
Plant Responses to Heavy Metal Stresses: Mechanisms, Defense Strategies, and Nanoparticle-Assisted Remediation
Abstract
Heavy metal (HM) contamination threatens environmental sustainability, food safety, and agricultural productivity worldwide. HM toxicity adversely affects plant growth, reducing germination rates by 20–50%, impairing seedling establishment, and inhibiting shoot and root development by 30–60% in various crops. HM disrupts key physiological processes, including photosynthesis, stomatal regulation, membrane integrity, nutrient uptake, and enzymatic and nonenzymatic antioxidant activities. These disruptions largely result from oxidative stress, caused by the excessive accumulation of reactive oxygen species, which damage cellular components. To counteract HM toxicity, plants deploy a complex defense network involving antioxidant enzymes, metal chelation by phytochelatins and metallothioneins, vacuolar sequestration, and symbiotic interactions with arbuscular mycorrhizal fungi, which can retain 40–70% of metals in roots and reduce translocation to shoots. At the molecular level, MAPK (Mitogen-Activated Protein Kinase) signaling pathways, transcription factors (e.g., WRKY, MYB, bZIP, and NAC), and phytohormonal crosstalk regulate the expression of stress-responsive genes expression to enhance HM stress tolerance. Advances in nanotechnology offer promising strategies for the remediation of HM-contaminated soils and water sources (HM remediation); engineered and biogenic nanoparticles (e.g., ZnO, Fe3O4) improve metal immobilization, reduce bioavailability, and enhance plant growth by 15–35% under HM stresses, although excessive doses may induce phytotoxicity. Future applications of nanotechnology in HM remediation should consider nanoparticle transformation (e.g., dissolution and agglomeration) and environmentally relevant concentrations to ensure efficacy and minimize phytotoxicity. Integrating phytoremediation with nanoparticle-enabled strategies provides a sustainable approach for HM remediation. This review emphasizes the need for a multidisciplinary framework linking plant science, biotechnology, and nanoscience to advance HM remediation and safeguard agricultural productivity.
Article type: Review Article
Keywords: heavy metal stress, nanoparticles, nanoparticle-assisted remediation, oxidative stress, phytotoxicity, signaling pathways
Affiliations: Department of Agronomy, Gazipur Agricultural University, Gazipur 1706, Bangladesh; ayshabsmrau@gmail.com (A.S.J.); munny@gau.edu.bd (M.A.); Institute of Genomics for Crop Abiotic Stress Tolerance, Department of Plant and Soil Science, Texas Tech University, Lubbock, TX 79409, USA; tanik@ttu.edu (T.R.A.); huong.nguyen@ttu.edu (H.M.N.); Agronomy and Farming Systems Division, Bangladesh Sugarcrop Research Institute, Ishwardi 6620, Bangladesh; aponbsmrau11@bsri.gov.bd; Department of Genetics and Plant Breeding, Gazipur Agricultural University, Gazipur 1706, Bangladesh; ashraful4225@stu.gau.edu.bd; Department of Agroforestry, Bangladesh Agricultural University, Mymensingh 2202, Bangladesh; alrahat.bsmrau@gmail.com; Vietnam National University of Agriculture, Ha Noi 100000, Vietnam; ntthuong355@gmail.com
License: © 2025 by the authors. CC BY 4.0 Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Article links: DOI: 10.3390/plants14243834 | PubMed: 41470716 | PMC: PMC12736845
Relevance: Moderate: mentioned 3+ times in text
Full text: PDF (2.1 MB)
1. Introduction
Heavy metals (HMs) are dense metallic elements with densities exceeding 5 g cm−3 and relatively high atomic weights [ref. 1,ref. 2,ref. 3]. HMs are non-biodegradable and can persist in the environment for long periods, leading to prolonged biological half-lives [ref. 1,ref. 2,ref. 3]. Arsenic (As), cadmium (Cd), chromium (Cr), mercury (Hg), and lead (Pb) are common HMs found in soils, water, and wastewater, primarily introduced via natural processes, anthropogenic activities, and agricultural practices [ref. 4,ref. 5,ref. 6,ref. 7]. In the rhizosphere, HMs interact with root exudates, altering their solubility and bioavailability, which may impair nutrient uptake and induce mineral deficiencies in plants [ref. 8,ref. 9]. HMs disrupt physiological and biochemical processes by generating oxidative stress, impairing protein synthesis and cell division, and causing symptoms such as leaf chlorosis and growth inhibition [ref. 9,ref. 10,ref. 11,ref. 12,ref. 13,ref. 14,ref. 15,ref. 16,ref. 17,ref. 18,ref. 19]. Cellular components, including plasma membranes, mitochondria, and chloroplasts are particularly vulnerable to HM-induced toxicity [ref. 15,ref. 20]. To cope with HM stress, plants have evolved several adaptive mechanisms, such as regulating metal uptake, enhancing antioxidant defenses, chelating and compartmentalizing HMs, and activating stress-responsive signaling pathways [ref. 8,ref. 9,ref. 21,ref. 22]. Detoxification strategies include symbiotic associations with mycorrhizal fungi, immobilization of metal ions via root exudates or cell wall binding, active efflux of metals across membranes, chelation by metal-binding peptides such as phytochelatins and metallothioneins, and sequestration into vacuoles to prevent cytosolic toxicity [ref. 23,ref. 24,ref. 25]. Phytoremediation represents a sustainable strategy for mitigating HM contamination, utilizing the metal-accumulating capabilities of plants and their associated rhizosphere microorganisms [ref. 24,ref. 25]. Furthermore, the incorporation of nanotechnology into remediation strategies also shows potential to enhance HM removal efficiency. However, its effectiveness depends on the type of metal, soil characteristics, and nanoparticle properties [ref. 8,ref. 26,ref. 27]. To fully harness the potential of nanoparticle-assisted remediation, a comprehensive understanding of the nanoscale interactions and the associated cellular and molecular mechanisms is essential.
This review aims to explore the morphological, physiological, biochemical, and molecular responses of plants to HM stress, focusing on their underlying defense mechanisms and current strategies in phytoremediation and nanoparticle-assisted remediation. Furthermore, it highlights critical knowledge gaps and outlines future research directions for the effective management of HM contamination.
2. Roles of Heavy Metals in Plants
HM stress negatively affects plant growth and development [ref. 28,ref. 29]. Table 1 presents the roles as well as the adequate and toxic levels of heavy metals in plants. While some HMs serve as essential micronutrients at trace levels, their excessive accumulation in plant tissues causes toxicity [ref. 30,ref. 31,ref. 32]. The severity of HM toxicity depends on the specific metal species, its chemical form, concentration, and plant type [ref. 33,ref. 34]. Non-essential HMs, such as As (≥5 mg kg−1 dry weight, DW), Cd (≥5 mg kg−1 DW), Cr (≥5 mg kg−1 DW), Hg (≥1 mg kg−1 DW), and Pb (≥30 mg kg−1 DW) are toxic to plants (Table 1). For example, Cd accumulation at toxic levels disrupts water and nutrient uptake, induces oxidative stress, impairs root development, and ultimately reduces crop productivity and food safety [ref. 35,ref. 36]. Hg predominantly accumulates in roots, with possible translocation to shoots, where it interferes with photosynthesis, respiration, and overall cellular metabolism by generating reactive oxygen species (ROS) and damaging cellular membranes [ref. 37,ref. 38]. Cr inhibits seed germination by 20–50%, reduces amylase activity, alters sugar transport to embryos, and decreases photosynthetic efficiency through oxidative damage [ref. 39,ref. 40]. Pb interferes with root and shoot growth (30–60%) in various crops as well as chlorophyll synthesis, stomatal regulation, and photosystem II efficiency, thereby impairing CO2 assimilation [ref. 41,ref. 42]. As, particularly arsenite (As3+, AsO33–), is primarily taken up via aquaporin channels and binds to sulfhydryl groups in proteins, inhibiting respiration by targeting dehydrogenases, promoting ROS production, and causing intrachromosomal recombination. In contrast, arsenate (As5+, AsO43–) is taken up mainly through phosphate transporters and is largely reduced to As3+ in roots, resulting in limited translocation to shoots, whereas As3+ can translocate more readily depending on the plant species and metal detoxification mechanisms [ref. 43]. Furthermore, combined exposure to HMs can exacerbate leaf and stem wilting, in purple loosestrife (Lythrum salicaria) and maize (Zea mays) [ref. 44,ref. 45]. Essential micronutrients, including copper (Cu), iron (Fe), manganese (Mn), molybdenum (Mo), and zinc (Zn), are vital for optimal vegetative and reproductive development in plants. Both deficiencies and excesses of these elements can disrupt physiological and biochemical processes, resulting in impaired growth, reduced photosynthetic efficiency, nutrient imbalances, and decreased crop productivity [ref. 46,ref. 47]. For instance, Cu (5–30 mg kg−1 DW) and Zn (25–150 mg kg−1 DW) concentrations enhance enzyme activity, photosynthetic efficiency, and antioxidant defense systems, thereby supporting overall plant growth and productivity [ref. 46,ref. 47,ref. 48,ref. 49]. Additionally, Cu and Zn nanoparticles have been reported to promote seed germination, root elongation, and stress tolerance by modulating ROS homeostasis and enhancing nutrient uptake [ref. 46,ref. 47,ref. 48,ref. 49,ref. 50,ref. 51]. Excess Cu (>30 mg kg−1 DW) inhibits seed germination and sugar metabolism by suppressing α-amylase and invertase activities, resulting in growth retardation and leaf chlorosis [ref. 51]. Similarly, excessive Fe (>500 mg kg−1 DW) and Mn (>1000 mg kg−1 DW) promote ROS generation, leading to membrane, protein, and DNA damage, which manifests as bronzing, chlorosis, and inhibited plant growth [ref. 52,ref. 53]. Elevated Mo interferes with nitrogen fixation in legumes, resulting in stunted growth and floral abnormalities [ref. 54]. High Zn concentrations (>150 mg kg−1 DW) suppress enzymatic activities (α-amylase, protease, ribonuclease), impair the mobilization of food reserves and carbonic anhydrase activity, increase malondialdehyde (MDA) content and electrolyte leakage, and ultimately reduce plant biomass [ref. 55,ref. 56,ref. 57,ref. 58,ref. 59]. Mung bean (Vigna radiata) and tomato (Solanum lycopersicum) exhibit enhanced growth at cobalt (Co) 0.02–2 mg kg−1 DW levels, whereas concentrations of >10 mg kg−1 DW induce toxicity, causing stunted growth and physiological disturbances [ref. 49,ref. 50]. Figure 1 illustrates the varying impacts of HMs on plants at toxic concentrations.
Table 1: Roles and adequate/toxic levels of heavy metals in plants.
| Metals | Essentiality and Roles in Plants | Adequate/Beneficial Levels (mg kg−1 DW) | Toxic Levels (mg kg−1 DW) | References |
|---|---|---|---|---|
| Arsenic (As) | Non-essential toxic heavy metal | – | ≥5 | [ref. 60] |
| Cadmium (Cd) | – | ≥5 | [ref. 60,ref. 61] | |
| Chromium (Cr) | – | ≥5 | [ref. 62] | |
| Mercury (Hg) | – | ≥1 | [ref. 60,ref. 63] | |
| Lead (Pb) | – | ≥30 | [ref. 62,ref. 64] | |
| Copper (Cu) | Essential micronutrient | 5–30 | >30 | [ref. 65,ref. 66] |
| Iron (Fe) | 50–250 | >500 | [ref. 65] | |
| Manganese (Mn) | 20–500 | >1000 | [ref. 65] | |
| Molybdenum (Mo) | 0.1–5 | >10 | [ref. 65] | |
| Zinc (Zn) | 25–150 | >150 | [ref. 67,ref. 68] | |
| Nickel (Ni) | 0.1–10 | >50 | [ref. 69,ref. 70] | |
| Cobalt (Co) | Beneficial for nitrogen fixation in legumes | 0.02–2 | >10 | [ref. 65] |
Adequate/beneficial levels indicate concentrations typically required for normal plant growth. Toxic levels indicate concentrations at which plants exhibit growth inhibition, chlorosis, or other toxicity symptoms.
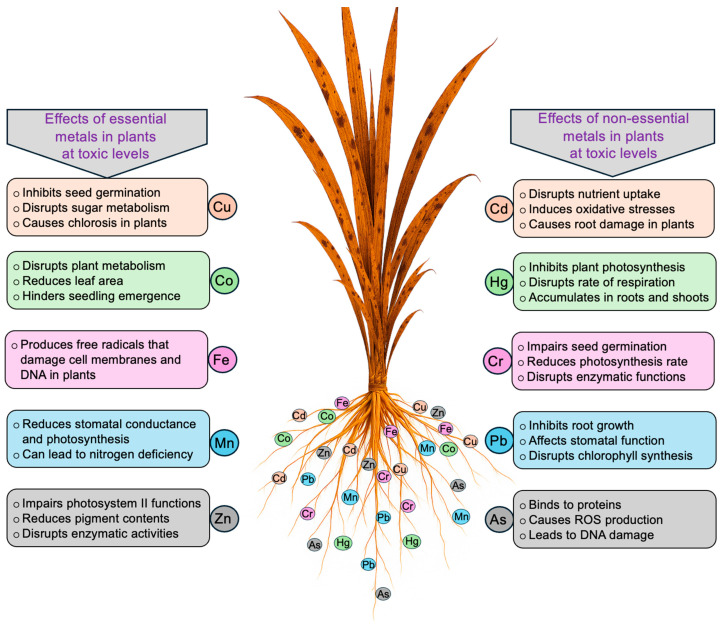
3. Morphological, Physiological, and Biochemical Responses to Heavy Metal Stresses
HM stress adversely affects morphological, physiological, and biochemical processes in plants. An overview of HM-induced changes in plant morphological, physiological, and biochemical processes is provided in Table 2. One of the primary effects of HM stress is a reduction in intra-leaf CO2 levels, which refers to the concentration of CO2 within the leaf mesophyll and directly affects photosynthetic carbon assimilation and leaf gas exchange characteristics [ref. 71,ref. 72]. HM stress adversely affects stomatal function through both direct accumulation in leaf tissues and the indirect disruption of key physiological processes, ultimately reducing the rates of photosynthesis and transpiration [ref. 72,ref. 73,ref. 74]. The regulation of ionic fluxes within guard cells plays a pivotal role in HM-induced stomatal dysfunction [ref. 73,ref. 74]. Since stomatal conductance directly influences CO2 uptake and water vapor loss, any alteration in stomatal morphology such as changes in size, density, or opening behavior can significantly impact leaf gas exchange, nutrient acquisition, and plant water balance [ref. 74]. For instance, Pb accumulation in soybean (Glycine max) guard cells disrupts cytoplasmic membrane integrity, resulting in potassium ion efflux, reduced guard cell turgor, and stomatal closure [ref. 75]. Additionally, Pb stress reduces transpiration in tobacco (Nicotiana tabacum) [ref. 76] and stomatal conductance in wheat (Triticum aestivum) and broadleaf plantain (Plantago major) [ref. 77,ref. 78]. Cd exposure induces stomatal closure, leading to decreased rates of photosynthesis, respiration, and water use efficiency, ultimately disrupting the plant’s energy [ref. 79,ref. 80]. In wheat, Cd stress results in lowered CO2 assimilation, reduced stomatal conductance, and diminished water use efficiency [ref. 81]. Cr stress significantly reduces stomatal conductance in maize (Zea mays), thereby limiting leaf gas exchange and negatively affecting photosynthetic activity [ref. 82]. Similarly, Zn stress reduces stomatal conductance in pigeon pea (Cajanus cajan) [ref. 83].
