
The Influence of Different Light Spectra on Broiler Chicken Endocrine Systems and Productivity
Abstract
Simple Summary:
Light plays a crucial role in regulating physiological processes in broiler chickens. It not only helps birds see but also reaches deep into their brains, influencing important body functions. In broiler chickens, different LED light colors (e.g., green and blue) can affect the hormone systems that control growth, metabolism, and stress. Green light stimulates early muscle development by increasing growth-related hormones, while blue light supports later muscle formation and helps maintain oxidative balance. Light also affects the hormone melatonin, which regulates daily rhythms and influences how the body responds to light. Changes in day length and exposure to artificial light at night can impact hormone activity, growth, and stress responses. Understanding how light affects the bird’s body and applying smart lighting strategies can help improve broiler health, growth, and welfare.
Abstract:
In birds, light can penetrate the cranial bones and reach deep brain regions, where non-visual photoreceptors, especially in the hypothalamus, detect spectral and photoperiodic cues. Alongside retinal photoreception, deep-brain light sensing contributes to circadian entrainment and regulates melatonin secretion by the pineal gland. These light-driven pathways modulate endocrine activity, playing a key role in muscle development. This review explores how monochromatic light-emitting diode (LED) illumination, particularly green and blue wavelengths, affects the somatotropic axis (growth hormone-releasing hormone [GHRH]-growth hormone [GH]-insulin-like growth factor 1 [IGF-1]), the gonadal axis (gonadotropin-releasing hormone [GnRH]-luteinizing hormone [LH]/follicle-stimulating hormone [FSH]-sex steroids [testosterone, estrogen, progesterone]), the thyroid axis (thyrotropin-releasing hormone [TRH]-thyroid-stimulating hormone [TSH]-thyroxine [T4]/triiodothyronine [T3]), and the hypothalamic-pituitary-adrenal (HPA) axis (corticotropin-releasing hormone [CRH]-adrenocorticotropic hormone [ACTH]-corticosterone). Green light enhances early-stage muscle growth via GHRH and IGF-1 upregulation, while blue light supports later myogenic activity and oxidative balance. Light schedules also influence melatonin dynamics, which in turn modulate endocrine axis responsiveness to photic cues. Furthermore, variations in photoperiod and exposure to artificial lights at night (ALAN) affect thyroid activity and HPA axis reactivity, influencing metabolism, thermoregulation, and stress resilience. Together, ocular and intracranial photoreception form a complex network that links environmental light to hormonal regulation and muscle growth. These insights support the strategic use of LED lighting to optimize broiler performance and welfare.
Article type: Review Article
Keywords: poultry management, monochromatic light, muscle development, somatotropic axis, hypothalamic-pituitary-gonadal axis, hypothalamic-pituitary-thyroid axis, hypothalamic-pituitary-adrenal axis
Affiliations: Preclinics Department, Faculty of Veterinary Medicine, Ion Ionescu de la Brad Iași University of Life Sciences, 8 M. Sadoveanu Alley, 700489 Iași, Romania; lenuta.galan@iuls.ro (L.G.); carmen.solcan@iuls.ro (C.S.); Clinics Department, Faculty of Veterinary Medicine, Ion Ionescu de la Brad Iași University of Life Sciences, 8 M. Sadoveanu Alley, 700489 Iași, Romania
License: © 2025 by the authors. CC BY 4.0 Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Article links: DOI: 10.3390/ani15213209 | PubMed: 41227539 | PMC: PMC12607636
Relevance: Moderate: mentioned 3+ times in text
Full text: PDF (1.6 MB)
1. Introduction
In contemporary poultry production systems, lighting represents a critical environmental factor that directly influences broiler growth performance and physiological development. Among the key parameters involved, the photoperiod, light intensity, and spectral compositions have been identified as major modulators of physiological responses and behavioral patterns in broiler chickens.
Optimizing lighting strategies in broiler housing has proven to be essential for enhancing not only production efficiency but also bird health status and overall welfare. In this context, the implementation of light-emitting diode (LED) technology has gained increasing attention due to its numerous advantages, including extended operational lifespan and improved energy efficiency [ref. 1], but also due to the fact that it positively influences behavior, resulting in higher quality meat with a better texture [ref. 2]. Previous studies on broiler production systems have demonstrated that the use of monochromatic LED lighting, particularly within the green and blue spectra, is associated with beneficial effects on growth performance [ref. 3] and muscle development [ref. 4,ref. 5,ref. 6,ref. 7,ref. 8,ref. 9,ref. 10,ref. 11,ref. 12,ref. 13].
Exposure to these wavelengths increases serum melatonin [ref. 14] and plasma testosterone levels and stimulates myofibril proliferation, thereby contributing to increased muscle mass and body weight [ref. 15,ref. 16]. Notably, green light has proven particularly effective in promoting early-stage muscle development, while blue light exerts a more pronounced stimulatory effect during the later stages [ref. 13]. Moreover, green monochromatic light has been reported to accelerate the myogenic activity of skeletal satellite cells [ref. 17,ref. 18], whereas blue light contributes to improved meat quality by enhancing broiler antioxidant capacity [ref. 8]. Consequently, it has been hypothesized that monochromatic light may promote growth and development through hypothalamic GHRH expression modulation in broiler chickens [ref. 19]. The combined application of green and blue LED lighting has been shown to exert synergistic effects on productive performance, resulting in improved feed conversion efficiency and higher carcass yield compared with the use of individual monochromatic lights or conventional white light sources [ref. 16,ref. 20].
The underlying molecular mechanisms responsible for these effects involve an upregulation of growth hormone receptor expression, as well as the stimulation of satellite cell proliferation within skeletal muscle, both of which are critical processes for muscle development [ref. 21,ref. 22]. The duration and timing of light exposure are critically important. The common use of broad-spectrum LED lighting at nighttime in poultry facilities leads to the continuous stimulation of avian photoreceptors, interfering with the normal hormonal rhythms that depend on periods of darkness. Proper light exposure management through regulating its intensity, duration, and spectral composition is essential in reducing the endocrine-disrupting effects of artificial light at night (ALAN) in broiler chickens [ref. 23,ref. 24,ref. 25,ref. 26].
Integrating internal physiological states with external cues, the endocrine system is especially susceptible and responsive to environmental variations. Hormonal levels fluctuate according to light and dark cycles, in turn impacting metabolic, immune, and reproductive functions [ref. 27,ref. 28]. Adjusting light spectra in managing the broiler rearing environment represents an effective strategy for enhancing growth performance. The significance of this approach is supported by evidence indicating that the use of green and blue monochromatic lighting does not exert adverse effects on blood biochemical parameters or on the health status of internal organs [ref. 15,ref. 16]. Controlled manipulation of the light environment may therefore serve as a valuable tool for optimizing productive performance and improving the overall efficiency of modern poultry production systems.
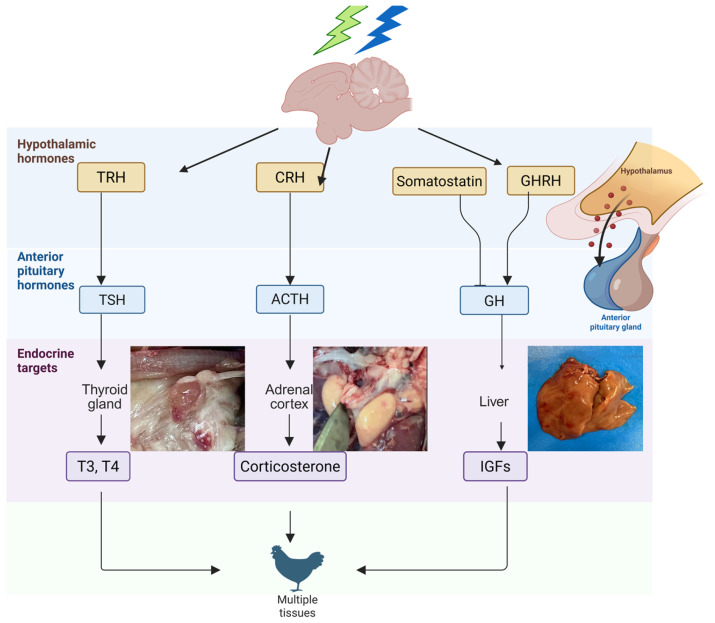
The aim of this article is to explore how light, particularly various monochromatic wavelengths (green and blue), influences the activity of the pineal–hypothalamic axis in broiler chickens. We highlight how this influence extends to the functioning of endocrine glands such as the thyroid and adrenal glands, as well as to pectoral musculature development (Figure 1). Our analysis is based on studies demonstrating that variations in lighting conditions can modulate melatonin expression and its receptor activity, with direct effects on the secretion of GH, IGF-1, and other hormones essential for poultry performance [ref. 19,ref. 29,ref. 30]. Through this approach, we aim to provide an integrated perspective on the relationship between light stimuli and the endocrine responses of the avian organism.
2. Light Detection Pathways: Visual and Non-Visual Mechanisms in Physiological Regulation
Diurnal animals’ ability to perceive and integrate light-related information is essential for appropriate environmental adaptation, allowing them to adjust to both circadian and seasonal variations [ref. 31,ref. 32]. Light functions as a major environmental cue, significantly shaping animal behavior and playing a vital role in regulating physiological functions, productivity, and overall welfare across various animal and avian species [ref. 33,ref. 34,ref. 35,ref. 36,ref. 37]. Among its many physiological roles, light supports visual perception, modulates reproductive hormone secretion, and influences social interactions. In birds, the most notable physiological responses to light are associated with changes in photoperiod and light intensity, which are known to affect reproductive cycles, health status, and behavioral patterns [ref. 21,ref. 35,ref. 38].
Poultry, in particular, possess a highly advanced visual system that plays a central role in mediating behaviors such as feeding, drinking, and social recognition [ref. 39]. Broiler chickens perceive light primarily through multiple photoreceptive sites, including the retina, pineal gland, and extraretinal photoreceptors [ref. 40,ref. 41,ref. 42] (Figure 2).
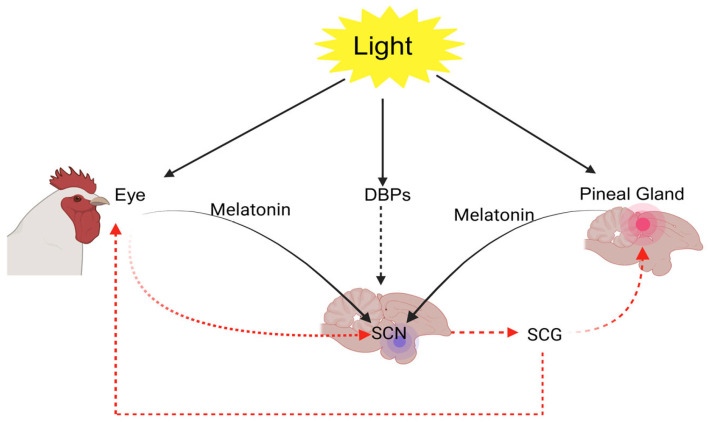
The chicken retina contains both rods, which facilitate vision under dim lighting, and cones. Notably, the retinal structure in poultry is among the most developed cone systems found in vertebrates. It includes five types of photoreceptor cones: four that are maximally sensitive to violet, blue, green, and red wavelengths of light [ref. 43,ref. 44], and one double cone that is thought to be involved in achromatic motion detection [ref. 16,ref. 42]. Beyond visual perception, light influences a wide spectrum of physiological and behavioral processes in poultry. The circadian energy homeostasis control is maintained via an intrinsic biological clock situated in the suprachiasmatic nuclei (SCN) of the hypothalamus. This clock becomes synchronized with environmental lighting through photic input from intrinsically photosensitive retinal ganglion cells, aligning physiological and behavioral rhythms with the light-dark cycle [ref. 45].
Light represents a powerful environmental cue for entraining the circadian system, although other factors such as feeding patterns can also modulate its signaling [ref. 45]. As a result, light exposure plays a vital role in shaping both growth and developmental trajectories in poultry. Responding to changes in photoperiod requires the coordinated function of three components: photoreceptors capable of detecting light, an internal timing mechanism that assesses day length, and an effector system that conveys photoperiodic information to the rest of the organism [ref. 46,ref. 47]. Numerous studies have shown that birds possess photoreceptors in various brain areas, including the pineal gland and septal regions of the telencephalon [ref. 48,ref. 49]. Birds’ extraretinal photoreceptors include several types of opsins. For instance, Opn4, Opsin 5, and vertebrate ancient opsin have been identified in the avian brain, where they respond to photoperiodic information and influence the onset and development of reproductive function. All three types of deep-brain photoreceptors (DBPs) appear to be involved in priming the neuroendocrine system to activate avian reproductive functions [ref. 46,ref. 47,ref. 50].
These opsins exhibit peak light sensitivity around 420, 480, and 490 nm, respectively [ref. 51,ref. 52,ref. 53,ref. 54]. For these deep-brain photoreceptors to be activated in non-mammalian vertebrates, light must pass through overlying tissues.
Consequently, the light spectrum that ultimately reaches these photoreceptors is largely shaped by the transmission characteristics of the intervening tissue layers [ref. 51,ref. 55]. As light passes through biological tissues, its properties are primarily altered by scattering, which favors the transmission of longer wavelengths over shorter ones. Additionally, the presence of light-absorbing molecules within the tissue further shapes the spectral profile of the transmitted light. Hemoglobin is the dominant absorber in this context, creating a spectral window that permits enhanced light transmission within the 460–540 nm range to reach internal photoreceptors [ref. 56,ref. 57].
