. Accordingly, this review aims to provide a comprehensive overview of the pharmacological and molecular mechanisms underlying the effects of CBD on wound healing and bone regeneration. Furthermore, it discusses dose–response relationships, delivery routes, formulation strategies, and the current legal and regulatory frameworks influencing CBD translation into clinical dental applications. These insights may support the rational design of next-generation bioactive materials incorporating CBD for oral and maxillofacial regenerative therapies.
Affiliations: † Department of Diagnosis and Surgery, School of Dentistry, São Paulo State University (UNESP), 14801-903 Araraquara, Brazil; ‡ Department of Physiology and Pathology, School of Dentistry, São Paulo State University (UNESP), 14801-903 Araraquara, Brazil; § Department of Dental Materials and Prosthodontics, São Paulo State University (UNESP), 16015-050 Araçatuba, Brazil; ∥ Postgraduate Program in Dentistry, Federal University of Piaui, 64049-550 Teresina, Brazil; ⊥ Department of Diagnostic and Surgery, Araçatuba School of Dentistry, São Paulo State University (UNESP), 16015-050 Araçatuba, Brazil; # Department of Dental Materials and Prosthodontics, São Paulo State University (UNESP), 14801-903 Araraquara, Brazil; ∇ School of Dentistry, Universidad Católica de Santa María (UCSM), 04013 Arequipa, Peru
License: © 2026 The Authors. Published by American Chemical Society CC BY 4.0 This article is licensed under CC-BY 4.0
Article links: DOI: 10.1021/acsomega.5c13259 | PMC: PMC13129851
Relevance: Relevant: mentioned in keywords or abstract
Full text: PDF (7.4 MB)
Introduction
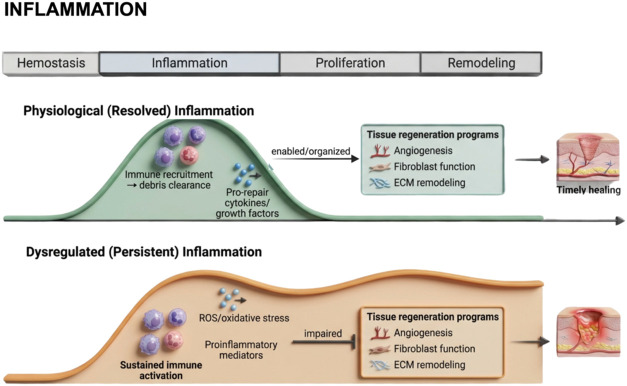
Tissue injury initiates a highly orchestrated and dynamic sequence of cellular, molecular, and biochemical events designed to restore structural integrity and functional homeostasis.ref1,ref2 Successful wound healing relies on the precise coordination of four interdependent and overlapping phases, hemostasis, inflammation, proliferation, and remodeling or resolution, each governed by a complex interplay of immune cells, cytokines, and growth factors.ref. ref3 The timely and balanced transition between these phases is critical for effective repair. However, dysregulation of the immune response can disrupt this sequence, leading to chronic inflammation, and impaired regeneration.ref. ref4
In the skeletal system, excessive or unresolved inflammation exerts particularly detrimental effects. An exacerbated inflammatory milieu can stimulate osteoclastogenesis, enhancing bone resorption and compromising the stability of the surrounding tissue.ref. ref4 Bone defects frequently arise from trauma, infection, tumor resection, or chronic inflammatory diseases and may surpass the intrinsic regenerative capacity of the host.ref. ref5 In extensive or complex lesions, spontaneous healing becomes unfeasible due to persistent inflammation, inadequate vascularization, and the absence of osteogenic or osteoinductive cues required for new bone formation.ref. ref5 Such critical-size defects, defined as defects incapable of spontaneous healing even after surgical stabilization, require adjunctive therapeutic strategies, including autologous bone grafting or engineered biomaterials to restore structural and functional continuity.ref. ref6 Consequently, bone regeneration remains one of the most demanding challenges in regenerative medicine, influenced by anatomical location, defect size, and local biological conditions.ref. ref7
To counteract inflammation-associated tissue destruction and promote regenerative healing, a variety of therapeutic strategies have been developed.ref8,ref9 Conventional interventions include the debridement of necrotic tissue, rigorous infection control, surgical revascularization, and bone grafting procedures employing autologous, allogeneic, or synthetic substitutes.ref8,ref9 Although clinically effective, these approaches are frequently limited by donor-site morbidity, immune incompatibility, infection risk, and suboptimal osteogenic potential. As a result, increasing attention has been directed toward alternative or complementary strategies, particularly those involving bioactive natural compounds and biomaterials endowed with intrinsic regenerative and immunomodulatory properties.ref10−ref11ref12
Among these emerging therapeutic approaches, the medicinal potential of Cannabis sativa L. has attracted renewed scientific interest. Historical evidence documents its therapeutic application as early as 1550 BCE in ancient Egyptian papyri.ref. ref13 The plant’s remarkable ecological adaptability has enabled its widespread cultivation across diverse geographic regions, leading to the identification of more than 450 bioactive constituents, including approximately 70 phytocannabinoids.ref14,ref15 Structurally, these compounds share a conserved dibenzopyran core linked to a hydrophobic alkyl side chain, a configuration that confers marked lipophilicity and facilitates their interaction with membrane-bound receptorsref. ref16 (Figure ).
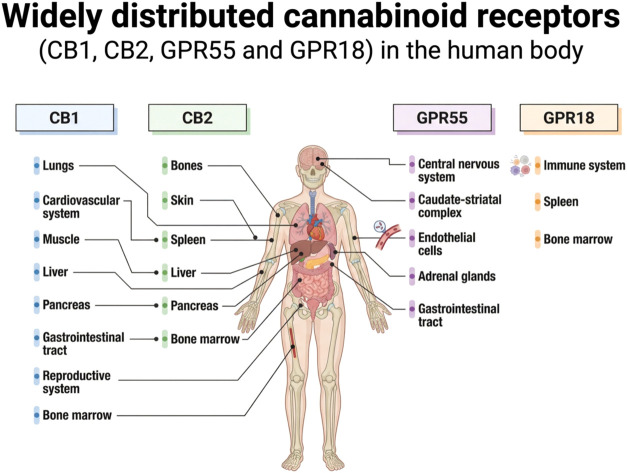
The biological effects of phytocannabinoids are primarily mediated through the endocannabinoid system (ECS), a complex signaling network composed of endogenous ligands, metabolic enzymes, and cannabinoid receptors CB1 and CB2.ref. ref17 CB1 receptors are predominantly expressed in the central and peripheral nervous systems but are also detected in peripheral tissues, including bone, adipose tissue, myocardium, reproductive organs, and retina.ref. ref18 CB2 receptors are more abundant in immune cells, yet they are also present in the liver, gastrointestinal tract, brain, and skeletal tissues.ref. ref19 Importantly, both CB1 and CB2 receptors have been identified in bone marrow–derived stromal cells and within the bone immune microenvironment.ref. ref20 Experimental evidence indicates that pharmacological activation or genetic modulation of these receptors influences osteoblast and osteoclast activity, attenuates pathological bone turnover, and regulates remodeling dynamicsref. ref20 (Figure ).
Beyond this classical CB1/CB2 framework, increasing evidence supports the existence of an expanded endocannabinoid network that includes noncanonical receptors operating through CB1/CB2-independent pathways.ref. ref21 Among these, GPR18 and GPR55 have emerged as putative cannabinoid-responsive receptors capable of being modulated by endocannabinoids, phytocannabinoids, and synthetic ligands. GPR18, also referred to as the N-arachidonoylglycine (NAGly) receptor, is predominantly expressed in immune cells and plays a role in neutrophil migration, macrophage phenotype modulation, and lymphocyte maturation.ref. ref22 It is also highly expressed in microglial cells, which are key components of the central nervous system immune network, where it participates in chemotactic signaling and leukocyte recruitment to sites of injury.ref. ref19 Cannabidiol (CBD) has been described as a modulator of orphan receptors, including GPR18, acting as an antagonist or inverse agonist under certain conditions. These interactions may partly explain CBD’s anti-inflammatory properties independent of CB1 activation, thereby avoiding the psychotropic effects associated with Δ9-tetrahydrocannabinol (Δ9-THC).ref. ref19
GPR55, in contrast, signals primarily via G 12/13 and G q proteins rather than Gi-coupled mechanisms typical of CB receptors. This distinction leads to activation of intracellular pathways such as RhoA and calcium mobilization, processes involved in cell proliferation, migration, and angiogenesis.ref. ref23 GPR55 expression has been identified in multiple central nervous system regions, including the caudate-striatal complex, as well as in peripheral tissues such as endothelial cells, adrenal glands, and the gastrointestinal tract.ref. ref24 Its wide distribution suggests involvement in innate and adaptive immune regulation.ref. ref25 Collectively, these findings underscore the complexity of cannabinoid-related signaling and reinforce the concept that phytocannabinoid effects extend beyond the classical CB1/CB2 paradigm, engaging a broader receptor repertoire relevant to immune modulation and tissue homeostasis.
Among phytocannabinoids, Δ9-THC and cannabidiol (CBD) are the most extensively studied due to their distinct pharmacological profiles.ref. ref26 Δ9-THC, isolated in 1964, is the principal psychoactive constituent of the plant and exerts its central effects primarily via CB1 activation.ref27,ref28 In contrast, CBD is nonpsychoactive and exhibits low affinity for CB1 and CB2 receptors. Instead, it modulates the ECS indirectly and interacts with additional molecular targets, including TRPV1, PPARγ, and 5-HT1A receptors.ref. ref29
CBD has garnered substantial attention due to its wide range of reported therapeutic effects, encompassing anti-inflammatory, antioxidant, neuroprotective, and regenerative properties.ref30,ref31 These pleiotropic actions suggest that CBD may act as a multimodal agent capable of modulating several signaling pathways relevant to tissue homeostasis and repair. From a regenerative perspective, an ideal bioactive compound should attenuate inflammation and oxidative stress, exhibit antimicrobial activity, and promote cellular proliferation, matrix deposition, and angiogenesis.ref32,ref33 In this context, preclinical evidence has demonstrated the pro-healing capacity of CBD. For instance, Yan et al.ref. ref34 reported that topical CBD administration enhanced wound closure and vascularization through the upregulation of vascular endothelial growth factor (VEGF) in granulation tissue. Additional studies have shown that CBD modulates inflammatory responses, promotes collagen synthesis, and stimulates neovascularization, thereby supporting its potential role in tissue regeneration.
Despite this promising evidence, the precise molecular and cellular mechanisms underlying CBD-mediated tissue remodeling remain incompletely understood. Considering the pivotal role of inflammation in regulating bone remodeling and the detrimental effects of excessive immune activation on bone integrity, the immunomodulatory functions of CBD are of particular relevance.ref. ref35 Notably, Li et al.ref. ref35 demonstrated that CBD reduced the mRNA expression of proinflammatory cytokines such as TNF-α and IL-6 in lipopolysaccharide-stimulated bone marrow mesenchymal stem cells, while concurrently enhancing the expression of osteogenic markers, including Runx2, alkaline phosphatase (ALP), and osteocalcin (OCN).
Given the multifactorial nature of wound healing and bone regeneration, elucidating the biological mechanisms through which natural compounds like CBD modulate inflammation and stimulate reparative processes is crucial for advancing translational regenerative strategies. The exploration of such bioactive molecules offers novel opportunities to overcome the limitations of conventional therapies and improve clinical outcomes. Accordingly, this review aims to provide a comprehensive analysis of the chemical and biological mechanisms underlying the effects of CBD in wound healing and bone regeneration, with particular emphasis on its immunomodulatory and osteogenic properties. Furthermore, it addresses key considerations regarding dosage, administration routes, formulation strategies, and regulatory aspects to optimize its therapeutic potential in regenerative medicine.
Natural versus Impaired Course of Wound Healing
Wound healing follows a tightly regulated physiological cascade involving overlapping cellular and molecular events that collectively restore tissue integrity and function.ref36,ref37 Under normal conditions, this process proceeds in a coordinated manner through the sequential phases of hemostasis, inflammation, proliferation, and remodeling, ultimately culminating in the re-establishment of tissue homeostasis. However, the persistence of inflammatory stimuli, microbial contamination, ischemia, or metabolic dysfunction can disrupt this orderly sequence, leading to delayed, chronic, or pathological healing outcomes. Such disturbances often result in excessive inflammation, impaired cell migration and proliferation, and aberrant extracellular matrix deposition, all of which compromise functional tissue restoration.
A clear understanding of the fundamental differences between physiological (natural) and dysregulated (non-natural or impaired) healing processes is essential for the rational design of therapeutic interventions targeting tissue regeneration. This distinction provides a critical conceptual framework for evaluating the pharmacological potential of bioactive compounds, such as cannabidiol (CBD), in modulating the wound-healing response. Prior to developing any therapeutic formulation aimed at enhancing repair in acute or chronic wounds, it is imperative to establish a comprehensive understanding of the molecular mechanisms that distinguish the natural from the impaired course of healing (Figure ).
Natural Course of Wound Healing: From Hemostasis to Remodeling
The physiological (natural) course of wound healing is a highly orchestrated process that progresses through four dynamic and overlapping phases: hemostasis, inflammation, proliferation, and remodelingref. ref38 (Figure ). Hemostasis, the immediate response to tissue injury, involves vascular constriction, platelet aggregation, and fibrin clot formation, establishing a provisional matrix and limiting blood loss. Inflammation follows, characterized by the sequential recruitment and activation of neutrophils and macrophages, which remove debris and pathogens while releasing cytokines and growth factors that orchestrate subsequent regenerative events. The proliferative phase entails fibroblast proliferation, extracellular matrix deposition, angiogenesis, and epithelial migration, collectively restoring tissue architecture. Finally, remodeling (maturation) involves the reorganization of collagen fibers, resolution of neovasculature, and restoration of tissue tensile strength, culminating in functional repair.