Table 2: Morphological, physiological, and biochemical responses of plants to heavy metal stresses.
| Plants | HM Stresses | Morphological, Physiological, and Biochemical Responses | References |
|---|---|---|---|
| Rice (Oryza sativa) | Cd | Reduces catalase activity, which impairs H2O2 scavenging, resulting in higher lipid peroxide levels | [ref. 84] |
| Ni | Increases the level of H2O2 and TBARS | [ref. 85] | |
| Pb | Increases lipid peroxidation | [ref. 86] | |
| Hg | Decreases canopy height, tillers number, panicle length, and yield | [ref. 87] | |
| Wheat (Triticum aestivum) | Ni | Increases electrolyte leakage and lipid peroxidation | [ref. 88] |
| Cd | Reduces shoot and root development | [ref. 89] | |
| Maize (Zea mays) | Zn and Ni | Intensifies lipid peroxidation and decrease in permeability of cell membranes | [ref. 90] |
| Sunflower (Helianthus annuus) | Cr | Increases lipid peroxidation by stimulating production of malondialdehyde and H2O2 | [ref. 91] |
| Ni | Inhibits mobilization of stored proteins and amino acids; reduces α-amylase and protease activity | [ref. 92] | |
| Lentil (Lens culinaris) | Cu | Increases lipid peroxidation in roots | [ref. 93] |
| Pigeon pea (Cajanus cajan) | Cd and Ni | Reduces photosynthetic activity | [ref. 94] |
| Siris tree (Albizia lebbeck) | Cd and Pb | Cd impairs seedling development and elongation; Pb disrupts stored food material and reduces germination rate | [ref. 95] |
| Indian mustard (Brassica juncea) | Cd | Reduces shoot and root biomass and decreases total chlorophyll content in the leaves | [ref. 96] |
| Silene compecta and Thalpsi ochrolucum | Cu | Damages the electron transport chain involved in photosynthesis | [ref. 97] |
| Thale cress (Arabidopsis thaliana) | Cd | Increases lipid peroxidation | [ref. 98] |
| Duckweed (Lemna minor) | Cu | [ref. 99] |
Cd, cadmium; Cr, chromium; Cu, copper; HM, heavy metal; Hg, mercury; H2O2, hydrogen peroxide; Ni, nickel; Pb, lead; TBARS, thiobarbituric acid reactive substances; Zn, zinc.
Plants counteract HM toxicity through the synthesis of phytochelatins (PCs) and metallothioneins (MTs), which chelate metals and reduce cytosolic toxicity. PCs preferentially bind Cd2+ and As3+, forming stable vacuolar complexes that limit cytosolic accumulation and oxidative damage, while MTs exhibit variable metal-binding affinities depending on the plant species. The specificity of PCs and MTs influences detoxification efficiency; for example, Cd2+ is rapidly chelated in roots, reducing shoot translocation, whereas As3+ complexes may be redistributed gradually. These mechanisms vary across species, with Arabidopsis (Arabidopsis thaliana) accumulating Cd–PC complexes mainly in roots, while rice (Oryza sativa) shows greater root-to-shoot Cd translocation. Differences in MT expression further modulate tolerance to multi-metal stress, highlighting species-specific strategies in HM detoxification. HM stress increases abscisic acid (ABA) synthesis, which activates the OST1 (Open Stomata 1) protein kinase while inhibiting protein phosphatase 2C (PP2C) activity [ref. 96,ref. 97]. Blocking PP2Cs prevents protein dephosphorylation, thus relieving the suppression of ABA signaling [ref. 98]. The Open Stomata 1 (OST1) represents one of the main targets of PP2Cs, with OST1 being the best-characterized member [ref. 100,ref. 101]. OST1 transmits the ABA signal by phosphorylating downstream targets and enhancing ROS production via NADPH oxidase activation [ref. 102]. Additionally, nitric oxide (NO) and reactive nitrogen species (RNS) act as secondary messengers, amplifying the signaling cascade under HM stress. As a redox-active free radical, NO can neutralize HM-induced ROS directly or by activating antioxidant defenses. However, an imbalance between NO, ROS, and antioxidant activity can trigger oxidative stress [ref. 103]. HM stress also disrupts the photosynthetic electron transport chain, leading to decreased Photosystem II activity. At elevated concentrations, HM stress disturbs the light-harvesting complex, particularly the transition between its functional states. This results in antenna complex disorganization and compromised photochemical efficiency [ref. 104]. HM stress disrupts glucose metabolism, tricarboxylic acid cycle, and photosynthesis, while inducing ABA, ROS, and NO production, leading to signaling cascades including PP2C, OST1, and 8-Nitro-cGMP that cause turgor loss and stomatal closure. These processes collectively reduce the rates of photosynthesis, transpiration, respiration, nutrient and water uptake, and leaf area, while causing ion imbalance and cell membrane damage. The impact of HM stress on the physiological functions of plants is illustrated in Figure 2.

4. Disruption of Cell Membrane Integrity, Oxidative Homeostasis, and Enzymatic Activities Under Heavy Metal Stresses
HMs can penetrate cell membranes and bind to essential components such as proteins and phospholipids [ref. 105]. HMs interfere with membrane integrity, disrupt transport processes, displace calcium ions from critical binding sites, and reduce the availability of ATPase substrates by binding to adenosine triphosphate (ATP) [ref. 106,ref. 107]. HM stress strongly promotes the formation of hydroxyl free radicals, triggering lipid peroxidation through primary initiation as well as via the Haber–Weiss and Fenton reactions [ref. 108]. HM stress also leads to the accumulation of methylglyoxal in plants, which impairs antioxidant defense by depleting glutathione levels [ref. 109,ref. 110,ref. 111]. Redox-active metals such as Cu, Fe, and Cr can directly generate ROS [ref. 112], while redox-inactive metals such as Cd, Pb, and Zn promote ROS accumulation indirectly by impairing the plant’s antioxidant defense system, triggering calcium-dependent signaling pathways, and disturbing iron metabolism [ref. 111,ref. 112,ref. 113,ref. 114,ref. 115,ref. 116]. On the other hand, Cd, Pb, Ni, and Cr can substitute for vital cofactors like Mg, Zn, Fe, and Ca in enzymes such as RuBisCO and superoxide dismutases, resulting in impaired enzymatic activity and disrupted cellular signaling [ref. 117,ref. 118]. For example, Cd disrupts cellular functions by displacing Zn in zinc-finger transcription factors and Ca in calmodulin, thereby altering gene expression and impairing stomatal regulation, whereas Pb interferes with chlorophyll structure and photosystem II activity, ultimately reducing photosynthetic efficiency [ref. 119,ref. 120]. Additionally, arsenate (AsO43–) stress can mimic phosphate and disrupt ATP synthesis [ref. 121]. It also binds strongly to protein sulfhydryl (–SH) and carboxyl (–COOH) groups, leading to enzyme denaturation [ref. 105,ref. 122,ref. 123,ref. 124,ref. 125,ref. 126,ref. 127].
5. Plants’ Resistance Mechanisms to Heavy Metal Stresses
Plants cope with HM stress through two main resistance strategies: avoidance and tolerance. Avoidance mechanisms, such as mycorrhizal immobilization, root exudate complexation, and rhizosphere pH adjustment, function to prevent metal uptake and entry into root tissues. Among these avoidance strategies, immobilization by mycorrhizal associations is particularly effective for its role in restricting metal translocation and mitigating toxicity. When metals do enter plant cells, tolerance mechanisms are triggered, including the enhancement of antioxidant defenses, production of stress-associated proteins, and gene regulation to detoxify and compartmentalize the metals into less harmful forms in plants.
5.1. Immobilization by Mycorrhizal Associations
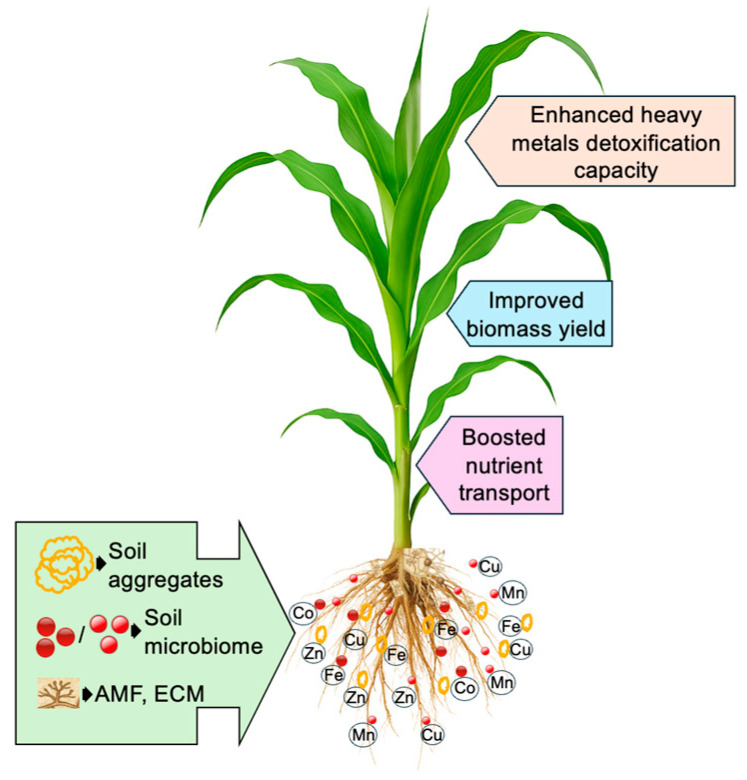
Mycorrhizal fungi, particularly arbuscular mycorrhizal fungi (AMF) and ectomycorrhizal fungi (ECM), play a vital role in enhancing plant tolerance to HM stress by forming symbiotic associations with roots [ref. 128,ref. 129]. These fungi reduce HM uptake through chelation, adsorption, and the release of organic acids and glomalin [ref. 130,ref. 131]. Extraradical hyphae of AMF extend beyond the root zone, improving nutrients such as N, P, and K uptake, which in turn enhances plant growth and HM stress resistance [ref. 132,ref. 133]. AMF can retain 40–70% of metals in root tissues via chelation, while extraradical and intraradical hyphae provide additional binding surfaces containing glucan, chitin, and galactosamine polymers [ref. 134]. They also release organic acids such as citric, malic, and oxalic acids that immobilize HMs by forming stable complexes [ref. 131]. A major contribution of AMF is the secretion of glomalin, a glycoprotein with high metal-binding affinity, which stabilizes soil aggregates and reduces HM bioavailability. Evidence shows that AMF inoculation promotes HM retention in roots and reduces their movement to aerial tissues, often resulting in significantly lower concentrations in edible parts [ref. 135]. For example, Adeyemi et al. [ref. 134] reported that AMF inoculation enhanced P uptake and restricted HM transport to shoots in soybean (Glycine max), while eggplants (Solanum melongena) inoculated with AMF accumulated less Pb in fruits, improving growth and food safety. Although most studies confirm reduced translocation, small amounts of metals may still reach aboveground tissues at lower concentrations, and the precise mechanisms controlling long-distance movement remain only partly understood. In addition to AMF, ECM form a fungal sheath around roots and secrete extracellular polymers and organic acids that immobilize metals in the rhizosphere. ECM are particularly important for woody plants and act primarily as barriers to HM entry, whereas AMF combine nutrient acquisition with HM detoxification. Together, these two groups of fungi represent complementary strategies for enhancing plant tolerance to metal-contaminated soils. Through these mechanisms, mycorrhizal associations enhance nutrient transport, detoxify HMs, and improve biomass accumulation, especially during active phases of plant growth. They also block pathogen invasion, enrich beneficial rhizosphere microbes, and modulate phytohormone levels (e.g., IAA, cytokinins), further supporting root health and plant vigor [ref. 136,ref. 137,ref. 138,ref. 139]. Figure 3 illustrates the enhancement of HM detoxification, nutrient transport, and biomass accumulation in plants through microbial functions. Thus, mycorrhizal symbiosis functions as a critical plant defense strategy in HM-contaminated environments, providing both direct detoxification and indirect growth benefits. Nonetheless, knowledge gaps remain regarding the regulation of HM translocation from roots to shoots, warranting further research to fully elucidate these processes.
