
Long term exposure to dietary cannabinoids inhibits rapid functional tolerance to ethanol in Drosophila melanogaster adults
Abstract
In rodents, exposure to cannabidiol (CBD) was shown to alter responses to alcohol, primarily via CB1/CB2 signaling. However, CBD also targets non-canonical receptors whose potential roles in modulating CBD-alcohol interactions remain unknown. We used a simple behavioral assay to compare responses to ethanol in Drosophila melanogaster , which lacks CB1/CB2 genes but has orthologs for non-canonical targets. Flies fed with a CBD-enriched diet retain baseline ethanol sensitivity but fail to develop rapid functional tolerance to ethanol. These findings support using Drosophila as a model to study non-canonical CBD signaling in ethanol behaviors relevant to the development of alcohol use disorder.
Affiliations: Biology, California State University, Northridge, Northridge, California, United States
License: Copyright: © 2025 by the authors CC BY 4.0 This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Article links: DOI: 10.17912/micropub.biology.001825 | PubMed: 41459254 | PMC: PMC12741748
Relevance: Relevant: mentioned in keywords or abstract
Full text: PDF (733 KB)
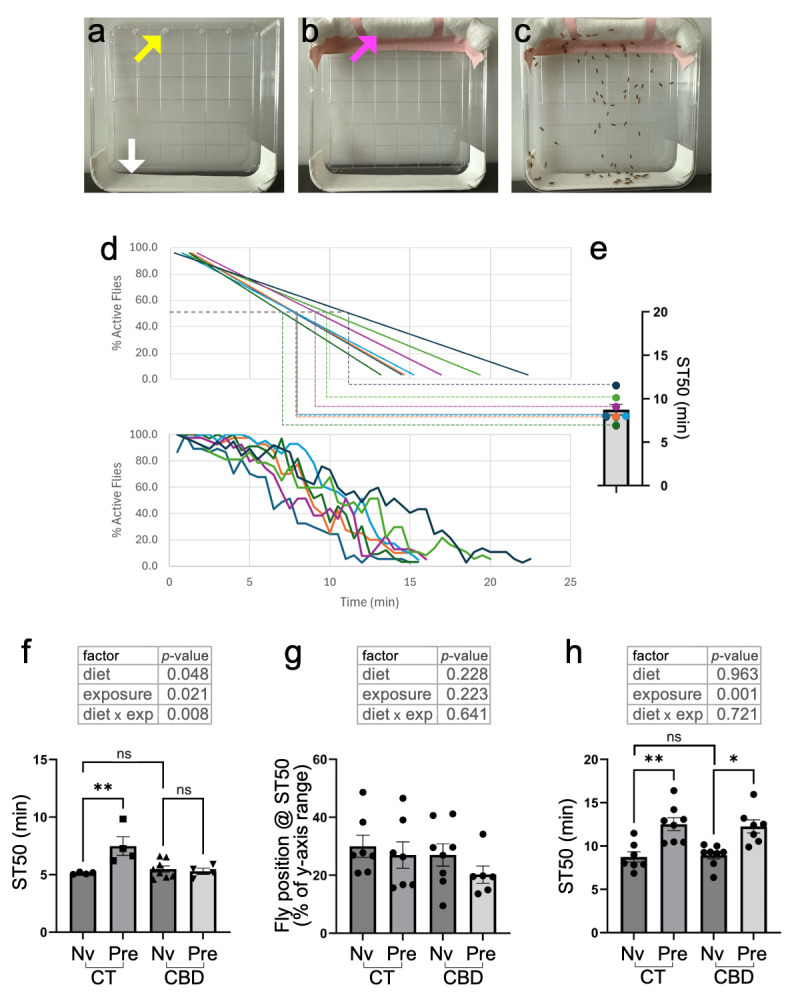
Figure 1.