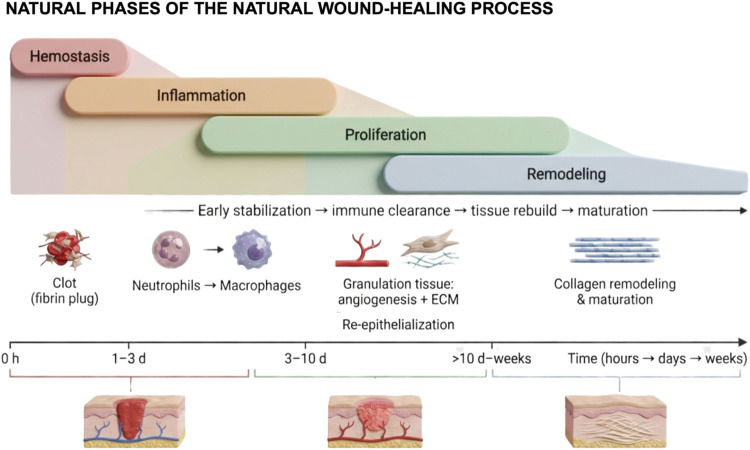
This well-regulated sequence provides the benchmark for assessing the therapeutic potential of bioactive compounds, such as CBD. By modulating inflammatory responses, promoting fibroblast and endothelial cell activity, and facilitating the transition from inflammation to proliferation and remodeling, CBD may enhance the efficiency and quality of tissue repair, particularly in chronic or nonhealing wounds. Understanding the natural course of healing is therefore essential for interpreting CBD’s regenerative mechanisms and optimizing its application in tissue engineering and regenerative medicine.
The initial phase of wound healing, hemostasis, functions as a protective mechanism that limits blood loss and establishes a provisional matrix for subsequent tissue repair.ref. ref39 Immediately following injury, vasoconstrictive mediators, including endothelin, released by damaged endothelium, and systemic factors such as epinephrine, stimulate contraction of vascular smooth muscle, transiently reducing blood flow at the injury site.ref. ref40 Concurrently, mediators such as bradykinin and fibrinopeptides initiate the coagulation cascade, leading to platelet aggregation and the formation of a primary hemostatic plug.ref. ref38
Coagulation proceeds via two principal pathways: the extrinsic pathway, triggered by tissue factor exposed at the site of vascular injury, and the intrinsic pathway, activated by contact between blood components and negatively charged surfaces, such as exposed collagen or matrix proteins.ref38,ref41 Both pathways converge on the activation of factor X, initiating the common pathway that leads to the conversion of prothrombin into thrombin.ref. ref42 Thrombin subsequently catalyzes the transformation of fibrinogen into fibrin, forming an insoluble network that stabilizes the platelet plug and establishes a secondary hemostatic structure, often referred to as the provisional matrix.ref. ref38
This fibrin-based matrix serves as a scaffold for cellular infiltration, initiating the inflammatory phase of wound healing. Neutrophils are among the first immune cells recruited to the wound site, where they perform essential functions including pathogen clearance, phagocytosis of debris, and secretion of proinflammatory mediators to orchestrate the subsequent reparative response.ref. ref43 The migration of inflammatory cells is guided by chemotactic signals generated by tissue-resident cells in response to damage-associated molecular patterns (DAMPs) and pathogen-associated molecular patterns (PAMPs), alerting neutrophils in the bone marrow to the site of injury and facilitating their directed movement into the wound.ref43,ref44
Upon arrival at the injury site, neutrophils release cytotoxic granules containing proteolytic enzymes and reactive oxygen species, which directly target and eliminate invading pathogens.ref. ref38 They also perform phagocytosis via surface antigen receptors, engulfing and degrading microbial and cellular debris.ref. ref36 Concurrently, macrophages infiltrate the wound, initially adopting a pro-inflammatory (M1) phenotype, characterized by the production of cytokines such as interleukin (IL)-6, tumor necrosis factor-α (TNF-α), and IL-1β, thereby amplifying the antimicrobial response.ref. ref45 Macrophages also contribute to resolution of inflammation by phagocytosing senescent neutrophils that fail to return to the bone marrow, effectively terminating the inflammatory phase.ref38,ref46 Following this, M1 macrophages undergo phenotypic switching to the anti-inflammatory, tissue-repair-promoting M2 phenotype, which supports angiogenesis, extracellular matrix (ECM) deposition, and immunoregulation.ref. ref45
The proliferative phase is initiated with the replacement of the fibrin clot by granulation tissue and the restoration of vascular supply via angiogenesis, ensuring delivery of oxygen and nutrients to the regenerating tissue.ref38,ref47 This phase involves coordinated activity of keratinocytes, fibroblasts, macrophages, and endothelial cells, aimed at reestablishing a functional tissue barrier.ref. ref48 Endothelial cells respond to hypoxia by upregulating vascular endothelial growth factor (VEGF), proliferating, and forming new capillary networks, thereby facilitating nutrient and oxygen delivery essential for effective tissue repair.ref38,ref49 Meanwhile, fibroblasts, stimulated by signals from platelets, endothelial cells, and M2 macrophages, deposit a provisional extracellular matrix, predominantly composed of type III collagen. A subset of fibroblasts differentiates into myofibroblasts, which generate contractile forces that reduce wound size and contribute to tissue integrity.ref. ref50
The remodeling (maturation) phase represents the final stage of wound healing, during which the granulation tissue is reorganized into mature, functional tissue. M2 macrophages transition to a phenotype (M2c) involved in ECM turnover, releasing matrix metalloproteinases (MMPs) that degrade excess collagen and remodel the provisional matrix.ref. ref45 During this phase, type III collagen is gradually replaced by type I collagen, restoring the tensile strength of the tissue.ref38,ref51 Additional structural components, such as elastin, are also reincorporated into the regenerating tissue, contributing to the functional and mechanical properties of the scar.ref. ref45 Together, these tightly regulated events ensure the successful resolution of the wound and the restoration of tissue homeostasis.
Non-Natural Course of Wound Healing: Persistence of Inflammatory Stimuli
In contrast to the tightly regulated sequence of cellular, humoral, and molecular events that characterize physiological wound healing, certain pathological conditions disrupt this process, leading to impaired regeneration. Such wounds may either form excessive scar tissue or develop into chronic lesions that exhibit minimal reduction in size (typically <40–50%) and fail to heal effectivelyref. ref52 (Figure ). In these scenarios, the anti-inflammatory and immunomodulatory properties of cannabidiol (CBD) may provide therapeutic benefit by attenuating persistent inflammation and promoting progression toward tissue repair.
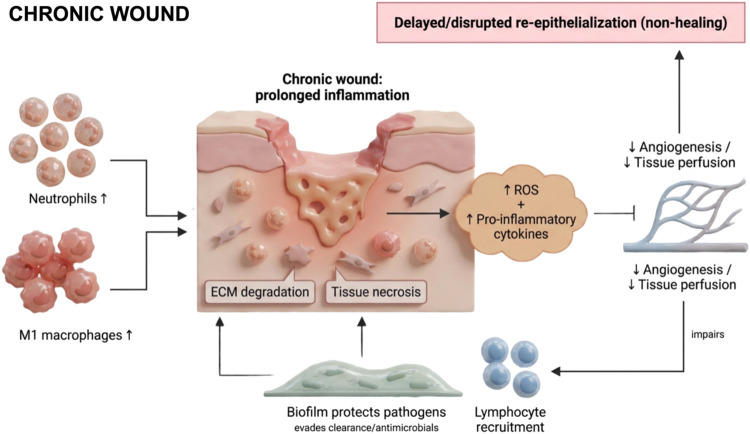
Chronic wound development is influenced by intrinsic factors, including age, malnutrition, diabetes, and immunosuppression, as well as extrinsic factors such as local temperature, humidity, and infection.ref53,ref54 Pathologically, chronic wounds are characterized by sustained inflammation, recurrent infections, necrosis, impaired re-epithelialization, reduced angiogenesis, and excessive production of reactive oxygen species (ROS).ref. ref55 Dysregulation may begin as early as the hemostatic phase; for example, hypercoagulable states can lead to thrombosis, which occludes blood vessels, restricts oxygen delivery, and induces tissue ischemia. Similarly, conditions such as hyperglycemia in diabetic individuals can elevate ROS levels, which, beyond their physiological role in vasoconstriction during hemostasis and vasodilation during proliferation, disrupt endothelial cell function, impair angiogenesis, and promote apoptosis.ref38,ref56
Persistent inflammation is the primary driver of the non-natural healing trajectory.ref. ref54 Chronic wounds are marked by prolonged infiltration of myeloid cellsincluding neutrophils, monocytes, and macrophagesinto the late inflammatory phase.ref. ref57 Within these wounds, there is an imbalance between pro-inflammatory (M1) and anti-inflammatory (M2) macrophages, with insufficient resolution of inflammation.ref. ref58 Impaired clearance of apoptotic neutrophils exacerbates the inflammatory milieu, sustaining high levels of cytokines such as TNF-α and IL-1β, which perpetuate tissue degradation.ref59,ref60 Additionally, macrophage-derived matrix metalloproteinases (e.g., MMP-2 and MMP-9) degrade extracellular matrix components, further delaying the proliferative phase.ref. ref59 Fibrocytes, a subset of macrophage-derived cells responsible for ECM deposition, may contribute to excessive fibrosis when dysregulated.ref. ref61
Crosstalk between immune cells and nonhematopoietic cells, such as keratinocytes, is also disrupted in chronic wounds. Aberrant expression of microRNAs, including miR-34a/c, miR-203, miR-19a/b, and miR-20a, alters keratinocyte-mediated immune regulation, delays re-epithelialization, and amplifies inflammation via upregulation of the NF-κB pathway, resulting in increased pro-inflammatory cytokine and chemokine production.ref35,ref62 Collectively, these failures in immune coordination prevent proper tissue repair, resulting in chronic wounds or pathological scarring.ref61,ref63
Moreover, microbial colonization plays a central role in perpetuating chronic inflammation and impairing tissue repair. A systematic review encompassing 185 chronic wounds reported that 78.2% contained polymicrobial biofilms, which provide structural and biochemical protection to pathogens against host immune defenses and antimicrobial therapies.ref63,ref64 In addition, coinfection with microorganisms such as Candida spp. and Porphyromonas gingivalis has been shown to inhibit cellular migration in vitro, underscoring the contribution of complex microbial communities to delayed healing, particularly in oral mucosal lesions following cancer therapy.ref. ref65
In contrast, the wound healing process differs substantially between cutaneous and oral mucosal tissues. Experimental models demonstrate that oral mucosal wounds exhibit reduced pro-fibrotic signaling and an expansion of fibroblast populations, which collectively promote earlier re-epithelialization, accelerated wound closure, and diminished scar formation compared with comparable skin lesions.ref66,ref67 Interleukin-1 (IL-1) signaling appears to play a tissue-specific role in the oral mucosa, being essential for efficient healing and protection of open wounds from bacterial invasion, while exerting comparatively limited effects on cutaneous wound closure under similar conditions.ref. ref68 Supporting the concept that oral tissues are primed for a heightened inflammatory responsiveness due to continuous microbial exposure, ex vivo gingival biopsies have been shown to secrete higher levels of pro-inflammatory cytokines, including IL-6, IL-8, IL-1β, IL-10, and TNF-α, than skin biopsies.ref. ref69 Furthermore, salivary secretions play a critical role in oral tissue repair, as evidenced by hyposalivation models demonstrating delayed palatal wound healing, thereby highlighting the importance of locally derived soluble factors in mucosal regeneration.ref. ref70 Collectively, these findings emphasize the distinct immunobiological environment of oral tissues and its implications for wound healing dynamics, particularly in the context of microbial burden and local regulatory factors.
Biological and Chemical Mechanisms of Cannabidiol in Wound Healing
CBD, a nonpsychoactive phytocannabinoid isolated from C. sativa L., has attracted considerable attention for its broad therapeutic potential, encompassing anti-inflammatory, analgesic, antioxidant, and pro-regenerative effects.ref. ref71 While early cannabinoid research predominantly addressed psychoactive properties, emerging evidence since the early 2000s highlights CBD’s capacity to modulate immune responses, cellular signaling pathways, and tissue repair mechanisms.ref. ref72 In the context of wound healing, CBD exerts effects across multiple overlapping phases, including the inflammatory, proliferative, and remodeling stages, by regulating immune cell function, oxidative stress, extracellular matrix deposition, and angiogenesis.
Cannabidiol and the Inflammatory Phase of Wound Healing
While inflammation is essential for initiating the natural wound-healing processref. ref73, its dysregulation can shift healing toward a nonphysiological trajectory, characterized by chronic inflammation, delayed repair, and fibrosis.ref. ref74 This underscores the therapeutic importance of modulating, rather than suppressing, the inflammatory response to promoting effective tissue regeneration.