When considering the impact of artificial light at night (ALAN), the spectral sensitivity of non-visual photoreceptors plays a key role in determining how vertebrate species perceive and respond to light stimuli. For a physiological or behavioral response to occur, the emitted spectrum of artificial light must coincide with the photoreceptor’s sensitivity range. If there is no such overlap, the light is unlikely to be perceived or to provoke any biological effects. Evidence of clock gene expression in both the mediobasal hypothalamus (MBH) and the suprachiasmatic nuclei (SCN) supports the hypothesis that birds’ photoperiodic timekeeping mechanism is located within the hypothalamus [ref. 58]. Alongside these insights, thyroid hormones have long been recognized as key regulators of the avian photoperiodic response [ref. 59].
Yoshimura et al. [ref. 60] demonstrated that in Japanese quail, the growth of gonads in response to day length is regulated by the light-stimulated conversion of the thyroid hormone thyroxine (T4) into its active form, triiodothyronine (T3), within the mediobasal hypothalamus (MBH). The enzyme responsible for this conversion, type 2 iodothyronine deiodinase (Dio2), is upregulated by light exposure. Increasing T3 levels in the brain replicates the gonadal growth normally triggered by long photoperiods, whereas blocking Dio2 prevents this development. Therefore, the light-induced activation of Dio2 and the resulting production of T3 in the MBH represent a key mechanism underlying photoperiodic reproductive function control in birds.
3. Interactions Between the Endocrine System and the Circadian Clock in Avian Physiology
Among vertebrates, circadian rhythms are predominantly governed by intracellular transcriptional and translational feedback loops involving core clock genes. These circadian processes play a critical role in the regulatory mechanism of reproductive functions in Japanese quails, as the reproductive system represents a primary endocrine function affected by light-driven rhythmic activity [ref. 61]. In contrast to mammals, the per1 gene (Period Circadian Regulator 1) is not expressed in birds, and avian cryptochromes, encoded by cry genes, may also possess light-sensing capabilities [ref. 62]. The principal molecular clock mechanism is supplemented by several auxiliary feedback loops that support physiological function regulation. One of these auxiliary loops establishes a connection between metabolic processes and the central circadian clock through metabolic sensors that interact with the core clock gene bmal1 (Basic Helix-Loop-Helix ARNT Like 1, also known as arntl). Another loop is linked to stress regulation; it modulates corticosteroid receptor activity via core clock gene interactions throughout the day [ref. 63]. Moreover, both central and peripheral feedback loops are tightly integrated with the immune system in a bidirectional regulatory relationship [ref. 64,ref. 65,ref. 66].
Photic information is transmitted to the molecular clock via cyclic adenosine monophosphate (cAMP) response elements that influence core clock and immediate early gene expression [ref. 67,ref. 68]. The main circadian loop is also coupled with melatonin signaling, as it controls the expression of enzymes involved in melatonin synthesis [ref. 69], while melatonin itself feeds back to modulate bmal1 expression [ref. 70].
Endocrine-circadian crosstalk is further reflected in the rhythmic secretion patterns of several hormones, as well as in the reciprocal influence of endocrine pathways on circadian regulation. For example, corticosterone-the primary glucocorticoid secreted through the hypothalamic-pituitary-adrenal (HPA) axis-follows a clear circadian rhythm, with peak concentrations occurring just before the onset of daily activity [ref. 71]. This pattern is generally conserved among bird species, though seasonal modulation and geographic variation (such as at higher latitudes) can influence its expression [ref. 72,ref. 73,ref. 74]. In addition, sex steroids like testosterone and estradiol display distinct circadian rhythms [ref. 75,ref. 76] and can, in turn, exert regulatory effects on circadian system functioning [ref. 77,ref. 78].
4. Pineal Gland
In birds, the pineal gland (epiphysis cerebri) is located on the dorsal surface of the brain, positioned within a triangular region between the cerebral hemispheres and the cerebellums. During embryonic development, it originates as a neuroepithelium outgrowth from the dorsal region of the posterior diencephalon, specifically between the habenular nuclei and the posterior commissure. Throughout evolution, the pineal gland has undergone significant functional and structural changes: while it served both secretory and photoreceptive roles in species like fish and amphibians, it evolved into a solely neuroendocrine organ in mammals, receiving light input indirectly via neural pathways. These evolutionary adaptations were accompanied by modifications in the gland’s structural and neuronal organization. Among birds, there is notable interspecific variation in both the anatomy and physiological roles of the pineal gland [ref. 79,ref. 80,ref. 81,ref. 82,ref. 83]. The avian pineal gland typically consists of a distal expansion called the pineal vesicle, which lies against the skull and is attached to the dura mater, and a narrower proximal part-the pineal stalk-connecting it to the dorsal wall of the third ventricle. In domestic chickens (Gallus domesticus), the pineal gland measures approximately 1.8 mm in width and 2.8 mm in length [ref. 84].
The pineal gland maintains bidirectional communication with the brain via efferent and afferent neural pathways [ref. 84,ref. 85]. In many bird species, the pineal gland receives dense innervation, primarily through noradrenergic postganglionic sympathetic fibers that originate from the superior cervical ganglia (SCG). However, unlike passerine birds, galliform species exhibit only limited neural projections from the pineal gland to the brain. The avian pineal gland consists of photoreceptor and ependymal (interstitial) cells, as well as intrinsic neurons [ref. 86]. Exhibiting characteristics similar to photoreceptors, pinealocytes are modified sensory cells that possess reduced outer segments. In numerous avian species, these cells lack synaptic connections with the gland’s intrinsic neurons. Nonetheless, immunohistochemical evidence of light-sensitive pigments such as pinopsin, which shares structural similarities with rhodopsin, has been identified in the outer segments of pinealocytes, indicating their photoreception capacity [ref. 87]. Experimental studies have further demonstrated that melatonin production in isolated pineal cells is strongly regulated by light exposure [ref. 88,ref. 89]. Reflecting the neuroendocrine nature of the pineal gland, pinealocytes contain dense-core secretory vesicles located in the Golgi apparatus and along the basal processes. The neuronal population within the gland mainly comprises bipolar cells, although pseudounipolar neurons are also present in certain bird species. Electron and light microscopic studies have shown that the pineal gland parenchyma includes not only secretory cells but also immune cells such as lymphocytes, plasma cells, basophilic granulocytes and eosinophils. This lymphoid component forms what is termed ‘lympho-pineal tissue’, suggesting an immunologic role for the gland in addition to its endocrine function. The number of proliferating cells in the pineal parenchyma is significantly higher than in the interstitial tissue, implying that pineal products may influence postthymic T cell differentiation. During development, the pineal gland in birds shows significant morphological and cytological changes, with the number and size of follicles increasing during embryonic stages and cells localizing to the outer layers as development progresses. These features emphasize the pineal gland’s dual role in endocrine melatonin secretion and potential immunomodulation in broiler chicks [ref. 90,ref. 91,ref. 92].
The pineal gland is the primary site of melatonin synthesis, a neurohormone that plays a critical role in regulating the organism’s daily and seasonal rhythms. Its production is stimulated during darkness and suppressed by light exposure [ref. 93,ref. 94], a process governed by a multisynaptic neural pathway that links the pineal gland to environmental light cues via the retina. In the avian pineal gland, melatonin synthesis is regulated through three interrelated mechanisms: the presence of direct photoreceptors, including light-sensitive photopigments, an intrinsic circadian oscillator, and noradrenergic transmission [ref. 95]. Melatonin levels are influenced not only by exposure to light and darkness but also by additional external factors such as ambient temperature and thermal stress [ref. 96]. It is estimated that approximately 80 percent of the circulating melatonin in birds originates from the pineal gland [ref. 97], while the remaining amount is produced by the retina and other structures, including the skin, gastrointestinal tract, and immune cells [ref. 98]. Due to its lipophilic nature, melatonin is capable of rapidly diffusing into various tissues and organs via both the bloodstream and the cerebrospinal fluid. Melatonin receptors are found in the central nervous system, particularly in the hypothalamus, the suprachiasmatic nucleus (SCN), and the preoptic area, as well as in peripheral organs such as the liver, lungs, ovaries, spleen, and gastrointestinal tract [ref. 99,ref. 100].
Monochromatic light has been shown to play a major role in regulating poultry growth and development [ref. 101,ref. 102]. More specifically, exposure to green monochromatic light through the action of melatonin enhances Pit-1 and GH mRNA expression and elevates GH levels in the adenohypophyseal cells of chicks, a process involving the melatonin membrane receptors Mel1b and Mel1c [ref. 103,ref. 104] (Figure 3).
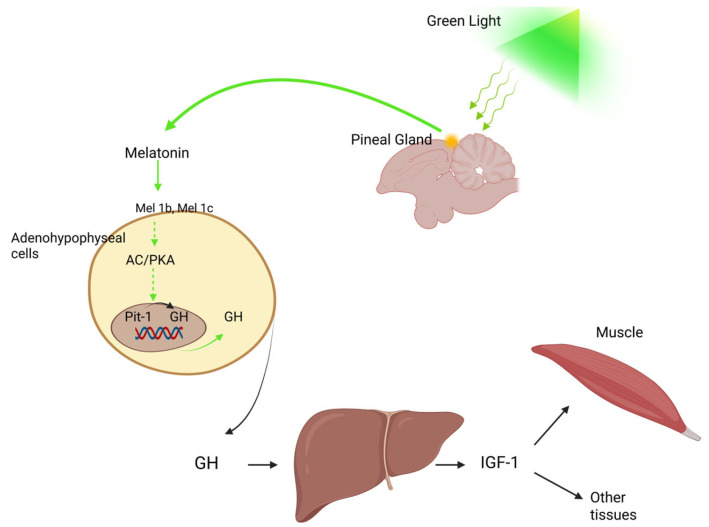
Melatonin can translate environmental cues into endocrine responses, a function that enables the synchronization of animal physiology, metabolism, and behavior with optimal environmental conditions. Aligned with photoperiodic changes, melatonin plays a fundamental role in regulating neuroendocrine processes, contributing to the biological activity of virtually all cells [ref. 105]. It also influences growth and health status [ref. 106], as well as circulating hormone levels in poultry [ref. 107]. Melatonin is closely linked with central serotonin metabolism, and several of its physiological effects are mediated through serotonergic neurotransmission modulation. In chickens, melatonin has been shown to increase serotonin concentrations [ref. 108]. Furthermore, the central adrenergic pathway in birds, which regulates growth hormone (GH) secretion [ref. 109], is also modulated by melatonin. In addition to its essential role in regulating circadian rhythms, melatonin is involved in a wide range of physiological processes, including immune function [ref. 110], reproduction, thermoregulation, energy metabolism, and antioxidant defense. This molecule directly influences lymphocyte production and antibody synthesis [ref. 111], while also contributing to the neutralization of free radicals [ref. 111,ref. 112], thereby preventing oxidative stress. Its antioxidant properties [ref. 113] are also evident in the context of liver protection and adaptation to heat stress [ref. 114,ref. 115]. Moreover, melatonin mediates the effects of monochromatic light on broiler chicken growth and development [ref. 116].
5. Hypothalamus
The hypothalamus is a crucial brain region located beneath the thalamus and dorsally to the anterior part of the pituitary gland. It plays a central role in linking the nervous and endocrine systems [ref. 117], primarily through the synthesis and release of specific neurohormones known as releasing hormones and neurotransmitters, which stimulate pituitary hormone secretion. The hypothalamus also contributes to regulating essential physiological functions, including hunger and thirst, circadian rhythms and sleep, body temperature, and fatigue levels [ref. 118,ref. 119]. The hypothalamus plays a pivotal role during developmental stages, primarily by facilitating the effects of growth hormone (GH), which in turn impacts somatic development, body composition, lipid metabolism, physical performance, and agility. It also governs intermediary metabolism through the secretion of essential regulatory hormones, including growth hormone-releasing hormone (GHRH), somatostatin (SST), and ghrelin (GHRL), which collectively influence GH activity. Unlike in mammals, where hypothalamic subregions can be functionally distinguished using Nissl staining, avian hypothalamic structures cannot be as readily dissected for individual function [ref. 117]. Therefore, as many elements of its structure and regulation remain incompletely understood, the avian hypothalamus continues to be a key area of neuroendocrine research.
Recognized as a central neuroendocrine integrative hub, the hypothalamus plays a crucial role in maintaining homeostasis by regulating key physiological processes through major endocrine axis control. The activity of this regulatory center is influenced by melatonin, which exerts its effects on hypothalamic neurons through specific receptors (Mel1a, Mel1b, Mel1c) located in critical areas such as the suprachiasmatic nuclei (SCN), the preoptic area (POA), and other regions involved in thermoregulation, metabolic control, and feeding behavior [ref. 120].
Hormonal axes represent complex networks of interactions involving the hypothalamus, the anterior pituitary, and downstream somatic target tissues, playing a crucial role in regulating growth and development [ref. 121]. In vertebrates, four major endocrine axes influence these processes: the somatotropic, gonadal, corticotropic, and thyrotropic axes [ref. 122,ref. 123].
The somatotropic axis consists of growth hormone-releasing hormone (GHRH), growth hormone (GH), the GH receptor (GHR), insulin-like growth factors (IGFs), the type 1 IGF receptor (IGFR1), and IGF-binding proteins (IGFBPs). Through its actions, this axis promotes skeletal and muscle mass development, primarily by inhibiting apoptosis and reducing lipid accumulation in adipose tissue [ref. 124,ref. 125,ref. 126,ref. 127,ref. 128,ref. 129,ref. 130,ref. 131,ref. 132,ref. 133,ref. 134,ref. 135].