: (a) Ethanol sedation cages are prepared by perforating four pinholes near the edge on the back side of a 10cm square Petri dish (yellow arrow), and affixing clear tape to the lateral side on the opposite end (white arrow). This needs to be done only once. (b) Prior to each sedation assay, a 10cm-wide gauze rod is secured with tape to the top of the cage, covering the pinholes (magenta arrow). (c) Example of a picture with flies being exposed to ethanol vapors by soaking the gauze rod with 4mL of ethanol and sealing the cage. All non-sedated flies across pictures were spotted using the multi-selection tool in ImageJ/Fiji, and Y-coordinate values were used to calculate average %Active flies and %Y-climb (as explained in Methods). (d) Raw line plots of %Active flies over time (bottom), and their corresponding linear regressions (top). A standard line equation was used to calculate ST50 values, i.e., the time at which 50% of the flies in a group are projected to become sedated. (e) ST50 values from each student group were pooled as biological replicates for each sample in subsequent statistical analyses. (f) ST50 values of adult flies on a 14-days control (CT) or cannabinoid (CBD) diet, and exposed to ethanol vapors for the first time (naïve, Nv) or following a prior intoxication, 4 hrs earlier (Pre-exposed, Pre). (g) Average position of flies along the y-axis (expressed as % of y-axis range) at their corresponding ST50 times. (h) Comparison of ST50 values across groups as in (f), except that the flies were exposed to cannabinoids (or not) as larvae, not as adults. The top tables in (f) thru (h) show the outcomes of 2-way ANOVAs, testing data across diet (CT or CBD), exposure (Nv or Pre) and their interaction. It should be noted, however, that not all samples passed a Shapiro-Wilk’s test of normality.
Description
There has been increased interest in the potential health benefits of cannabidiol (CBD) and other non-psychotropic cannabinoids, particularly in the context of their anti-inflammatory, neuroprotective and analgesic effects (Bhunia et al., 2022; Cásedas et al., 2024; Voicu et al., 2023). In mammals, cannabinoids are thought to function primarily via their cognate G protein-coupled receptors CB1 and CB2 (reviewed in Shabazi et al., 2020 and Coelho et al., 2023). However, cannabinoids have also been shown to exert a broad range of physiological and neurological phenotypes via so-called non-canonical molecular targets, such as TRPV1-4 and TRPA1 (Etemad et al., 2022), GPR55 (Rosenberg et al., 2023), PPARγ (Khosropoor et al., 2023; Puighermanal et al., 2024), the 5-HT1A serotonin receptor (Linge et al., 2016; Liu et al., 2024) and the Hedgehog pathway intermediate Smoothened (Khaliullina et al., 2015). The Drosophila melanogaster genome does not encode homologs of the CB1 and CB2 receptors (McPartland et al., 2001; McPartland et al., 2006). However, previous studies have already shown effects of cannabis extracts and synthetic cannabinoids on diverse aspects of D. melanogaster biology (Ahn et al., 2021; Candib et al., 2024; He, Tan, et al., 2021; Kawasaki et al., 2025). Therefore, fruit flies could serve as an excellent experimental model to dissect the genetic mechanisms of cannabinoid function via any of the CB1/2-independent targets listed above, for all of which there exist Drosophila homologs. For instance, Jacobs and Sehgal showed that a metabolite derived from the endocannabinoid anandamide protects flies against seizures, and used a targeted genetic screen to determine that this effect was at least partially mediated by the TRP channel water witch (Jacobs & Sehgal, 2020).
Given that both alcohol and cannabis use are associated with a significant risk of developing into corresponding use disorders and addiction, it is not surprising that there has been a long-standing interest in understanding their potential interactions at both the neuropharmacological and molecular level. Much of the previous work in this area has focused on the role of endocannabinoids and CB1-dependent signaling on sensitivity and tolerance development to ethanol (reviewed in Pava & Woodward, 2012), with relatively less known about the potential role of non-canonical cannabinoid signaling on an organism’s response to alcohol. Of note, exposure to CBD and other non-psychoactive cannabinoids has been shown to attenuate behaviors related to ethanol tolerance and addiction in rodent models (reviewed in Turna et al., 2019), and diverse clinical trials have been designed to explore the use of cannabidiol and related substances for the treatment of substance use disorders (reviewed in Morel et al., 2021).