Cannabidiol (CBD) offers a promising strategy for controlling excessive inflammation due to its multifaceted biological and chemical mechanisms. Unlike classical anti-inflammatory agents, which typically act via cyclooxygenase inhibition, CBD does not entirely suppress inflammation; instead, it modulates the response by reducing excessive cytokine release, limiting oxidative stress, and attenuating aberrant immune cell infiltration, thereby facilitating the resolution phase of healing.ref75,ref76
Mechanistically, cannabidiol (CBD) exerts its biological effects primarily through inhibition of the nuclear factor kappa B (NF-κB) signaling pathway, a key regulator of pro-inflammatory gene interleukin-1 receptor antagonist (IL-1Ra) and selected cytokinesref75,ref77 (Figure ). In vitro studies further elucidate these molecular mechanisms. For example, Sangiovanni et al.ref. ref76 demonstrated in HaCaT keratinocytes that CBD significantly reduced the release of vascular endothelial growth factor (VEGF), a central mediator of angiogenesis, and downregulated matrix metalloproteinase-9 (MMP-9), an enzyme critically involved in extracellular matrix degradation, in a dose-dependent manner. Additionally, CBD attenuated tumor necrosis factor-α (TNF-α) expression via NF-κB inhibition, highlighting its ability to modulate pro-inflammatory signaling without completely suppressing physiological inflammatory responses.ref. ref76 Beyond canonical pathway regulation, CBD also interacts with targets within the expanded endocannabinoid system, including GPR18. Emerging evidence indicates that CBD may function as a partial agonist of GPR18, promoting activation of the MAPK p44/42 signaling cascade, which is implicated in cellular processes essential for tissue repair and regeneration, such as proliferation, migration, and the regulation of apoptosis and autophagy.ref. ref78 Collectively, these findings underscore the pleiotropic signaling profile of CBD and its potential therapeutic relevance in modulating inflammation while supporting regenerative tissue outcomes. Collectively, these findings suggest that CBD acts as a regulatory modulator of inflammation, creating a favorable environment for the subsequent proliferative and remodeling phases of wound healing.
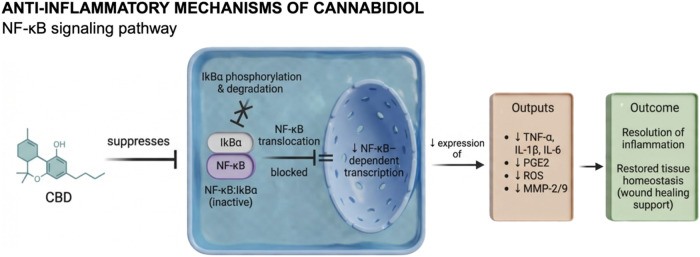
Kongkadee et al.ref. ref75 and Tran et al.ref. ref79 corroborated these findings by demonstrating that CBD downregulates pro-inflammatory mediators, particularly TNF-α and IL-1β, in macrophage-like and monocyte-derived cells. Both studies showed that CBD effectively inhibits phosphorylation of the p65 subunit of NF-κB, a critical step for its nuclear translocation and activation of inflammatory gene transcription. Specifically, Kongkadee et al.ref. ref75 identified CBD as the most potent bioactive compound within hemp extract, showing that treatment with 1 μg/mL maximally suppressed TNF-α production in RAW 264.7 macrophages. Moreover, CBD at concentrations ranging from 5 to 50 μg/mL significantly decreased IL-1β secretion, further confirming its dose-dependent immunomodulatory potential. Similarly, Tran et al.ref. ref79 reported that treatment of THP-1 cells with 6.6 μM CBD markedly reduced IL-6 and TNF-α production, concomitant with inhibition of IκB degradationan inhibitory protein that prevents NF-κB activation. In agreement with these findings, Kozela et al.ref. ref80 demonstrated that CBD impedes phosphorylation of the NF-κB p65 subunit, thereby preventing its nuclear translocation and subsequent transcription of pro-inflammatory cytokines (Figure ).
Evidence from in vivo studies further substantiates the anti-inflammatory properties of CBD. Klein et al.ref. ref81 investigated the effects of CBD on oral wound healing in Wistar rats and observed significantly lower inflammatory scores in CBD-treated animals compared to vehicle controls by the third day postinjury, indicating accelerated resolution of inflammation. Likewise, Genovese et al.ref. ref77 demonstrated that oral administration of CBD (10 mg/kg) reduced endometriosis-associated inflammation in Sprague–Dawley rats. This was characterized by increased IκB-α expression, decreased cytosolic cyclooxygenase-2 (COX-2) levels, and reduced nuclear localization of NF-κB within lesion tissues. Moreover, the treatment led to reduced levels of TNF-α, IL-1β, and prostaglandin E2 (PGE2) in the peritoneal environment, collectively confirming CBD’s ability to modulate both systemic and localized inflammatory responses.
Expanding on these findings, Zhou et al.ref. ref82 recently developed a multifunctional hydrogel incorporating CBD to promote the repair of radiation-induced and cutaneous wounds. The hydrogel markedly reduced inflammation, enhanced collagen deposition, and accelerated tissue regeneration. Mechanistically, CBD treatment was associated with modulation of key cytokines and chemokines, including IL-6, IL-17A, IL-22, CCL3, and CCL11, which are closely linked to the regulation of macrophage polarization and attenuation of chronic inflammatory signaling cascades. The authors proposed that these effects highlight CBD’s capacity to promote a shift toward a pro-resolving immune environment, thereby facilitating wound closure and tissue remodeling.
Taken together, the current body of evidence underscores CBD’s potential as a natural bioactive compound capable of attenuating inflammation and promoting wound repair through NF-κB pathway modulation and immune homeostasis restoration. Nonetheless, further studies are warranted to optimize therapeutic dosing, elucidate biodistribution dynamics, and assess long-term safety to ensure the translational viability of CBD-based interventions in regenerative medicine.
Cannabidiol as an Antioxidant: Mitigating Inflammation through Reduction of Oxidative Stress
Persistent inflammation is often accompanied by oxidative stress due to an imbalance between reactive oxygen species (ROS) and antioxidant defenses.ref. ref83 High levels of ROS impair angiogenesis, damage DNA, and disrupt cellular signaling pathways essential for healing.ref. ref84 CBD acts as a potent antioxidant by scavenging ROS and enhancing endogenous antioxidant systems such as glutathione peroxidase and superoxide dismutase (SOD).ref85,ref86
Antioxidant enzymes like SOD, glutathione peroxidase, and catalase help to protect the cells in the human body from harmful reactive oxygen substances.ref. ref87 In this context, previous studies have shown the beneficial effects of CBD on the reduction of the oxidation process.ref33,ref77,ref85,ref86 For instance, Genovese et al.ref. ref77 demonstrated that daily administration of CBD to Sprague–Dawley rats with endometriosis reduced oxidative stress. The thiobarbituric acid reactive substances (TBARS) assay revealed a decrease in lipid peroxidation, while levels of glutathione (GSH) and SOD activity were restored. Furthermore, Western blot analysis showed elevated expression of NADPH oxidase 1 (Nox-1) and Nox-4 in control rats, which was significantly reduced with CBD treatment.ref. ref77
Additionally, recent studies have focused on new biomaterials loaded with CBD, including a hydrogel made from alginate-zinc infused with CBD (CBD/Alg-Zn), which demonstrated effective antioxidant activity in reducing ROS in an in vivo model.ref. ref86 In another study, Chelminiak-Dudkiewicz et al.ref. ref88 examined chitosan-based orodispersible films enriched with cannabis oil as a source of CBD on L929 mouse fibroblasts, revealing noteworthy results in the context of antioxidant activity. The antioxidant activity of pure chitosan film was modest (9.5%), but the inclusion of CBD markedly enhanced radical-scavenging potentialup to 63.2%. DPPH assays revealed CBD concentration-dependent increases in antioxidant activity: 42.5% (1% CBD), 64.8% (5% CBD), and 72.7% (10% CBD). These findings highlight the strong antioxidant capacity of CBD-infused biomaterials for potential wound healing applications.
Cannabidiol and the Proliferative phase of Wound Healing
As previously discussed, the proliferative phase involves granulation tissue formation, and collagen depositon to create proteins in the extracellular matrix.ref. ref73 The presence of fibroblasts in the wound area is crucial for the formation of granulation tissue and the synthesis and deposition of collage.ref. ref89 A study shows that CBD may also support this phase by promoting fibroblast migration and matrix remodeling without impairing cellular proliferation.ref. ref90
Styrczewska et al.ref. ref90 developed a flax fiber dressing infused with CBD, which demonstrated promising outcomes in promoting cell proliferation and migration in vitro. Although CBD did not significantly influence fibroblast or keratinocyte morphology or proliferation, it enhanced matrix metalloproteinase activity and exerted notable anti-inflammatory effects. In wound healing assays, fibroblasts treated with CBD achieved full wound coverage within 48 h, with significantly fewer unhealed areas compared to controls.ref. ref81 Similarly, Kongkadee et al.ref. ref75 used an in vitro scratch assay to evaluate CBD’s effect on wound closure. At a concentration of 0.5 μg/mL, CBD significantly enhanced wound closure at 24-, 36-, and 48 h post-treatment, supporting the potential of CBD in wound healing.ref. ref75 Futhermore, granulation tissue formation is also critical for wound stabilization.
Effects of Cannabidiol on Granulation Tissue Formation
During the granulation phase of wound healing, fibroblasts play a pivotal role in synthesizing and depositing extracellular matrix (ECM) components, thereby providing structural integrity to the developing tissue. Concurrently, keratinocytes at the wound margins proliferate and migrate centripetally to re-establish the epithelial barrier.ref. ref73 Recent advances in biomaterial-based delivery systems have sought to potentiate these cellular events through the incorporation of bioactive molecules such as cannabidiol (CBD). In this context, Zheng et al.ref. ref86 developed a CBD-loaded alginate–zinc hydrogel (CBD/Alg–Zn) and evaluated its regenerative potential in a full-thickness excisional wound model in Sprague–Dawley rats. The CBD/Alg–Zn-treated wounds exhibited markedly enhanced granulation tissue formation, accelerated re-epithelialization, and reduced inflammatory cell infiltration compared with both the Alg–Zn-only and untreated control groups. These findings indicate that CBD, when combined with zinc and alginate, exerts synergistic effects likely attributable to its anti-inflammatory and antioxidant activities, as well as its potential to modulate fibroblast proliferation and ECM remodeling. Despite these promising outcomes, the precise molecular pathways by which CBD influences fibroblast activity, collagen deposition, and keratinocyte migration during granulation remain insufficiently characterized. Thus, further mechanistic studies, particularly those integrating molecular profiling and advanced biomaterial systems, are warranted to fully elucidate CBD’s role in this critical phase of wound repair.
Effect of Cannabidiol on Collagen Deposition
Collagen constitutes the primary structural protein of the extracellular matrix (ECM) in skin and connective tissues, providing mechanical strength, structural integrity, and a scaffold for cell adhesion and migration during tissue repair.ref91,ref92 The synthesis, organization, and remodeling of collagen fibers are tightly regulated processes that determine the quality and strength of the healed tissue. Dysregulation of collagen deposition may result in impaired healing or excessive scar formation, underscoring the importance of modulating this pathway for optimal wound repair.
In this context, cannabidiol (CBD), a nonpsychoactive phytocannabinoid derived from C. sativa, has emerged as a bioactive compound with potential regulatory effects on ECM remodeling. Zheng et al.ref. ref86 demonstrated that topical application of a CBD-loaded alginate-zinc hydrogel (CBD/Alg-Zn) significantly enhanced collagen deposition during the wound healing process in Sprague–Dawley rats. Histological analysis using Masson’s trichrome staining revealed a denser and more organized collagen network in both CBD/Alg-Zn- and Alg-Zn-treated groups at days 7 and 14 postinjury, with the CBD-enriched hydrogel exhibiting superior performance compared to the zinc-only formulation. These findings suggest that CBD may potentiate the reparative effects of zinc and alginate by synergistically promoting fibroblast activity and ECM synthesis.
Further supporting this notion, Qi et al.ref. ref93 investigated the osteogenic and extracellular matrix-modulatory effects of CBD in human dental pulp stem cells (hDPSCs). After 6 h of CBD exposure, there was a marked upregulation in the expression of collagen type I and II genes, indicating that CBD can stimulate the early stages of matrix protein synthesis in mesenchymal-derived cells. Type I collagen is the predominant fibrillar form found in dermal and bone tissues, while type II collagen is characteristic of cartilaginous structures, highlighting CBD’s broad influence across different tissue types.
Although the precise molecular mechanisms underlying CBD-induced collagen deposition remain incompletely understood, emerging evidence points toward the involvement of cannabinoid receptors (CB1 and CB2) and peroxisome proliferator-activated receptor γ (PPARγ) pathways, which can regulate fibroblast proliferation, differentiation, and ECM turnover. Additionally, CBD’s antioxidant and anti-inflammatory activities may create a microenvironment favorable to collagen maturation by reducing oxidative stress and pro-inflammatory cytokine release, both of which are known to impair matrix assembly.
Despite these encouraging findings, comprehensive mechanistic and translational studies are still required to confirm the direct targets of CBD in collagen biosynthesis, fiber organization, and cross-linking. Future research should integrate molecular, histological, and biomechanical analyses to elucidate how CBD influences ECM dynamics during various phases of wound healing and tissue regeneration.
Effects of Cannabidiol on Wound Contraction
Wound contraction constitutes a fundamental component of the proliferative phase of tissue repair, contributing to wound closure through centripetal movement of the wound edges. This process is predominantly mediated by the differentiation of fibroblasts into contractile myofibroblasts, a transition that typically occurs within 10–14 days postinjury.ref. ref89 Myofibroblasts express α-smooth muscle actin (α-SMA) within stress fibers, generating contractile forces that reorganize the collagenous matrix and promote mechanical closure of the wound bed.ref. ref47 The fibroblast-to-myofibroblast trans differentiation is orchestrated by a complex interplay of cytokines and growth factors, including transforming growth factor-β1 (TGF-β1), platelet-derived growth factor (PDGF), and connective tissue growth factor (CTGF), which collectively regulate cytoskeletal assembly, matrix remodeling, and integrin-mediated cell–matrix interactions.ref. ref94
Although the role of cannabidiol (CBD) in modulating wound contraction remains incompletely elucidated, growing evidence suggests it may influence fibroblast activity, ECM remodeling, and inflammatory resolutionfactors indirectly linked to contractile efficiency. Gangopadhyay et al.ref. ref95 demonstrated that a polyherbal Ayurvedic formulation containing C. sativa accelerated wound contraction in full-thickness excision wounds in Wistar rats, achieving a significantly smaller wound area within 6 days of treatment. While this study did not isolate CBD as the active component, the findings imply a potential cannabinoid-mediated enhancement of granulation tissue formation and matrix remodeling.