The hypothalamic-pituitary-gonadal (HPG) axis constitutes a central neuroendocrine system that tightly regulates growth and reproductive function in poultry. Within this regulatory network, gonadotropin-releasing hormone (GnRH) produced by the hypothalamus binds to specific receptors on pituitary gonadotropes, stimulating the synthesis and secretion of follicle-stimulating (FSH) and luteinizing hormones (LH). Once released into the bloodstream, FSH and LH act on the gonads to promote sex hormone production. These sex hormones, in turn, exert feedback control on the hypothalamus and pituitary, modulating the ongoing synthesis and release of GnRH, FSH, and LH, and thus maintaining precise reproductive activity regulation [ref. 136].
Within the corticotropic axis, the main hormone is corticosterone (CORT), which is secreted by the adrenal cortex under the influence of adrenocorticotropic hormone (ACTH), itself stimulated by corticotropin-releasing hormone (CRH) produced in the hypothalamus. The physiological effects of CORT are mediated through its binding to the glucocorticoid receptor (NR3C1) and its association with corticosteroid-binding globulin (CBG). The activation of this system leads to energy mobilization in response to stress, increased food intake, decreased feed conversion ratio (FCR), and enhanced lipogenesis [ref. 137,ref. 138,ref. 139,ref. 140,ref. 141,ref. 142,ref. 143,ref. 144].
The thyrotropic axis involves the hypothalamic production of thyrotropin-releasing hormone (TRH), which stimulates the secretion of thyroid-stimulating hormone (TSH) [ref. 145,ref. 146,ref. 147,ref. 148,ref. 149,ref. 150]. In turn, TSH promotes the release of thyroid hormones (THs) T4 and T3 [ref. 151,ref. 152,ref. 153], the activity of which is regulated locally in target tissues through deiodinases (DIOs), enzymes responsible for the conversion and inactivation of thyroid hormones [ref. 154,ref. 155].
6. Implications of Hormone Signaling in Muscle Tissue Development
In their mature form, skeletal muscle fibers are encased and structurally supported by three connective tissue layers: the endomysium, perimysium, and epimysium. This connective tissue consists of cellular elements and an extracellular matrix (ECM), which contains both fibrous and non-fibrous proteins, notably collagens and proteoglycans. Healthy muscle tissue is characterized by clearly delineated muscle fibers, distinct endomysial spaces separating individual fibers, and well-organized perimysial spaces between fiber bundles. The preservation of this spatial organization is essential for preventing degenerative changes in muscle fibers [ref. 156]. The pectoralis muscle primarily consists of type II glycolytic fibers, which rely on anaerobic metabolism. This metabolic pathway leads to the production of lactic acid, which is predominantly cleared from the muscle via the circulatory system and subsequently reconverted to glycogen in the liver [ref. 157]. Various growth factors play critical roles as either promoters or suppressors of myoblast and satellite cell proliferation and differentiation. Among the key regulators influencing muscle hyperplasia and hypertrophy are hepatocyte growth factor (HGF), fibroblast growth factor 2 (FGF2), insulin-like growth factor (IGF), transforming growth factor-beta (TGF-β), and myostatin [ref. 156].
Given the economic significance of skeletal muscle tissue, understanding the cellular mechanisms underlying muscle growth and regeneration in broiler chickens is essential. Muscle development predominantly occurs through satellite cell proliferation [ref. 158]. Multiple studies have shown that the regenerative activity of satellite cells, specifically their proliferation and differentiation, is strongly regulated by neural innervation, vascular supply, hormonal signals, nutritional status, and the degree of tissue injury [ref. 159,ref. 160,ref. 161,ref. 162,ref. 163]. Notably, satellite cells that remain in contact with the sarcolemma of intact myofibers display reduced sensitivity to mitogenic signals [ref. 164]. Under physiological conditions, satellite cells remain in a dormant state and do not undergo cell division [ref. 165]. Nevertheless, their proliferation can be activated either to replenish the satellite stem cell pool or initiate muscle tissue repair [ref. 166]. Once activated, these cells and their descendants are known as myoblasts [ref. 165,ref. 167], which are distinguished by the prompt expression of key myogenic transcription factors such as myoblast determination protein 1 (MyoD1) and myogenic factor 5 (MYF5) [ref. 168,ref. 169]. For muscle fiber development to occur, myoblasts must differentiate into contractile structures, a process that involves the formation of myotubes—elongated, multinucleated, post-mitotic cells resulting from the fusion of several myoblasts [ref. 158]. This fusion process requires the withdrawal of myoblasts from the cell cycle and the upregulation of myogenic regulatory factors (MRFs) [ref. 170,ref. 171,ref. 172]. Initial myotubes continue to develop as they fuse with additional myoblasts, resulting in the formation of mature myotubes [ref. 173]. In adult animals, the number of muscle fibers composed of mature myotubes remains generally stable, and muscle growth occurs via increases in the size of these existing fibers [ref. 174].
According to Orna Halevy [ref. 17], exposure to green and blue lighting promoted superior body and muscle development compared with red and white lighting, a result that was strongly associated with increased satellite cell proliferation in chickens. In broiler chicks, muscle fiber formation is mostly completed by the time of hatching. During embryonic development, myoblasts proliferate and differentiate into myotubes, determining the final number of muscle fibers present at hatch [ref. 156,ref. 175]. After hatching, muscle growth relies on the fusion of satellite cells with existing myotubes and the subsequent increase in the number of nuclei [ref. 176]. Additionally, enhanced protein synthesis is necessary to ensure that protein degradation does not exceed synthesis rates [ref. 156]. Rapid muscle growth in newly hatched chicks is supported by the high proliferation rate of satellite cells, their DNA synthesis, and fusion with myotubes [ref. 177]. However, this growth process is dependent on sufficient dietary energy. Chicks deprived of feed for 24 to 48 h post-hatch show significantly reduced muscle development compared with those fed immediately after hatching [ref. 177,ref. 178]. This period of intense muscle growth typically ends around three to four weeks of age, though growth continues at a slower pace beyond this point [ref. 179]. The immediate post-hatch phase is thus the most crucial window for muscle development in chicks.
Satellite cell activity is regulated by the expression of several key genes, particularly myogenic regulatory factors (MRFs) such as myogenic determination factor 1 (MyoD1) and myogenin. MyoD1 is expressed in activated and proliferating satellite cells and plays a vital role in directing their developmental pathway [ref. 180,ref. 181]. As satellite cells progress toward terminal differentiation, myogenin expression increases, which is essential for the completion of this process [ref. 180,ref. 182]. MRF expression can be negatively influenced by feed deprivation or restricted feeding regimens in chicks [ref. 183,ref. 184] and in satellite cell cultures [ref. 185,ref. 186], which, in turn, affects breast muscle development and growth.
Hormonal signaling plays a significant role in regulating myogenesis, and muscle tissue development is influenced by the function of the somatotropic [ref. 187] and thyrotropic hormonal axes. While most research in this field has been carried out on mammalian models, several studies have also investigated chickens. For instance, treating chicken satellite cells with human IGF-1 and IGF-2 stimulates DNA synthesis [ref. 188]. This finding indicates that IGFs promote muscle hypertrophy by activating satellite cells, which later fuse with existing myotubes. Conversely, IGF-binding proteins (IGFBPs) reduce these effects; by binding growth factors, they limit their availability and lead to lower DNA synthesis in vitro [ref. 189].
Thyroid hormones (THs) can also stimulate muscle growth. Their actions are, however, context-dependent and vary with developmental stage. Embryonic chicken thigh myoblasts cultured in vitro did not divide when exposed to T4; instead, they differentiated into myotubes and remained in this state longer than untreated cells [ref. 190]. This suggests that during embryonic development, T4 plays a key role in promoting myoblast differentiation. Nyoman Suthama et al. [ref. 191] investigated the effects of dietary T4 supplementation on growth performance and muscle protein metabolism in broiler chickens. Moderate doses (0.4–1.2 ppm) promoted feed efficiency, muscle mass deposition, and reduction in abdominal fat, with sex-related differences (females showed reduced fat accumulation, while males exhibited enhanced muscle protein synthesis). In contrast, higher doses (3.6 ppm) had negative effects, leading to body weight, muscle mass, and liver weight decreases, although protein metabolism was intensified in males. Therefore, moderate T4 administration may contribute to optimizing growth and improving carcass quality by stimulating protein synthesis and reducing fat accumulation. In young chickens, dietary supplementation with T3 (0.1 mg/kg) increased thigh muscle growth from hatch to six weeks. However, the same treatment suppressed growth between six and eight weeks [ref. 192]. These observations indicate that thyroid hormones are essential for proper muscle development in early life but, nevertheless, their positive effects decline as birds grow older. Altogether, these findings confirm that both the somatotropic and thyrotropic axes contribute to muscle development, with their impact strongly dependent on the developmental stage. Light is another external factor that interacts with these endocrine influences. Light color, in particular, has a marked effect on hormonal balance in broilers. Green or green-blue light increases plasma concentrations of T3 and T4 [ref. 193,ref. 194]; in contrast, exposure to blue light lowers these hormones, along with melatonin and corticosterone [ref. 195,ref. 196]. Overall, green and blue light are generally considered more beneficial than red or white light. They reduce stress levels and help maintain calmer behavior in broiler chickens [ref. 197,ref. 198].
7. The Somatotropic Axis
7.1. Hormonal Influence on Tissues
Somatotropic hormones play a crucial role in regulating muscle, bone, and fat tissue development. The hypothalamus secretes GHRH [ref. 126], a peptide that is part of the adenylate cyclase-activating polypeptide superfamily and highly conserved across vertebrate species [ref. 199]. GHRH acts by promoting growth hormone (GH) production in the somatotroph cells of the anterior pituitary. In contrast, hypothalamic somatostatin (SST) inhibits GH synthesis by binding to the somatostatin receptor 2 (SSTR2) located in the anterior pituitary, thereby counteracting GHRH’s stimulatory effects [ref. 200,ref. 201].
Growth hormone secreted by the pituitary gland serves as a central effector in the somatotropic axis. It is a relatively unstable and straightforward protein, characterized by a single intrachain disulfide bond and a molecular weight of approximately 22 kDa [ref. 202]. The liver serves as a primary target organ for GH, producing the key effector molecules IGF-1 and IGF-2 in response to its stimulation [ref. 132,ref. 133]. In addition to its role in the GH–IGF signaling axis, GH can also exert direct metabolic effects independent of IGFs. Both GH- and IGF-dependent pathways are crucial for promoting longitudinal bone growth and increasing skeletal muscle mass.
One of GH’s main functions is enhancing hepatic IGF production, though IGFs are also locally expressed in skeletal muscle and other tissues [ref. 203] (Figure 3). The anabolic effects of IGFs on bones and muscles are primarily mediated through cell proliferation stimulation and apoptosis inhibition, mechanisms triggered by their binding to the type 1 IGF receptor (IGFR1) [ref. 204]. Unlike mammals, which possess two IGF receptors, only one has been characterized so far for avian species [ref. 205]. Both IGF-1 and IGF-2 can interact with IGFR1, although the latter binds with lower affinity than the former [ref. 206]. A critical regulatory component of the IGF-IGFR1 interaction is the insulin-like growth factor binding protein (IGFBP) family. In mammals, this group consists of seven members (IGFBP1-IGFBP7), while six have been identified in avian species. Despite being highly conserved across vertebrates [ref. 135,ref. 207,ref. 208,ref. 209,ref. 210], IGFBP6 has not been detected in any bird species to date [ref. 211,ref. 212]. IGFBP7 structurally differs from the other members and exhibits low IGF affinity; however, it binds insulin efficiently and can inhibit insulin receptor activation [ref. 213]. IGFBPs synthesized in the liver circulate in the bloodstream and can modulate IGF activity by enhancing or inhibiting receptor binding, extending IGF half-life, or altering tissue-specific IGF effects [ref. 214,ref. 215]. In addition to their role in IGF regulation, IGFBPs can also function independently. For instance, IGFBP2 has been shown to promote apoptosis in the absence of IGFs [ref. 216,ref. 217], whereas IGFBP5 can stimulate osteogenic cell proliferation [ref. 218].
IGFBPs contribute to IGF signaling fine-tuning by interacting with other elements of the somatotropic axis, such as the insulin-like growth factor acid-labile subunit (IGFALS), a key modulator of IGF activity [ref. 219]. The formation of a ternary complex involving an IGF, IGFBP3, and IGFALS significantly prolongs the circulating half-life of IGFs in the plasma [ref. 220,ref. 221]. Although IGFALS’s function has not yet been confirmed in chickens, its strong structural and functional conservation in humans and rats [ref. 222,ref. 223] suggests a likely similar role in avian species. Like IGFs and IGFBPs, IGFALS is mainly synthesized in the liver, with its gene expression induced by GH signaling [ref. 224]. Furthermore, IGFBPs are subject to regulation by specific proteases [ref. 225,ref. 226], which serve as a mechanism for controlling IGF bioavailability. The proteolytic cleavage of IGFBPs lowers their affinities for IGFs, thus releasing the growth factors to bind their receptors and exert biological effects.
IGFBPs influence growth regulation by interacting with IGFs to either increase or decrease receptor binding affinity, prolong hormone stability, or modify tissue-specific actions [ref. 214,ref. 215]. For example, IGFBP1 inhibits protein synthesis within skeletal muscles [ref. 227], whereas IGFBP2 and IGFBP4 act to suppress long bone elongation [ref. 218,ref. 228]. In myoblast cultures, IGFBP5 stimulates cell proliferation when complexed with IGF-1 but inhibits it when bound to IGF-2, while IGFBP4 restricts myoblast proliferation only in the presence of IGF-1 [ref. 229]. Additionally, certain IGFBPs have signaling capabilities independent of IGF binding; IGFBP2 can independently modulate apoptosis [ref. 216,ref. 217], while IGFBP5 can promote bone cell proliferation without IGF involvement [ref. 218]. IGFBPs thus offer an additional regulatory mechanism within the IGF system and possess direct biological functions beyond IGF mediation.