Foundational work by Scholz and colleagues demonstrated that flies can develop rapid functional tolerance to ethanol (RFTE; Scholz et al., 2000). Briefly, flies that have been pre-exposed to intoxicating doses of ethanol will show decreased sensitivity to a second ethanol exposure shortly after (hence, ‘rapid’), even when the internal ethanol concentration after the second exposure is comparable to that achieved after the first (hence, ‘functional’). Since this seminal discovery, Drosophila has served as an excellent model to study the neuronal, molecular and genetic mechanisms behind the development of RFTE (for reviews highlighting different advances in the field see Chvilicek et al., 2020; Engel et al., 2019 and Park et al., 2017). These previous studies have not only led to a rich theoretical framework for future studies, but also to the development of simple and scalable behavioral assays that can be easily implemented in the context of a Course-based Undergraduate Research Experience (CURE) class (e.g., Bhandari et al., 2009 and Sandhu et al., 2015). Therefore, we decided to explore whether long-term exposure to dietary phytocannabinoids might affect the ability of flies to develop rapid functional ethanol tolerance through a 300-level CURE offered at our institution. The work presented here was inspired by a previous study by He and colleagues, who reported that young males exposed to dietary cannabinoids for 2 and 4 days did not significantly affect their baseline sensitivity to ethanol, nor their ability to develop RFTE (He, Ng, et al., 2021). In our study, we decided to explore if exposing female flies to food containing cannabinoids for a longer period (14 days) might affect their ability to develop RFTE.
When adult females were maintained on a CBD diet for 14 days, their baseline sensitivity to ethanol sedation was comparable to that of flies on a control diet (CBD_Nv vs. CT_N; Fig. 1f). As expected, control flies pre-exposed to an intoxicating dose of ethanol showed a significant increase in ST50 (CT_N vs. CT_Pre; Fig. 1f), reflecting their ability to develop RFTE. Surprisingly, flies on a 14-days CBD diet did not show a significant increase in ST50 following a pre-exposure to ethanol (CBD_N vs. CBD_Pre; Fig. 1f), indicating that they had lost such ability. We also tested the mean climbing percent of flies across groups at their corresponding ST50; i.e., how high, on average, the remaining 50% of unsedated flies in a group could climb at the ST50. This comparison revealed no significant differences across all groups (Fig. 1g), suggesting that prior intoxications and pre-exposure to dietary CBD affected the onset of sedation and the ability of flies to develop RFTE, but not the climbing capacity of those that remain unsedated.
Lastly, we sought to determine if flies exposed to CBD during larval development, instead of as adults, would also lose their ability to develop RFTE. Adult parents were kept for 4 days to lay eggs in bottles containing regular food (CT) or supplemented with CBD oil (CBD), and then removed. Their progeny was then left to develop into adulthood in either diet, and adults were tested 1-4 days after eclosion for their ability to develop RFTE. We did not observe any noticeable differences in larval size, eclosion rates or the size and overall behavior of adults under either diet. In addition, there was no difference in the baseline ethanol sensitivity of flies that developed in regular vs. CBD food (CT_N vs. CBD_N; Fig. 1h), and flies exposed to CBD during larval development retained their ability to develop RTFE, as shown by a significant increase in ST50 following pre-exposure to ethanol in both groups and a non-significant interaction between diet and pre-exposure to ethanol in a 2-way ANOVA (Fig. 1h).
Our findings are inconsistent with a report by He et al., who showed that pre-exposure to cannabinoids did not significantly affect the ability of adult flies to develop RFTE (He, Ng, et al., 2021). However, there exist two important differences between our studies that may warrant deeper exploration: i) the sex and genetic strain of the flies used ( Canton S males in He, Ng et al., 2021 vs. w 1118 females in our study); ii) the time of exposure to cannabinoids (2-4 days for He, Ng et al., 2021 vs. 14-days in this study).