In contrast, Klein et al.ref. ref81 evaluated the effect of pure CBD in an oral mucosal injury model in Wistar rats. Although topical CBD application did not significantly reduce lesion size on days 3 and 7, histological analyses revealed markedly reduced inflammatory cell infiltration and lower inflammatory scores at day 3. These results indicate that CBD does not inhibit inflammation outright but rather modulates its resolution, likely through suppression of excessive pro-inflammatory cytokine release (e.g., TNF-α, IL-1β) and promotion of the transition to the proliferative phase. This immunomodulatory action may indirectly favor wound contraction by establishing a microenvironment conducive to fibroblast proliferation and differentiation, even if immediate contraction effects were not evident in oral tissue.
Further insights were provided by McIver et al.,ref. ref96 who investigated the topical application of a CBD-manuka honey formulation in equine limb wounds. Despite the known pro-healing and antimicrobial properties of manuka honey, the combination did not significantly alter wound contraction rates or overall healing time compared to controls. These findings highlight the complexity of translating rodent and in vitro data to large-animal and clinical models, where differences in skin architecture, wound tension, and pharmacokinetic profiles can markedly influence therapeutic outcomes.
At the molecular level, preliminary studies have suggested that CBD may modulate the TGF-β/Smad signaling axis, a central pathway in myofibroblast differentiation and matrix contraction. In addition, CBD’s antioxidant and endocannabinoid receptor-mediated actionsparticularly through CB2 and PPARγ activationcould attenuate excessive oxidative stress and inflammatory signaling, thereby optimizing the wound microenvironment for balanced matrix remodeling and contraction.ref97,ref98 However, direct evidence linking CBD to α-SMA expression or mechanical wound contraction remains scarce.
Altogether, these findings suggest that CBD’s influence on wound contraction is likely indirect, mediated through its effects on inflammatory resolution, fibroblast activation, and ECM dynamics, rather than through direct stimulation of contractile mechanisms. Future research should aim to clarify these interactions through well-controlled dose–response studies, standardized delivery systems, and mechanistic analyses involving molecular markers of myofibroblast differentiation (e.g., α-SMA, vimentin, and FAK phosphorylation). Understanding these pathways will be critical to harnessing CBD’s potential as a regulator of balanced tissue repair and fibrosis prevention.
Cannabidiol and the Remodeling Phase of Wound Healing
The remodeling or maturation phase represents the final stage of wound healing, typically commencing 2 to 3 weeks after injury and persisting for several months. This phase is characterized by the replacement of the provisional extracellular matrix (ECM) with a mature, functionally organized matrix, predominantly composed of type I collagen, and by the re-establishment of tissue architecture and mechanical strength.ref. ref91 Key cellular processes include re-epithelialization, collagen fiber cross-linking and alignment, regression of neovasculature, and apoptosis of myofibroblasts once sufficient wound tension is achieved. The interplay between fibroblasts, keratinocytes, endothelial cells, and immune cells determines the structural and functional outcome of the healed tissue.
Emerging evidence suggests that cannabidiol (CBD) exerts modulatory effects during the remodeling phase through its influence on epithelial differentiation, ECM reorganization, and vascular stabilization. In a study by Klein et al.,ref. ref81 topical administration of CBD on traumatic tongue ulcers in Wistar rats resulted in enhanced epithelial tissue structuring and organization during the late stages of healing. Histological evaluation revealed increased epithelial thickness, acanthosis, and hyperkeratinization, accompanied by more organized collagen fiber deposition and pronounced neovascularization. These histopathological features collectively indicate a stimulatory effect of CBD on tissue maturation and ECM remodeling.
Complementary findings from noncutaneous model further support CBD’s role in tissue remodeling. For instance, Zhang et al.ref. ref21 demonstrated that CBD treatment attenuated tissue swelling, fiber rupture, and inflammatory cell infiltration in a murine model of myocardial injury (C57BL/6 mice). These improvements were associated with reduced oxidative stress markers and enhanced organization of myocardial fibers, suggesting a broader role of CBD in postinjury tissue restoration and ECM homeostasis.
Mechanistically, CBD’s regulatory influence on remodeling is thought to involve several convergent pathways. Through activation of cannabinoid receptor type 2 (CB2) and peroxisome proliferator-activated receptor γ (PPARγ), CBD modulates fibroblast activity, promotes collagen maturation, and attenuates excessive matrix metalloproteinase (MMP) activity, thereby preventing aberrant degradation of newly synthesized collagen.ref. ref98 Additionally, CBD’s antioxidant capacity mitigates reactive oxygen species (ROS)-induced collagen cross-link disruption, while its anti-inflammatory effects contribute to the resolution of the wound milieu, allowing for balanced fibroblast-to-myofibroblast transition and appropriate ECM turnover.
CBD may also influence keratinocyte behavior during re-epithelialization. Studies have shown that cannabinoids can regulate keratinocyte proliferation and differentiation through TRPV1, CB1, and CB2 receptor-mediated pathways.ref99,ref100 By modulating intracellular calcium signaling and cytokine release, CBD may foster a microenvironment conducive to epithelial barrier restoration and stratified epidermal regenerationprocesses essential for complete tissue closure and long-term wound stability.
Despite these promising indications, current literature remains insufficient to establish a definitive understanding of CBD’s role in ECM remodeling and re-epithelialization. The available studies are limited by heterogeneous experimental designs, variations in CBD concentrations, and lack of standardization regarding formulation type (topical vs systemic) and administration timing. Furthermore, long-term effects on scar quality, tensile strength, and fibrosis risk have not been systematically evaluated.
Future investigations should integrate quantitative biomechanical assessments, molecular profiling of ECM components (e.g., type I/III collagen ratio, MMP/TIMP balance), and high-resolution imaging techniques to clarify CBD’s influence on the architectural and functional outcomes of tissue remodeling. Elucidating the optimal dosage and delivery method will be crucial for translating CBD’s reparative potential into therapeutic strategies for chronic wounds, fibrotic disorders, and implant-tissue interfaces.
Biological and Chemical Mechanisms of CBD on Bone Cells
Bone healing is a dynamic and tightly regulated process comprising three overlapping phases: inflammation, repair, and remodeling.ref. ref101 Intraoral bone remodeling is also influenced by the mechanical load from mastication.ref. ref102 Unlike extraoral skeletal sites where remodeling is predominantly regulated by systemic factors such as estrogen deficiency, parathyroid hormone levels, aging, and biomechanical stimuli, intraoral bone is persistently challenged by microbial stimuli.ref103−ref104ref105 Continuous exposure to microbial challenge decisively modulates the cellular and molecular mechanisms involved in tissue repair, characterizing bone healing in the intraoral region as a unique osteoimmune microenvironment.ref. ref106
During the remodeling phase, bone turnover depends on the rigorously coordinated coupling between osteoclast-mediated resorption and osteoblast-driven bone formation.ref. ref107 In intraoral sites, this coupling is frequently compromised by chronic low-grade inflammation triggered by dysbiotic biofilms enriched with key pathogens such as P. gingivalis, Treponema denticola, and Tannerella forsythia.ref108,ref109 These microorganisms activate host pattern recognition receptors, particularly Toll-like receptors (TLR2 and TLR4), in resident immune cells, osteoblasts, and periodontal ligament fibroblasts, leading to NF-κB activation and the transcription of pro-inflammatory cytokines such as TNF-α, IL-1β, and IL-6.ref. ref110
Another fundamental mechanism for intraoral bone resorption is the dysregulation of the RANK/RANKL/OPG axis.ref. ref109 In situations such as periodontal inflammation, there is an increase in the expression of receptor activator of nuclear factor kappa B (RANKL) ligand by activated T and B lymphocytes, osteoblastic lineage cells, and periodontal ligament cells, while simultaneously reducing the levels of its decoy receptor, osteoprotegerin (OPG).ref. ref109 This increased RANKL/OPG ratio intensifies the binding of RANKL to RANK in osteoclastic precursors, activating intracellular signaling pathways such as TRAF6, NF-κB, MAPKs, and NFATc.ref. ref111 This signaling cascade promotes the differentiation, polarization, and fusion of multinucleated osteoclasts, increasing the expression of tartrate-resistant acid phosphatase (TRAP), cathepsin K, and integrin αvβ3, culminating in extracellular matrix degradation and mineral dissolution. In addition, mechanisms initiated by virulence factors such as lipopolysaccharides (LPS) from Gram-negative anaerobes can directly or indirectly stimulate the differentiation of osteoclastic precursors through the activation of stromal and immune cells.ref. ref112 Furthermore, Th17 cells and IL-17 have been shown to amplify osteoclastogenic signaling, connecting adaptive immunity to alveolar bone destruction.ref. ref113 Collectively, evidence from osteoimmunology demonstrates that intraoral bone remodeling represents a paradigmatic model of inflammation-induced bone loss, in which host-microorganism interactions critically determine the balance between tissue regeneration and destruction.
Previous studies have demonstrated that CBD, the main nonpsychoactive component of cannabis, can enhance healing and recovery after bone injuries.ref93,ref114−ref115ref116 The current knowledge is rather limited and partly contradictory regarding the effect of exogenous cannabinoids on bone cells, and the more elaborate knowledge on the endocannabinoid system with respect to bone. Bone wellness depends on the coordinated action of osteoclast (OC) and osteoblast (OB) cells. The OC is a multinucleated bone cell derived from the hematopoietic lineage and formed through fusion of mononucleated precursors of the myeloid lineage. While OC cells can resorb bone matrix, OB are the bone-forming cells originating from the mesenchymal lineage. From an inflammatory perspective, OC and OB cells have been shown to express CB1 and CB2 receptors, both involved in the pathways of the endocannabinoid system.ref. ref117 CBD has been demonstrated to function as a molecule that acts like a noncompetitive antagonist for CB1 and CB2, primarily for CB,ref118,ref119 inhibiting the receptor’s activity without directly blocking the agonist binding site. However, inconsistent information has been reported regarding the effects of endocannabinoids on OCs. In a controlled in vitro study, Idris et al.ref. ref120 observed a stimulation of RANKL-induced OC formation by the addition of the substances: HU308, 2-AG or AEA, in the cell culture medium. Conversely, in 2016, Ofek et al.ref. ref121 observed an opposite effect, i.e., the inhibition of RANKL-induced OC formation by HU308, as a selective CB2 endocannabinoid agonist.ref. ref121 In fact, AEA reveals a conflicting role, since this endocannabinoid agonist may stimulate human OCs to form actin rings as well as to resorb bone in vitro, depending on its concentration. More recently, Nielsen et al.ref. ref122 raised an important question regarding the need to ensure that the dosage of CBD used to treat cultured cells is of pharmacological relevance. In this sense, the authors found that no or only weak inhibition of OC differentiation occurred in the lower doses tested (≤10 μM). Indeed, the number of nuclei per osteoclast cell significantly impacts its function, particularly in bone resorption. Previous studies have already indicated a strong correlation between number of nuclei and OC morphology, with bone formation/resorption, strongly suggesting that multinucleated osteoclast improves resorption efficiencyref. ref123 (Figure ).
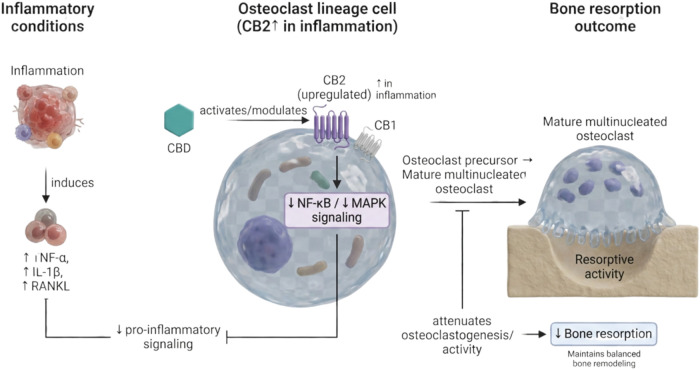
Importantly, recent mechanistic study supports that CBD can directly promote osteogenesis under inflammatory conditions in a receptor- and pathway-dependent manner. Li et al. demonstrated that CBD enhanced osteogenic differentiation of bone marrow mesenchymal stem cells (BMSCs) exposed to inflammatory stimulation, restoring osteogenic markers and mineralization while reducing inflammatory mediator expression; critically, these effects were linked to CB2-dependent activation of p38 MAPK signaling, reinforcing the concept that CBD may “re-couple” osteogenesis during inflammation rather than acting solely as an antiresorptive agent.ref. ref124 This is consistent with evidence that CBD can activate or modulate MAPK-related programs associated with osteogenic differentiation, including increased ALP activity, upregulation of osteoblast lineage markers, and enhanced mineral deposition in osteoblast-like cells.ref. ref115
Extending these observations to oral craniofacial contexts, CBD has also been shown to promote odonto/osteogenic responses in dental-derived progenitors. Qi et al. reported that CBD induced odonto/osteogenesis in human dental pulp cells (HDPCs), supporting the feasibility of CBD-mediated hard-tissue regenerative programs in oral settings.ref. ref93 In an inflammatory mimic, Yu et al. further demonstrated that CBD rescued TNF-α–inhibited proliferation, migration, and osteogenic/odontogenic differentiation of dental pulp stem cells (DPSCs), while concomitantly reducing TNF-α–induced expression of pro-inflammatory cytokines (TNF-α, IL-1β, IL-6) and upregulating pro-angiogenic VEGF expressionfeatures highly relevant to bone regeneration and vascularized repair in infected or inflamed microenvironments.ref. ref125 Collectively, these studies strengthen the biological plausibility of CBD as a dual-action modulator in oral bone regeneration, simultaneously counteracting inflammatory suppression of osteogenesis and supporting pro-reparative cellular behaviors (migration, angiogenic signaling) in MSC-like populations.