5.2. Root-Mediated Mechanisms and Phytochelatin-Driven Detoxification Under Heavy Metal Stresses
Among the diverse strategies employed by plants to withstand HM toxicity, root-mediated mechanisms and PC-driven detoxification play pivotal roles. Cellular mechanisms of HM detoxification and sequestration in plants are illustrated in Figure 4. Roots serve as the primary site for HM perception and uptake. One of the key defense strategies involves the secretion of root exudates, which include diffusates (e.g., peptides, carboxylic acids, and carbohydrates), excretions (e.g., protons, bicarbonates, and CO2), and secretions (e.g., mucilage, siderophores, and allelochemicals) [ref. 140]. These compounds contribute to the formation of stable metal complexes in the rhizosphere, thereby reducing HM solubility, mobility, and toxicity near the root surface [ref. 141]. Organic acids chelate metal ions and reduce their toxicity. Additionally, amino acids like histidine and nicotianamine act as metal chelators, enhancing the solubility and translocation of micronutrients such as Fe and Zn. Root-derived sugars and amino acids can also increase rhizosphere pH and modify redox conditions, encouraging metal precipitation and improving nutrient availability [ref. 142]. PCs and MTs play critical roles in chelating metal ions and mediating their compartmentalization particularly into vacuoles thereby minimizing cytosolic toxicity [ref. 143,ref. 144,ref. 145,ref. 146]. PCs can form stable complexes with a wide range of metal ions including Cd2+, As3+, Pb2+, and Zn2+ [ref. 147]. Their synthesis is rapidly upregulated following HM exposure, utilizing cytosolic glutathione present in millimolar concentrations as a substrate [ref. 148]. Once formed, PC-metal complexes are actively transported into the vacuole via tonoplast-localized ATP-Binding cassette transporters or metal/H+ antiporters, effectively sequestering toxic metals from the cytosol and protecting sensitive enzymatic processes [ref. 145]. Inside the vacuole, these complexes may undergo further stabilization through interactions with inorganic sulfide or sulfite ions, which shield them from proteolytic degradation. Over time, this compartmentalization may result in the formation of insoluble crystalline metal deposits, contributing to long-term immobilization and detoxification of HMs [ref. 148]. In addition to their detoxification function, PCs are also involved in metal homeostasis, particularly in regulating the uptake and distribution of essential micronutrients such as Zn and Cu [ref. 149,ref. 150]. In parallel, MTs serve as metal-binding ligands that assist in detoxifying excess metal ions while also regulating the availability of essential elements such as Zn and Cu. In plants, MTs are categorized into subfamilies (e.g., MT1 and MT2) based on the arrangement of their cysteine domains, which determine their metal-binding specificity and biological roles [ref. 151]. In addition to PC- and MT-mediated chelation, plants utilize several cellular barriers and strategies to limit HMs entry and toxicity:
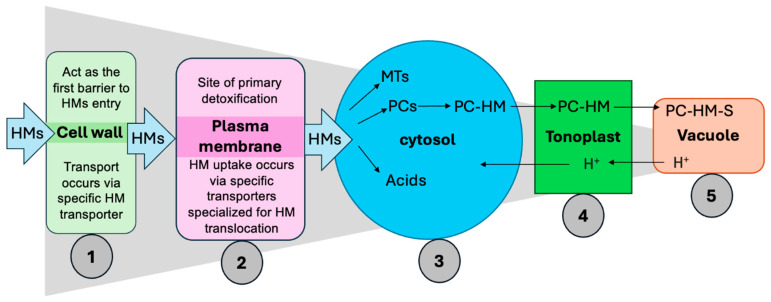
- Plant’s cell wall, rich in pectin and lignin, can adsorb HMs through ion exchange, thereby reducing metal mobility and entry into cells [ref. 152].
- Specific membrane transporters and channel proteins regulate HM uptake. Under HM stress, plants often downregulate these transporters to reduce metal influx [ref. 118].
- Chelation is a widespread detoxification mechanism involving intracellular ligands such as PCs, MTs, organic acids (e.g., citric, malic, oxalic acids), and amino acids (e.g., histidine, nicotianamine), which bind free metal ions and prevent them from interfering with vital metabolic processes [ref. 153].
- The tonoplast (vacuolar membrane) minimizes HM movement back into the cytoplasm via active permeability mechanisms, serving as a dynamic barrier against metal recirculation [ref. 147].
- Once PC-metal complexes are formed, they are transported into vacuoles and ultimately converted into insoluble metal deposits, stabilizing the intracellular environment and preventing oxidative damage [ref. 149,ref. 153].
5.3. Antioxidant Responses to Heavy Metal Stresses
Antioxidant enzymes play a central role in mitigating oxidative damage in plants exposed to HM stresses [ref. 112,ref. 113,ref. 114]. HM exposure induces the production of heat shock proteins (HSPs), which act as molecular chaperones, helping stabilize proteins and preserve membrane integrity. Studies show that various HSPs, including low-molecular-weight forms and HSP70, are upregulated in different plant species under HM stress [ref. 154,ref. 155,ref. 156,ref. 157]. Cd, Pb, Cr, and Hg induce overproduction of ROS, including superoxide radicals, hydrogen peroxide, and hydroxyl radicals, which disrupt cellular homeostasis [ref. 158,ref. 159]. In response, plants activate enzymatic defense mechanisms involving superoxide dismutase (SOD), catalase (CAT), peroxidases (POD), ascorbate peroxidase (APX), and glutathione reductase (GR) [ref. 160,ref. 161,ref. 162,ref. 163,ref. 164]. However, the intensity and efficiency of these responses vary with plant species, developmental stage, metal type, and exposure level. An overview of plant antioxidant enzyme responses to HM stress is provided in Table 3. In addition to enzymatic antioxidants, plants also accumulate secondary metabolites which further support ROS detoxification and improve stress resilience [ref. 161,ref. 165,ref. 166,ref. 167,ref. 168]. These metabolites help mitigate HM toxicity by chelating metals in the rhizosphere and scavenging ROS within the cells.
Table 3: Responses of antioxidant enzymes in plants exposed to heavy metal stresses.
| Plants | HM | Antioxidant Enzymes Response | References |
|---|---|---|---|
| Maize (Zea mays) | Cd | Increases APX and GPX activities | [ref. 162] |
| Zn | SOD and POD activities increase, while CAT activity decreases at higher Zn levels | [ref. 163] | |
| Barley (Hordeum vulgare) | Cd | Increases activities of APX and GPX | [ref. 164] |
| Rice (Oryza sativa) | Pb | Elevates guaiacol peroxidase, SOD, and GR activities | [ref. 86] |
| Mung bean (Vigna radiata) | Cr | APX activity increases, which helps reduce H2O2 accumulation | [ref. 169] |
| Tomato (Lycopersicon esculentum) | Cu | Increases the activities of SOD, POD, and CAT | [ref. 170] |
| Indian mustard (Brassica juncea) | Zn | Increases CAT activity, which scavenges H2O2 and reduces oxidative stress | [ref. 171] |
| Okra (Abelmoschus esculentus) | Hg | Increases SOD, APX, and GR activities and decreases CAT activity | [ref. 172] |
| Peregrina (Jatropha integerrima) | Zn | POD and CAT activities increase with Zn concentration | [ref. 173] |
| Coffee (Coffea arabica) | Increases GR activity, which supports GSH levels for PC biosynthesis | [ref. 174] | |
| Water hyacinth (Eichhornia crassipes) | CAT activity increases with Ag, Cd, Cr, Pb, and Cu | [ref. 175] | |
| Camelthorn (Alhagi camelorum) | Cu | Induces PC synthesis and depletes total GSH activity | [ref. 176] |
Ag, silver; APX, ascorbate peroxidase; CAT, catalase; Cd, cadmium; Cr, chromium; Cu, copper; GPX, glutathione peroxidase; GR, glutathione reductase; GSH, reduced glutathione; HM, heavy metal; Hg, mercury; H2O2, hydrogen peroxide; PC, phytochelatins; Pb, lead; POD, peroxidase; SOD, superoxide dismutase; Zn, zinc.
5.4. Regulation of Signaling Pathways and Gene Expression in Plants Under Heavy Metal Stresses
HM exposure triggers complex signaling pathways in plants that regulate gene expression and activate defense mechanisms. A major downstream signaling route is the mitogen-activated protein kinase cascade, which integrates signals from ROS, nitric oxide, and phytohormones to regulate stress-responsive genes [ref. 177]. These coordinated responses help initiate detoxification and stress adaptation processes. Crosstalk among signaling pathways ensures fine-tuned regulation, enabling plants to maintain metal homeostasis and resist toxicity [ref. 178]. Key signaling pathways and associated molecular players implicated in plant adaptation to HM stress are summarized in Table 4. Understanding these mechanisms is essential for advancing crop resilience to HM-contaminated environments.
Table 4: Key signaling pathways and molecular components involved in heavy metal stress responses in plants.
| Signaling Pathways | Key Components | Heavy Metals | Responses | Genes Involved | References |
|---|---|---|---|---|---|
| Calcium-dependent signaling | Ca2+ channels, calmodulins (CaMs), calmodulin-like proteins (CMLs), calcium-dependent protein kinases (CDPKs), calcineurin B-like proteins (CBLs)/CBL-interacting protein kinases (CIPKs) | Cr, As, Pb, Cu | Calcium influx triggers antioxidant enzyme activation (e.g., SOD, APX); regulates redox homeostasis; CDPKs and CaMs modulate downstream responses | AtCBL1, CDPK-like kinases, CAMs | [ref. 178,ref. 179,ref. 180] |
| MAPK cascade signaling | MAPKKK → MAPKK → MAPK (MPKs) | Cd, Cu, As, Cr | Phosphorylation of TFs (WRKY, DREB, bZIP, MYB); modulation of stress-responsive genes; interaction with HSPs for defense | OsMAPK2, ZmMPK3/6, WRKY, ERF, bZIP, MYB | [ref. 177,ref. 181,ref. 182] |
| ROS signaling | ROS (O2–, H2O2, OH−), antioxidant enzymes (SOD, CAT, APX), thiol metabolism enzymes | Cd, Cr, As, Pb | Low ROS levels act as signaling molecules; high ROS induce PCD; upregulation of antioxidant genes maintains ROS balance | OsGSTL2, OsMATE1/2, DHAR, GR, SOD, CAT | [ref. 178,ref. 183] |
| Hormonal Signaling | ABA, JA, ET, SA, EIN2/3, JAZ, AP2/ERF transcription factors | Cd, Cr, As | Phytohormones regulate transcription and crosstalk with MAPK cascades; influence root development and HM detoxification | AP2/ERF, ACS, OsARM1, AtMYB, AB15, TGAL3 | [ref. 180,ref. 184] |
| Crosstalk and integration | Interactions among Ca2+, ROS, MAPKs, hormones, nitric oxide | Cd, Pb, As, Cr, Cu | Synergistic and antagonistic interactions among signaling pathways coordinate stress responses; modulate TF networks | Multigene families: WRKY, bZIP, HSF, MYB, ERF | [ref. 177,ref. 178] |
ABA, abscisic acid; AP2/ERF, APETALA2/ethylene-responsive factor; APX, ascorbate peroxidase; As, arsenic; Ca2+, calcium ion; CAT, catalase; Cd, cadmium; CBL, calcineurin B-like protein; CDPK, calcium-dependent protein kinase; CIPK, CBL-interacting protein kinase; Cr, chromium; Cu, copper; DHAR, dehydroascorbate reductase; ET, ethylene; GR, glutathione reductase; H2O2, hydrogen peroxide; HSP, heat shock protein; JA, jasmonic acid; MAPK, mitogen-activated protein kinase; MAPKK, MAPK kinase; MAPKKK, MAPK kinase kinase; OH−, hydroxyl radical; O2−, superoxide anion; PCD, programmed cell death; ROS, reactive oxygen species; SA, salicylic acid; SOD, superoxide dismutase; TF, transcription factor; Zn, zinc.
Transcription factors (TFs) play a pivotal role in regulating plant responses to HM stress by modulating the expression of stress-responsive genes [ref. 185,ref. 186,ref. 187]. An overview of transcription factors activated during HM stress in plants is provided in Table 5. TFs act as molecular switches that activate or repress target genes involved in metal uptake, transport, sequestration, and detoxification. Key TF families, such as bZIP (basic leucine zipper transcription factor), MYB (myeloblastosis transcription factor), WRKY (tryptophan–arginine–lysine–tyrosine transcription factor), NAC, and AP2/ERF, coordinate a wide range of physiological and biochemical processes, including antioxidant defense, synthesis of metal-binding ligands (e.g., PCs and MTs), and regulation of ion transporters [ref. 187,ref. 188,ref. 189,ref. 190,ref. 191,ref. 192,ref. 193]. Mechanistically, WRKY TFs are frequently upregulated under Cd stress, where they activate downstream genes encoding antioxidant enzymes and phytochelatin synthases, thereby enhancing detoxification capacity. MYB TFs preferentially respond to Pb stress, regulating secondary metabolism and ion transport to limit Pb translocation to shoots [ref. 182,ref. 194,ref. 195]. bZIP TFs are strongly linked to redox homeostasis, integrating ROS signaling with metal-induced oxidative stress responses, while NAC and AP2/ERF TFs coordinate hormonal signaling pathways (e.g., ABA, ethylene) that fine-tune stress-responsive gene expression [ref. 182,ref. 195]. Under combined metal exposure (e.g., Cd + Pb), crosstalk among these TFs integrates overlapping yet distinct signaling networks, synchronizing antioxidant defenses, metal chelation, and ion homeostasis [ref. 194,ref. 195,ref. 196,ref. 197,ref. 198,ref. 199]. This integrated regulation highlights how plants orchestrate both metal-specific and multi-metal tolerance strategies, providing mechanistic insights valuable for engineering crops with enhanced resilience to HM stress.
Table 5: Roles of transcription factors in plant adaptation to heavy metal stresses.
| Plants | TFs | Gene(s) | Key Findings | References |
|---|---|---|---|---|
| Wheat (Triticum aestivum) | HSF | TaHsfA4a | Upregulates metallothionein genes under Cd stress | [ref. 194] |
| Rice (Oryza sativa) | MYB | OsMYB45 | Downregulation increases Cd sensitivity; regulates antioxidant activity | [ref. 182,ref. 195] |
| bZIP | – | Involved in auxin and HM signaling crosstalk | ||
| WRKY | – | Activated by MAPK pathways under HM stress | ||
| Sorghum (Sorghum bicolor) | MYB | SbMYB15 | Confers Cd and Ni stress tolerance | [ref. 196] |
| Walnut (Juglans regia) | MYB | JrMYB2 | Improves tolerance to Cd stress | [ref. 197] |
| Tomato (Solanum lycopersicum) | HSF | HSF1A | Induces melatonin biosynthesis for Cd tolerance | [ref. 198] |
| Rapeseed (Brassica napus) | bZIP | BnbZIP2, BnbZIP3 | Upregulated under drought and Cd; involved in stress signaling | [ref. 199] |
| Arabidopsis (Arabidopsis thaliana) | MYB | AtMYB4 | Improves antioxidant defense under Cd stress | [ref. 200] |
| WRKY | AtWRKY12 | Downregulated under Cd; represses GSH1 to negatively regulate Cd tolerance | [ref. 201] | |
| WRKY | AtWRKY13 | Upregulated under Cd; activates PDR8 to positively regulate Cd tolerance | [ref. 202] | |
| WRKY | WRKY33 | Regulates HM uptake via IRT1 regulation under Cd stress | [ref. 203] | |
| bZIP | AB15 | Interacts with MYB49 to reduce Cd uptake via IRT1 inactivation | [ref. 204] |
Cd, cadmium; Ni, nickel; GSH1, glutathione synthetase 1; HM, heavy metal; PDR8, pleiotropic drug resistance 8; IRT1, iron-regulated transporter 1; MAPK, mitogen-activated protein kinase; HSF, heat shock transcription factor; MYB, myeloblastosis transcription factor; TFs, transcription factors; WRKY, tryptophan-arginine-lysine-tyrosine transcription factor; bZIP, basic leucine zipper transcription factor.