7.2. Hormone-Mediated Regulatory Signaling
The somatotropic axis is activated when growth hormone-releasing hormone (GHRH) is synthesized and secreted from the hypothalamic paraventricular nucleus (PVN), where it binds to its specific receptor (GHRHR) located on somatotroph cells in the anterior pituitary [ref. 127,ref. 128,ref. 129]. GHRHR is a G protein-coupled receptor (GPCR) embedded in the cell membrane [ref. 230,ref. 231,ref. 232]. In chickens, GHRHR shows predominant expression in the pituitary gland, while only low levels are present in certain extrapituitary tissues, including the brain, pancreas, kidneys, and testes [ref. 233]. Likewise, in rats, GHRHR and its splice variants are chiefly localized in the pituitary, where they play a key role in regulating growth hormone synthesis and release [ref. 234]. Upon ligand binding, the receptor activates the stimulatory Gα subunit of the associated heterotrimeric G protein, which, in turn, triggers adenylate cyclase to convert ATP into cAMP [ref. 235]. cAMP functions as a secondary messenger that activates protein kinase A (PKA), leading to transcription factor CREB (cAMP response element-binding protein) phosphorylation [ref. 236,ref. 237]. Once phosphorylated, CREB binds to DNA and initiates growth hormone (GH) gene transcription, ultimately promoting GH synthesis and release from somatotrophs.
This synthesis can be inhibited by the action of hypothalamic somatostatin (SST), which binds to somatostatin receptor 2 (SSTR2) on pituitary cells, blocking the PKA-dependent activation of CREB, and thus, suppressing GH transcription [ref. 200,ref. 201]. Similar to GHRHR, SSTR2 is also a GPCR [ref. 238]. This receptor exists in two isoforms-SSTR2A and SSTR2B-generated via alternative splicing [ref. 239]. Both isoforms couple to a G protein composed of α, β, and γ subunits that upon activation, inhibit adenylate cyclase activity [ref. 240]. The reciprocal release of GHRH and SST from the hypothalamus generates rhythmic fluctuations in GH output, resulting in a pulsatile pattern of secretion into the bloodstream.
Xiaojing Qin [ref. 241] demonstrated that the light spectrum influences SST levels in both the hypothalamus and pituitary of chicks, with the highest concentrations observed under red light (RL) and the lowest under green light (GL), findings that are consistent with the results of Zhang et al. [ref. 19] and Yue et al. [ref. 103]. Green light stimulates hypothalamic GHRH expression and increases plasma GH secretion, whereas red light has the opposite effect. In the hypothalamus, SST regulates GH and GHRH, while in the pituitary gland, it modulates both SST and GH. These mechanisms are essential for chick growth and development under monochromatic light. The biological action of SST is mediated by five receptor subtypes, SSTR1–5. According to Bossis and Porter [ref. 200], selective SSTR2 agonists strongly suppress both basal and GHRH-stimulated GH release, even at low nanomolar concentrations, whereas selective SSTR5 agonists reduce GH secretion only under basal conditions.
GH promotes IGF-1 synthesis in hepatocytes by interacting with the growth hormone receptor (GHR), a class I cytokine receptor that activates signaling via the JAK2/STAT5 and Src/ERK pathways [ref. 242,ref. 243,ref. 244]. In chickens, GHR is expressed not only in the liver but also in muscle and lymphoid tissues [ref. 234,ref. 245]. Akt is capable of activating various downstream targets involved in cell growth pathways [ref. 246]. One important target is the mechanistic target of rapamycin (mTOR) [ref. 247]. However, this activation is indirect and requires tuberous sclerosis complex 2 (TSC2) phosphorylation [ref. 248,ref. 249], which allows mTOR to form a complex with RPTOR, a regulatory protein [ref. 250]. This assembly, known as mTOR complex 1 (mTORC1), plays a central role in promoting protein synthesis, lipid formation, and mitochondrial development [ref. 251,ref. 252]. These cellular processes are driven by mTORC1 through the activation of transcription factors and translational regulators such as sterol regulatory element-binding proteins (SREBPs) and S6 kinases (S6Ks) [ref. 253,ref. 254].
7.3. The Effect of Light on the Somatotropic Axis
Post-hatch growth in broiler chickens primarily results from muscle fiber hypertrophy. This cellular enlargement is regulated not only by endocrine signals but also by environmental factors such as light [ref. 7,ref. 194,ref. 255]. Light influences the somatotropic axis through mechanisms involving extraretinal photoreception (notably by the hypothalamus and pineal gland), as well as melatonin-GH/IGF-1 pathway regulation. In chickens, hypothalamic photoreceptors are capable of directly detecting light stimuli.
Experimental evidence has shown that muscle fiber size increases in response to specific light wavelengths, such as green light (GL) and blue light (BL), but at different developmental stages. More specifically, green light was associated with hypertrophy during the early growth phase (days 0–26), whereas blue light had a more pronounced effect in the later phase (days 27–49) [ref. 7]. These effects may reflect underlying cellular processes, such as satellite cell proliferation prior to 3 weeks of age, and cellular degeneration thereafter [ref. 7,ref. 18].
The early post-hatch period appears critical for satellite cell activity. According to Liu et al. [ref. 18], the first three days post-hatching may represent a pivotal window for mitotic activity, which peaks at day 3. Similarly, Halevy et al. [ref. 17,ref. 256] reported that exposure to green light either during incubation or immediately after hatching enhances satellite cell proliferation, particularly evident at postnatal day 5; however, data for other time points remain unclear.
Further studies have demonstrated that green light enhances multiple physiological and molecular parameters in broilers. For instance, GL significantly increased hepatic IGF-1 secretion [ref. 29], upregulated PCNA expression in the liver of young chicks, and stimulated skeletal satellite cell proliferation [ref. 17,ref. 257]. In addition, GL was associated with increased GHRH expression in the hypothalamus [ref. 19], enhanced muscle growth [ref. 7,ref. 18], and improved meat quality [ref. 8,ref. 258]. These effects appear to be synergistic with those of growth hormone (GH), which plays a central role in promoting overall growth and productivity in broilers. Consistent with these findings, Helva [ref. 259] observed that from days 14 to 42, broilers exposed to GL and BL exhibited higher body weight compared with those exposed to white light (WL). These observations aligned with Liu et al. [ref. 18], who reported increased growth rates under GL and BL between days 5 and 21. Interestingly, in Helva’s study, green light further enhanced body weight gain between days 35 and 42. Additionally, the highest plasma IGF-1 concentration was recorded on the day of hatching, followed by a gradual decline up to day 42. Notably, light color significantly influenced IGF-1 concentration only on day 7, when WL-exposed chicks showed higher IGF-1 levels compared with those under GL and BL. However, no significant effects of light color were observed on plasma GH concentrations between days 7 and 42.
The lighting program also plays a crucial role in broiler chicken development and overall performance, as adequate feed intake during the first days of life is essential for achieving optimal growth. For this reason, near-continuous lighting schedules during the first week of life have become a common strategy for encouraging early feeding [ref. 260]. Nevertheless, many studies have shown that in broilers, prolonged exposure to almost continuous light can negatively influence growth rate, feed intake, feed conversion efficiency, mortality, and welfare [ref. 261,ref. 262]. It is therefore reasonable to consider that these adverse effects may also appear during the first week of life, when such lighting regimes are typically applied [ref. 263].
Supporting these observations, Yang Yang et al. [ref. 264] reported that constant light exposure during the early stages of life induces fear-related behaviors in broiler chickens, accompanied by elevated corticosterone levels and reduced blood melatonin/5-HT concentrations. Moreover, early-life exposure to constant light disrupts the expression of genes involved in circadian clock regulation, oxidative stress response, and the BDNF/TrκB/ERK signaling pathway in the avian hippocampus.
On the other hand, providing longer dark periods of about four to six hours during the growing phase does not negatively affect broiler performance or welfare [ref. 263,ref. 265]. Furthermore, recent findings indicate that extending the dark period even further, up to twelve hours, may actually enhance growth performance and welfare [ref. 266].
Overall, light management remains a key component of commercial broiler production systems. Research has demonstrated that extending the photoperiod or applying intermittent lighting schedules can significantly improve productive parameters such as feed intake, body weight, and feed conversion ratio, compared with constant or nearly continuous light exposure [ref. 267].
8. Hypothalamic–Pituitary–Gonadal Axis
8.1. Hormonal Influence on Tissues
The hypothalamus acts as an integrative center, receiving signals from the external environment, the gonads, and the body to coordinate reproductive system development and maturation by regulating the production of gonadotropins from the pituitary gland, thereby promoting growth and maintaining gonadal function [ref. 268].
GnRH is the primary hypothalamic neuropeptide responsible for stimulating the synthesis and release of the gonadotropins, luteinizing hormone (LH) and follicle-stimulating hormone (FSH) from the anterior pituitary, which, in turn, stimulate follicular maturation and steroidogenesis, triggering ovulation in females [ref. 269]. In this way, GnRH serves as a key link between the nervous and endocrine systems in regulating avian reproduction [ref. 270].
Synthesized by the external thecal layer of small ovarian follicles, estradiol promotes reproductive tract development, as well as secondary sexual trait and reproductive behavior manifestations [ref. 269,ref. 271]. It also contributes to the negative feedback mechanisms acting on the hypothalamus and pituitary gland [ref. 272]. Additionally, estradiol enhances the hepatic production of key yolk precursors, shifts bone metabolism toward medullary formation rather than cortical growth, and increases calcitriol activity, thereby elevating blood calcium levels and ensuring calcium availability for eggshell formation [ref. 273]. In contrast, progesterone secreted by the granulosa cells of large follicles plays a crucial role in regulating ovulation [ref. 269,ref. 271,ref. 274].
Avian reproduction is governed by a dynamic balance between GnRH stimulation and GnIH inhibition. These two systems operate antagonistically but remain interconnected and self-regulating, enabling precise gonadotropin secretion adjustment according to internal and external conditions [ref. 270].
GnIH fibers form direct synapses with GnRH neurons, modulating their electrical activity and suppressing GnRH-I gene expression. Consequently, the central administration of GnIH rapidly decreases LH levels, suggesting that GnIH may inhibit gonadal development and maintenance, as well as sexual maturation in birds, by suppressing gonadotropin synthesis and release [ref. 275].
Melatonin coordinates the GnRH-GnIH balance: during long nights, increased melatonin enhances GnIH expression in the PVN, inhibiting GnRH neurons, reducing gonadotropin secretion, and causing gonadal regression [ref. 276]. In contrast, during long days, decreased nocturnal melatonin reduces GnIH and increases GnRH activity, supporting reproductive maturation [ref. 277]. In this way, melatonin acts as a neuroendocrine translator, converting photoperiodic signals into changes in the GnRH/GnIH ratio and controlling the transition between reproductive activity and inactivity.
The GnRH-GnIH balance is further influenced by gonadal steroid levels (providing negative feedback on GnRH secretion and positive GnIH expression regulation), social and stress signals (activating GnIH via the HPA axis), and thyroid hormones, which contribute to reactivating hypothalamic photosensitivity [ref. 60,ref. 270].
These multiple interactions enable the complex integration of hormonal, seasonal, and behavioral information, ensuring precise reproductive cycle synchronization with optimal environmental conditions.
8.2. Hormone-Mediated Regulatory Signaling
The activity of neurons secreting type I gonadotropin-releasing hormone (cGnRH-I) is a pivotal factor in regulating reproductive function in birds. It is modulated by a complex network of neurotransmitters, neuropeptides, and glial cells, among which the glutamatergic system plays a key excitatory role in the hypothalamo-pituitary-gonadal (HPG) axis. N-methyl-D-aspartate (NMDA) administration has been shown to promote luteinizing hormone (LH) release, though its effect on cGnRH-I neurons appears to be indirect [ref. 278].
Vasoactive intestinal peptide (VIP) represents another crucial neuromodulator involved in regulating prolactin secretion. In quail, VIP interacts with cGnRH-I neurons and may indirectly influence their activity according to photoperiodic cues [ref. 279]. The proximity of VIP and cGnRH-I nerve terminals in the median eminence suggests the presence of fine local regulation between these neuroendocrine systems.
Opiate pathways and neuropeptide Y (NPY) also contribute to cGnRH-I modulation during the ovulatory cycle. While naloxone administration stimulates cGnRH-I secretion, β-endorphin exerts an inhibitory effect, particularly near the preovulatory LH peak [ref. 280]. NPY gene expression increases under food restriction [ref. 281], correlating with decreased cGnRH-I expression [ref. 282]. In the median eminence, NPY promotes cGnRH-I release during the preovulatory LH surge, likely under progesterone influence, which reduces β-endorphin’s inhibitory effect [ref. 283].
Dopamine is another major regulator of the GnRH system, acting at both the hypothalamic and pituitary levels to stimulate LHβ subunit expression [ref. 284]. Dopaminergic neurons near cGnRH-I neurons are activated after photostimulation [ref. 285], and direct contacts between dopaminergic fibers and cGnRH-I neurons have been observed in sexually precocious birds or under food restriction [ref. 286,ref. 287].