On the other hand, CBD also influences bone biology through nonclassical targets. Previously, CBD has been reported to modulate the orphan G protein-coupled receptor GPR55, which plays a functional role in bone remodeling.ref. ref126 Whyte et al.ref. ref127 showed that activation of GPR55 by its agonists increased OC polarization and intensified resorptive activity, whereas these effects were markedly reduced in OC derived from GPR55–/– mice and after CBD treatment, consistent with CBD acting as a functional antagonist of GPR55. In addition, CBD has been reported to promote OB migration and mesenchymal stem cell differentiation in vitro via antagonism of GPR55.ref. ref128 More recent evidence reinforces the relevance of the LPI–GPR55 axis as a regulator of osteoclast differentiation and function and provides additional mechanistic support for CBD-mediated antagonism in this pathway. Mosca et al. (2021) demonstrated that GPR55 signaling modulates osteoclast activity in RANKL-driven differentiation systems, where lysophosphatidylinositol (LPI) enhanced osteoclast activity and GPR55 antagonism reduced resorptive function.ref. ref129
Indeed, the stimulatory and inhibitory effects of cannabinoids on bone cells is a complex matter. Although cannabis is portrayed as a dangerous drug, also due to its ability to reduce bone mineral density, CBD, as a nonpsychoactive component of C. sativa, also presents therapeutic properties in bone repair/remodeling that are not yet fully understood. In 2020, Kang et al.ref. ref115 investigated the impact of CBD on osteoblastic differentiation and found that the CBD treatment upregulated the expression of angiopoietin-1, enhanced alkaline phosphatase (ALP) activity, and stimulated cell migration and calcium deposition. The results also demonstrated a time-dependent increase in the expression of osteoblast-related proteins that induce bone, including distal-less homeobox 5 (DLX5), bone sialoprotein (BSP), osteocalcin (OCN), type I collagen, Runx2, osterix (OSX), and ALP matrix-related mineralization.ref. ref115
Mechanistically, Kang et al. (2020) further linked these osteogenic effects to strengthened interactions among RUNX2/OSX and phosphorylated p38 MAPK, suggesting that CBD can facilitate transcriptional control of osteoblast lineage commitment via MAPK-driven osteogenic signaling.ref. ref115 Together with the CB2–p38-dependent osteogenic rescue observed in inflammatory BMSC models,ref. ref130 these findings support a convergent theme in which CBD may promote osteoblastogenesis through p38 MAPK–associated pathways, with receptor dependence likely varying by cell type and inflammatory context.
During the repair phase, vascular invasion and collagen matrix deposition lead to callus formation. Fibroblasts contribute by forming a stroma conducive to vascular growth.ref. ref102 In this sense, Kogan et al.ref. ref116 examined the influence of CBD on fracture healing in a rat model and demonstrated that CBD enhanced the biomechanical strength of the fracture callus without affecting its volume or mineral content. The increased mechanical integrity was attributed to upregulation of PLOD1, an enzyme responsible for collagen cross-linking, in osteoblast cultures treated with CBD. These findings highlight the specific role of CBD in improving fracture healing in long bones, increasing the biomechanical quality of the newly formed bone.ref. ref116
This observation is particularly relevant for craniofacial and peri-implant regeneration, where collagen cross-linking and matrix maturation influence the mechanical quality of newly formed bone and the stability of the implant–bone interface. Although long-bone fracture models cannot be directly extrapolated to intraoral bone, the mechanistic emphasis on collagen cross-linking enzymes suggests that CBD may affect not only osteogenic differentiation but also the material properties of repair tissue, an aspect often overlooked in oral regenerative discussions.
The final stage of fracture healing is the remodeling phase, during which bone regains its original architecture, function, and mechanical strength. This process, which can span several months to years, is modulated by mechanical stimuli.ref. ref131 Under axial loading, bone is formed in regions experiencing stress and resorbed in areas where it is not required.ref. ref123 A recent study by Kamali et al.ref. ref114 reported that CBD accelerated healing and improved biomechanical properties in a rat model of critical-sized bone defect through the mesenchymal stem cell migration and osteogenic differentiation, leading to a more effective bone bridge formation at the defect site.ref. ref114 Although literature regarding the effect of CBD on this process remains scarce, the found information reinforces the idea that CBD could be a promising therapeutic option for promoting bone healing and remodeling.
Notably, emerging oral-focused literature highlights an additional translational layer: beyond host cell modulation, CBD may also influence the microbial drivers of osteoimmune dysfunction. For instance, CBD demonstrated antimicrobial activity against multispecies subgingival biofilms in vitro, supporting the possibility that CBD-based local delivery platforms might provide dual benefits: reducing microbial burden while modulating host inflammation and bone remodeling pathways.ref. ref132 This is particularly aligned with periodontal and peri-implant pathogenesis, where persistent biofilm challenge sustains NF-κB signaling, elevates RANKL, and prolongs osteoclastogenic cues.
Mind the Gap: From Laboratorial Studies to Clinical Application
Regarding the commercial availability of cannabinoid-based medicines, only a few have been rigorously tested to assess their safety and efficacy. They have therefore been approved for use at the national level by regulatory agencies. Epidiolex is an oral solution of 98% pure cannabidiol (CBD). The medicine is approved for the treatment of seizures in pediatric patients with Lennox-Gastaut syndrome or Dravet syndrome.ref. ref14 The biological effects of cannabinoids, the major constituents of the ancient medicinal plant C. sativa (marijuana) are mediated by two members of the G-protein coupled receptor family, cannabinoid receptors 1 (CB1R) and 2. The CB1R is the prominent subtype in the central nervous system (CNS) and has drawn great attention as a potential therapeutic avenue in several pathological conditions, including neuropsychological disorders and neurodegenerative diseases. Furthermore, cannabinoids also modulate signal transduction pathways and exert profound effects at peripheral sites. Despite the therapeutic potential of cannabinoids, their clinical application has been significantly hindered by their psychoactive effects. In this review, we briefly summarized our knowledge of cannabinoids and the endocannabinoid system, focusing on the CB1R and the CNS, with emphasis on recent breakthroughs in the field. We aim to define several potential roles of cannabinoid receptors in the modulation of signaling pathways and in association with several pathophysiological conditions. We believe that the therapeutic significance of cannabinoids is masked by the adverse effects and here alternative strategies are discussed to take therapeutic advantage of cannabinoids.ref. ref14 Sativex is also a mouth spray formulated from the extract of the C. sativa L. plant, containing mainly Δ9-THC and CBD in almost equal proportions. It is indicated for the treatment of spasticity.ref. ref133 However, although there are CBD-based medicines, none of them are indicated for promoting tissue and bone regeneration.ref. ref134
Now, CBD is being studied in several preclinical studies, showing surprising results. However, CBD has limitations due to its highly lipophilic nature with a log P of 6.3, which represents the logarithm of the partition coefficient of a drug between n-octanol and water and, it has low water solubility, measuring at 12.6 mg/L. These characteristics allow CBD as a Class II substance in the Biopharmaceutical Classification System (BCS), characterizing it as a substance with low water solubility.ref. ref135 CBD is extensively metabolized in the liver, mainly through hydroxylation, forming 7-OH–CBD. This compound undergoes further metabolization, generating various metabolites, which are eliminated from the body via feces and urine.ref. ref136
Bioavailability refers to the proportion of a drug that reaches systemic circulation unchanged. Intravenous administration provides 100% bioavailability, while oral, inhaled, or transdermal routes reduce it due to incomplete absorption. CBD’s bioavailability clearly varies depending on the method of administration.ref. ref137 When clinically applied, the bioavailability of aerosolized CBD has been reported to be capable of generating rapid peaks in plasma concentration between five and 10 min, with significantly higher bioavailability (around 31%) compared to oral administration.ref. ref138 Oral bioavailability was estimated at 6%, mainly due to extensive first-pass metabolism, where drug molecules are metabolized before entering systemic circulation, which can reduce their bioavailability.ref. ref139 Thus, administering cannabinoids via inhalation or oral mucosal routes provides an alternative method that by passes or minimizes extensive first-pass metabolism, as seen with oral cannabinoid administration.ref. ref140
The effective dose of CBD shows great variability between individuals, influenced by factors such as metabolism, body weight, age, gender, and clinical conditions, which represents a challenge for the development of universal release systems.ref. ref141 One study showed that in humans, after ingesting an oral capsule containing 5.4 mg of CBD, the average maximum plasma concentration (Cmax) was 0.93 ng/mL, which was higher in women than in men.ref. ref116 Furthermore, in one experiment, coadministration of lipids with oral CBD increased systemic availability by almost three times in rats.ref. ref142 These findings reinforce the need for personalized strategies to optimize its administration in different patient profiles.
In the past decade, there has been a growing number of studies focused on enhancing the solubility of class II drugs. Schedule II drugs, according to the Biopharmaceutical Classification, substances or chemicals are defined as drugs with a high potential for abuse, with use potentially leading to severe psychological or physical dependence.ref. ref143 The bioavailability of these drugs is directly influenced by their dissolution rate, which is closely related to solubility, consequently, an improved solubility leads to enhanced bioavailability. Various strategies can be employed to optimize the dissolution rate, such as nanonization, which involves reducing the particle size of the active pharmaceutical ingredient (API) to a nanometric scale.ref. ref143 According to the Noyes-Whitney equation, which indicates the dissolution rate of a solid in a solvent, reducing the particle size of a drug increases its surface area, resulting in a proportional increase in the dissolution rate. Consequently, this leads to better absorption of drugs with low solubility.ref. ref144 In this context, the use of nanotechnology as a drug delivery system has gained prominence due to its unique properties, which include advantages in controlling the physicochemical behavior of the drug, such as solubility and release, as well as directing the drug to the target site, reducing adverse effects.ref. ref145 Nanosystems play a vital role in safeguarding drugs from damage in the gastrointestinal region, thereby facilitating the efficient administration of class II drugs to their intended targets.ref. ref146 One notable characteristic of these nanodrug delivery systems is their remarkable versatility in terms of application routes. These routes include parenteral, oral, nasal, pulmonary, ocular, and transdermal routes.ref. ref146 Among various nanomaterials, liposomes, vesicles, micelles, nanoparticles, nanosuspensions, microemulsions, and nanoemulsions are notableref. ref147 to enhance the solubility of CBD.
Tran et al. developed a CBD-based nanoemulsion to evaluate its regenerative potential and bioavailability using an in vitro human corneal substitute model. The formulation demonstrated superior physicochemical stability, retaining 93.57% of cannabidiol (CBD) content, whereas pure CBD exhibited only 53.58% retention after 4 h of exposure.ref. ref79 This difference was attributed to degradation and metabolic susceptibility of the free compound, while the nanoemulsion matrix provided a protective environment that enhanced CBD chemical stability during topical application.ref. ref79
In the context of dental applications, recent investigations have explored CBD-loaded biomaterials as local delivery platforms, particularly in scenarios requiring prolonged retention and controlled release within the oral cavity. For example, a chitosan-based mucoadhesive hydrogel incorporating CBD-loaded poly(lactic-co-glycolic acid) (PLGA) nanospheres was designed to improve mucosal permeability and local drug availability. In vitro analyses demonstrated a pro-reparative profile characterized by downregulation of inflammatory cytokines, suggesting potential therapeutic applicability in oral inflammatory conditions such as gingivitis and periodontitis.ref. ref148 Furthermore, CBD has been incorporated into multifunctional regenerative hydrogels targeting bone repair. An alginate-based copper-CBD hydrogel exhibiting antibacterial properties was shown to enhance both osteogenic differentiation and angiogenic responses in vitro.ref. ref149 Collectively, these biomaterial systems highlight the potential of nanotechnology-enabled approaches to optimize local CBD delivery to oral tissues; however, robust clinical validation remains lacking.
To date, relatively few studies have investigated nanotechnology-based strategies to enhance the biological performance of CBD, and the available literature is limited by the scarcity of comprehensive preclinical and clinical investigations. Therefore, further well-designed in vitro and in vivo studies are warranted to elucidate the therapeutic benefits of CBD and to clarify its underlying mechanisms of action in wound healing and bone regeneration, thereby facilitating translation into clinical practice.
Legal Status of Cannabidiol
In November 2017, the World Health Organization (WHO)ref. ref150 Expert Committee on Drug Dependence (ECDD) concluded that pure CBD does not appear to pose a risk of abuse or cause harm. The current evidence does not justify reclassifying this substance. However, the decision on its legal status is the responsibility of the legislators in each country. Some countries, such as Australia, Canada, Switzerland, the United Kingdom, and the United States, have loosened regulations on CBD, classifying CBD-containing products as medicines.
Furthermore, in 2018, the Food and Drug Administration (FDA) approved the use of Epidiolex, a CBD-based drug, for the treatment of seizures associated with tuberous sclerosis complex in patients aged one year and older after concluding that the drug is safe and effective for this indication. Following its approval, Epidiolex was classified as a Schedule V substance, the least restrictive category, reserved for medicines with a low potential for abuse. This removed it from the Schedule I classification, which includes substances with a high potential for abuse and no accepted medicinal value under the Federal Controlled Substances Act of 1970.
In 2019, the European Medicines Agency (EMA) approved the first cannabinoid-derived medicine. This drug, composed of isolated CBD, has been authorized for the treatment of children with intractable epilepsy and has been designated an orphan drug because there are no similar therapeutic alternatives available.
Also, in 2019, the National Health Surveillance Agency (Anvisa)ref. ref151 in Brazil regulated the manufacture and sale of CBD-based products in pharmacies on prescription to treat serious conditions such as refractory epilepsy, autism, and degenerative neurological diseases. The agency has imposed limits on the THC content: products with up to 0.2% can be prescribed for a wide range of patients, while those with a higher content are restricted to patients in palliative care or with no other therapeutic options.