Transgenic approaches have proven effective in improving plant tolerance HM stress. Overexpression of genes such as phytochelatin synthase or metal transporters (HMA3, Nramp) enhances HM sequestration and limits translocation to shoots, reducing cytosolic toxicity [ref. 185,ref. 186,ref. 187,ref. 188,ref. 189,ref. 190,ref. 191]. Antioxidant genes further mitigate oxidative damage and preserve photosynthetic efficiency [ref. 158,ref. 159,ref. 160,ref. 165,ref. 166,ref. 167,ref. 168]. Combining these genetic strategies with nanoparticle-assisted remediation offers synergistic benefits: nanoparticles immobilize HMs in the rhizosphere, while transgenic plants efficiently chelate and compartmentalize metals intracellularly. This integrated approach enhances plant growth, stress tolerance, and HM removal efficiency, highlighting the potential of biotechnology in conjunction with nanotechnology for HM-contaminated soils.
CRISPR/Cas9 genome editing offers a precise approach to improve plant tolerance to HM stress. By targeting key genes, transcription factors (e.g., WRKY, MYB, bZIP), and metal transporters, CRISPR/Cas9 can reduce metal uptake and translocation, enhance antioxidant defenses, and strengthen PC or MT-mediated detoxification [ref. 182,ref. 194]. Studies in rice, Arabidopsis, and wheat have shown that such modifications can improve growth, stress tolerance, and yield under HM exposure, providing an efficient strategy to develop HM-resilient crops.
6. Nanoparticle-Mediated Alleviation of Heavy Metal Stresses in Plants
Nanoparticles (NPs) have demonstrated significant potential in alleviating the adverse effects of HM stress in plants [ref. 205]. At low concentrations, NPs can stimulate plant growth, enhance stress resistance, and improve photosynthetic efficiency. However, excessive exposure may cause phytotoxicity, including oxidative damage, growth inhibition, and impaired nutrient uptake [ref. 206]. Understanding the dose-dependent behavior of NPs is therefore critical for their safe and effective application. NPs can affect HM root-to-shoot translocation by immobilizing metals in the rhizosphere or binding them within root tissues, thus reducing their translocation to shoots [ref. 207,ref. 208]. For example, Fe3O4 NPs reduce Cd mobility in wheat roots and shoots, while mercapto-functionalized silica nanoparticles (SiNPs) stabilize Cd in less bioavailable forms [ref. 209,ref. 210]. ZnO NPs enhance antioxidant defenses in white leadtree (Leucaena leucocephala), mitigating Cd and Pb toxicity, and can interact with endogenous phytochelatins and metallothioneins to form stable metal-ligand complexes, further reducing cytosolic toxicity [ref. 211,ref. 212,ref. 213]. These interactions are often concentration-dependent, with optimal NP levels promoting growth and metal detoxification, whereas excessive doses can disrupt cellular redox homeostasis and metal chelation, highlighting the need to determine safe application thresholds. Beyond plant-level effects, the environmental fate of nanoparticles must be considered. Uncontrolled or excessive NP application may affect soil microbial communities, bioaccumulate in non-target organisms, or contaminate soil and water ecosystems. Therefore, careful monitoring, dose optimization, and ecological risk assessment are essential for the sustainable use of nanoparticles in HM remediation. Table 6 presents different nanoparticles used to mitigate HM stress in plants, highlighting their roles in enhancing tolerance and reducing toxicity.
Table 6: Nanoparticles for heavy metal stress mitigation in plants.
| Nanoparticles | Plant Species | HMs | Reduction of HMs (%) | Key Findings | References |
|---|---|---|---|---|---|
| Zinc oxide (ZnO) | Rice, fenugreek, and Leucaena leucocephala | Pb, Cd, Cr, Cu | Pb: 79–85; Cd: 80–87; Cr: 38–81; Cu: 60 | Improves growth and Zn uptake; reduces HM accumulation | [ref. 213,ref. 214] |
| Cerium oxide (CeO2) | Rice | Cd | 8.4 | Reduces growth inhibition and oxidative stress | [ref. 152] |
| Astaxanthin-functionalized gold (Ast-Au) NPs | 26–86 | Enhances chlorophyll content and amino acid metabolism; scavenges ROS | [ref. 215] | ||
| Titanium dioxide (TiO2) | Rice and cucumber | Pb, As, Al | 34–97 | Reduces HM contamination and toxicity | [ref. 216] |
| Iron oxide (Fe3O4) | Wheat | Pb, Zn, Cd, Cu | Roots: 24–68; Shoots: 11–100 | Reduces oxidative stress and growth suppression | [ref. 217,ref. 218] |
| Selenium NPs (Se, Bio-Se) | Coriander and pak choi | Cd, Pb, Hg | Cd: 21–31; Pb: 5–30; Hg: 3–23 | Enhances antioxidant defense; reduces HM uptake | [ref. 219,ref. 220] |
| Graphene oxide | Lettuce | Cd | – | Reduces Cd toxicity; improves photosynthesis, chlorophyll content, antioxidant enzymes, and biomass | [ref. 221] |
Al, aluminum; As, arsenic; Bio-Se, biologically synthesized selenium; Cd, cadmium; Cr, chromium; Cu, copper; Fe, iron; HM, heavy metal; Hg, mercury; NPs, nanoparticles; Pb, lead; ROS, reactive oxygen species; Zn, zinc.
7. Nanoparticle-Assisted Bioremediation Against Heavy Metal Stresses
Nanoparticle-assisted bioremediation represents a promising and eco-friendly strategy that merges nanotechnology with biological approaches, particularly the use of microorganisms to detoxify HMs stress [ref. 222,ref. 223]. For example, bacteria such as Pseudomonas aeruginosa, Bacillus subtilis, Rhizobium spp., and Staphylococcus aureus can biosynthesize NPs with high adsorption capacity, enabling effective HM immobilization [ref. 224]. Fungal genera like Aspergillus, Rhizopus, and Penicillium contribute to HM remediation [ref. 225]. Plant-derived NPs, synthesized using extracts from Camellia sinensis, Citrus limon, and Mangifera indica, benefit from stabilization by secondary metabolites, enhancing their biocompatibility and remediation potential [ref. 226]. Polyvinylpyrrolidone (PVP)-coated iron oxide NPs combined with Halomonas sp. achieved 100% removal of Cd2+ and 80% removal of Pb2+ within 24 h, far outperforming NPs used alone [ref. 227]. Biogenic manganese oxide (MnO) NPs synthesized by bacteria and fungi have demonstrated excellent cation exchange and redox capacity, enabling effective reduction of Cd, Cu, Pb, and Zn in contaminated soils [ref. 228,ref. 229]. Remarkably, biogenic MnO from Pseudomonas putida MnB1 exhibits 7–8 times greater HM adsorption across diverse environmental conditions. However, the potential ecological risks of nanoparticles should not be overlooked. Excessive or uncontrolled NP application may lead to phytotoxicity, disruption of soil microbial communities, bioaccumulation in non-target organisms, and contamination of water and soil ecosystems. Therefore, careful dose optimization, environmental risk assessment, and monitoring are essential for the safe and sustainable implementation of nanoparticle-assisted remediation strategies. Table 7 summarizes the microbial isolates used for NP biosynthesis and their applications in mitigating HM stress in plants.
Table 7: Microbial and plant-based biosynthesis of nanoparticles and their role in mitigating heavy metal phytotoxicity.
| Nanoparticles | Microorganisms/Plant Extracts Used | Alleviation of Phytotoxicity/Key Findings | References |
|---|---|---|---|
| Silver NPs | Escherichia coli | Rapid reduction of Ag+ ions within minutes | [ref. 230] |
| Pseudomonas stutzeri | Silver-resistant; accumulates silver and reduces its toxicity | [ref. 231] | |
| Solanum xanthocarpum (berry extract) | Enhances Ag+ ion reduction rate via phytochemicals | [ref. 232] | |
| Convolvulus arvensis (leaf extract) | Achieves 98.99% Cu2+ ion removal via adsorption | [ref. 233] | |
| Gold NPs | Bacillus subtilis | Acts as a biocontrol agent with antifungal properties | [ref. 234] |
| Aspergillus japonicus | Reduces Au(III) to Au(0); immobilized AuNPs | [ref. 235] | |
| Colletotrichum sp. | Reduces and caps gold NPs | [ref. 236] | |
| ZnO NPs | Green algae | Converts metal ions into zero-valent metals via phytochemicals | [ref. 237] |
| Citrus limon (leaf extract) | Non-toxic synthesis; biomolecule-rich extract enhances safety | [ref. 238] | |
| Lead NPs | Clostridium pasteurianum | Reduces Cr(VI) to Cr(III); ~70% remediation efficiency | [ref. 239] |
| Iron NPs | Chlorococcum (alga) | Biosynthesized Fe NPs removed 92% Cr vs. 25% by bulk Fe | [ref. 240] |
| Aspergillus oryzae | Cost-effective and eco-friendly NP synthesis for remediation | [ref. 241] | |
| TiO2 NPs | Aspergillus niger | Reduces Cr(VI) toxicity and DNA damage in Helianthus annuus by minimizing total Cr uptake | [ref. 242] |
| Biogenic Fe–Mn oxides (BFMO) | Pseudomonas sp. | Converts As(III) to less mobile As(V); enhances arsenic remediation | [ref. 243] |
| CdS NPs | P. aeruginosa | EPS-enriched CdS NPs enhance cadmium ion adsorption and stabilization | [ref. 244] |
Ag+, silver ions; As, arsenic; As(III), arsenite; As(V), arsenate; AuNPs, gold nanoparticles; CdS, cadmium sulfide; Cr, chromium; Cr(III), trivalent chromium; Cr(VI), hexavalent chromium; EPS, extracellular polymeric substances; Fe–Mn, iron–manganese; NPs, nanoparticles; TiO2, titanium dioxide.
8. Conclusions and Future Research Directions
Growing prevalence of HM contamination in agricultural ecosystems poses a significant threat to ecosystem health and the sustainability of food production systems. HM stress disrupts essential plant functions by restricting shoot and root development, delaying seedling establishment, while impairing key physiological and metabolic processes including photosynthesis, respiration, water and nutrient uptake, enzymatic activities, and hormonal signaling pathways. These disruptions lead to the excessive generation of ROS, resulting in oxidative stress that causes cellular damage, growth inhibition, and substantial reductions in crop yield and quality. To counteract HM toxicity, plants have evolved a complex network of defense mechanisms operating at physiological, biochemical, and molecular levels. Physiological strategies include root exudation, modulation of ion transport, and regulation of stomatal conductance to limit metal uptake and mitigate stress impacts. Biochemical responses to HM stress involve the synthesis of phytochelatins and metallothioneins for metal chelation and sequestration, alongside the activation of enzymatic (e.g., SOD, CAT, POD) and non-enzymatic (e.g., AsA, GSH) antioxidant systems to scavenge ROS and maintain cellular redox balance. At the molecular level, plants employ transcriptional reprogramming, signal transduction cascades that regulate stress-responsive genes and pathways. Key transcription factors, including WRKY, MYB, bZIP, and NAC families, along with MAPK signaling cascades, play pivotal roles in orchestrating gene expression and enhancing plant resilience under HM stress. Moreover, the advent of engineered NPs and nanoparticle-assisted bioremediation offers promising avenues for enhancing HM immobilization, reducing bioavailability, and promoting safer crop production. Nanoparticle-assisted bioremediation, which integrates biologically synthesized nanoparticles with traditional bioremediation approaches, holds promise for increasing reactivity, improving microbial resilience, and targeting specific contaminants. To bridge existing gaps and advance sustainable HM remediation strategies, future research should focus on:
- Deciphering complex plant–microbiome–NPs interactions to optimize rhizosphere processes for HM detoxification.
- Applying synthetic biology and CRISPR/Cas9-based gene editing to enhance key regulatory genes, transcription factors, and transporters for improved HM stress tolerance.
- Integrating multi-omics tools to unravel the regulatory networks and crosstalk between physiological, biochemical, and molecular pathways involved in HM stress tolerance.
- Evaluating the long-term ecological risks and field performance of NPs, with emphasis on safe design, environmental fate, and regulatory frameworks.
- Developing scalable, field-applicable nanoparticle-assisted phytoremediation protocols that combine engineered plants, beneficial microbes, and smart nanomaterials for site-specific remediation.
Collectively, these efforts will help transform laboratory advances into practical, sustainable solutions for managing HM stress in agroecosystems, ensuring resilient food production systems and safeguarding the environment for future generations.