Given that CBD acts as a natural antagonist of CB1R, and CB1R antagonists have been shown to reduce the self-administration and appetitive value of ethanol in rodent models (Arnone et al., 1997; Economidou et al., 2006), as well as inhibit the development of rapid tolerance to ethanol in rats (Lemos et al., 2007), CBD had long been viewed as a promising pharmacotherapy candidate to treat alcohol used disorders (AUD). Accordingly, CBD treatment was shown to inhibit the context-dependent relapse into alcohol seeking in rats with a history of alcohol consumption (Gonzalez-Cuevas et al., 2018), and their ability to develop tolerance to sedation by alcohol following re-exposure (Szulc et al., 2023). Not surprisingly, most previous work has focused on the potential role of CB1/2R-dependent signaling in the cross-talk between ethanol and CBD. Notably, Viudez-Martinez and colleagues showed that a 5HT1A serotonin receptor antagonist could block the reduction in alcohol self-administration triggered by the combined treatment with CBD and naltrexone (Viudez-Martínez et al., 2018), establishing a precedent for the potential involvement of non-canonical cannabinoid signaling in the cross-talk between CBD and alcohol.
It should be noted that we cannot rule out indirect effects of CBD on a fly’s response to ethanol. For instance, exposure of adults to a CBD diet for more than two days was shown to slightly but significantly reduce appetite in flies (He, Tan et. al, 2021). If our longer-term exposure to CBD effectively reduced their appetite even further, the observed effect of CBD could potentially be explained by a hidden connection between reduced nutrition and a diminished capacity for RFTE. Similar hypotheses could be explored for a diverse range of physiological axes, making this experimental paradigm particularly appealing in the context of a CURE class, in which students without prior Drosophila experience can still formulate and test a wide range of their own mechanistic hypotheses (e.g. effect of sex, age, genetic background, CBD and ethanol dosing, temperature, co-treatments, social stress, etc).
More broadly, and whether through more direct or indirect mechanisms, the findings reported here suggest that Drosophila could be added as a powerful experimental model to further explore the role of non-canonical pathways in the development of alcohol tolerance, and their potential use as new therapeutic targets to treat AUD.
Methods
Two vials containing 45 females and 15 males from a standard w 1118 stock were maintained for 14 days in 2mL of control or CBD food, and flipped every 2-3 days (see Reagents for details). CBD food was prepared by dissolving 120mg of the original oil in 1mL ethanol (used as a carrier), and diluted 1/230 into molten fly food at or below 65°C during preparation (for a 0.52mg/mL, or approximately 1.65mM final concentration). On day 15, flies were pooled and females from each group (CT or CBD) were re-sorted into regular food vials (40 flies/vial) and left to recover overnight.
Ethanol sedation cages were prepared once by perforating 4 small (~2-3mm) equidistant holes on the back side of a 10cm-square plastic Petri dish, approximately 5mm from the edge and adding a strip of white tape on the opposite side, to aid in distinguishing between sedated and non-sedated flies (Fig. 1a). Before each sedation assay, a rolled-up 10x10cm medical gauze is taped to the top, covering the pinholes (Fig. 1b).
On the morning of day 16, both vials from each group were randomly labeled as ‘naïve’ (Nv) or ‘pre-exposed’ (Pre), thus generating four experimental groups: CT_Nv; CT_Pre; CBD_Nv and CBD_Pre. Flies in both pre-exposed groups were transferred by tapping into empty vials capped with half-plugs soaked in 1mL of pure ethanol. Flies were exposed to ethanol vapors for 10 minutes and transferred back into regular food vials by tapping and left to recover for 4hrs (keeping the vials horizontal to prevent the sticking of sedated flies to the food surface).