On the other hand, in most Latin American countries, the medicinal use of cannabidiol (CBD) is authorized, especially for the treatment of specific diseases such as epilepsy and other neurodegenerative conditions. However, regulations regarding the production, sale, and import of CBD-based products increase considerably between countries. In addition, the recreational use of cannabis remains illegal in almost the entire region, of one except Uruguay. The trend is toward a gradual relaxation of the laws, accompanied by the implementation of standards that ensure the safe and regulated use of the substance.ref. ref152
Despite the advances in legislation, there is still a need to carry out more research, establish clear and accessible regulations, and maintain continuous monitoring to guarantee the efficacy and safety of CBD use as a therapeutic medicine.
Conclusion
In conclusion, this critical review examined the biological mechanisms through which cannabidiol (CBD) may contribute to wound healing and bone repair/regeneration. Current preclinical evidence supports the therapeutic potential of CBD in enhancing both soft- and hard-tissue repair by modulating key molecular pathways involved in inflammation, cellular proliferation, angiogenesis, and extracellular matrix remodeling. Despite these promising findings, important translational challenges remain. In particular, the minimum effective dose required to achieve therapeutic benefits while minimizing adverse effects has not yet been clearly established, underscoring the need for rigorous dose–response and safety studies to enable clinical application in humans.
With respect to bone repair and regeneration, several mechanistic aspects require further clarification, including the interaction of CBD with osteoblast and osteoclast receptor systems, its effects on osteoclast multinucleation and activity, and the resulting impact on the balance between bone formation and resorption. The studies discussed in this review collectively highlight substantial knowledge gaps regarding both the local and systemic effects of CBD, as well as its integration within complex intracellular signaling networks. Therefore, additional well-designed mechanistic investigations, followed by robust preclinical and clinical studies, are necessary before routine clinical implementation can be considered. Concurrently, comprehensive evaluation of optimal dosage regimens, pharmacokinetics, bioavailability, and bio efficacy of CBD-based drugs and biomaterials will be essential to support their safe and effective translation into oral and craniofacial regenerative therapies.
References
- T. J. Shaw, P. Martin. Wound Repair at a Glance. J. Cell Sci., 2009. [DOI | PubMed]
- S. A. Eming, B. Brachvogel, T. Odorisio, M. Koch. Regulation of Angiogenesis: Wound Healing as a Model. Prog. Histochem. Cytochem., 2007. [DOI | PubMed]
- A. Gosain, L. A. DiPietro. Aging and Wound Healing. World J. Surg., 2004. [DOI | PubMed]
- M. A. Terkawi, T. Ebata, S. Yokota, D. Takahashi, T. Endo, G. Matsumae, T. Shimizu, K. Kadoya, N. Iwasaki. Low-Grade Inflammation in the Pathogenesis of Osteoarthritis: Cellular and Molecular Mechanisms and Strategies for Future Therapeutic Intervention. Biomedicines, 2022. [DOI | PubMed]
- A. Nauth, E. Schemitsch, B. Norris, Z. Nollin, J. T. Watson. Critical-Size Bone Defects: Is There a Consensus for Diagnosis and Treatment?. J. Orthop. Trauma, 2018. [DOI | PubMed]
- E. Gómez-Barrena, P. Rosset, D. Lozano, J. Stanovici, C. Ermthaller, F. Gerbhard. Bone Fracture Healing: Cell Therapy in Delayed Unions and Nonunions. Bone, 2015. [DOI | PubMed]
- J. Henkel, M. A. Woodruff, D. R. Epari, R. Steck, V. Glatt, I. C. Dickinson, P. F. M. Choong, M. A. Schuetz, D. W. Hutmacher. Bone Regeneration Based on Tissue Engineering Conceptions – A 21st Century Perspective. Bone Res., 2013. [DOI | PubMed]
- R. Dimitriou, E. Jones, D. McGonagle, P. V. Giannoudis. Bone Regeneration: Current Concepts and Future Directions. BMC Med., 2011. [DOI | PubMed]
- V. Ap, H. K, S. A, S. Ad, B. Ab. Therapeutic Strategies for Enhancing Angiogenesis in Wound Healing. Adv. Drug Delivery Rev., 2019. [DOI]
- I. B. do Valle, P. H. D. M. Prazeres, R. A. Mesquita, T. A. Silva, H. M. de Castro Oliveira, P. R. Castro, I. D. P. Freitas, S. R. Oliveira, N. A. Gomes, R. F. de Oliveira, L. F. Marquiore, S. Macari, F. A. do Amaral, H. Jácome-Santos, L. S. Barcelos, G. B. Menezes, M. M. Marques, A. Birbrair, I. M. A. Diniz. Photobiomodulation Drives Pericyte Mobilization towards Skin Regeneration. Sci. Rep., 2020. [DOI | PubMed]
- S. Geahchan, P. Baharlouei, A. Rahman. Marine Collagen: A Promising Biomaterial for Wound Healing, Skin Anti-Aging, and Bone Regeneration. Mar. Drugs, 2022. [DOI | PubMed]
- S. Geahchan, P. Baharlouei, A. Rahman. Marine Collagen: A Promising Biomaterial for Wound Healing, Skin Anti-aging, and Bone Regeneration. Mar. Drugs, 2022. [DOI | PubMed]
- E. B. Russo. History of Cannabis and Its Preparations in Saga, Science, and Sobriquet. Chem. Biodivers., 2007. [DOI | PubMed]
- S. Zou, U. Kumar. Cannabinoid Receptors and the Endocannabinoid System: Signaling and Function in the Central Nervous System. Int. J. Mol. Sci., 2018. [DOI | PubMed]
- S. Pisanti, M. Bifulco. Medical Cannabis: A Plurimillennial History of an Evergreen. J. Cell. Physiol., 2019. [DOI | PubMed]
- M. A. ElSohly, Z. Mehmedic, S. Foster, C. Gon, S. Chandra, J. C. Church. Changes in Cannabis Potency Over the Last 2 Decades (1995–2014): Analysis of Current Data in the United States. Biol. Psychiatry, 2016. [DOI | PubMed]
- P. Pacher, S. Bátkai, G. Kunos. The Endocannabinoid System as an Emerging Target of Pharmacotherapy. Pharmacol. Rev., 2006. [DOI | PubMed]
- P. H. Reggio. Endocannabinoid Binding to the Cannabinoid Receptors: What Is Known and What Remains Unknown. Curr. Med. Chem., 2010. [DOI | PubMed]
- D. McHugh. GPR18 in Microglia: Implications for the CNS and Endocannabinoid System Signalling. Br. J. Pharmacol., 2012. [DOI | PubMed]
- M. Omi, Y. Mishina. Roles of Osteoclasts in Alveolar Bone Remodeling. Genesis, 2022. [DOI | PubMed]
- J. Zhang, Z. Luo, Z. Zhang, M. Zhao, C. Tong, P. Cong, S. Mao, Y. Zhao, M. Hou, Y. Piao, Y. Liu. Protective Effect and Mechanism of Cannabidiol on Myocardial Injury in Exhaustive Exercise Training Mice. Chem. Biol. Interact., 2022. [DOI | PubMed]
- E. Honkisz-Orzechowska, D. Łażewska, G. Baran, K. Kieć-Kononowicz. Uncovering the Power of GPR18 Signalling: How RvD2 and Other Ligands Could Have the Potential to Modulate and Resolve Inflammation in Various Health Disorders. Molecules, 2024. [DOI | PubMed]
- Structural insight into GPR55 ligand recognition and G-protein coupling | Cell Research.
- H. Yang, J. Zhou, C. Lehmann. GPR55 – a Putative “Type 3” Cannabinoid Receptor in Inflammation. J. Basic Clin. Physiol. Pharmacol., 2016. [DOI | PubMed]
- V. Chiurchiù, M. Lanuti, M. De Bardi, L. Battistini, M. Maccarrone. The Differential Characterization of GPR55 Receptor in Human Peripheral Blood Reveals a Distinctive Expression in Monocytes and NK Cells and a Proinflammatory Role in These Innate Cells. Int. Immunol., 2015. [DOI | PubMed]
- K. S. Ladha, P. Ajrawat, Y. Yang, H. Clarke. Understanding the Medical Chemistry of the Cannabis Plant Is Critical to Guiding Real World Clinical Evidence. Molecules, 2020. [DOI | PubMed]
- Y. Gaoni, R. Mechoulam. Isolation, Structure, and Partial Synthesis of an Active Constituent of Hashish. J. Am. Chem. Soc., 1964. [DOI]
- L. M. Borgelt, K. L. Franson, A. M. Nussbaum, G. S. Wang. The Pharmacologic and Clinical Effects of Medical Cannabis. Pharmacotherapy, 2013. [DOI | PubMed]
- Constituents of Cannabis Sativa . In. Advances in Experimental Medicine and Biology;, 2021. [DOI | PubMed]
- K. Iffland, F. Grotenhermen. An Update on Safety and Side Effects of Cannabidiol: A Review of Clinical Data and Relevant Animal Studies. Cannabis Cannabinoid Res., 2017. [DOI | PubMed]
- C. Pagano, G. Navarra, L. Coppola, G. Avilia, M. Bifulco, C. Laezza. Cannabinoids: Therapeutic Use in Clinical Practice. Int. J. Mol. Sci., 2022. [DOI | PubMed]
- D. Simões, S. P. Miguel, M. P. Ribeiro, P. Coutinho, A. G. Mendonça, I. J. Correia. Recent Advances on Antimicrobial Wound Dressing: A Review. Eur. J. Pharm. Biopharm., 2018. [DOI | PubMed]
- J. M. Baron, M. Glatz, E. Proksch. Optimal Support of Wound Healing: New Insights. Dermatology, 2020. [DOI | PubMed]
- X. Yan, B. Chen, Y. Lin, Y. Li, Z. Xiao, X. Hou, Q. Tan, J. Dai. Acceleration of Diabetic Wound Healing by Collagen-Binding Vascular Endothelial Growth Factor in Diabetic Rat Model. Diabetes Res. Clin. Pract., 2010. [DOI | PubMed]
- D. Li, H. Peng, L. Qu, P. Sommar, A. Wang, T. Chu, X. Li, X. Bi, Q. Liu, I. Gallais Sérézal, O. Rollman, W. Lohcharoenkal, X. Zheng, S. Eliasson Angelstig, J. Grünler, A. Pivarcsi, E. Sonkoly, S.-B. Catrina, C. Xiao, M. Ståhle, Q.-S. Mi, L. Zhou, N. Xu Landén. miR-19a/b and miR-20a Promote Wound Healing by Regulating the Inflammatory Response of Keratinocytes. J. Invest. Dermatol., 2021. [DOI | PubMed]
- M. Takeo, W. Lee, M. Ito. Wound Healing and Skin Regeneration. Cold Spring Harb. Perspect. Med., 2015. [DOI | PubMed]
- R. Richardson, K. Slanchev, C. Kraus, P. Knyphausen, S. Eming, M. Hammerschmidt. Adult Zebrafish as a Model System for Cutaneous Wound-Healing Research. J. Invest. Dermatol., 2013. [DOI | PubMed]
- M. Rodrigues, N. Kosaric, C. A. Bonham, G. C. Gurtner. Wound Healing: A Cellular Perspective. Physiol. Rev., 2019. [DOI | PubMed]
- S. A. Smith, R. J. Travers, J. H. Morrissey. How It All Starts: Initiation of the Clotting Cascade. Crit. Rev. Biochem. Mol. Biol., 2015. [DOI | PubMed]
- S. Godo, H. Shimokawa. Endothelial Functions. Arterioscler. Thromb., Vasc. Biol., 2017. [DOI | PubMed]
- K. Neubauer, B. Zieger. Endothelial Cells and Coagulation. Cell Tissue Res., 2022. [DOI | PubMed]
- R. G. Macfarlane. An Enzyme Cascade in the Blood Clotting Mechanism, and Its Function as a Biochemical Amplifier. Nature, 1964. [DOI | PubMed]
- S. Guo, L. A. Dipietro. Factors Affecting Wound Healing. J. Dent. Res., 2010. [DOI | PubMed]
- M. Lech, H.-J. Anders. Macrophages and Fibrosis: How Resident and Infiltrating Mononuclear Phagocytes Orchestrate All Phases of Tissue Injury and Repair. Biochim. Biophys. Acta, Mol. Basis Dis., 2013. [DOI]
- H. N. Wilkinson, M. J. Hardman. Wound Healing: Cellular Mechanisms and Pathological Outcomes. Open Biol., 2020. [DOI | PubMed]
- D. M. Mosser, J. P. Edwards. Exploring the Full Spectrum of Macrophage Activation. Nat. Rev. Immunol., 2008. [DOI | PubMed]