References
- A.J.M. Baker. Accumulators and excluders-strategies in the response of plants to heavy metals. J. Plant Nutr., 1981. [DOI]
- M. Hanikenne, C. Nouet. Metal hyperaccumulation and hypertolerance: A model for plant evolutionary genomics. Curr. Opin. Plant Biol., 2011. [DOI | PubMed]
- X.S. Luo, S. Yu, Y.G. Zhu, X.D. Li. Trace metal contamination in urban soils of China. Sci. Total Environ., 2012. [DOI | PubMed]
- Z. Fu, W. Guo, Z. Dang, Q. Hu, F. Wu, C. Feng, X. Zhao, W. Meng, B. Xing, J.P. Giesy. Refocusing on nonpriority toxic metals in the aquatic environment in China. Environ. Sci. Technol., 2017. [DOI]
- A.E. Burakov, E.V. Galunin, I.V. Burakova, A.E. Kucherova, S. Agarwal, A.G. Tkachev, V.K. Gupta. Adsorption of heavy metals on conventional and nanostructured materials for wastewater treatment purposes: A review. Ecotoxicol. Environ. Saf., 2017. [DOI | PubMed]
- S. Singh, D. Kapoor, S. Khasnabis, J. Singh, P.C. Ramamurthy. Mechanism and kinetics of adsorption and removal of heavy metals from wastewater using nanomaterials. Environ. Chem. Lett., 2021. [DOI]
- R. Nazar, N. Iqbal, A. Masood, M.I.R. Khan, S. Syeed, N.A. Khan. Cadmium toxicity in plants and role of mineral nutrients in its alleviation. Am. J. Plant Sci., 2012. [DOI]
- Q. Ali, M.A. Zia, M. Kamran, M. Shabaan, U. Zulfiqar, M. Ahmad, M.F. Maqsood. Nanoremediation for heavy metal contamination: A review. Hybrid Adv., 2023. [DOI]
- M. Saeed, N. Ilyas, F. Bibi, S. Shabir, S. Mehmood, N. Akhtar, S.M. Eldin. Nanoremediation approaches for the mitigation of heavy metal contamination in vegetables: An overview. Nanotechnol. Rev., 2023. [DOI]
- E. Małkowski, K. Sitko, P. Zieleźnik-Rusinowska, Ż. Gieroń, M. Szopiński. Heavy metal toxicity: Physiological implications of metal toxicity in plants. Plant Metallomics and Functional Omics, 2019. [DOI]
- N. Ghori, T. Ghori, M.Q. Hayat, S.R. Imadi, A. Gul, V. Altay, M. Ozturk. Heavy metal stress and responses in plants. Int. J. Environ. Sci. Technol., 2019. [DOI]
- U. Krämer. Metal hyperaccumulation in plants. Annu. Rev. Plant Biol., 2010. [DOI | PubMed]
- N. Verbruggen, C. Hermans, H. Schat. Mechanisms to cope with arsenic or cadmium excess in plants. Curr. Opin. Plant Biol., 2009. [DOI | PubMed]
- H. Schat, M. Llugany, R. Bernhard. Metal-specific patterns of tolerance, uptake, and transport of heavy metals in hyperaccumulating and nonhyperaccumulating metallophytes. Phytoremediation of Contaminated Soil and Water, 2020. [DOI]
- S. Khan, S. Rehman, A.Z. Khan, M.A. Khan, M.T. Shah. Soil and vegetables enrichment with heavy metals from geological sources in Gilgit, northern Pakistan. Ecotoxicol. Environ. Saf., 2010. [DOI | PubMed]
- M. Shahid, E. Pinelli, B. Pourrut, J. Silvestre, C. Dumat. Lead-induced genotoxicity to Vicia faba L. roots in relation with metal cell uptake and initial speciation. Ecotoxicol. Environ. Saf., 2010. [DOI | PubMed]
- S. Wu, X. Zhang, B. Chen, Z. Wu, T. Li, Y. Hu, Y. Sun, Y. Wang. Chromium immobilization by extraradical mycelium of arbuscular mycorrhiza contributes to plant chromium tolerance. Environ. Exp. Bot., 2016. [DOI]
- C. Bini, M. Wahsha, S. Fontana, L. Maleci. Effects of heavy metals on morphological characteristics of Taraxacum officinale Web growing on mine soils in NE Italy. J. Geochem. Explor., 2012. [DOI]
- A. Khan, S. Khan, M.A. Khan, Z. Qamar, M. Waqas. The uptake and bioaccumulation of heavy metals by food plants, their effects on plants nutrients, and associated health risk: A review. Environ. Sci. Pollut. Res., 2015. [DOI]
- J.J. Mellem, H. Baijnath, B. Odhav. Translocation and accumulation of Cr, Hg, As, Pb, Cu and Ni by Amaranthus dubius (Amaranthaceae) from contaminated sites. J. Environ. Sci. Health Part A, 2009. [DOI]
- M. Maleki, M. Ghorbanpour, K. Kariman. Physiological and antioxidative responses of medicinal plants exposed to heavy metals stress. Plant Gene, 2017. [DOI]
- A. Michalak. Phenolic compounds and their antioxidant activity in plants growing under heavy metal stress. Pol. J. Environ. Stud., 2006
- J.A. Delmar, C.C. Su, E.W. Yu. Heavy metal transport by the CusCFBA efflux system. Protein Sci., 2015. [DOI | PubMed]
- R. Dhalaria, D. Kumar, H. Kumar, E. Nepovimova, K. Kuča, M. Torequl Islam, R. Verma. Arbuscular mycorrhizal fungi as potential agents in ameliorating heavy metal stress in plants. Agronomy, 2020. [DOI]
- P. Agarwal, R. Vibhandik, R. Agrahari, A. Daverey, R. Rani. Role of root exudates on the soil microbial diversity and biogeochemistry of heavy metals. Appl. Biochem. Biotechnol., 2024. [DOI | PubMed]
- Y. Ma, C. Zhang, R.S. Oliveira, H. Freitas, Y. Luo. Bioaugmentation with endophytic bacterium E6S homologous to Achromobacter piechaudii enhances metal Rhizoaccumulation in host Sedum plumbizincicola. Front. Plant Sci., 2016. [DOI | PubMed]
- S. Muthusaravanan, N. Sivarajasekar, J.S. Vivek, T. Paramasivan, M. Naushad, J. Prakashmaran, V. Gayathri, O.K. Al-Duaij. Phytoremediation of heavy metals: Mechanisms, methods and enhancements. Environ. Chem. Lett., 2018. [DOI]
- A. Hussain, N. Abbas, F. Arshad, M. Akram, Z.I. Khan, K. Ahmad, M. Mansha, F. Mirzaei. Effects of diverse doses of lead (Pb) on different growth attributes of Zea mays L. Agric. Sci., 2013. [DOI]
- R. Bharti, R. Sharma. Effect of heavy metals: An overview. Mater. Today Proc., 2022. [DOI]
- R. Djingova, I. Kuleff. Instrumental techniques for trace analysis. Trace Metals in the Environment, 2000. [DOI]
- I. Raskin, B.D. Ensley. Phytoremediation of Toxic Metals: Using Plants to Clean up the Environment, 2000
- C.D. Jadia, M.H. Fulekar. Phytoremediation of heavy metals: Recent techniques. Afr. J. Biotechnol., 2009
- M.K. Upadhyay, A. Majumdar. Heavy metals’ stress responses in field crops. Response of Field Crops to Abiotic Stress, 2022. [DOI]
- L. Taiz, E. Zeiger. Plant Physiology, 2002
- M.A. Farooq, A. Detterbeck, S. Clemens, K.J. Dietz. Silicon-induced reversibility of cadmium toxicity in rice. J. Exp. Bot., 2016. [DOI | PubMed]
- M. Ruyter-Hooley, A.C. Larsson, B.B. Johnson, O.N. Antzutkin, M.J. Angove. The effect of inositol hexaphosphate on cadmium sorption to gibbsite. J. Colloid Interface Sci., 2016. [DOI | PubMed]
- R. Fernández-Martínez, R. Larios, I. Gómez-Pinilla, B. Gómez-Mancebo, S. López-Andrés, J. Loredo, A. Ordóñez, I. Rucandio. Mercury accumulation and speciation in plants and soils from abandoned cinnabar mines. Geoderma, 2015. [DOI]
- R.L. Messer, P.E. Lockwood, W.Y. Tseng, K. Edwards, M. Shaw, G.B. Caughman, J.B. Lewis, J.C. Wataha. Mercury (II) alters mitochondrial activity of monocytes at sublethal doses via oxidative stress mechanisms. J. Biomed. Mater. Res. Part B Appl. Biomater., 2005. [DOI | PubMed]
- I. Zeid. Responses of Phaseolus vulgaris to chromium and cobalt treatments. Biol. Plant., 2001. [DOI]
- D. Srivastava, M. Tiwari, P. Dutta, P. Singh, K. Chawda, M. Kumari, D. Chakrabarty. Chromium stress in plants: Toxicity, tolerance and phytoremediation. Sustainability, 2021. [DOI]
- B. Pourrut, S. Jean, J. Silvestre, E. Pinelli. Lead-induced DNA damage in Vicia faba root cells: Potential involvement of oxidative stress. Mutat. Res. Genet. Toxicol. Environ. Mutagen., 2011. [DOI]
- L.H. Dao, J. Beardall. Effects of lead on two green microalgae Chlorella and Scenedesmus: Photosystem II activity and heterogeneity. Algal Res., 2016. [DOI]
- T. Helleday, R. Nilsson, D. Jenssen. Arsenic [III] and heavy metal ions induce intrachromosomal homologous recombination in the hprt gene of V79 Chinese hamster cells. Environ. Mol. Mutagen., 2000. [DOI | PubMed]
- A.M. Nicholls, T.K. Mal. Effects of lead and copper exposure on growth of an invasive weed, Lythrum salicaria L. (Purple Loosestrife). Ohio J. Sci., 2003
- G. Abdul. Toxic effects of heavy metals on plant growth and metal accumulation in maize (Zea mays L.). Iran. J. Toxicol., 2010
- D. Van Nguyen, H.M. Nguyen, N.T. Le, K.H. Nguyen, H.T. Nguyen, H.M. Le, A.T. Nguyen, N.T.T. Dinh, S.A. Hoang, C. Van Ha. Copper nanoparticle application enhances plant growth and grain yield in maize under drought stress conditions. J. Plant Growth Regul., 2022. [DOI]
- R. Hänsch, R.R. Mendel. Physiological functions of mineral micronutrients (Cu, Zn, Mn, Fe, Ni, Mo, B, Cl). Curr. Opin. Plant Biol., 2009. [DOI | PubMed]
- T. Rahman Anik, M.G. Mostofa, M.M. Rahman, S.S. Keya, C.V. Ha, M.A.R. Khan, M. Abdelrahman, M.N.K. Dao, H.D. Chu, L.-S.P. Tran. Comparative effects of ZnSO4 and ZnO-NPs in improving cotton growth and yield under drought stress at early reproductive stage. Plant Sci., 2025. [DOI | PubMed]
- J. Chatterjee, C. Chatterjee. Management of phytotoxicity of cobalt in tomato by chemical measures. Plant Sci., 2003. [DOI]
- K. Jayakumar, C.A. Jaleel, A. Municipality, E. Emirates, A. Emirates, S. Arabia. Phytochemical changes in green gram (Vigna radiata) under cobalt stress. Glob. J. Mol. Sci., 2008
- J. Katare, M. Pichhode, N. Kumar. Growth of Terminalia bellirica [(Gaertn.) Roxb.] on the Malanjkhand copper mine overburden dump spoil material. Int. J. Res.-Granthaalayah, 2015. [DOI]
- A. Arora, R.K. Sairam, G.C. Srivastava. Oxidative stress and antioxidative system in plants. Curr. Sci., 2002
- S. De Dorlodot, S. Lutts, P. Bertin. Effects of ferrous iron toxicity on the growth and mineral composition of an interspecific rice. J. Plant Nutr., 2005. [DOI]
- J. Yang, Z. Song, J. Ma, H. Han. Toxicity of molybdenum-based nanomaterials on the soybean–rhizobia symbiotic system: Implications for nutrition. ACS Appl. Nano Mater., 2020. [DOI]
- M.S.A. Ahmad, M. Ashraf. Essential roles and hazardous effects of nickel in plants. Rev. Environ. Contam. Toxicol., 2011. [DOI | PubMed]
- G.R. Shi, Q.S. Cai. Photosynthetic and anatomic responses of peanut leaves to zinc stress. Biol. Plant., 2009. [DOI]
- M.H. Siddiqui, M.H. Al-Whaibi, M.O. Basalah. Interactive effect of calcium and gibberellin on nickel tolerance in relation to antioxidant systems in Triticum aestivum L. Protoplasma, 2011. [DOI | PubMed]
- A. Schutzendubel, P. Schwanz, T. Teichmann, K. Gross, R. Langenfeld-Heyser, D.L. Godbold, A. Polle. Cadmium-induced changes in antioxidative systems, H2O2 content and differentiation in pine (Pinus sylvestris) roots. Plant Physiol., 2001. [DOI | PubMed]
- S.S. Gill, N. Tuteja. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem., 2010. [DOI | PubMed]
- N. Sarwar, Saifullah, S.S. Malhi, M.H. Zia, A. Naeem, S. Bibi, G. Farid. Role of mineral nutrition in minimizing cadmium accumulation by plants. J. Sci. Food Agric., 2010. [DOI | PubMed]
- A.A. Zunaidi, L.H. Lim, F. Metali. Heavy metal tolerance and accumulation in the Brassica species (Brassica chinensis var. parachinensis and Brassica rapa L.): A pot experiment. Heliyon, 2024. [DOI | PubMed]
- B. Pourrut, M. Shahid, C. Dumat, P. Winterton, E. Pinelli. Lead uptake, toxicity, and detoxification in plants. Rev. Environ. Contam. Toxicol., 2011. [DOI | PubMed]
- M.F. Dawood, M.R. Sofy, H.I. Mohamed, A.R. Sofy, H.A. Abdel-Kader. N-or/and P-deprived Coccomyxa chodatii SAG 216–2 extracts instigated mercury tolerance of germinated wheat seedlings. Plant Soil, 2023. [DOI]
- H.E. Osman, R.S. Fadhlallah. Impact of lead on seed germination, seedling growth, chemical composition, and forage quality of different varieties of Sorghum. J. Umm Al-Qura Univ. Appl. Sci., 2023. [DOI]
- H. Marschner. Marschner’s Mineral Nutrition of Higher Plants, 2011. [DOI]
- U. Aqeel, T. Aftab, M.M.A. Khan, M. Naeem. Excessive copper induces toxicity in Mentha arvensis L. by disturbing growth, photosynthetic machinery, oxidative metabolism and essential oil constituents. Plant Stress, 2023. [DOI]