In our CURE class, students transferred the flies to empty vials and “ice-nesthetized” them by incubating the vials on ice for 10 minutes. Ice-nesthetized flies were transferred by gentle tapping into sedation cages, which were immediately sealed with parafilm. Flies were left to recover from ice-nesthesia for 15 minutes at room temperature. After the 15min recovery, 4mL of ethanol were pipetted onto the gauze rod at the top of the cage through the pinholes in the back (1mL/pinhole, to facilitate even distribution). The pinholes were then taped over, the cages were placed in front of a white background and a timer was started. Students took frontal pictures of the cage every 30 seconds (Fig. 1c), until all flies were observed at the bottom of the cage in two consecutive pictures. It should be noted that the sedation cages were not tapped at regular intervals before taking the pictures. Therefore, our assay did not measure sedation as a reduction in the evoked negative gravotaxis reflex triggered by startling in flies, but on a more general loss of motor control and adhesion to vertical surfaces.
Pictures were opened as an image sequence using ImageJ/Fiji, and the multi-point selection tool was used to “spot” all flies, except those visibly laying on their side or backs at the bottom of the cage. The Analyze>Measure function in ImageJ/Fiji was then used to generate a table with X/Y coordinates of all the spotted flies, which were copy/pasted into an MS Excel spreadsheet. Using the MIN and MAX formulas across all time points, the relative position of each fly was re-calculated as (Y val -Y MIN )/(Y MAX -Y MIN )*100, to convert all Y-climb values to a 0-100 scale and standardize measurements across groups. The re-scaled Y-climb values were copied to a cross-tab table (with columns for time points and rows for Y-climb values of spotted flies). Each column was then used to calculate a mean Y-climb for each time point, and the COUNT function was used to calculate the percent of active flies at each time point (as #spotted flies/total flies in the cage*100), both of which were used to generate line plots of mean Y-climb or %Act vs. time (e.g., Fig. 1d). A linear regression was then used to calculate ST50 values for each group (i.e. the time at which 50% of the flies were sedated), by solving (50-Y intercept ) / slope, based on the corresponding line equation. The calculated ST50 were treated as biological replicates across groups (Fig. 1e), and ANOVA was used to compare sedation rates across treatments. The climbing capacity for a sample at ST50 (Fig. 1g), was calculated by solving %Y-climb ST50 = slope * ST50 + Y intercept , based on a linear regression of %Y-climb values over time.
Each experiment was repeated via three independent trials throughout the semester, and only the results consistent across the three trials are reported here. Results were analyzed via parametric or non-parametric 1- and 2-way ANOVAs (followed by pairwise post-hoc comparisons when applicable), using Prism (GraphPad) and the open-source statistical package JASP (ver 0.95.0; https://jasp-stats.org/). Note that only the outcomes of more relevant pairwise comparisons are depicted by brackets in the graphs: i.e., Nv vs. Pre for each diet (reflective of effects on RFTE), and Nv vs. Nv between diets (reflective of effects on baseline sensitivity to ethanol).