- I. A. Darby, B. Laverdet, F. Bonté, A. Desmoulière. Fibroblasts and Myofibroblasts in Wound Healing. Clin. Cosmet. Investig. Dermatol., 2014. [DOI]
- J. Li, J. Chen, R. Kirsner. Pathophysiology of Acute Wound Healing. Clin. Dermatol., 2007. [DOI | PubMed]
- G. C. Gurtner, S. Werner, Y. Barrandon, M. T. Longaker. Wound Repair and Regeneration. Nature, 2008. [DOI | PubMed]
- H. E. Talbott, S. Mascharak, M. Griffin, D. C. Wan, M. T. Longaker. Wound Healing, Fibroblast Heterogeneity, and Fibrosis. Cell Stem Cell, 2022. [DOI | PubMed]
- M. B. Witte, A. Barbul. General Principles of Wound Healing. Surg. Clin. North Am., 1997. [DOI | PubMed]
- X. Zhu, M. M. Olsson, R. Bajpai, K. Järbrink, W. E. Tang, J. Car. Health-Related Quality of Life and Chronic Wound Characteristics among Patients with Chronic Wounds Treated in Primary Care: A Cross-Sectional Study in Singapore. Int. Wound J., 2022. [DOI | PubMed]
- G. Han, R. Ceilley. Chronic Wound Healing: A Review of Current Management and Treatments. Adv. Ther., 2017. [DOI | PubMed]
- R. Zhao, H. Liang, E. Clarke, C. Jackson, M. Xue. Inflammation in Chronic Wounds. Int. J. Mol. Sci., 2016. [DOI | PubMed]
- J. Larouche, S. Sheoran, K. Maruyama, M. M. Martino. Immune Regulation of Skin Wound Healing: Mechanisms and Novel Therapeutic Targets. Adv. Wound Care, 2018. [DOI]
- C. Dunnill, T. Patton, J. Brennan, J. Barrett, M. Dryden, J. Cooke, D. Leaper, N. T. Georgopoulos. Reactive Oxygen Species (ROS) and Wound Healing: The Functional Role of ROS and Emerging ROS-Modulating Technologies for Augmentation of the Healing Process. Int. Wound J., 2017. [DOI | PubMed]
- K. Raziyeva, Y. Kim, Z. Zharkinbekov, K. Kassymbek, S. Jimi, A. Saparov. Immunology of Acute and Chronic Wound Healing. Biomolecules, 2021. [DOI | PubMed]
- A. Saleh, A. Macia, A. R. Muotri. Transposable Elements, Inflammation, and Neurological Disease. Front. Neurol., 2019. [DOI | PubMed]
- P. Krzyszczyk, R. Schloss, A. Palmer, F. Berthiaume. The Role of Macrophages in Acute and Chronic Wound Healing and Interventions to Promote Pro-Wound Healing Phenotypes. Front. Physiol., 2018. [DOI | PubMed]
- T. J. Koh, L. A. DiPietro. Inflammation and Wound Healing: The Role of the Macrophage. Expert Rev. Mol. Med., 2011. [DOI | PubMed]
- M. Xue, C. J. Jackson. Extracellular Matrix Reorganization During Wound Healing and Its Impact on Abnormal Scarring. Adv. Wound Care, 2015. [DOI]
- X. Nie, J. Zhao, H. Ling, Y. Deng, X. Li, Y. He. Exploring microRNAs in Diabetic Chronic Cutaneous Ulcers: Regulatory Mechanisms and Therapeutic Potential. Br. J. Pharmacol., 2020. [DOI | PubMed]
- C. K. Sen. Human Wound and Its Burden: Updated 2020 Compendium of Estimates. Adv. Wound Care, 2021. [DOI]
- M. Malone, T. Bjarnsholt, A. J. McBain, G. A. James, P. Stoodley, D. Leaper, M. Tachi, G. Schultz, T. Swanson, R. D. Wolcott. The Prevalence of Biofilms in Chronic Wounds: A Systematic Review and Meta-Analysis of Published Data. J. Wound Care, 2017. [DOI | PubMed]
- T. M. Haverman, A. M. G. A. Laheij, J. J. de Soet, J. de Lange, F. R. Rozema. Candida and Porphyromonas Gingivalis: The Effect on Wound Closure in Vitro. J. Oral Microbiol., 2017. [DOI | PubMed]
- A. M. Szpaderska, E. I. Egozi, R. L. Gamelli, L. A. DiPietro. The Effect of Thrombocytopenia on Dermal Wound Healing. J. Invest. Dermatol., 2003. [DOI | PubMed]
- J. E. Glim, M. van Egmond, F. B. Niessen, V. Everts, R. H. J. Beelen. Detrimental Dermal Wound Healing: What Can We Learn from the Oral Mucosa?. Wound Repair Regener., 2013. [DOI]
- C. Garlanda, I. Di Ceglie, S. Jaillon. IL-1 Family Cytokines in Inflammation and Immunity. Cell. Mol. Immunol., 2025. [DOI | PubMed]
- I. J. Kosten, R. van de Ven, M. Thon, S. Gibbs, T. D. de Gruijl. Comparative Phenotypic and Functional Analysis of Migratory Dendritic Cell Subsets from Human Oral Mucosa and Skin. PLoS One, 2017. [DOI | PubMed]
- M. Waasdorp, B. P. Krom, F. J. Bikker, P. P. M. van Zuijlen, F. B. Niessen, S. Gibbs. The Bigger Picture: Why Oral Mucosa Heals Better Than Skin. Biomolecules, 2021. [DOI | PubMed]
- R. G. Pertwee. The Diverse CB1 and CB2 Receptor Pharmacology of Three Plant Cannabinoids: Delta9-Tetrahydrocannabinol, Cannabidiol and Delta9-Tetrahydrocannabivarin. Br. J. Pharmacol., 2008. [DOI | PubMed]
- S. Burstein. Cannabidiol (CBD) and Its Analogs: A Review of Their Effects on Inflammation. Bioorg. Med. Chem., 2015. [DOI | PubMed]
- A. J. Singer, R. A. Clark. Cutaneous Wound Healing. N. Engl. J. Med., 1999. [DOI | PubMed]
- P. Martin, R. Nunan. Cellular and Molecular Mechanisms of Repair in Acute and Chronic Wound Healing. Br. J. Dermatol., 2015. [DOI | PubMed]
- K. Kongkadee, W. Wisuitiprot, K. Ingkaninan, N. Waranuch. Anti-Inflammation and Gingival Wound Healing Activities of Cannabis Sativa L. Subsp. Sativa (Hemp) Extract and Cannabidiol: An in Vitro Study. Arch. Oral Biol., 2022. [DOI | PubMed]
- E. Sangiovanni, M. Fumagalli, B. Pacchetti, S. Piazza, A. Magnavacca, S. Khalilpour, G. Melzi, G. Martinelli, M. Dell’Agli. Cannabis sativa L. extract and cannabidiol inhibit in vitro mediators of skin inflammation and wound injury. Phytother. Res., 2019. [DOI | PubMed]
- T. Genovese, M. Cordaro, R. Siracusa, D. Impellizzeri, S. Caudullo, E. Raffone, F. Macrí, L. Interdonato, E. Gugliandolo, C. Interlandi, R. Crupi, R. D’Amico, R. Fusco, S. Cuzzocrea, R. Di Paola. Molecular and Biochemical Mechanism of Cannabidiol in the Management of the Inflammatory and Oxidative Processes Associated with Endometriosis. Int. J. Mol. Sci., 2022. [DOI | PubMed]
- W. Zeng, Y. Wang, R. Gao, H. Wen, M. Yu. Unlocking the Reverse Targeting Mechanisms of Cannabidiol: Unveiling New Therapeutic Avenues. J. Med. Chem., 2024. [DOI | PubMed]
- V. N. Tran, O. Strnad, J. Šuman, T. Veverková, A. Sukupová, P. Cejnar, R. Hynek, O. Kronusová, J. Šach, P. Kaštánek, T. Ruml, J. Viktorová. Cannabidiol Nanoemulsion for Eye Treatment – Anti-Inflammatory, Wound Healing Activity and Its Bioavailability Using in Vitro Human Corneal Substitute. Int. J. Pharm., 2023. [DOI | PubMed]
- E. Kozela, M. Pietr, A. Juknat, N. Rimmerman, R. Levy, Z. Vogel. Cannabinoids Delta(9)-Tetrahydrocannabinol and Cannabidiol Differentially Inhibit the Lipopolysaccharide-Activated NF-kappaB and Interferon-Beta/STAT Proinflammatory Pathways in BV-2 Microglial Cells. J. Biol. Chem., 2010. [DOI | PubMed]
- M. Klein, J. de Quadros De Bortolli, F. S. Guimarães, F. G. Salum, K. Cherubini, M. A. Z. de Figueiredo. Effects of Cannabidiol, a Cannabis Sativa Constituent, on Oral Wound Healing Process in Rats: Clinical and Histological Evaluation. Phytother. Res. PTR, 2018. [DOI | PubMed]
- W. Zhou, L. Zhu, X. Zu, Y. Ma, P. Shen, Y. Hu, P. Zhang, Z. Ni, N. Wang, D. Sun, Z. Bai, X. Gao, C. Huangfu, Y. Gao. A Novel Antioxidant and Anti-Inflammatory Carboxymethylcellulose/Chitosan Hydrogel Loaded with Cannabidiol Promotes the Healing of Radiation-Combined Wound Skin Injury in the 60Co γ-Irradiated Mice. Phytomedicine, 2025. [DOI | PubMed]
- M. Levine, P.-C. Violet. Data Triumph at C. Cancer Cell, 2017. [DOI | PubMed]
- J. Qu, X. Zhao, Y. Liang. Degradable conductive injectable hydrogels as novel antibacterial, anti-oxidant wound dressings for wound healing. Chem. Eng. J., 2019. [DOI]
- M. Dos-Santos-Pereira, F. S. Guimarães, E. Del-Bel, R. Raisman-Vozari, P. P. Michel. Cannabidiol Prevents LPS-Induced Microglial Inflammation by Inhibiting ROS/NF-κB-Dependent Signaling and Glucose Consumption. Glia, 2020. [DOI | PubMed]
- Z. Zheng, J. Qi, L. Hu, D. Ouyang, H. Wang, Q. Sun, L. Lin, L. You, B. Tang. A Cannabidiol-Containing Alginate Based Hydrogel as Novel Multifunctional Wound Dressing for Promoting Wound Healing. Biomater. Adv., 2022. [DOI | PubMed]
- J. M. MatÉs, C. Pérez-Gómez, I. N. De Castro. Antioxidant enzymes and human diseases. Clin. Biochem., 1999. [DOI | PubMed]
- D. Chelminiak-Dudkiewicz, A. Smolarkiewicz-Wyczachowski, K. Mylkie, M. Wujak, D. T. Mlynarczyk, P. Nowak, S. Bocian, T. Goslinski, M. Ziegler-Borowska. Chitosan-Based Films with Cannabis Oil as a Base Material for Wound Dressing Application. Sci. Rep., 2022. [DOI | PubMed]
- W. M. Petroll, H. D. Cavanagh, P. Barry, P. Andrews, J. V. Jester. Quantitative Analysis of Stress Fiber Orientation during Corneal Wound Contraction. J. Cell Sci., 1993. [DOI | PubMed]
- M. Styrczewska, A. Kulma, K. Ratajczak, R. Amarowicz, J. Szopa. Cannabinoid-like Anti-Inflammatory Compounds from Flax Fiber. Cell. Mol. Biol. Lett., 2012. [DOI | PubMed]
- M. Dai, X. Zheng, X. Xu, X. Kong, X. Li, G. Guo, F. Luo, X. Zhao, Y. Q. Wei, Z. Qian. Chitosan-Alginate Sponge: Preparation and Application in Curcumin Delivery for Dermal Wound Healing in Rat. J. Biomed. Biotechnol., 2009. [DOI | PubMed]
- M. D. Shoulders, R. T. Raines. Collagen Structure and Stability. Annu. Rev. Biochem., 2009. [DOI | PubMed]
- X. Qi, C. Liu, G. Li, H. Luan, S. Li, D. Yang, Z. Zhou. Investigation of in Vitro Odonto/Osteogenic Capacity of Cannabidiol on Human Dental Pulp Cell. J. Dent., 2021. [DOI | PubMed]
- I. V. Yannas, D. S. Tzeranis, P. T. C. So. Regeneration of Injured Skin and Peripheral Nerves Requires Control of Wound Contraction, Not Scar Formation. Wound Repair Regener., 2017. [DOI]
- K. S. Gangopadhyay, M. Khan, S. Pandit. Pharmacological Evaluation and Chemical Standardization of an Ayurvedic Formulation for Wound Healing Activity. Int. J. Lower Extremity Wounds, 2014. [DOI]
- V. C. McIver, A. S. Tsang, N. E. Symonds, N. R. Perkins, E. Uquillas, C. M. Dart, L. B. Jeffcott, A. J. Dart. Effects of Topical Treatment of Cannabidiol Extract in a Unique Manuka Factor 5 Manuka Honey Carrier on Second Intention Wound Healing on Equine Distal Limb Wounds: A Preliminary Study. Aust. Vet. J., 2020. [DOI | PubMed]
- E. A. Formukong, A. T. Evans, F. J. Evans. Analgesic and Antiinflammatory Activity of Constituents of Cannabis Sativa L. Inflammation, 1988. [DOI | PubMed]
- S. Atalay, I. Jarocka-Karpowicz, E. Skrzydlewska. Antioxidative and Anti-Inflammatory Properties of Cannabidiol. Antioxidants, 2020. [DOI]
- B. I. Tóth, N. Dobrosi, A. Dajnoki, G. Czifra, A. Oláh, A. G. Szöllosi, I. Juhász, K. Sugawara, R. Paus, T. Bíró. Endocannabinoids Modulate Human Epidermal Keratinocyte Proliferation and Survival via the Sequential Engagement of Cannabinoid Receptor-1 and Transient Receptor Potential Vanilloid-1. J. Invest. Dermatol., 2011. [DOI | PubMed]
- Epigenetic control of skin differentiation genes by phytocannabinoids. Br. J. Pharmacol.,, 2013. [DOI | PubMed]