- M.R. Broadley, P.J. White, J.P. Hammond, I. Zelko, A. Lux. Zinc in plants. New Phytol., 2007. [DOI | PubMed]
- J. Mathur, R. Panwar. Synergistic effect of pyrene and heavy metals (Zn, Pb, and Cd) on phytoremediation potential of Medicago sativa L. (alfalfa) in multi-contaminated soil. Environ. Sci. Pollut. Res., 2024. [DOI]
- A. Kabata-Pendias. Trace Elements in Soils and Plants, 2010. [DOI]
- A. Shahzad, U. Aslam, S. Ferdous, M. Qin, A. Siddique, M. Billah, M. Naeem, Z. Mahmood, S. Kayani. Combined effect of endophytic Bacillus mycoides and rock phosphate on the amelioration of heavy metal stress in wheat plants. BMC Plant Biol., 2024. [DOI | PubMed]
- J. Dong, F.-B. Wu, G.-P. Zhang. Effect of cadmium on growth and photosynthesis of tomato seedlings. J. Zhejiang Univ. Sci. B, 2005. [DOI | PubMed]
- T. Gao, H. Wang, C. Li, M. Zuo, X. Wang, Y. Liu, Y. Yang, D. Xu, Y. Liu, X. Fang. Effects of heavy metal stress on physiology, hydraulics, and anatomy of three desert plants in the Jinchang mining area, China. Int. J. Environ. Res. Public Health, 2022. [DOI | PubMed]
- L.N. Liu. Distribution and dynamics of electron transport complexes in cyanobacterial thylakoid membranes. Biochim. Biophys. Acta (BBA)-Bioenerg, 2016. [DOI]
- Z. Guo, Y. Gao, X. Yuan, M. Yuan, L. Huang, S. Wang, C. Duan. Effects of heavy metals on stomata in plants: A review. Int. J. Mol. Sci., 2023. [DOI | PubMed]
- E. Weryszko-Chmielewska, M. Chwil. Lead-induced histological and ultrastructural changes in the leaves of soybean (Glycine max (L.) Merr.). Soil Sci. Plant Nutr., 2005. [DOI]
- R. Alkhatib, J. Maruthavanan, S. Ghoshroy, R. Steiner, T. Sterling, R. Creamer. Physiological and ultrastructural effects of lead on tobacco. Biol. Plant., 2012. [DOI]
- A. Kanwal, M. Farhan, F. Sharif, M.U. Hayyat, L. Shahzad, G.Z. Ghafoor. Effect of industrial wastewater on wheat germination, growth, yield, nutrients and bioaccumulation of lead. Sci. Rep., 2020. [DOI | PubMed]
- A. Kosobrukhov, I. Knyazeva, V. Mudrik. Plantago major plants responses to increased content of lead in soil: Growth and photosynthesis. Plant Growth Regul., 2004. [DOI]
- S. Yang, J. Zhang, L. Chen. Growth and physiological responses of Pennisetum sp. to cadmium stress under three different soils. Environ. Sci. Pollut. Res., 2021. [DOI]
- E.T. Hamanishi, B.R. Thomas, M.M. Campbell. Drought induces alterations in the stomatal development program in Populus. J. Exp. Bot., 2012. [DOI | PubMed]
- E. Abo-Kassem, A. Sharaf-El-Din, J. Rozema, E.A. Foda. Synergistic effects of cadmium and NaCl on the growth, photosynthesis and ion content in wheat plants. Biol. Plant., 1995. [DOI]
- S.A. Anjum, U. Ashraf, I. Khan, M.F. Saleem. Chromium toxicity induced alterations in growth, photosynthesis, gas exchange attributes and yield formation in maize. Pak. J. Agric. Sci., 2016. [DOI]
- T. Khudsar, A. Arshi, T.O. Siddiqi, Mahmooduzzafar, M. Iqbal. Zinc-induced changes in growth characters, foliar properties, and Zn-accumulation capacity of pigeon pea at different stages of plant growth. J. Plant Nutr., 2008. [DOI]
- J. Chen, C. Zhu, D. Lin, Z.X. Sun. The effects of Cd on lipid peroxidation, hydrogen peroxide content and antioxidant enzyme activities in Cd-sensitive mutant rice seedlings. Can. J. Plant Sci., 2007. [DOI]
- R. Maheshwari, R.S. Dubey. Nickel-induced oxidative stress and the role of antioxidant defence in rice seedlings. Plant Growth Regul., 2009. [DOI]
- S. Verma, R.S. Dubey. Lead toxicity induces lipid peroxidation and alters the activities of antioxidant enzymes in growing rice plants. Plant Sci., 2003. [DOI]
- M.G. Kibra. Effects of mercury on some growth parameters of rice (Oryza sativa L.). Soil Environ., 2008
- E. Gajewska, P. Bernat, J. Długoński, M. Skłodowska. Effect of nickel on membrane integrity, lipid peroxidation and fatty acid composition in wheat seedlings. J. Agron. Crop Sci., 2012. [DOI]
- I. Ahmad, M.J. Akhtar, Z.A. Zahir, A. Jamil. Effect of cadmium on seed germination and seedling growth of four wheat (Triticum aestivum L.) cultivars. Pak. J. Bot., 2012
- A.S. Lukatkin, N.N. Kashtanova, P. Duchovskis. Changes in maize seedlings growth and membrane permeability under the effect of epibrassinolide and heavy metals. Russ. Agric. Sci., 2013. [DOI]
- D. Kumar, C.S. Seth. Photosynthesis, lipid peroxidation, and antioxidative responses of Helianthus annuus L. against chromium (VI) accumulation. Int. J. Phytoremediat., 2022. [DOI | PubMed]
- M.Y. Ashraf, R. Sadiq, M. Hussain, M. Ashraf, M.S.A. Ahmad. Toxic effect of nickel (Ni) on growth and metabolism in germinating seeds of sunflower (Helianthus annuus L.). Biol. Trace Elem. Res., 2011. [DOI | PubMed]
- K.M. Janas, J. Zielińska-Tomaszewska, D. Rybaczek, J. Maszewski, M.M. Posmyk, R. Amarowicz, A. Kosińska. The impact of copper ions on growth, lipid peroxidation, and phenolic compound accumulation and localization in lentil (Lens culinaris Medic.) seedlings. J. Plant Physiol., 2010. [DOI | PubMed]
- I.S. Sheoran, H.R. Singal, R. Singh. Effect of cadmium and nickel on photosynthesis and the enzymes of the photosynthetic carbon reduction cycle in pigeonpea (Cajanus cajan L.). Photosynth. Res., 1990. [DOI | PubMed]
- Z.R. Farooqi, M.Z. Iqbal, M. Kabir, M. Shafiq. Toxic effects of lead and cadmium on germination and seedling growth of Albizia lebbeck (L.) Benth. Pak. J. Bot., 2009
- J.-K. Zhu. Abiotic stress signaling and responses in plants. Cell, 2016. [DOI | PubMed]
- K. Vishwakarma, N. Upadhyay, N. Kumar, G. Yadav, J. Singh, R.K. Mishra, V. Kumar, R. Verma, R.G. Upadhyay, M. Pandey. Abscisic acid signaling and abiotic stress tolerance in plants: A review on current knowledge and future prospects. Front. Plant Sci., 2017. [DOI | PubMed]
- S.Y. Park, P. Fung, N. Nishimura, D.R. Jensen, H. Fujii, Y. Zhao, S. Lumba, J. Santiago, A. Rodrigues, T.-F.F. Chow. Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. Science, 2009. [DOI | PubMed]
- S.K. Panda. Impact of copper on reactive oxygen species, lipid peroxidation and antioxidants in Lemna minor. Biol. Plant., 2008. [DOI]
- A.C. Mustilli, S. Merlot, A. Vavasseur, F. Fenzi, J. Giraudat. Arabidopsis OST1 protein kinase mediates the regulation of stomatal aperture by abscisic acid and acts upstream of reactive oxygen species production. Plant Cell, 2002. [DOI | PubMed]
- A.E. Postiglione, G.K. Muday. The role of ROS homeostasis in ABA-induced guard cell signaling. Front. Plant Sci., 2020. [DOI | PubMed]
- Z.M. Pei, Y. Murata, G. Benning, S. Thomine, B. Klusener, G.J. Allen, E. Grill, J.I. Schroeder. Calcium channels activated by hydrogen peroxide mediate abscisic acid signalling in guard cells. Nature, 2000. [DOI | PubMed]
- S. Sahay, M. Gupta. An update on nitric oxide and its benign role in plant responses under metal stress. Nitric Oxide, 2017. [DOI | PubMed]
- P. Linger, A. Ostwald, J. Haensler. Cannabis sativa L. growing on heavy metal contaminated soil: Growth, cadmium uptake and photosynthesis. Biol. Plant., 2005. [DOI]
- A. Emamverdian, Y. Ding. Effects of heavy metals’ toxicity on plants and enhancement of plant defense mechanisms of Si-mediation “review”. Int. J. Environ. Agric. Res., 2017
- A. Llamas, C.I. Ullrich, A. Sanz. Cd2+ effects on transmembrane electrical potential difference, respiration and membrane permeability of rice (Oryza sativa L.) roots. Plant Soil, 2000. [DOI]
- S. Amiri, M. Mehrnia, H. Azami, D. Barzegari, M. Shavandi, M. Sarrafzadeh. Effect of heavy metals on fouling behavior in membrane bioreactors. J. Environ. Health Sci. Eng., 2010
- R.K. Sharma, M. Agrawal. Biological effects of heavy metals: An overview. J. Environ. Biol., 2005. [PubMed]
- M.A. Hossain, M.Z. Hossain, M. Fujita. Stress-induced changes of methylglyoxal level and glyoxalase I activity in pumpkin seedlings and cDNA cloning of glyoxalase I gene. Aust. J. Crop Sci., 2009
- M.A. Hossain, M.Z. Hossain, M. Fujita. Up-regulation of antioxidant and glyoxalase systems by exogenous glycinebetaine and proline in mung bean confer tolerance to cadmium stress. Physiol. Mol. Biol. Plants, 2010. [DOI | PubMed]
- K. Shah, R.G. Kumar, S. Verma, R.S. Dubey. Effect of cadmium on lipid peroxidation, superoxide anion generation and activities of antioxidant enzymes in growing rice seedlings. Plant Sci., 2001. [DOI]
- M. Gill. Heavy metal stress in plants: A review. Int. J. Adv. Res., 2014
- M.J. Rao, M. Duan, C. Zhou, J. Jiao, P. Cheng, L. Yang, W. Wei, Q. Shen, P. Ji, Y. Yang. Antioxidant defense system in plants: Reactive oxygen species production, signaling, and scavenging during abiotic Stress-Induced oxidative damage. Horticulturae, 2025. [DOI]
- X. Shi, N.S. Dalal. Evidence for a Fenton-type mechanism for the generation of OH radicals in the reduction of Cr (VI) in cellular media. Arch. Biochem. Biophys., 1990. [DOI | PubMed]
- M. Zhang, Z.Q. Jin, J. Zhao, G. Zhang, F. Wu. Physiological and biochemical responses to drought stress in cultivated and Tibetan wild barley. Plant Growth Regul., 2015. [DOI]
- H. Wu, N. Tito, J.P. Giraldo. Anionic cerium oxide nanoparticles protect plant photosynthesis from abiotic stress by scavenging reactive oxygen species. ACS Nano, 2017. [DOI | PubMed]
- H.I. Mohamed, I. Ullah, M.D. Toor, N.A. Tanveer, M.M.U. Din, A. Basit, Y. Sultan, M. Muhammad, M.U. Rehman. Heavy metals toxicity in plants: Understanding mechanisms and developing coping strategies for remediation: A review. Bioresour. Bioprocess., 2025. [DOI | PubMed]
- G. DalCorso, S. Farinati, S. Maistri, A. Furini. How plants cope with cadmium: Staking all on metabolism and gene expression. J. Integr. Plant Biol., 2008. [DOI | PubMed]
- M. Patra, N. Bhowmik, B. Bandopadhyay, A. Sharma. Comparison of mercury, lead and arsenic with respect to genotoxic effects on plant systems and the development of genetic tolerance. Environ. Exp. Bot., 2004. [DOI]
- F. Van Assche, H. Clijsters. Inhibition of photosynthesis in Phaseolus vulgaris by treatment with toxic concentrations of zinc: Effects on electron transport and photophosphorylation. Physiol. Plant., 1986. [DOI]
- A.A. Meharg, J. Hartley-Whitaker. Arsenic uptake and metabolism in arsenic resistant and nonresistant plant species. New Phytol., 2002. [DOI]
- D. Quig. Cysteine metabolism and metal toxicity. Altern. Med. Rev., 1998. [PubMed]
- K.P.C.B. Chander, P.C. Brookes, S.A. Harding. Microbial biomass dynamics following addition of metal-enriched sewage sludges to a sandy loam. Soil Biol. Biochem., 1995. [DOI]
- D. Chillé, V. Mollica-Nardo, O. Giuffrè, R.C. Ponterio, F. Saija, J. Sponer, S. Trusso, G. Cassone, C. Foti. Binding of arsenic by common functional groups: An experimental and quantum-mechanical study. Appl. Sci., 2022. [DOI]
- J. Kucharski, E. Boros, J. Wyszkowska. Biochemical activity of nickel-contaminated soil. Pol. J. Environ. Stud., 2009
- J. Kumchai, J.Z. Huang, C.Y. Lee, F.C. Chen, S.W. Chin. Proline partially overcomes excess molybdenum toxicity in cabbage seedlings grown in vitro. Genet. Mol. Res., 2013. [DOI | PubMed]
- J. Singh, A.S. Kalamdhad. Effects of heavy metals on soil, plants, human health and aquatic life. Int. J. Res. Chem. Environ., 2011
- G. Jentschke, D.L. Godbold. Metal toxicity and ectomycorrhizas. Physiol. Plant., 2000. [DOI]
- C. Leyval, K. Turnau, K. Haselwandter. Effect of heavy metal pollution on mycorrhizal colonization and function: Physiological, ecological and applied aspects. Mycorrhiza, 1997. [DOI]