Glial cells further modulate interactions between neurotransmitters, neuropeptides, and cGnRH-I neurons by shaping synaptic input, particularly in response to photostimulation. Studies in quail, chickens, and mammals indicate increased cFos expression, a marker of cellular activation, in the basal hypothalamus after light exposure, in both neurons and glial cells of the median eminence [ref. 288]. In quail, post-photostimulation cGnRH-I release is proposed to result from a local increase in T3 within the basal hypothalamus. Elevated T3 may induce glial retraction around GnRH-I terminals in the median eminence, facilitating hormone release [ref. 289].
The anterior pituitary contains gonadotropes, which respond to cGnRH-I released from the median eminence. These cells synthesize and secrete LH and FSH, each comprising a shared α-subunit and a hormone-specific β-subunit (LHβ or FSHβ). In chicks, LH and FSH are produced in distinct cell populations [ref. 290], providing an important basis for studying the separate regulatory mechanisms controlling each gonadotropin.
A critical component of reproductive regulation is the interaction between GnRH and its specific receptor (cGnRH-R) expressed on gonadotropes. Receptor binding activates intracellular signaling pathways, altering gene expression for gonadotropin subunits. Two GnRH isoforms (cGnRH-I and cGnRH-II) are present in the chicken hypothalamus [ref. 291]; both are capable of stimulating LH release in vitro, and in vivo studies have confirmed that either form can induce its secretion in chickens [ref. 292]. However, gonadotropin synthesis primarily relies on cGnRH-I receptor activation, as cGnRH-II is nearly absent from the median eminence [ref. 293].
Sex-specific differences exist in pituitary responses to cGnRH-I and cGnRH-II. In domestic chickens, LH release is greater in roosters than in hens. In roosters, both GnRH forms have similar potency, whereas in hens, cGnRH-II is more effective than cGnRH-I in stimulating LH release [ref. 294]. These differences likely arise from the differential regulation of cGnRH-RI and cGnRH-RII gene expression across the reproductive cycle, explaining the sexually dimorphic response to these peptides.
Unlike mammalian receptors, avian GnRH receptors have a cytoplasmic C-terminal tail that is involved in receptor internalization, ligand binding, phosphorylation, desensitization, and expression regulation [ref. 295]. In cGnRH-RII, this tail is longer than in cGnRH-RI, with conserved internalization regions [ref. 296]. Intracellular segments linked to transmembrane domains 3 and 7, crucial for G-protein coupling, are similar between both receptor types [ref. 297,ref. 298]. cGnRH-RI internalizes through a dynamin-dependent mechanism [ref. 299].
Receptor desensitization occurs with prolonged stimulation via receptor phosphorylation, which stabilizes β-arrestin binding, inhibits G-protein coupling, and inactivates downstream signaling [ref. 300,ref. 301].
In birds, GnRH receptors primarily couple with Gq proteins, activating phospholipase C (PLC) and triggering inositol phosphate signaling. This hydrolyzes phosphatidylinositol-4,5-bisphosphate (PIP2) into inositol-1,4,5-trisphosphate (IP3) and diacylglycerol (DAG). IP3 induces intracellular Ca2+ release, while DAG activates protein kinase C (PKC), initiating cascades that regulate gonadotropin subunit gene expression [ref. 302]. GnRH receptors may also interact with Gi and Gs proteins, affecting adenylate cyclase activity. Gs activation produces cAMP, which activates PKA, phosphorylating nuclear and cytoplasmic proteins, and modulating transcription through cAMP response elements. To date, both cGnRH-RI and cGnRH-RII signal mainly through Gq [ref. 303]. In addition, a third receptor subtype, GnRH-RIII, has been identified predominantly in the pituitary gland, where it shows a distinct expression and regulation pattern closely linked to reproductive activity and its functional interplay with the GnIH system [ref. 304].
Gonadotropin receptors are localized in the gonads, particularly in ovarian granulosa and theca cells and testicular Sertoli cells. In chickens, FSH receptor expression correlates with follicular responsiveness, peaking in the granulosa cells of small yellow follicles (6–8 mm), while theca cell expression remains relatively stable across follicular stages [ref. 305]. Preovulatory follicle hierarchy is established from small yellow follicles and involves reduced epidermal growth factor expression in selected follicles, which decreases its inhibitory effect on FSH-induced steroidogenesis in granulosa cells [ref. 306].
LH stimulates the expression of its own receptor but does not significantly affect FSH receptor expression. This is counterbalanced by activin A, particularly in mid-hierarchy preovulatory follicles undergoing rapid growth, which acts paracrinely to regulate LH sensitivity [ref. 307]. In prehierarchical follicles, activin A induces LH receptor expression and, together with FSH, promotes FSH receptor expression [ref. 308].
Long-term cell culture studies show LH receptor induction in granulosa cells exposed to both FSH and LH [ref. 309]. In vivo, LH receptor mRNA is expressed in granulosa cells of hierarchical follicles, increasing progressively with follicular maturation. In theca cells, LH receptor expression is present in prehierarchical follicles and increases up to the F2 follicle [ref. 310]. This enhanced LH receptor expression in granulosa cells reflects increased progesterone production, with the highest levels in mature preovulatory follicles.
8.3. The Effect of Light on the Hypothalamic-Pituitary-Gonadal (HPG) Axis
In controlled settings, the reproductive axis can be modulated by adjusting the photoperiod. Increasing the day length elevates both mRNA and GnRH-I peptide levels, which in turn activates pituitary gonadotropes and stimulates ovarian steroidogenesis [ref. 285,ref. 304], a response primarily mediated by deep-brain photoreceptors.
Conversely, the inhibitory pathway involving GnIH is regulated by melatonin [ref. 277,ref. 311], a hormone produced during the scotophase and predominantly present under short, non-stimulatory photoperiods (<12 h). Following photostimulation, rising levels of gonadal steroids (estradiol and progesterone) provide feedback to the pituitary, reducing GnIH receptor mRNA expression [ref. 312]. Additionally, longer day lengths lower melatonin secretion, which decreases GnIH levels and relieves its inhibitory effect on the stimulatory reproductive pathway [ref. 313].
Numerous studies have reported the influence of light on hormone levels in poultry. For instance, Han et al. [ref. 314] found that extending the photoperiod enhances FSH and LH secretion in pre-laying hens. Similarly, in 20-week-old Cobb breeder chicks, Lewis et al. [ref. 315] observed that longer photoperiods were associated with increased LH levels, with the most pronounced rise at 11 h of light and a plateau at 13 h. The findings of Mengqian Liu et al. [ref. 316] are generally consistent with this, although variations may arise due to factors such as age at light stimulation initiation, differences in lighting schedules, or the chicken breeds examined.
In line with this, the authors of [ref. 316] investigated the effects of varying photoperiods on reproductive hormone levels in the hypothalamus and pituitary of male Chahua Chickens No. 2. A transcriptomic analysis was also conducted to clarify the molecular mechanisms through which the photoperiod regulates hormone secretion. The findings indicated that extending the photoperiod led to significant increases in blood levels of GnRH, FSH, and LH, confirming its strong stimulatory effect on reproductive hormone production.
Beyond photoperiod, light quality-specifically the number of photons that reach the bird and the emission spectrum-also plays a key role. Longer wavelengths penetrate the skull and brain tissue more effectively, stimulating hypothalamic photoreceptors [ref. 317]. As a result, red light is the most effective in activating the reproductive axis [ref. 41,ref. 318]. Shorter wavelengths, such as blue or green light, require higher intensities to elicit the same hypothalamic response [ref. 319], and evidence suggests that green light may even inhibit reproduction via retinal photoreceptors [ref. 320]. While the impact of green light on growth in laying hens is less clear, studies in both hens and roosters have shown that green light exposure can enhance body weight gain [ref. 41,ref. 321].
Overall, these findings highlight that both photoperiod and light characteristics critically influence the avian reproductive axis. Longer day lengths and light of longer wavelengths, particularly red light, stimulate hypothalamic photoreceptors, enhance GnRH-I activity, and promote gonadal steroidogenesis. In contrast, shorter photoperiods and inhibitory signals mediated by melatonin and GnIH suppress reproductive activity. Understanding these photic and hormonal interactions provides valuable insights for optimizing reproductive performance and growth in poultry under controlled environmental conditions.
9. Hypothalamic-Pituitary-Thyroid Axis
9.1. Anatomy and Histology of the Thyroid Gland
The avian thyroid gland is an oval-shaped, highly vascularized endocrine organ. Unlike mammals, birds possess two distinct lobes-left and right-located on either side of the trachea, near the posterior larynx, at the junction where the carotid arteries meet the subclavian arteries [ref. 322]. Histologically, the thyroid structure in birds shows no major differences compared with other vertebrates. The gland is predominantly composed of thyroid follicles, which make up around 80% of its mass, consisting of a single layer of epithelial cells called thyrocytes, which have a three-dimensional oval shape. When stimulated by thyroid-stimulating hormone (TSH), these cells become columnar; in a resting state, they appear flattened. The center of each follicle is filled with a protein-rich colloid, surrounded by the epithelial layer. The main component of this colloid is thyroglobulin (TG), a glycoprotein with two major roles: (1) it acts as a precursor for thyroid hormone (TH) synthesis due to the presence of tyrosyl residues and (2) serves as a large intrathyroidal T3 and T4 reservoir. This hormone storage mechanism is a unique adaptation that helps the organism cope with periodic iodine deficiency. The outer boundary of the follicle is formed by the basal membrane of the thyrocytes, which is in direct contact with an extensive capillary network, facilitating efficient exchange between the blood and follicular lumen. Tight junctions between these cells create a selective barrier, preventing the unwanted diffusion of transmembrane proteins and uncontrolled hormone release into circulation. In addition, parafollicular or C cells are found between follicles. These cells are involved in the production and secretion of calcitonin, a hormone that plays a role in calcium and phosphate metabolism [ref. 323].
9.2. Hormonal Influence on Tissues
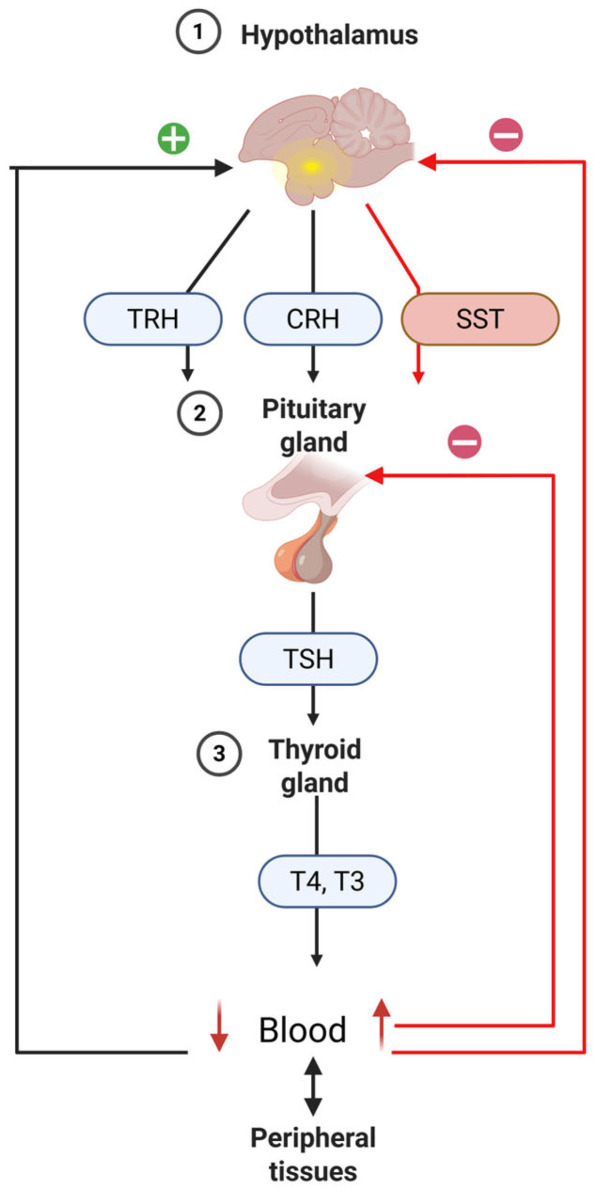
The hypothalamic-pituitary-thyroid (HPT) axis is an endocrine system regulated by a negative feedback loop. When the body experiences a deficiency in thyroid hormones (THs), the hypothalamus-specifically the paraventricular nucleus (PVN)-releases thyrotropin releasing hormone (TRH). This hormone travels through the hypothalamo-hypophyseal portal circulation to the pituitary gland where it promotes thyroid stimulating hormone (TSH) production and release into the bloodstream. TSH binds to its specific receptor on thyroid cells, triggering thyroid hormone production and secretion [ref. 151]. During embryonic development and growth, TRH influences pituitary cell function [ref. 324,ref. 325]. In adult birds, however, its effect on thyroid function becomes less pronounced, while its role in stimulating GH secretion increases [ref. 325]. TSH encourages thyroid gland enhancement and boosts the synthesis and release of T4 and, to a lesser extent, T3. These hormones circulate in the blood both freely and bound to transport proteins. Once the concentration of T4 and T3 in the blood reaches appropriate levels, they exert negative feedback on the hypothalamus and pituitary, reducing the secretion of TRH and TSH (Figure 4). In birds, corticotropin-releasing hormone (CRH) also acts as a significant thyrotropic factor, working via CRHR2 receptors to stimulate TSH synthesis and release from anterior pituitary thyrotropes [ref. 326]. Besides TRH and CRH, the HPT axis is further regulated by somatostatin (SST), a hypothalamic peptide that suppresses TSH production in the pituitary [ref. 327]. TSH activates thyroid cells by binding to its receptor and promotes TH production and release [ref. 151].