Reagents
| Reagent | Description | Source | Pdt/Cat # |
| Fly strain | w 1118 ; + ; + ; + | BDSC | RRID:BDSC_3605 |
| Control food | Drosophila Agar = 1% w/vCornmeal = 5.9% w/vMolasses = 5.0% v/vYeast Extract = 2.4% w/vTegosept = 0.13% w/vPropionic acid = 542mMPhosphoric acid = 2.95mM | GeneseeScientificFisherScientific | 66-10362-10062-11762-10620-258AC149300010A242-4 |
| 29:1 CBD:THC food | Control food + Full spectrum Cannabis Oil (724mg/mL CBD mix [cannabidiol, cannabigerol, cannabinol, cannabichromene, cannabidoilic acid] + 23mg/mL THC) | Proof Operations, Inc | F0103-111523 |
| Square TC plates | 10cm Square Petri Dish | Amazon/BIPEE | B01DBBBZO4 |
| Square gauze pads | 10cm Square 4-ply wipes | Amazon/Perfect Stix | B0791Y5WS6 |
| Vials & Plugs | Narrow PS vials + Flugs | Genesee Scientific | 32-109BF |
| Ethanol | Standard 200-proof ethanol | Pharmco | 111000200 |
References
- Yejin Ahn, Sung Hee Han, Min Guk Kim, Ki-Bae Hong, Woo Jung Kim, Hyung Joo Suh, Kyungae Jo. Anti-depressant effects of ethanol extract from Cannabis sativa (hemp) seed in chlorpromazine-induced Drosophila melanogaster depression model. Pharmaceutical Biology, 2021. [DOI]
- Michèle Arnone, Jeanne Maruani, Frédérique Chaperon, Marie-Hélène Thiébot, M. Poncelet, Philippe Soubrié, Gérard Le Fur. Selective inhibition of sucrose and ethanol intake by SR 141716, an antagonist of central cannabinoid (CB1) receptors. Psychopharmacology, 1997. [DOI | PubMed]
- Poonam Bhandari, Kenneth S. Kendler, Jill C. Bettinger, Andrew G. Davies, Mike Grotewiel. An Assay for Evoked Locomotor Behavior in Drosophila Reveals a Role for Integrins in Ethanol Sensitivity and Rapid Ethanol Tolerance. Alcoholism: Clinical and Experimental Research, 2009. [DOI | PubMed]
- Sukanya Bhunia, Nagesh Kolishetti, Adriana Yndart Arias, Arti Vashist, Madhavan Nair. Cannabidiol for neurodegenerative disorders: A comprehensive review. Frontiers in Pharmacology, 2022. [DOI]
- Alec Candib, Nicholas Lee, Natasha Sam, Eddie Cho, Jesse Rojas, Reina Hastings, Kyle DeAlva, Diana Khon, Andrea Gonzalez, Brandon Molina, Gina Torabzadeh, Josephine Vu, Kyle Hasenstab, Karylin Sant, Joy A. Phillips, Kim Finley. The Influence of Cannabinoids on Drosophila Behaviors, Longevity, and Traumatic Injury Responses of the Adult Nervous System. Cannabis and Cannabinoid Research, 2024. [DOI | PubMed]
- Guillermo Cásedas, Martín de Yarza-Sancho, Víctor López. Cannabidiol (CBD): A Systematic Review of Clinical and Preclinical Evidence in the Treatment of Pain. Pharmaceuticals, 2024. [DOI | PubMed]
- Maggie M. Chvilicek, Iris Titos, Adrian Rothenfluh. The Neurotransmitters Involved in Drosophila Alcohol-Induced Behaviors. Frontiers in Behavioral Neuroscience, 2020. [DOI]
- Mariana Pinto Coelho, Patrícia Duarte, Marta Calado, António J. Almeida, Catarina Pinto Reis, M. Manuela Gaspar. The current role of cannabis and cannabinoids in health: A comprehensive review of their therapeutic potential. Life Sciences, 2023. [DOI | PubMed]
- Daina Economidou, Laura Mattioli, Carlo Cifani, Marina Perfumi, Maurizio Massi, Vincenzo Cuomo, Luigia Trabace, Roberto Ciccocioppo. Effect of the cannabinoid CB1 receptor antagonist SR-141716A on ethanol self-administration and ethanol-seeking behaviour in rats. Psychopharmacology, 2005. [DOI | PubMed]