- J.Á. Hall. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot., 2002. [DOI | PubMed]
- W. Shi, Y. Zhang, S. Chen, A. Polle, H. Rennenberg, Z.B. Luo. Physiological and molecular mechanisms of heavy metal accumulation in nonmycorrhizal versus mycorrhizal plants. Plant Cell Environ., 2019. [DOI | PubMed]
- N.S. Millar, A.E. Bennett. Stressed out symbiotes: Hypotheses for the influence of abiotic stress on arbuscular mycorrhizal fungi. Oecologia, 2016. [DOI | PubMed]
- D. Liao, C. Sun, S. Li, A. Tauqeer, X. Bian, J. Shen, S. Wu. The utilization and molecular mechanism of arbuscular mycorrhizal symbiosis in vegetables. Veg. Res., 2023. [DOI]
- N.O. Adeyemi, M.O. Atayese, O.S. Sakariyawo, J.O. Azeez, S.P.A. Sobowale, A. Olubode, S. Adeoye. Alleviation of heavy metal stress by arbuscular mycorrhizal symbiosis in Glycine max (L.) grown in copper, lead and zinc contaminated soils. Rhizosphere, 2021. [DOI]
- R. Chaturvedi, P.J. Favas, J. Pratas, M. Varun, M.S. Paul. Effect of Glomus mosseae on accumulation efficiency, hazard index and antioxidant defense mechanisms in tomato under metal(loid) stress. Int. J. Phytoremediat., 2018. [DOI]
- M. Riaz, M. Kamran, Y. Fang, Q. Wang, H. Cao, G. Yang, L. Deng, Y. Wang, Y. Zhou, I. Anastopoulos. Arbuscular mycorrhizal fungi-induced mitigation of heavy metal phytotoxicity in metal contaminated soils: A critical review. J. Hazard. Mater., 2021. [DOI | PubMed]
- V.A. Borowicz. Do arbuscular mycorrhizal fungi alter plant–pathogen relations?. Ecology, 2001. [DOI]
- E.F. Abd_Allah, A. Hashem, A.A. Alqarawi, A.H. Bahkali, M.S. Alwhibi. Enhancing growth performance and systemic acquired resistance of medicinal plant Sesbania sesban (L.) Merr using arbuscular mycorrhizal fungi under salt stress. Saudi J. Biol. Sci., 2015. [DOI | PubMed]
- Y. Bi, J. Zhang, Z. Song, Z. Wang, L. Qiu, J. Hu, Y. Gong. Arbuscular mycorrhizal fungi alleviate root damage stress induced by simulated coal mining subsidence ground fissures. Sci. Total Environ., 2019. [DOI | PubMed]
- H. Marschner. Mineral Nutrition of Higher Plants, 1995. [DOI]
- A.A. Dalvi, S.A. Bhalerao. Response of plants towards heavy metal toxicity: An overview of avoidance, tolerance and uptake mechanism. Ann. Plant Sci., 2013
- A.S. Jarin, M.M. Islam, A. Rahat, S. Ahmed, P. Ghosh, Y. Murata. Drought stress tolerance in rice: Physiological and biochemical insights. Int. J. Plant Biol., 2024. [DOI]
- E. Grill, S. Löffler, E.L. Winnacker, M.H. Zenk. Phytochelatins, the heavy-metal-binding peptides of plants, are synthesized from glutathione by a specific γ-glutamylcysteine dipeptidyl transpeptidase (phytochelatin synthase). Proc. Natl. Acad. Sci. USA, 1989. [DOI | PubMed]
- W.E. Rauser. Phytochelatins. Annu. Rev. Biochem., 1990. [DOI | PubMed]
- C.S. Cobbett. Phytochelatin biosynthesis and function in heavy-metal detoxification. Curr. Opin. Plant Biol., 2000. [DOI | PubMed]
- Y. Tong, R. Kneer, Y. Zhu. Vacuolar compartmentalization: A second-generation approach to engineering plants for phytoremediation. Trends Plant Sci., 2003. [DOI | PubMed]
- S. Clemens, E.J. Kim, D. Neumann, J.I. Schroeder. Tolerance to toxic metals by a gene family of phytochelatin synthases from plants and yeast. EMBO J., 1999. [DOI | PubMed]
- M. Bertrand, J.C. Guary, B. Schoefs. How plants adapt their physiology to an excess of metals. Handbook of Plant and Crop Physiology, 2001. [DOI]
- D.E. Salt, W.E. Rauser. MgATP-dependent transport of phytochelatins across the tonoplast of oat roots. Plant Physiol., 1995. [DOI | PubMed]
- S. Ebbs, I. Lau, B. Ahner, L. Kochian. Phytochelatin synthesis is not responsible for Cd tolerance in the Zn/Cd hyperaccumulator Thlaspi caerulescens (J. & C. Presl). Planta, 2002. [DOI | PubMed]
- P. Goldsbrough. Metal tolerance in plants: The role of phytochelatins and metallothioneins. Phytoremediation of Contaminated Soil and Water, 2020. [DOI]
- Y. Wang, L. Wang, C. Ma, K. Wang, Y. Hao, Q. Chen, Y. Mo, Y. Rui. Effects of Cerium Oxide on Rice Seedlings as Affected by Co-Exposure of Cadmium and Salt. Environ. Pollut., 2019. [DOI | PubMed]
- S.S. Sharma, K.J. Dietz. The relationship between metal toxicity and cellular redox imbalance. Trends Plant Sci., 2009. [DOI | PubMed]
- T.S. Tseng, S.S. Tzeng, K.W. Yeh, C.H. Yeh, F.C. Chang, Y.M. Chen, C.Y. Lin. The heat-shock response in rice seedlings: Isolation and expression of cDNAs that encode class I low-molecular-weight heat-shock proteins. Plant Cell Physiol., 1993. [DOI]
- R. Wollgiehn, D. Neumann. Metal stress response and tolerance of cultured cells from Silene vulgaris and Lycopersicon peruvianum: Role of heat stress proteins. J. Plant Physiol., 1999. [DOI]
- D. Neumann, O. Lichtenberger, D. Günther, K. Tschiersch, L. Nover. Heat-shock proteins induce heavy-metal tolerance in higher plants. Planta, 1994. [DOI]
- S. Lewis, M.E. Donkin, M.H. Depledge. Hsp70 expression in Enteromorpha intestinalis (Chlorophyta) exposed to environmental stressors. Aquat. Toxicol., 2001. [DOI | PubMed]
- K.M. Rao, T.V.S. Sresty. Antioxidative parameters in the seedlings of pigeonpea (Cajanus cajan (L.) Millspaugh) in response to Zn and Ni stresses. Plant Sci., 2000. [DOI | PubMed]
- A. Schützendübel, A. Polle. Plant responses to abiotic stresses: Heavy metal-induced oxidative stress and protection by mycorrhization. J. Exp. Bot., 2002. [DOI | PubMed]
- A.M. Bhaduri, M.H. Fulekar. Antioxidant enzyme responses of plants to heavy metal stress. Rev. Environ. Sci. Bio/Technol., 2012. [DOI]
- A. Okem, W.A. Stirk, R.A. Street, C. Southway, J.F. Finnie, J. Van Staden. Effects of Cd and Al stress on secondary metabolites, antioxidant and antibacterial activity of Hypoxis hemerocallidea Fisch. & C.A. Mey. Plant Physiol. Biochem., 2015. [DOI | PubMed]
- P. Malekzadeh, J. Khara, S. Farshian, A.K. Jamal-Abad, S. Rahmatzade. Cadmium toxicity in maize seedlings: Changes in antioxidant enzyme activities and root growth. Pak. J. Biol. Sci., 2007. [DOI | PubMed]
- Y. Cui, N. Zhao. Oxidative stress and change in plant metabolism of maize (Zea mays L.) growing in contaminated soil with elemental sulfur and toxic effect of zinc. Plant Soil Environ., 2011. [DOI]
- F. Chen, F. Wang, F. Wu, W. Mao, G. Zhang, M. Zhou. Modulation of exogenous glutathione in antioxidant defense system against Cd stress in two barley genotypes differing in Cd tolerance. Plant Physiol. Biochem., 2010. [DOI | PubMed]
- V. Lobo, A. Patil, A. Phatak, N. Chandra. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev., 2010. [DOI | PubMed]
- P. Sharma, A.B. Jha, R.S. Dubey, M. Pessarakli. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot., 2012. [DOI]
- A.S. Bali, G.P.S. Sidhu, V. Kumar. Root exudates ameliorate cadmium tolerance in plants: A review. Environ. Chem. Lett., 2020. [DOI]
- A. Bartwal, R. Mall, P. Lohani, S.K. Guru, S. Arora. Role of secondary metabolites and brassinosteroids in plant defense against environmental stresses. J. Plant Growth Regul., 2013. [DOI]
- K.A. Shanker, M. Djanaguiraman, R. Sudhagar, C.N. Chandrashekar, G. Pathmanabhan. Differential antioxidative response of ascorbate-glutathione pathway enzymes and metabolites to chromium speciation stress in green gram (Vigna radiata L. cv. CO 4) roots. Plant Sci., 2004. [DOI]
- S. Mazhoudi, A. Chaoui, M.H. Ghorbal, E. El Ferjani. Response of antioxidant enzymes to excess copper in tomato (Lycopersicon esculentum Mill.). Plant Sci., 1997. [DOI]
- K.V.S.K. Prasad, P.P. Saradhi, P. Sharmila. Concerted action of antioxidant enzymes and curtailed growth under zinc toxicity in Brassica juncea. Environ. Exp. Bot., 1999. [DOI]
- A. Hameed, T.N. Qadri, T.O. Siddiqi, M. Iqbal. Differential activation of the enzymatic antioxidant system of Abelmoschus esculentus L. under CdCl2 and HgCl2 exposure. Braz. J. Plant Physiol., 2011. [DOI]
- Z.-B. Luo, X.-J. He, L. Chen, L. Tang, S. Gao, F. Chen. Effects of zinc on growth and antioxidant responses in Jatropha curcas seedlings. Int. J. Agric. Biol., 2010
- R.A. Gomes-Junior, C.A. Moldes, F.S. Delite, P.L. Gratão, P. Mazzafera, P.J. Lea, R.A. Azevedo. Nickel Elicits a Fast Antioxidant Response in Coffea arabica Cells. Plant Physiol. Biochem., 2006. [DOI | PubMed]
- V.J. Odjegba, I.O. Fasidi. Changes in antioxidant enzyme activities in Eichhornia crassipes (Pontederiaceae) and Pistia stratiotes (Araceae) under heavy metal stress. Rev. De Biol. Trop., 2007. [DOI | PubMed]
- M.M.A. Boojar. Role of antioxidant enzyme responses and phytochelatins in tolerance strategies of Alhagi camelorum Fisch growing on copper mine. Acta Bot. Croat., 2010
- L.A. Niekerk, A. Gokul, G. Basson, M. Badiwe, M. Nkomo, A. Klein, M. Keyster. Heavy metal stress and mitogen activated kinase transcription factors in plants: Exploring heavy metal–ROS influences on plant signalling pathways. Plant Cell Environ., 2024. [DOI | PubMed]
- S. Kumar, P.K. Trivedi. Heavy metal stress signaling in plants. Plant Metal Interaction, 2016. [DOI]
- S. Ali, R.A. Mir, A. Tyagi, N. Manzar, A.S. Kashyap, M. Mushtaq, A. Raina, S. Park, S. Sharma, Z.A. Mir. Chromium toxicity in plants: Signaling, mitigation, and future perspectives. Plants, 2023. [DOI | PubMed]
- D. Chakrabarty, P.K. Trivedi, P. Misra, M. Tiwari, M. Shri, D. Shukla, S. Kumar, A. Rai, A. Pandey, D. Nigam. Comparative transcriptome analysis of arsenate and arsenite stresses in rice seedlings. Chemosphere, 2009. [DOI | PubMed]
- Y. Liu, L. Liu, J. Qi, P. Dang, T. Xia. Cadmium activates ZmMPK3-1 and ZmMPK6-1 via induction of reactive oxygen species in maize roots. Biochem. Biophys. Res. Commun., 2019. [DOI | PubMed]
- F.Y. Zhao, F. Hu, S.Y. Zhang, K. Wang, C.R. Zhang, T. Liu. MAPKs regulate root growth by influencing auxin signaling and cell cycle-related gene expression in cadmium-stressed rice. Environ. Sci. Pollut. Res., 2013. [DOI]
- S. Mansoor, A. Ali, N. Kour, J. Bornhorst, K. AlHarbi, J. Rinklebe, D. Abd El Moneim, P. Ahmad, Y.S. Chung. Heavy Metal Induced Oxidative Stress Mitigation and ROS Scavenging in Plants. Plants, 2023. [DOI | PubMed]
- N.-N. Trinh, T.-L. Huang, W.-C. Chi, S.-F. Fu, C.-C. Chen, H.-J. Huang. Chromium stress response effect on signal transduction and expression of signaling genes in rice. Physiol. Plant., 2013. [DOI]
- M. Mitra, P. Agarwal, S. Roy. Plant response to heavy metal stress: An insight into the molecular mechanism of transcriptional regulation. Plant Transcription Factors, 2022. [DOI]
- M.K. Hasan, Y. Cheng, M.K. Kanwar, X.Y. Chu, G.J. Ahammed, Z.Y. Qi. Responses of plant proteins to heavy metal stress—A review. Front. Plant Sci., 2017. [DOI | PubMed]
- Ö. Çelik, A. Ayan, S. Meriç, Ç. Atak. Heavy metal stress-responsive phyto-miRNAs. Cellular and Molecular Phytotoxicity of Heavy Metals, 2020. [DOI]
- H. Kaur, S.K. Kohli, K. Khanna, S. Dhiman, J. Kour, T. Bhardwaf, R. Bhardwaf. Deciphering the role of metal binding proteins and metal transporters for remediation of toxic metals in plants. Bioremediation of Toxic Metal(loid)s, 2022. [DOI]
- W.J. Osborne, S. Sharma. Genomic instability in medicinal plants in response to heavy metal stress. Stress-Responsive Factors and Molecular Farming in Medicinal Plants, 2023. [DOI]
- B. Iqbal, N. Ahmad, G. Li, A. Jalal, A.R. Khan, X. Zheng, D. Du. Unlocking plant resilience: Advanced epigenetic strategies against heavy metal and metalloid stress. Plant Sci., 2024. [DOI | PubMed]
- S. Lanciano, M. Mirouze. DNA methylation in rice and relevance for breeding. Epigenomes, 2017. [DOI]