Thyroid hormones (THs), namely, thyroxine (T4) and triiodothyronine (T3), are derived from thyroglobulin and secreted by the thyroid gland [ref. 328]. These hormones are critical basal metabolic rate and systemic homeostasis regulators. They are indispensable for normal development and growth and contribute significantly to the proper function of the nervous, muscular, circulatory, and reproductive systems. In birds, T4 is the primary hormone synthesized by the thyroid gland, while T3 is mainly produced outside the thyroid through the enzymatic conversion (deiodination) of T4. Evidence shows that under both resting conditions and during hormonal stimulation, the avian thyroid predominantly secretes T4, which accounts for over 99% of total TH output, whereas T3 is released in only minimal amounts [ref. 329].
T4 is considered less potent than T3 due to its reduced thyroid hormone receptor binding affinity [ref. 152]. In circulation, both hormones are bound to carrier proteins such as transthyretin (TTR), albumin, and thyroxine-binding globulin (TBG) [ref. 330]. These transport proteins enhance solubility in plasma and facilitate hormone delivery to target tissues by preventing their passive diffusion across lipid membranes. Once inside the target tissue, T4 is locally converted to T3 by deiodinase enzymes DIO1 and DIO2, which remove an iodine atom from its outer ring [ref. 154,ref. 155]. The liver is also a major site of T3 production, representing its primary source [ref. 331]. T3 has a much higher binding affinity for receptors compared with T4. It acts primarily through nuclear thyroid hormone receptors, THRA and THRB, which activate either after direct cellular uptake or following intracellular conversion from T4.
To maintain appropriate hormonal signaling, T3 can be deactivated locally in target tissues by DIO1 or DIO3 through its conversion into 3,5-diiodo-L-thyronine (T2). Additionally, DIO3 may block the formation of T3 from T4 by converting the latter into reverse T3 (rT3) [ref. 332,ref. 333]. Deiodinase enzymes provide a mechanism for tissue-specific thyroid hormone action regulation. Thus, plasma THs levels do not always accurately reflect the TH activity at the tissue level due to the localized nature of these enzymes. Type I deiodinase (DIO1) is predominantly found in the liver, kidneys, and muscle, while Type II deiodinase (DIO2) is primarily expressed in the brain [ref. 334], particularly in the pituitary, where it is responsible for producing over 75% of the local T3. In contrast, Type III deiodinase (DIO3) is widely distributed in many tissues. During embryonic development, its highest activity is seen in the liver and kidneys, and in adult birds, it is also present in the brain, pituitary, heart, skeletal muscle, thyroid, and skin [ref. 322]. Thyroid hormone receptors (THRs) are located in the cell nucleus and act as T3 binding-activated transcription factors [ref. 335].
Free thyroid hormones (THs) travel through the bloodstream to reach their target tissues. To regulate gene expression, they must cross the cell membrane. Research has shown that specific transmembrane proteins facilitate iodothyronine transport into and out of cells [ref. 336]. These transporters belong to three families: (i) monocarboxylate transporters (MCTs), (ii) organic anion transporting polypeptides (OATPs), and (iii) L-type amino acid transporters (LATs). These systems primarily transport T4, which is then converted to T3 inside the cell. All THRs contain a nuclear localization signal [ref. 337], which ensures their proper transport into the nucleus, often with the help of N-terminal peptides [ref. 338]. Once T3 binds to THRs at thyroid hormone response elements on DNA, conformational changes allow co-regulator and transcriptional machinery recruitment, modulating gene expression [ref. 339].
9.3. Hormone-Mediated Regulatory Signaling
The thyrotropic axis is centrally regulated by neurons in the paraventricular nucleus (PVN) that secrete thyrotropin-releasing hormone (TRH). These neurons play a critical role in maintaining the normal activity of the axis. TRH expression is influenced by various hormones, most notably from the thyroid, which exert negative feedback control: elevated TH levels suppress TRH production, while low levels enhance it [ref. 340]. Other key modulators include leptin [ref. 341], α-melanocyte-stimulating hormone (α-MSH), and noradrenaline (NA) [ref. 342]. Once produced in the hypothalamus, TRH enters the portal circulation and stimulates TSH secretion from the anterior pituitary thyrotrophs by binding to TRH receptor 1 (TRHR-1), a GPCR [ref. 343]. In chickens, both TRHR-1 and TRHR-3 have been identified [ref. 344]. TRHR-1 activation triggers signaling cascades via cAMP or IP3, leading to the recruitment of transcription factors such as CREB, AP-1, and Elk-1 [ref. 146,ref. 147].
TSH primarily acts on the thyroid gland, promoting TH synthesis. Its receptor, TSHR, is located on the basolateral membrane of thyroid follicular cells [ref. 345] and is also a GPCR. When activated, thyroglobulin is mobilized from the colloid and internalized into follicular cells [ref. 328]. This glycoprotein serves as both a scaffold and a precursor for thyroid hormone synthesis [ref. 346], and is present in chickens both before and after hatching [ref. 347,ref. 348]. TSH enhances thyroglobulin gene expression [ref. 349]. Within the follicular cells, tyrosine residues on thyroglobulin undergo iodination, forming monoiodotyrosine (MIT) and diiodotyrosine (DIT), which subsequently couple to produce THs. T4 results from two DITs, while T3 is formed from a DIT and an MIT [ref. 350,ref. 351]. Although thyroid hormone release was once believed to occur through passive diffusion, studies show that low plasma TH levels occur when transport proteins like MCT8 are dysfunctional [ref. 352]. This suggests that a dual mechanism involving both passive and active transport is required to maintain TH homeostasis.
Thyroid hormone signaling involves multiple mechanisms. THRs mediate genomic responses by binding T3 intracellularly and regulating gene expression through interaction with thyroid hormone response elements (TREs) [ref. 353,ref. 354]. Ligand binding induces structural changes that enhance receptor affinity for TREs [ref. 355]. These elements may contain palindromic half-sites, and THRs can form monomers, homodimers, or heterodimers (THRA/THRB) to modulate transcription differently [ref. 356,ref. 357,ref. 358]. Their action is further strengthened by coactivator proteins (TRAPs) [ref. 359].
A second essential component of TH regulation is the deiodinase (DIO) enzyme system, which, although not directly involved in receptor signaling, modulates hormone bioavailability by removing iodine atoms from THs, thereby fine-tuning their activity in a tissue-specific manner [ref. 360].
In conclusion, the proper functioning of the thyrotropic axis depends on an integrated network of regulatory mechanisms, including the release of TRH and TSH, the synthesis and iodination of thyroglobulin for thyroid hormone production, their transport and secretion into the circulation, and deiodinase (DIO) enzymatic activity. Collectively, these processes establish a highly coordinated and dynamic regulatory system that is essential for development, growth, and metabolic homeostasis maintenance in both birds and mammals.
9.4. The Effect of Light on the Hypothalamic-Pituitary-Thyroid (HPT) Axis
Thyroid hormones are characterized by distinct circadian and/or seasonal secretion patterns and are highly sensitive to photoperiodic changes. As such, exposure to artificial light at night (ALAN) may significantly affect their secretion, thereby influencing a wide range of physiological processes. This section provides a brief overview of THs’ involvement in regulating circadian and seasonal rhythms, outlining the potential effects of ALAN on thyroid function.
T3 and T4 play critical roles in growth regulation [ref. 361]. Their levels are positively associated with feed intake in chicks [ref. 362,ref. 363], and they have been implicated in both growth suppression and compensatory growth stimulation in broilers [ref. 364]. Beyond their role in growth, THs are essential regulators of basal metabolic rate and are actively involved in thermoregulatory processes. They are indispensable for the proper development and function of nearly all tissues. Moreover, they contribute to photoperiodic regulation, modulate the hypothalamic-pituitary-gonadal axis, and play a part in the molting cycle.
At the cellular level, the physiological effects of THs are mediated by specific nuclear and/or membrane receptors and depend on hormone availability. In birds, since the thyroid gland produces only limited amounts of T3, its circulating levels are largely determined by deiodinase expression and activity, enzymes responsible for the local conversion and metabolism of this iodothyronine in peripheral tissues. Seasonal variation in the timing of peak T3 and T4 levels suggests a strong photoperiodic influence on thyroid activity [ref. 365]. This variation highlights the fundamental role of thyroid hormones in the photoperiodic responses of vertebrates, acting as central mediators of physiological adaptation to changes in day length [ref. 366,ref. 367]. ALAN can interfere with the organism’s ability to accurately perceive natural day length, leading the neuroendocrine system to interpret such lighting conditions as an extension of the photoperiod and mimicking a long-day environment. As a result, ALAN exposure may profoundly reshape endocrine and physiological states, promoting a shift toward long-day phenotypes. Within the context of thyroid function, extended photoperiods initiate a physiological cascade that enhances the peripheral conversion of T4 to the more biologically active T3, a process that plays a key role in gonadal activation. Elevated TSH and T3 levels under long-day conditions have been documented across several species, further supporting the involvement of this mechanism [ref. 368].
In addition to photoperiod duration, the spectral composition of light can modulate thyroid hormone dynamics in birds. This is exemplified by studies reporting significantly higher plasma T4 concentrations in birds exposed to GL compared with those maintained under WL, despite identical photoperiodic conditions [ref. 369].
Shahabodin Gharahveysi [ref. 13] reported that broiler chicks reared under green light achieved greater body weight compared with those exposed to yellow or red. This outcome was linked to modifications in thyroid hormone and testosterone secretion, both of which are known to directly influence metabolic activity and muscle development.
Green light is thought to reduce metabolic rate by regulating thyroid hormone levels, thereby improving energy efficiency and facilitating increased body mass accumulation [ref. 370]. Moreover, this specific light spectrum has been shown to enhance muscle growth, particularly during the early phases of development, as evidenced by several previous studies [ref. 7,ref. 197,ref. 262]. Nonetheless, it is important to note that changes in circulating levels of thyroid hormones and other blood metabolites were not consistently significant across all experimental conditions. Rozenboim [ref. 194] explored the effects of age-dependent combinations of green and blue light on growth performance and T3 concentrations, though no clear and consistent relationship was established. Although the highest T3 concentration was found in the heaviest chicks from a group exposed to a transition from blue to green LED lighting at 20 days of age, this correlation was not uniformly observed. In contrast, environmental temperature appeared to have a more pronounced effect on metabolic function than lighting conditions, with a significant correlation reported between ambient temperature and T3 levels, as demonstrated in earlier research [ref. 364,ref. 371]. Liu et al. [ref. 370] provided evidence that various combinations of monochromatic light can substantially influence the circulating thyroid hormone levels in broiler chickens. More specifically, exposure to green-blue light was associated with significantly lower concentrations of T4, FT4, FT3, and TSH, while the red–red light combination led to the highest hormonal levels.
The variations observed were accompanied by shifts in gene expression related to muscle fiber types. A negative association was found between FT3 levels and genes linked to oxidative fibers, whereas a positive association emerged with those associated with glycolytic fibers. These patterns indicate that thyroid hormones may directly contribute to muscle fiber-type transitions triggered by light stimuli. This aligns with the broader understanding that broiler muscle phenotype development is governed by intricate interactions between genetic determinants and external modulators such as neural signaling, contractile activity, and hormonal milieu. Among the endocrine regulators involved, T3 plays a central role by acting through its specific nuclear receptors, influencing the transcription of genes coding for different myosin heavy chain (MHC) isoforms [ref. 372]. Reduced TH availability tends to favor slow-twitch fiber trait expression, while elevated levels promote fast-twitch, glycolytic fiber development. This regulatory mechanism has been further substantiated by thyroidectomy experiments, which demonstrated that in the absence of THs, the modulatory effects of light on muscle fiber-type differentiation are eliminated.
The enzymes DIO2 and DIO3 play essential roles in intracellular TH activity regulation, functioning as key modulators of hormonal activation and inactivation pathways [ref. 373]. DIO2 facilitates prohormone T4 conversion into its active form, T3, thereby enhancing its local bioavailability and intensifying its physiological effects. Conversely, DIO3 acts as an inactivating enzyme by converting T4 into reverse T3 (rT3) and T3 into T2, effectively dampening T3′s classical nuclear actions [ref. 374]. Sagliocchi et al. [ref. 375] further linked DIO2 upregulation to metabolic adjustments favoring the expression of faster and more glycolytic MHC isoforms, suggesting that these deiodinases play a direct role in shaping muscle fiber characteristics. Light environment appears to influence these enzymatic pathways. Exposure to green-blue light was associated with increased DIO3 expression and decreased DIO2 expression, while the opposite pattern was observed under red–red lighting conditions. These molecular alterations mirrored the observed differences in serum thyroid hormone concentrations, indicating that specific light spectra may modulate intramuscular T3 levels through differential deiodinase activity regulation. This mechanism likely contributes to the observed shifts in muscle fiber phenotype under varying light conditions.
TH levels are also tightly linked to energy metabolism and body mass regulation [ref. 376]. Elevated TH levels, such as those seen in hyperthyroid states, increase basal metabolic rate and resting energy expenditure, often leading to weight loss despite higher food intake. This paradox arises from enhanced futile metabolic cycling and reduced energy utilization efficiency [ref. 155]. Beyond their metabolic role, thyroid hormones are involved in regulating oxidative stress by influencing reactive oxygen species production in target tissues [ref. 377]. Interestingly, slightly lower TH levels have been associated with protective effects, potentially due to reduced oxidative stress and improved cellular resilience [ref. 378]. The results emphasize that the influence of light on broiler development is closely linked to interactions with other environmental factors, such as temperature, and largely mediated by the thyroid axis. Through the TH-DIO2/DIO3 signaling pathway, thyroid hormones play a central role in regulating muscle fiber type conversion and growth efficiency. In particular, illumination with blue-green light modulates this hormonal pathway, favoring oxidative fiber development, muscle hypertrophy and antioxidant capacity. These findings support lighting regime optimization as a viable strategy for improving meat quality and productive performance in intensive rearing systems.