- Gregory L. Engel, Kreager Taber, Elizabeth Vinton, Amanda J. Crocker. Studying alcohol use disorder using Drosophila melanogaster in the era of ‘Big Data’. Behavioral and Brain Functions, 2019. [DOI]
- Leila Etemad, Gholamreza Karimi, Mohaddeseh Sadat Alavi, Ali Roohbakhsh. Pharmacological effects of cannabidiol by transient receptor potential channels. Life Sciences, 2022. [DOI | PubMed]
- Gustavo Gonzalez-Cuevas, Remi Martin-Fardon, Tony M. Kerr, David G. Stouffer, Loren H. Parsons, Dana C. Hammell, Stan L. Banks, Audra L. Stinchcomb, Friedbert Weiss. Unique treatment potential of cannabidiol for the prevention of relapse to drug use: preclinical proof of principle. Neuropsychopharmacology, 2018. [DOI | PubMed]
- Jianzheng He, Si Yun Ng, Alice Mei Xien Tan, Wei Lin Yong, Fengwei Yu. Cannabinoids differentially modulate behavioral and developmental responses to ethanol in Drosophila. 2021. [DOI]
- Jianzheng He, Alice Mei Xien Tan, Si Yun Ng, Menglong Rui, Fengwei Yu. Cannabinoids modulate food preference and consumption in Drosophila melanogaster. Scientific Reports, 2021. [DOI]
- Jack A. Jacobs, Amita Sehgal. Anandamide Metabolites Protect against Seizures through the TRP Channel Water Witch in Drosophila melanogaster. Cell Reports, 2020. [DOI | PubMed]
- Haruhisa Kawasaki, Toshihiko Sato, Norio Ishida. Effects of cannabidiol to circadian period, sleep, life span, close-proximity rhythm, egg reproduction and motor function in Drosophila melanogaster. Biogerontology, 2025. [DOI]
- Helena Khaliullina, Mesut Bilgin, Julio L. Sampaio, Andrej Shevchenko, Suzanne Eaton. Endocannabinoids are conserved inhibitors of the Hedgehog pathway. Proceedings of the National Academy of Sciences, 2015. [DOI]
- Sara Khosropoor, Mohaddeseh Sadat Alavi, Leila Etemad, Ali Roohbakhsh. Cannabidiol goes nuclear: The role of PPARγ. Phytomedicine, 2023. [DOI | PubMed]
- Jose Inácio Lemos, Reinaldo Naoto Takahashi, Gina Struffaldi Morato. Effects of SR141716 and WIN 55,212-2 on tolerance to ethanol in rats using the acute and rapid procedures. Psychopharmacology, 2007. [DOI | PubMed]
- Raquel Linge, Laura Jiménez-Sánchez, Leticia Campa, Fuencisla Pilar-Cuéllar, Rebeca Vidal, Angel Pazos, Albert Adell, Álvaro Díaz. Cannabidiol induces rapid-acting antidepressant-like effects and enhances cortical 5-HT/glutamate neurotransmission: role of 5-HT1A receptors. Neuropharmacology, 2016. [DOI | PubMed]
- Yu-Meng Liu, Jin-Cao Li, Yong-Fang Gu, Ren-Hong Qiu, Jia-Ying Huang, Rui Xue, Shuo Li, Yang Zhang, Kuo Zhang, You-Zhi Zhang. Cannabidiol Exerts Sedative and Hypnotic Effects in Normal and Insomnia Model Mice Through Activation of 5-HT1A Receptor. Neurochemical Research, 2024. [DOI | PubMed]
- J. M. Mcpartland, J. Agraval, D. Gleeson, K. Heasman, M. Glass. Cannabinoid receptors in invertebrates. Journal of Evolutionary Biology, 2006. [DOI | PubMed]
- John McPartland, Vincenzo Di Marzo, Luciano De Petrocellis, Alison Mercer, Michelle Glass. Cannabinoid receptors are absent in insects. Journal of Comparative Neurology, 2001. [DOI | PubMed]
- Alix Morel, Pierre Lebard, Alexandra Dereux, Julien Azuar, Frank Questel, Frank Bellivier, Cynthia Marie-Claire, Mélina Fatséas, Florence Vorspan, Vanessa Bloch. Clinical Trials of Cannabidiol for Substance Use Disorders: Outcome Measures, Surrogate Endpoints, and Biomarkers. Frontiers in Psychiatry, 2021. [DOI]