- J.J. Gallo-Franco, C.C. Sosa, T. Ghneim-Herrera, M. Quimbaya. Epigenetic control of plant response to heavy metal stress: A new view on aluminum tolerance. Front. Plant Sci., 2020. [DOI | PubMed]
- Y.Y. Zhou, Y.S. Wang, C.C. Sun. Molecular cloning and expression analysis of the typical class III chitinase genes from three mangrove species under heavy metal stress. Plants, 2023. [DOI | PubMed]
- M. Zhou, S. Zheng, R. Liu, J. Lu, L. Lu, C. Zhang, Z. Liu, C. Lue, L. Zhang, L. Yant. Genome-wide identification, phylogenetic and expression analysis of the heat shock transcription factor family in bread wheat (Triticum aestivum L.). BMC Genom., 2019. [DOI | PubMed]
- S. Hu, Y. Yu, Q. Chen, G. Mu, Z. Shen, L. Zheng. OsMYB45 plays an important role in rice resistance to cadmium stress. Plant Sci., 2018. [DOI | PubMed]
- K.K. Sapara, J. Khedia, P. Agarwal, D.R. Gangapur, P.K. Agarwal. SbMYB15 transcription factor mitigates cadmium and nickel stress in transgenic tobacco by limiting uptake and modulating antioxidative defence system. Functi. Plant Biol., 2019. [DOI]
- Z. Xu, Y. Ge, W. Zhang, Y. Zhao, G. Yang. The walnut JrVHAG1 gene is involved in cadmium stress response through ABA-signal pathway and MYB transcription regulation. BMC Plant Biol., 2018. [DOI | PubMed]
- Q. Xing, M.K. Hasan, Z. Li, T. Yang, W. Jin, Z. Qi, P. Yang, G. Wang, G.J. Ahammed, J. Zhou. Melatonin-induced plant adaptation to cadmium stress involves enhanced phytochelatin synthesis and nutrient homeostasis in Solanum lycopersicum L. J. Hazard. Mater., 2023. [DOI | PubMed]
- Y. Zhou, D. Xu, L. Jia, X. Huang, G. Ma, S. Wang, M. Zhu, A. Zhang, M. Guan, K. Lu. Genome-wide identification and structural analysis of bZIP transcription factor genes in Brassica napus. Genes, 2017. [DOI | PubMed]
- P. Agarwal, M. Mitra, S. Banerjee, S. Roy. MYB4 transcription factor, a member of R2R3-subfamily of MYB domain protein, regulates cadmium tolerance via enhanced protection against oxidative damage and increases expression of PCS1 and MT1C in Arabidopsis. Plant Sci., 2020. [DOI | PubMed]
- Y. Han, T. Fan, X. Zhu, X. Wu, J. Ouyang, L. Jiang, S. Cao. WRKY12 represses GSH1 expression to negatively regulate cadmium tolerance in Arabidopsis. Plant Mol. Biol., 2019. [DOI | PubMed]
- Y. Sheng, X. Yan, Y. Huang, Y. Han, C. Zhang, Y. Ren, T. Fan, F. Xiao, Y. Liu, S. Cao. The WRKY transcription factor, WRKY13, activates PDR8 expression to positively regulate cadmium tolerance in Arabidopsis. Plant Cell Environ., 2019. [DOI | PubMed]
- C. Zhang, C. Tong, L. Cao, P. Zheng, X. Tang, L. Wang, M. Miao, Y. Liu, S. Cao. Regulatory module WRKY33–ATL31–IRT1 mediates cadmium tolerance in Arabidopsis. Plant Cell Environ., 2023. [DOI | PubMed]
- P. Zhang, R. Wang, Q. Ju, W. Li, L.-S.P. Tran, J. Xu. The R2R3-MYB transcription factor MYB49 regulates cadmium accumulation. Plant Physiol., 2019. [DOI | PubMed]
- M. Moharem, E. Elkhatib, M. Mesalem. Remediation of chromium and mercury polluted calcareous soils using nanoparticles: Sorption–desorption kinetics, speciation and fractionation. Environ. Res., 2019. [DOI | PubMed]
- F. Cao, H. Dai, P.F. Hao, F. Wu. Silicon regulates the expression of vacuolar H⁺-pyrophosphatase 1 and decreases cadmium accumulation in rice (Oryza sativa L.). Chemosphere, 2020. [DOI | PubMed]
- M.N. Khan, M. Mobin, Z.K. Abbas, K.A. AlMutairi, Z.H. Siddiqui. Role of nanomaterials in plants under challenging environments. Plant Physiol. Biochem., 2017. [DOI | PubMed]
- S. Mingyu, H. Fashui, L. Chao, W. Xiao, L. Xiaoqing, C. Liang, L. Zhongrui. Effects of nano-anatase TiO2 on absorption, distribution of light, and photoreduction activities of chloroplast membrane of spinach. Biol. Trace Elem. Res., 2007. [DOI | PubMed]
- A. Sebastian, A. Nangia, M.N.V. Prasad. Cadmium and sodium adsorption properties of magnetite nanoparticles synthesized from Hevea brasiliensis bark: Relevance in amelioration of metal stress in rice. J. Hazard. Mater., 2019. [DOI | PubMed]
- G. Wang, X. Pan, S. Zhang, Q. Zhong, W. Zhou, X. Zhang, W.J. Peijnenburg. Remediation of heavy metal-contaminated soil by biodegradable chelator–induced washing: Efficiencies and mechanisms. Environ. Res., 2020. [DOI | PubMed]
- P. Zhou, M. Adeel, N. Shakoor, M. Guo, Y. Hao, I. Azeem, Y. Rui. Application of nanoparticles alleviates heavy metals stress and promotes plant growth: An overview. Nanomaterials, 2020. [DOI | PubMed]
- J. Singh, B.K. Lee. Influence of nano-TiO2 particles on the bioaccumulation of Cd in soybean plants (Glycine max), a possible mechanism for the removal of Cd from the contaminated soil. J. Environ. Manag., 2016. [DOI]
- P. Venkatachalam, M. Jayaraj, R. Manikandan, N. Geetha, E.R. Rene, N.C. Sharma, S.V. Sahi. Zinc oxide nanoparticles (ZnONPs) alleviate heavy metal-induced toxicity in Leucaena leucocephala seedlings: A physiochemical analysis. Plant Physiol. Biochem., 2017. [DOI | PubMed]
- A.M. Shaik, M. David Raju, D. Rama Sekhara Reddy. Green synthesis of zinc oxide nanoparticles using aqueous root extract of Sphagneticola trilobata and its role in toxic metal removal and plant growth promotion. Inorg. Nano-Met. Chem., 2020. [DOI]
- S. Dai, B. Wang, Y. Song, Z. Xie, C. Li, S. Li, M. Jiang. Astaxanthin and its gold nanoparticles mitigate cadmium toxicity in rice by inhibiting cadmium translocation and uptake. Sci. Total Environ., 2021. [DOI | PubMed]
- F. Cai, X. Wu, H. Zhang, X. Shen, M. Zhang, W. Chen, X. Wang. Impact of TiO2 nanoparticles on lead uptake and bioaccumulation in rice (Oryza sativa L.). NanoImpact, 2017. [DOI]
- C. Song, F. Ye, H. Zhang, J. Hong, C. Hua, B. Wang, L. Zhao. Metal(loid) oxides and metal sulfides nanomaterials reduced heavy metals uptake in soil cultivated cucumber plants. Environ. Pollut., 2019. [DOI | PubMed]
- A. Konate, X. He, Z. Zhang, Y. Ma, P. Zhang, G.M. Alugongo, Y. Rui. Magnetic (Fe3O4) nanoparticles reduce heavy metals uptake and mitigate their toxicity in wheat seedling. Sustainability, 2017. [DOI]
- R. Sardar, S. Ahmed, A.A. Shah, N.A. Yasin. Selenium nanoparticles reduced cadmium uptake, regulated nutritional homeostasis and antioxidative system in Coriandrum sativum grown in cadmium toxic conditions. Chemosphere, 2022. [DOI | PubMed]
- Y. Zhu, Y. Dong, N. Zhu, H. Jin. Foliar application of biosynthetic nano-selenium alleviates the toxicity of Cd, Pb, and Hg in Brassica chinensis by inhibiting heavy metal adsorption and improving antioxidant system in plant. Ecotoxicol. Environ. Saf., 2022. [DOI | PubMed]
- M. Gao, X. Chang, Y. Yang, Z. Song. Foliar graphene oxide treatment increases photosynthetic capacity and reduces oxidative stress in cadmium-stressed lettuce. Plant Physiol. Biochem., 2020. [DOI | PubMed]
- Z. Bayat, M. Hassanshahian, S. Cappello. Immobilization of microbes for bioremediation of crude oil polluted environments: A mini review. Open Microbiol. J., 2015. [PubMed]
- D.S. Ken, A. Sinha. Recent developments in surface modification of nano zero-valent iron (nZVI): Remediation, toxicity and environmental impacts. Environ. Nanotechnol. Monit. Manag., 2020. [DOI]
- P. Gnanaprakasam, S.E. Jeena, D. Premnath, T. Selvaraju. Simple and robust green synthesis of Au NPs on reduced graphene oxide for the simultaneous detection of toxic heavy metal ions and bioremediation using bacterium as the scavenger. Electroanalysis, 2016. [DOI]
- S. Ghosh, I. Rusyn, O.V. Dmytruk, K.V. Dmytruk, H. Onyeaka, M. Gryzenhout, Y. Gafforov. Filamentous fungi for sustainable remediation of pharmaceutical compounds, heavy metal and oil hydrocarbons. Front. Bioeng. Biotechnol., 2023. [DOI | PubMed]
- M. Nakhjavani, V. Nikkhah, M.M. Sarafraz, S. Shoja, M. Sarafraz. Green synthesis of silver nanoparticles using green tea leaves: Experimental study on the morphological, rheological and antibacterial behaviour. Heat Mass Transf., 2017. [DOI]
- X. Cao, A. Alabresm, Y.P. Chen, A.W. Decho, J. Lead. Improved metal remediation using a combined bacterial and nanoscience approach. Sci. Total Environ., 2020. [DOI | PubMed]
- L. Della Puppa, M. Komárek, F. Bordas, J.C. Bollinger, E. Joussein. Adsorption of copper, cadmium, lead and zinc onto a synthetic manganese oxide. J. Colloid Interface Sci., 2013. [DOI | PubMed]
- J.I. Watanabe, Y. Tani, J. Chang, N. Miyata, H. Naitou, H. Seyama. As (III) oxidation kinetics of biogenic manganese oxides formed by Acremonium strictum strain KR21-2. Chem. Geol., 2013. [DOI]
- M. Rafique, I. Sadaf, M.S. Rafique, M.B. Tahir. A review on green synthesis of silver nanoparticles and their applications. Artif. Cells Nanomed. Biotechnol., 2017. [DOI | PubMed]
- T. Klaus, R. Joerger, E. Olsson, C.G. Granqvist. Silver-based crystalline nanoparticles, microbially fabricated. Proc. Natl. Acad. Sci. USA, 1999. [DOI | PubMed]
- M. Amin, F. Anwar, M.R.S.A. Janjua, M.A. Iqbal, U. Rashid. Green synthesis of silver nanoparticles through reduction with Solanum xanthocarpum L. berry extract: Characterization, antimicrobial and urease inhibitory activities against Helicobacter pylori. Int. J. Mol. Sci., 2012. [DOI | PubMed]
- G.M. Al-Senani, N. Al-Kadhi. The synthesis and effect of silver nanoparticles on the adsorption of Cu2+ from aqueous solutions. Appl. Sci., 2020. [DOI]
- U. Shedbalkar, R. Singh, S. Wadhwani, S. Gaidhani, B.A. Chopade. Microbial synthesis of gold nanoparticles: Current status and future prospects. Adv. Colloid Interface Sci., 2014. [DOI | PubMed]
- A. Bhargava, N. Jain, S. Gangopadhyay, J. Panwar. Development of gold nanoparticle-fungal hybrid based heterogeneous interface for catalytic applications. Process Biochem., 2015. [DOI]
- S.S. Shankar, A. Ahmad, R. Pasricha, M. Sastry. Bioreduction of chloroaurate ions by geranium leaves and its endophytic fungus yields gold nanoparticles of different shapes. J. Mater. Chem., 2003. [DOI]
- H. Agarwal, S.V. Kumar, S. Rajeshkumar. A review on green synthesis of zinc oxide nanoparticles—An eco-friendly approach. Resour.-Effic. Technol., 2017. [DOI]
- A. Hossain, Y. Abdallah, M.A. Ali, M.M.I. Masum, B. Li, G. Sun, Q. An. Lemon-fruit-based green synthesis of zinc oxide nanoparticles and titanium dioxide nanoparticles against soft rot bacterial pathogen Dickeya dadantii. Biomolecules, 2019. [DOI | PubMed]
- D. Chidambaram, T. Hennebel, S. Taghavi, J. Mast, N. Boon, W. Verstraete, J.P. Fitts. Concomitant microbial generation of palladium nanoparticles and hydrogen to immobilize chromate. Environ. Sci. Technol., 2010. [DOI | PubMed]
- V. Subramaniyam, S.R. Subashchandrabose, P. Thavamani, M. Megharaj, Z. Chen, R. Naidu. Chlorococcum sp. MM11—A novel phyco-nanofactory for the synthesis of iron nanoparticles. J. Appl. Phycol., 2015. [DOI]
- J.C. Tarafdar, R. Raliya. Rapid, low-cost, and ecofriendly approach for iron nanoparticle synthesis using Aspergillus oryzae TFR9. J. Nanopart., 2013. [DOI]
- D. Kumar, O.P. Dhankher, R.D. Tripathi, C.S. Seth. Titanium dioxide nanoparticles potentially regulate the mechanisms for photosynthetic attributes, genotoxicity, antioxidants defense machinery, and phytochelatins synthesis in relation to hexavalent chromium toxicity in Helianthus annuus L. J. Hazard. Mater., 2023. [DOI | PubMed]
- Y. Bai, T. Yang, J. Liang, J. Qu. The role of biogenic Fe-Mn oxides formed in situ for arsenic oxidation and adsorption in aquatic ecosystems. Water Res., 2016. [DOI | PubMed]
- R. Raj, K. Dalei, J. Chakraborty, S. Das. Extracellular polymeric substances of a marine bacterium mediated synthesis of CdS nanoparticles for removal of cadmium from aqueous solution. J. Colloid Interface Sci., 2016. [DOI | PubMed]