- L. Claes, S. Recknagel, A. Ignatius. Fracture Healing under Healthy and Inflammatory Conditions. Nat. Rev. Rheumatol., 2012. [DOI | PubMed]
- I. H. Kalfas. Principles of Bone Healing. Neurosurg. Focus, 2001. [DOI]
- J. A. Siddiqui, N. C. Partridge. Physiological Bone Remodeling: Systemic Regulation and Growth Factor Involvement. Physiology, 2016. [DOI | PubMed]
- P.-L. Chung, S. Zhou, B. Eslami, L. Shen, M. S. LeBoff, J. Glowacki. Effect of Age on Regulation of Human Osteoclast Differentiation. J. Cell. Biochem., 2014. [DOI | PubMed]
- M. N. Weitzmann, R. Pacifici. Estrogen Deficiency and Bone Loss: An Inflammatory Tale. J. Clin. Invest., 2006. [DOI | PubMed]
- G. Hajishengallis, R. J. Lamont. Beyond the Red Complex and into More Complexity: The Polymicrobial Synergy and Dysbiosis (PSD) Model of Periodontal Disease Etiology. Mol. Oral Microbiol., 2012. [DOI | PubMed]
- W. J. Boyle, W. S. Simonet, D. L. Lacey. Osteoclast Differentiation and Activation. Nature, 2003. [DOI | PubMed]
- S. C. Holt, J. L. Ebersole. Porphyromonas Gingivalis, Treponema Denticola, and Tannerella Forsythia: The “Red Complex”, a Prototype Polybacterial Pathogenic Consortium in Periodontitis. Periodontology 2000, 2005. [DOI | PubMed]
- M. Usui, S. Onizuka, T. Sato, S. Kokabu, W. Ariyoshi, K. Nakashima. Mechanism of Alveolar Bone Destruction in Periodontitis – Periodontal Bacteria and Inflammation. Jpn. Dent. Sci. Rev., 2021. [DOI | PubMed]
- H. Takayanagi. Osteoimmunology: Shared Mechanisms and Crosstalk between the Immune and Bone Systems. Nat. Rev. Immunol., 2007. [DOI | PubMed]
- H. Takayanagi, S. Kim, T. Koga, H. Nishina, M. Isshiki, H. Yoshida, A. Saiura, M. Isobe, T. Yokochi, J. Inoue, E. F. Wagner, T. W. Mak, T. Kodama, T. Taniguchi. Induction and Activation of the Transcription Factor NFATc1 (NFAT2) Integrate RANKL Signaling in Terminal Differentiation of Osteoclasts. Dev. Cell, 2002. [DOI | PubMed]
- S. Zhu, M.-Q. Yan, A. Masson, W. Chen, Y.-P. Li. Cell Signaling and Transcriptional Regulation of Osteoclast Lineage Commitment, Differentiation, Bone Resorption and Diseases. Cell Discovery, 2026. [DOI | PubMed]
- K. Sato, A. Suematsu, K. Okamoto, A. Yamaguchi, Y. Morishita, Y. Kadono, S. Tanaka, T. Kodama, S. Akira, Y. Iwakura, D. J. Cua, H. Takayanagi. Th17 Functions as an Osteoclastogenic Helper T Cell Subset That Links T Cell Activation and Bone Destruction. J. Exp. Med., 2006. [DOI | PubMed]
- A. Kamali, A. Oryan, S. Hosseini, M. H. Ghanian, M. Alizadeh, M. Baghaban Eslaminejad, H. Baharvand. Cannabidiol-Loaded Microspheres Incorporated into Osteoconductive Scaffold Enhance Mesenchymal Stem Cell Recruitment and Regeneration of Critical-Sized Bone Defects. Mater. Sci. Eng.:C, 2019. [DOI]
- M.-A. Kang, J. Lee, S.-H. Park. Cannabidiol Induces Osteoblast Differentiation via Angiopoietin1 and P38 MAPK. Environ. Toxicol., 2020. [DOI | PubMed]
- N. M. Kogan, E. Melamed, E. Wasserman, B. Raphael, A. Breuer, K. S. Stok, R. Sondergaard, A. V. V. Escudero, S. Baraghithy, M. Attar-Namdar, S. Friedlander-Barenboim, N. Mathavan, H. Isaksson, R. Mechoulam, R. Müller, A. Bajayo, Y. Gabet, I. Bab. Cannabidiol, a Major Non-Psychotropic Cannabis Constituent Enhances Fracture Healing and Stimulates Lysyl Hydroxylase Activity in Osteoblasts. J. Bone Miner. Res., 2015. [DOI | PubMed]
- D. Apostu, O. Lucaciu, A. Mester, H. Benea, D. Oltean-Dan, F. Onisor, M. Baciut, S. Bran. Cannabinoids and Bone Regeneration. Drug Metab. Rev., 2019. [DOI | PubMed]
- S. A. Bonini, M. Premoli, S. Tambaro, A. Kumar, G. Maccarinelli, M. Memo, A. Mastinu. Cannabis Sativa: A Comprehensive Ethnopharmacological Review of a Medicinal Plant with a Long History. J. Ethnopharmacol., 2018. [DOI | PubMed]
- R. B. Laprairie, A. M. Bagher, M. E. M. Kelly, E. M. Denovan-Wright. Cannabidiol Is a Negative Allosteric Modulator of the Cannabinoid CB1 Receptor. Br. J. Pharmacol., 2015. [DOI | PubMed]
- A. I. Idris, R. J. van ‘t Hof, I. R. Greig, S. A. Ridge, D. Baker, R. A. Ross, S. H. Ralston. Regulation of Bone Mass, Bone Loss and Osteoclast Activity by Cannabinoid Receptors. Nat. Med., 2005. [DOI | PubMed]
- O. Ofek, M. Karsak, N. Leclerc, M. Fogel, B. Frenkel, K. Wright, J. Tam, M. Attar-Namdar, V. Kram, E. Shohami, R. Mechoulam, A. Zimmer, I. Bab. Peripheral Cannabinoid Receptor, CB2, Regulates Bone Mass. Proc. Natl. Acad. Sci. U.S.A., 2006. [DOI | PubMed]
- S. S. R. Nielsen, J. A. Z. Pedersen, N. Sharma. Human osteoclasts in vitro are both inhibited and stimulated dose-dependently by cannabidiol (CBD) and Δ9-tetrahydrocannabinol (THC). Bone Rep., 2022. [DOI]
- Y. Zhang, S. E. Chen, J. Shao, J. J. J. P. Van Den Beucken. Combinatorial Surface Roughness Effects on Osteoclastogenesis and Osteogenesis. ACS Appl. Mater. Interfaces, 2018. [DOI | PubMed]
- L. Li, J. Feng, L. Sun, Y.-W. Xuan, L. Wen, Y.-X. Li, S. Yang, B. Zhu, X.-Y. Tian, S. Li, L.-S. Zhao, R.-J. Dang, T. Jiao, H.-S. Zhang, N. Wen. Cannabidiol Promotes Osteogenic Differentiation of Bone Marrow Mesenchymal Stem Cells in the Inflammatory Microenvironment via the CB2-Dependent P38 MAPK Signaling Pathway. Int. J. Stem Cells, 2022. [DOI | PubMed]
- L. Yu, L. Zeng, Z. Zhang, G. Zhu, Z. Xu, J. Xia, J. Weng, J. Li, J. L. Pathak. Cannabidiol Rescues TNF-α-Inhibited Proliferation, Migration, and Osteogenic/Odontogenic Differentiation of Dental Pulp Stem Cells. Biomolecules, 2023. [DOI | PubMed]
- E. Ryberg, N. Larsson, S. Sjögren, S. Hjorth, N.-O. Hermansson, J. Leonova, T. Elebring, K. Nilsson, T. Drmota, P. J. Greasley. The Orphan Receptor GPR55 Is a Novel Cannabinoid Receptor. Br. J. Pharmacol., 2007. [DOI | PubMed]
- L. S. Whyte, E. Ryberg, N. A. Sims, S. A. Ridge, K. Mackie, P. J. Greasley, R. A. Ross, M. J. Rogers. The Putative Cannabinoid Receptor GPR55 Affects Osteoclast Function in Vitro and Bone Mass in Vivo. Proc. Natl. Acad. Sci. U.S.A., 2009. [DOI | PubMed]
- E. Schmuhl, R. Ramer, A. Salamon, K. Peters, B. Hinz. Increase of Mesenchymal Stem Cell Migration by Cannabidiol via Activation of P42/44 MAPK. Biochem. Pharmacol., 2014. [DOI | PubMed]
- M. G. Mosca, M. Mangini, S. Cioffi, P. Barba, S. Mariggiò. Peptide Targeting of Lysophosphatidylinositol-Sensing GPR55 for Osteoclastogenesis Tuning. Cell Commun. Signal. CCS, 2021. [DOI | PubMed]
- L. Li, J. Feng, L. Sun, Y.-W. Xuan, L. Wen, Y.-X. Li, S. Yang, B. Zhu, X.-Y. Tian, S. Li, L.-S. Zhao, R.-J. Dang, T. Jiao, H.-S. Zhang, N. Wen. Cannabidiol Promotes Osteogenic Differentiation of Bone Marrow Mesenchymal Stem Cells in the Inflammatory Microenvironment via the CB2-dependent P38 MAPK Signaling Pathway. Int. J. Stem Cells, 2022. [DOI | PubMed]
- M. Pucci, C. Rapino, A. Di Francesco, E. Dainese, C. D’Addario, M. Maccarrone. Epigenetic Control of Skin Differentiation Genes by Phytocannabinoids. Br. J. Pharmacol., 2013. [DOI | PubMed]
- P. H. M. P. Tolentino, R. G. Dinelli, H. S. Garzón, D. R. Suárez, M. A. T. Corral, E. P. Christoff, M. R. Bueno, U. M. Sandi, L. J. Suárez, B. Bueno-Silva. Evaluation of the Antimicrobial Effect of Cannabidiol (CBD) in a Multispecies Subgingival Biofilm Model. J. Oral Microbiol., 2026. [DOI | PubMed]
- EMEA-000181-PIP02–13-M01 – paediatric investigation plan, European Medicines Agency (EMA) 2026 https://www.EMA.europa.eu/en/medicines/human/paediatric-investigation-plans/13-m01. (accessed February 24, 2026).
- C. A. Legare, W. M. Raup-Konsavage, K. E. Vrana. Therapeutic Potential of Cannabis, Cannabidiol, and Cannabinoid-Based Pharmaceuticals. Pharmacology, 2022. [DOI | PubMed]
- T. E. Gaston, D. Friedman. Pharmacology of Cannabinoids in the Treatment of Epilepsy. Epilepsy Behav., 2017. [DOI | PubMed]
- M. A. Huestis. Human Cannabinoid Pharmacokinetics. Chem. Biodivers., 2007. [DOI | PubMed]
- M. H, T. Sr, B. Sk, B. A, C. F, M. N. The Transdermal Delivery of Therapeutic Cannabinoids. Pharmaceutics, 2022. [DOI | PubMed]
- A. Ohlsson, J. E. Lindgren, S. Andersson, S. Agurell, H. Gillespie, L. E. Hollister. Single-Dose Kinetics of Deuterium-Labelled Cannabidiol in Man after Smoking and Intravenous Administration. Biomed. Environ. Mass Spectrom., 1986. [DOI | PubMed]
- F. Grotenhermen. Pharmacokinetics and Pharmacodynamics of Cannabinoids. Clin. Pharmacokinet., 2003. [DOI | PubMed]
- C. J. Lucas, P. Galettis, J. Schneider. The Pharmacokinetics and the Pharmacodynamics of Cannabinoids. Br. J. Clin. Pharmacol., 2018. [DOI | PubMed]
- S. A. Millar, N. L. Stone, A. S. Yates, S. E. O’Sullivan. A Systematic Review on the Pharmacokinetics of Cannabidiol in Humans. Front. Pharmacol., 2018. [DOI]
- A. Zgair, J. C. Wong, J. B. Lee, J. Mistry, O. Sivak, K. M. Wasan, I. M. Hennig, D. A. Barrett, C. S. Constantinescu, P. M. Fischer, P. Gershkovich. Dietary Fats and Pharmaceutical Lipid Excipients Increase Systemic Exposure to Orally Administered Cannabis and Cannabis-Based Medicines. Am. J. Transl. Res., 2016. [PubMed]
- Preuss, C. V. ; Kalava, A. ; King, K. C. Prescription of Controlled Substances: Benefits and Risks; StatPearls Publishing: Treasure Island (FL), 2025.
- A. Charalabidis, M. Sfouni, C. Bergström, P. Macheras. The Biopharmaceutics Classification System (BCS) and the Biopharmaceutics Drug Disposition Classification System (BDDCS): Beyond Guidelines. Int. J. Pharm., 2019. [DOI | PubMed]
- S. Sim, N. K. Wong. Nanotechnology and Its Use in Imaging and Drug Delivery (Review). Biomed. Rep., 2021. [DOI | PubMed]
- A. Yusuf, A. R. Z. Almotairy, H. Henidi, O. Y. Alshehri, M. S. Aldughaim. Nanoparticles as Drug Delivery Systems: A Review of the Implication of Nanoparticles’ Physicochemical Properties on Responses in Biological Systems. Polymers, 2023. [DOI | PubMed]
- D. Bhattacharya, B. Ghosh, M. Mukhopadhyay. Development of Nanotechnology for Advancement and Application in Wound Healing: A Review. IET Nanobiotechnol., 2019. [DOI | PubMed]
- B.-A. Chuluunbaatar, J. Park, J. Song, S. Mun, J.-H. Kang, K. H. Min. Cannabidiol-Loaded Mucoadhesive PLGA Nanosphere-Chitosan Hydrogel Patch for Oral Therapeutic Applications. Int. J. Mol. Sci., 2026. [DOI | PubMed]
- J. Qi, Z. Zheng, L. Hu, H. Wang, B. Tang, L. Lin. Development and Characterization of Cannabidiol-Loaded Alginate Copper Hydrogel for Repairing Open Bone Defects in Vitro. Colloids Surf., B, 2022. [DOI]
- Cannabidiol,. 2026
- Pereira, P. R. T. Ministério da Saúde..
- P. Cáceres Guido, N. Riva, G. Calle, M. Dell’Orso, M. Gatto, N. Sberna, P. Schaiquevich. Medicinal Cannabis in Latin America: History, Current State of Regulation, and the Role of the Pharmacist in a New Clinical Experience with Cannabidiol Oil. J. Am. Pharm. Assoc., 2020. [DOI]