10. Hypothalamus-Pituitary-Adrenal Axis
10.1. Anatomy and Histology of the Adrenal Glands
Present in pairs, the adrenal glands are positioned anterior and medial to the cranial pole of the kidney. Despite considerable variation in their shape across species, they are typically flattened and lie in close proximity, occasionally fusing in some animals. These glands receive arterial blood primarily from the cranial renal arteries, and sometimes directly from the aorta. Each adrenal gland drains through a single adrenal vein into the caudal (posterior) vena cava. Additionally, in chicks, one or two lymphatic vessels are associated with each gland [ref. 379]. Preganglionic sympathetic fibers from the thoracic and sacral splanchnic nerves [ref. 380] converge toward cranial and caudal ganglia, which are either located on the pericapsular sheath or integrated within the glandular tissue. These predominantly cholinergic preganglionic fibers bypass the ganglia and directly innervate chromaffin cell clusters within the gland. Each cluster typically contains 12 or more chromaffin cells [ref. 381]. In the adrenal gland, the subcapsular area consists exclusively of medullary tissue.
Originating from the ganglia, postganglionic sympathetic fibers, which are noradrenergic in nature, supply the adrenal blood vessels and may also target chromaffin cells [ref. 382]. These fibers also contribute to the formation of renal and gonadal nerve plexuses. Some evidence indicates that, as in mammals, the avian adrenal gland may also receive parasympathetic innervation, likely mediated by the vagus nerve through the celiac plexus [ref. 380]. Notably, adrenal blood vessels are innervated by both cholinergic and adrenergic terminals [ref. 382]. While synaptic endings are located close to adrenocortical cells, direct synaptic contacts are infrequent [ref. 383].
In chicks, the adrenal gland is surrounded by a dense fibrous connective tissue capsule, which is rich in collagen, reticular fibers, and blood vessels, but contains few elastic fibers [ref. 384,ref. 385]. The capsule’s thickness shows sexual dimorphism, as is significantly greater in females (22.09 ± 2.17 µm) than in males (18.14 ± 1.82 µm) [ref. 385]. Trabeculae extend from the capsule into the glandular parenchyma, contributing to its internal structure [ref. 384].
The avian adrenal parenchyma is composed of a combination of adrenocortical and chromaffin tissues. Chromaffin cells are predominantly concentrated in the central region of the gland, an area characterized by rich vascularization. Similar to in mammals, chromaffin cells are organized into small clusters of approximately 10–12. These cells are basophilic, polygonal or round, with basophilic cytoplasm and a large spherical nucleus containing two to three nucleoli [ref. 386]. Each cluster is typically innervated by a single nerve fiber bundle. However, in birds, these clusters do not merge to form the distinct adrenal medulla seen in mammals. A single nerve terminal may establish synaptic contact with up to three chromaffin cells of the same type-either NA or adrenaline (AD) secreting [ref. 381].
In broiler chicks, male adrenal cells exhibit slightly greater cell height (8.72 ± 0.231 µm) and nuclear diameter (4.39 ± 0.359 µm) compared with those from females (8.67 ± 0.218 and 3.84 ± 0.326 µm, respectively) [ref. 387]. The cytoplasm of these cells contains mitochondria with tubular cristae, ribosomes, endoplasmic reticulum, lipid droplets, and secretory granules [ref. 384,ref. 388]. Ganglionic cells are also observed interspersed between the medullary chromaffin cells [ref. 384]. In chick adrenal glands, approximately 70% of the chromaffin cells are AD type [ref. 389]. A consistent pattern has been observed in the spatial distribution of these cells: NA chromaffin cells are typically found at the edges of clusters, while AD cells are generally located toward the center [ref. 390,ref. 391].
Adrenocortical cells are organized into cords that extend outward from the gland’s center, forming frequent branches and connections. At the periphery, these cords loop along the inner side of the connective tissue capsule. Each cord comprises two rows of adrenocortical cells aligned with their long axes positioned perpendicular to the cord’s direction. Typically, though with some variations, the nuclei of these cells are eccentrically positioned near the cord’s shared, non-sinusoidal basal lamina [ref. 304]. The cellular heterogeneity-seen both microscopically and ultrastructurally-suggests the presence of a primitive form of zonation. Peripheral adrenocortical cells tend to be larger, often binucleated, and rich in lipid droplets, whereas those closer to the gland’s center are usually smaller, more elongated, and contain fewer lipids [ref. 392]. In birds, the structural organization of cortical (interrenal) and medullary (chromaffin) cell strands defines the zonation pattern within the adrenal parenchyma [ref. 323]. In hens, distinct peripheral (subcapsular) and central (internal) zones are observable upon adrenal gland sectioning [ref. 384]. Humayun et al. [ref. 393] also identified a subcapsular layer, as well as peripheral and central zones in the chick adrenal gland, consistent with similar observations in partridges [ref. 394] and quails [ref. 395].
10.2. Hormonal Influence on Tissues
The adrenocorticotropic axis is composed of CRH, ACTH, and CORT. CRH is synthesized by the PVN of the hypothalamus in response to environmental stressors [ref. 396], such as ambient temperature in broiler housing, stocking density, and limited access to water or feed. CRH release stimulates the anterior pituitary to secrete ACTH [ref. 138,ref. 141], which then promotes CORT production by the adrenal cortex [ref. 142]. This regulatory pathway is highly conserved across both mammals and avian species [ref. 397,ref. 398]. As the primary glucocorticoid in birds, CORT plays a fundamental role in triggering cellular mechanisms that mediate the stress response and modulating metabolic activity in reaction to stressors. Glucocorticoids are a diverse group of steroid hormones produced from cholesterol within the adrenal cortex [ref. 399]. While cortisol serves as the main circulating glucocorticoid in most mammals (with the exception of rodents), CORT fulfills this role in birds [ref. 400]. Due to its lipophilic nature, CORT does not circulate freely in the bloodstream; instead, it binds to corticosteroid-binding globulin (CBG), also known as transcortin [ref. 401], which is primarily synthesized in the liver [ref. 402]. In humans, CBG transports cortisol by enclosing it within a hydrophobic binding pocket [ref. 403,ref. 404], enabling its delivery to various target organs such as muscles, bones, liver, intestines, and kidneys [ref. 405,ref. 406]. Despite only a fifty percent similarity in amino acid composition at the steroid-binding site, avian CBG performs a comparable function to its mammalian counterpart [ref. 407].
10.3. Hormone-Mediated Regulatory Signaling
In the presence of an external stressor [ref. 137], the PVN of the hypothalamus releases CRH [ref. 139], which binds to specific CRH receptors (CRHRs). In chickens, two receptor subtypes have been identified: CRHR1 and CRHR2 [ref. 140,ref. 144]. Both are G protein-coupled receptors (GPCRs) that transmit CRH signals, although the latter has a notably higher binding affinity for CRH than the former [ref. 140]. Similar to GHRH receptors, CRHRs initiate intracellular signaling cascades, predominantly the cAMP/PKA pathway, which activates ACTH synthesis through the stepwise cleavage of the precursor proopiomelanocortin (POMC) by prohormone convertase 1/3 (PC1/3) [ref. 408,ref. 409,ref. 410]. It is also suggested that CRHRs may promote ACTH production by forming dimers with the vasotocin receptor subtype VT2R [ref. 411].
Following ACTH release from the anterior pituitary, it fulfills two primary roles: stimulating CORT secretion from the adrenal cortex and inhibiting further CRH release from the hypothalamus [ref. 412,ref. 413]. Similar to CRH, ACTH exerts its effects through a GPCR known as the melanocortin-2 receptor (MC2R) [ref. 414]. Although MC2R signaling mechanisms in chickens remain understudied, its structural similarity to mammalian MC2R implies a comparable function [ref. 415]. For functional activity, MC2R requires the presence of melanocortin receptor accessory proteins (MRAPs) [ref. 416], of which three essential ones have been identified in zebrafish and one in chickens [ref. 417,ref. 418]. These proteins facilitate MC2R transport from the endoplasmic reticulum to the plasma membrane, enabling ACTH signaling [ref. 419,ref. 420].
Corticosterone exerts its effects through the glucocorticoid receptor NR3C1, a ligand-dependent transcription factor. Upon reaching its target tissue, CORT diffuses into the cell and binds NR3C1 in the cytoplasm. At rest, NR3C1 is part of a chaperone complex including essential proteins HSP90 and HSP70, along with auxiliary components such as Hop, HSP40, and p23 [ref. 421]. Receptor inactivation is initiated by HSP70-induced partial unfolding, while reactivation depends on ATP hydrolysis by HSP90, a process regulated by histone deacetylase 6 (HDAC6) [ref. 422,ref. 423]. Upon ligand binding, NR3C1 dissociates from the chaperone complex and translocates into the nucleus, where it binds to glucocorticoid response elements (GREs) in regulatory regions of specific genes, leading to either transcriptional activation or repression depending on the tissue context [ref. 424,ref. 425]. In addition to its cellular effects, CORT contributes to a classical negative feedback loop by suppressing CRH and ACTH secretion at hypothalamus and anterior pituitary levels, respectively [ref. 426].
10.4. The Effect of Light on the Hypothalamic–Pituitary–Adrenal (HPA) Axis
Both within individual cells and across organ systems, circadian rhythms play a fundamental role in regulating numerous physiological activities by ensuring synchronized functioning between various biological systems. The adrenergic system is one such example, relying on the catecholamine neurotransmitters-adrenaline and noradrenaline-to perform essential biological tasks. The synthesis and release of these catecholamines are governed by both neural inputs from the sympathetic nervous system and hormonal signals involving the HPA axis. Notably, both regulatory pathways are influenced by signals from the central circadian clock, which contributes to the observed rhythmic fluctuations in catecholamine concentrations in the bloodstream and peripheral tissues. These oscillations can affect a range of target cells expressing adrenergic receptors, including those within the immune system [ref. 427].
The HPA axis is a central regulator of the body’s response to environmental stressors and external challenges [ref. 428]. Glucocorticoid hormones produced by the adrenal glands are essential for maintaining energy homeostasis in vertebrates [ref. 429]. These hormones enable the organism to respond to both routine environmental variations and sudden, acute stressors. As such, glucocorticoids serve as key modulators of phenotypic plasticity, allowing individuals to adapt to changing ecological conditions [ref. 430].
ALAN represents a relatively recent environmental stressor, the effects of which on HPA axis functioning are not yet fully understood. In particular, the extent to which ALAN influences the expression, localization, and sensitivity of glucocorticoid receptors (GRs) remains unclear. A thorough understanding of ALAN’s impact on the HPA axis requires an examination of both axis reactivity-meaning its activation and the resulting increase in circulating glucocorticoids (GCs) under stress-and the efficiency of its negative feedback regulation, through which GCs inhibit further hormone secretion by acting on GRs located in central structures such as the hypothalamus and anterior pituitary. It is possible that ALAN disrupts this feedback mechanism or alters GR expression or functionality, potentially leading to impaired glucocorticoid regulation and persistent HPA axis activation. Such dysregulation may have negative physiological and behavioral consequences. Therefore, only an integrated investigation of the entire HPA axis-from stress perception to hormone production and receptor-mediated feedback control-will allow for a comprehensive understanding of the mechanisms through which ALAN interferes with endocrine balance and for the development of effective mitigation strategies [ref. 24].
In chickens, as in other vertebrate species, the HPA axis functions as the primary system responsible for mediating long-term responses to stress. Upon activation, it stimulates the pituitary gland to secrete ACTH. However, inconsistent findings regarding the influence of light color on the physiological stress response have been reported in the scientific literature [ref. 259]. Prayitno et al. [ref. 5] and Hesham et al. [ref. 431] reported that broiler chickens exposed to red light exhibited increased aggressive behavior compared with those housed under blue, green, or white light. Additionally, Archer [ref. 432] demonstrated that exposing eggs to light, particularly white and red, during incubation subsequently reduced fear responses in chicks compared with incubation in darkness or under green light. Olanrewaju et al. [ref. 433,ref. 434] found no significant differences in corticosterone levels between birds raised under different LED light colors. Xie et al. [ref. 435] proposed that blue light might help mitigate stress responses in broilers, potentially through reducing serum interleukin-1β levels. Similarly, the findings of Helva [ref. 259] and Franco et al. [ref. 436] suggest that blue and green light may alleviate the negative effects of stress, supporting the conclusions drawn by Xie et al. [ref. 435]. Conversely, Mehaisen et al. [ref. 437] noted that glucocorticoid hormones suppress growth and feed intake. The elevated ACTH levels observed under white light conditions may be an additional factor contributing to reduced feed consumption in broilers exposed to this type of lighting.
11. Conclusions
Hypothalamic-pituitary-somatotropic (HPS) axis: Exposure to green LED light stimulates hypothalamic GHRH expression, pituitary GH secretion, and hepatic and muscular IGF-1 production, activating the JAK2/STAT5 and mTORC1 pathways that promote satellite cell proliferation and early muscle hypertrophy. Furthermore, melatonin receptors Mel1b and Mel1c participate in mediating the growth hormone secretion induced by green light in chicken adenohypophysis cells through the AC/PKA and ERK1/2 signaling pathways. In contrast, red light is associated with increased hypothalamic somatostatin expression and reduced pulsatile GH secretion, thereby limiting early muscle growth stimulation.