- Annie Park, Alfredo Ghezzi, Thilini P. Wijesekera, Nigel S. Atkinson. Genetics and genomics of alcohol responses in Drosophila. Neuropharmacology, 2017. [DOI | PubMed]
- Matthew J. Pava, John J. Woodward. A review of the interactions between alcohol and the endocannabinoid system: Implications for alcohol dependence and future directions for research. Alcohol, 2012. [DOI | PubMed]
- Emma Puighermanal, Marta Luna-Sánchez, Alejandro Gella, Gunter van der Walt, Andrea Urpi, María Royo, Paula Tena-Morraja, Isabella Appiah, Maria Helena de Donato, Fabien Menardy, Patrizia Bianchi, Anna Esteve-Codina, Laura Rodríguez-Pascau, Cristina Vergara, Mercè Gómez-Pallarès, Giovanni Marsicano, Luigi Bellocchio, Marc Martinell, Elisenda Sanz, Sandra Jurado, Francesc Xavier Soriano, Pilar Pizcueta, Albert Quintana. Cannabidiol ameliorates mitochondrial disease via PPARγ activation in preclinical models. Nature Communications, 2024. [DOI]
- Evan C. Rosenberg, Simon Chamberland, Michael Bazelot, Erica R. Nebet, Xiaohan Wang, Sam McKenzie, Swati Jain, Stuart Greenhill, Max Wilson, Nicole Marley, Alejandro Salah, Shanice Bailey, Pabitra Hriday Patra, Rebecca Rose, Nicolas Chenouard, Simón(e) D. Sun, Drew Jones, György Buzsáki, Orrin Devinsky, Gavin Woodhall, Helen E. Scharfman, Benjamin J. Whalley, Richard W. Tsien. Cannabidiol modulates excitatory-inhibitory ratio to counter hippocampal hyperactivity. Neuron, 2023. [DOI | PubMed]
- Simran Sandhu, Arnavaz P. Kollah, Lara Lewellyn, Robin F. Chan, Mike Grotewiel. An Inexpensive, Scalable Behavioral Assay for Measuring Ethanol Sedation Sensitivity and Rapid Tolerance in <em>Drosophila</em>. Journal of Visualized Experiments, 2015. [DOI]
- Henrike Scholz, Jennifer Ramond, Carol M. Singh, Ulrike Heberlein. Functional Ethanol Tolerance in Drosophila. Neuron, 2000. [DOI | PubMed]
- Fred Shahbazi, Victoria Grandi, Abhinandan Banerjee, John F. Trant. Cannabinoids and Cannabinoid Receptors: The Story so Far. iScience, 2020. [DOI | PubMed]
- Michał Szulc, Radosław Kujawski, Amanda Pacholak, Marta Poprawska, Kamila Czora-Poczwardowska, Bogna Geppert, Przemysław Ł. Mikołajczak. Cannabidiol as a Modulator of the Development of Alcohol Tolerance in Rats. Nutrients, 2023. [DOI | PubMed]
- Jasmine Turna, Sabrina K. Syan, Benicio N. Frey, Brian Rush, Mary Jean Costello, Mark Weiss, James MacKillop. Cannabidiol as a Novel Candidate Alcohol Use Disorder Pharmacotherapy: A Systematic Review. Alcoholism: Clinical and Experimental Research, 2019. [DOI | PubMed]
- Adrián Viudez‐Martínez, María S García‐Gutiérrez, Ana Isabel Fraguas‐Sánchez, Ana Isabel Torres‐Suárez, Jorge Manzanares. Effects of cannabidiol plus naltrexone on motivation and ethanol consumption. British Journal of Pharmacology, 2018. [DOI | PubMed]
- Victor Voicu, Felix-Mircea Brehar, Corneliu Toader, Razvan-Adrian Covache-Busuioc, Antonio Daniel Corlatescu, Andrei Bordeianu, Horia Petre Costin, Bogdan-Gabriel Bratu, Luca-Andrei Glavan, Alexandru Vlad Ciurea. Cannabinoids in Medicine: A Multifaceted Exploration of Types, Therapeutic Applications, and Emerging Opportunities in Neurodegenerative Diseases and Cancer Therapy. Biomolecules, 2023. [DOI | PubMed]