Hypothalamic-pituitary-gonadal (HPG) axis: The hypothalamus regulates reproductive function in birds by integrating hormonal and photic signals, maintaining the GnRH and GnIH balance that controls gonadotropin secretion. The activity of these neurohormones is influenced by melatonin, steroid and thyroid hormones, and environmental cues, ensuring reproductive cycle synchronization with seasonal conditions. Photoperiod and light quality play a crucial role: long days and red light stimulate hypothalamic and gonadal activity, while short days and melatonin inhibit reproduction. The interaction between endocrine and light-derived signals thus ensures precise hypothalamic-pituitary-gonadal axis regulation in birds.
Hypothalamic-pituitary-thyroid (HPT) axis: Variations in photoperiod and ALAN adjust TRH-TSH-T4/T3 cascades and deiodinase-mediated local T3 availability, modulating basal metabolism, seasonal reproductive cues, and tissue-specific thyroid hormone action through THRA/THRB signaling.
Hypothalamic-pituitary-adrenal (HPA) axis: Environmental stressors and the light spectrum interact in the CRH-ACTH-CORT feedback system: red light amplifies HPA activation and stress behaviors, whereas blue/green illumination mitigates corticosterone release and inflammation, indicating that spectral tuning can buffer stress responses.
In conclusion, exposure to green and blue light exerts a markedly superior enhancement of poultry productivity compared with red and white light. Furthermore, the controlled manipulation of LED spectral quality and exposure regimes provides a precise means of optimizing broiler growth, metabolic homeostasis, and stress resilience, thereby supporting sustainable and welfare-oriented poultry production practices.
References
- R. Parvin, M. Mushtaq, M. Kim, H. Choi. Light Emitting Diode (LED) as a Source of Monochromatic Light: A Novel Lighting Approach for Behaviour, Physiology and Welfare of Poultry. World Poult. Sci. J., 2014. [DOI]
- T.I. Pap, R.T. Szabó, Á. Bodnár, F. Pajor, I. Egerszegi, B. Podmaniczky, M. Pacz, D. Mezőszentgyörgyi, M. Kovács-Weber. Effect of Lighting Methods on the Production, Behavior and Meat Quality Parameters of Broiler Chickens. Animals, 2024. [DOI | PubMed]
- G. Luo, Y. Cheng, Y. Xu, J. Liu, W. Yang, J. Liu, B. Guo, H. Zhu. Monochromatic Light Impacts the Growth Performance, Intestinal Morphology, Barrier Function, Antioxidant Status, and Microflora of Yangzhou Geese. Animals, 2025. [DOI | PubMed]
- C. Wathes, H. Spechter, T. Bray. The Effects of Light Illuminance and Wavelength on the Growth of Broiler Chickens. J. Agric. Sci., 1982. [DOI]
- D. Prayitno, C. Phillips, H. Omed. The Effects of Color of Lighting on the Behavior and Production of Meat Chickens. Poult. Sci., 1997. [DOI | PubMed]
- I. Rozenboim, I. Biran, Z. Uni, B. Robinzon, O. Halevy. The Effect of Monochromatic Light on Broiler Growth and Development. Poult. Sci., 1999. [DOI | PubMed]
- J. Cao, W. Liu, Z. Wang, D. Xie, L. Jia, Y. Chen. Green and Blue Monochromatic Lights Promote Growth and Development of Broilers via Stimulating Testosterone Secretion and Myofiber Growth. J. Appl. Poult. Res., 2008. [DOI]
- Y. Ke, W. Liu, Z. Wang, Y. Chen. Effects of Monochromatic Light on Quality Properties and Antioxidation of Meat in Broilers. Poult. Sci., 2011. [DOI | PubMed]
- R. Mohamed, M. Eltholth, N. El-Saidy. Rearing Broiler Chickens under Monochromatic Blue Light Improve Performance and Reduce Fear and Stress during Pre-Slaughter Handling and Transportation. Biotechnol. Anim. Husb., 2014. [DOI]
- F.N. Soliman, K. El-Sabrout. Light Wavelengths/Colors: Future Prospects for Broiler Behavior and Production. J. Vet. Behav., 2020. [DOI]
- E.A.-M. Abdel-Moneim, S.A. Siddiqui, A.M. Shehata, A. Biswas, M.S. Abougabal, A.M. Kamal, N.M. Mesalam, M.A. Elsayed, B. Yang, T.A. Ebeid. Impact of lIght Wavelength on Growth and Welfare of broIler chIckens–overvIew and Future perspectIve. Ann. Anim. Sci., 2024. [DOI]
- I. Rozenboim, M. El Halawani, Y. Kashash, Y. Piestun, O. Halevy. The Effect of Monochromatic Photostimulation on Growth and Development of Broiler Birds. Gen. Comp. Endocrinol., 2013. [DOI | PubMed]
- S. Gharahveysi, M. Irani, T.A. Kenari, K.I. Mahmud. Effects of Colour and Intensity of Artificial Light Produced by Incandescent Bulbs on the Performance Traits, Thyroid Hormones, and Blood Metabolites of Broiler Chickens. Ital. J. Anim. Sci., 2020. [DOI]
- E. Jin, L. Jia, J. Li, G. Yang, Z. Wang, J. Cao, Y. Chen. Effect of Monochromatic Light on Melatonin Secretion and Arylalkylamine N-acetyltransferase mRNA Expression in the Retina and Pineal Gland of Broilers. Anat. Rec. Adv. Integr. Anat. Evol. Biol., 2011. [DOI]
- K. Hayat, R. Zheng, L. Zeng, Z. Ye, J. Pan. Impact of Full-Spectrum and Infrared Lighting on Growth, Oxidative Stress, and Cecal Microbiota in Broilers. Antioxidants, 2024. [DOI | PubMed]
- Y. Yang, Y. Yu, J. Pan, Y. Ying, H. Zhou. A New Method to Manipulate Broiler Chicken Growth and Metabolism: Response to Mixed LED Light System. Sci. Rep., 2016. [DOI | PubMed]
- O. Halevy, I. Biran, I. Rozenboim. Various Light Source Treatments Affect Body and Skeletal Muscle Growth by Affecting Skeletal Muscle Satellite Cell Proliferation in Broilers. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol., 1998. [DOI]
- W. Liu, Z. Wang, Y. Chen. Effects of Monochromatic Light on Developmental Changes in Satellite Cell Population of Pectoral Muscle in Broilers during Early Posthatch Period. Anat. Rec. Adv. Integr. Anat. Evol. Biol., 2010. [DOI]
- L. Zhang, J. Cao, Z. Wang, Y. Dong, Y. Chen. Melatonin Modulates Monochromatic Light-Induced GHRH Expression in the Hypothalamus and GH Secretion in Chicks. Acta Histochem., 2016. [DOI | PubMed]
- A.R.N. Reyad, O.A. Mohamed. A Comparative Study of The Effect of White And Blue-Green LED Light Wavelengths On Growth Performance, Feed Efficiency, And Behavioural Traits In Cobb Broiler Chickens. Egypt. Poult. Sci. J., 2024. [DOI]
- H. Olanrewaju, W. Miller, W. Maslin, S. Collier, J. Purswell, S. Branton. Effects of Light Sources and Intensity on Broilers Grown to Heavy Weights. Part 1: Growth Performance, Carcass Characteristics, and Welfare Indices. Poult. Sci., 2016. [DOI | PubMed]
- J.R. Nelson, J.L. Bray, J. Delabbio, G.S. Archer. Light Emitting Diode (LED) Color and Broiler Growth: Effect of Supplementing Blue/Green LED to White LED Light on Broiler Growth, Stress, and Welfare. Poult. Sci., 2020. [DOI | PubMed]
- K.L. Russart, R.J. Nelson. Light at Night as an Environmental Endocrine Disruptor. Physiol. Behav., 2018. [DOI | PubMed]
- J.Q. Ouyang, S. Davies, D. Dominoni. Hormonally Mediated Effects of Artificial Light at Night on Behavior and Fitness: Linking Endocrine Mechanisms with Function. J. Exp. Biol., 2018. [DOI | PubMed]
- J. Falcón, A. Torriglia, D. Attia, F. Viénot, C. Gronfier, F. Behar-Cohen, C. Martinsons, D. Hicks. Exposure to Artificial Light at Night and the Consequences for Flora, Fauna, and Ecosystems. Front. Neurosci., 2020. [DOI | PubMed]
- M.L. Grunst, A.S. Grunst. Endocrine Effects of Exposure to Artificial Light at Night: A Review and Synthesis of Knowledge Gaps. Mol. Cell. Endocrinol., 2023. [DOI | PubMed]
- H.C. Atkinson, B.J. Waddell. Circadian Variation in Basal Plasma Corticosterone and Adrenocorticotropin in the Rat: Sexual Dimorphism and Changes across the Estrous Cycle. Endocrinology, 1997. [DOI | PubMed]
- P. Guchhait, C. Haldar. Circadian Rhythms of Melatonin and Sex Steroids in a Nocturnal Bird, Indian Spotted Owlet Athene Brama during Reproductively Active and Inactive Phases. Biol. Rhythm Res., 1999. [DOI]
- S. Li, J. Cao, Z. Wang, Y. Dong, W. Wang, Y. Chen. Melatonin Mediates Monochromatic Light-induced Insulin-like Growth Factor 1 Secretion of Chick Liver: Involvement of Membrane Receptors. Photochem. Photobiol., 2016. [DOI | PubMed]
- S. Çalişlar, B. Yeter, A. Şahin. Importance of Melatonin on Poultry. Tarim Doga Derg., 2018. [DOI]
- H.E. Chmura, J.C. Wingfield, T.P. Hahn. Non-photic Environmental Cues and Avian Reproduction in an Era of Global Change. J. Avian Biol., 2020. [DOI]
- A.A. Hussein, E. Bloem, I. Fodor, E.-S. Baz, M.M. Tadros, M.F. Soliman, N.S. El-Shenawy, J.M. Koene. Slowly Seeing the Light: An Integrative Review on Ecological Light Pollution as a Potential Threat for Mollusks. Environ. Sci. Pollut. Res., 2021. [DOI | PubMed]
- S. Wilson, F. Cunningham. Effect of Increasing Day Length and Intermittent Lighting Schedules in the Domestic Hen on Plasma Concentrations of Luteinizing Hormone (LH) and the LH Response to Exogenous Progesterone. Gen. Comp. Endocrinol., 1980. [DOI | PubMed]
- C.E. Manser, H. Elliott, T. Morris, D. Broom. The Use of a Novel Operant Test to Determine the Strength of Preference for Flooring in Laboratory Rats. Lab. Anim., 1996. [DOI | PubMed]
- A. Deep, K. Schwean-Lardner, T. Crowe, B. Fancher, H. Classen. Effect of Light Intensity on Broiler Production, Processing Characteristics, and Welfare. Poult. Sci., 2010. [DOI | PubMed]
- A.M. Fernandes, K. Fero, W. Driever, H.A. Burgess. Enlightening the Brain: Linking Deep Brain Photoreception with Behavior and Physiology. Bioessays, 2013. [DOI | PubMed]
- A.E. Aulsebrook, R.D. Johnsson, J.A. Lesku. Light, Sleep and Performance in Diurnal Birds. Clocks Sleep, 2021. [DOI | PubMed]
- D. ViviD, G.E. Bentley. Seasonal Reproduction in Vertebrates: Melatonin Synthesis, Binding, and Functionality Using Tinbergen’s Four Questions. Molecules, 2018. [DOI | PubMed]
- S. Collins, B. Forkman, H.H. Kristensen, P. Sandøe, P.M. Hocking. Investigating the Importance of Vision in Poultry: Comparing the Behaviour of Blind and Sighted Chickens. Appl. Anim. Behav. Sci., 2011. [DOI]
- A. Dawson, V.M. King, G.E. Bentley, G.F. Ball. Photoperiodic Control of Seasonality in Birds. J. Biol. Rhythm., 2001. [DOI]
- M. Baxter, N. Joseph, V. Osborne, G. Bédécarrats. Red Light Is Necessary to Activate the Reproductive Axis in Chickens Independently of the Retina of the Eye. Poult. Sci., 2014. [DOI | PubMed]
- Y. Wu, J. Huang, S. Quan, Y. Yang. Light Regimen on Health and Growth of Broilers: An Update Review. Poult. Sci., 2022. [DOI | PubMed]
- J.K. Bowmaker, A. Knowles. The Visual Pigments and Oil Droplets of the Chicken Retina. Vis. Res., 1977. [DOI | PubMed]
- Y.A. Kram, S. Mantey, J.C. Corbo. Avian Cone Photoreceptors Tile the Retina as Five Independent, Self-Organizing Mosaics. PLoS ONE, 2010. [DOI | PubMed]
- D.A. Golombek, R.E. Rosenstein. Physiology of Circadian Entrainment. Physiol. Rev., 2010. [DOI | PubMed]
- S. Halford, S.S. Pires, M. Turton, L. Zheng, I. González-Menéndez, W.L. Davies, S.N. Peirson, J.M. García-Fernández, M.W. Hankins, R.G. Foster. VA Opsin-Based Photoreceptors in the Hypothalamus of Birds. Curr. Biol., 2009. [DOI | PubMed]
- W.J. Kuenzel, S.W. Kang, Z.J. Zhou. Exploring Avian Deep-Brain Photoreceptors and Their Role in Activating the Neuroendocrine Regulation of Gonadal Development. Poult. Sci., 2015. [DOI | PubMed]
- G. Wang, J.C. Wingfield. Immunocytochemical Study of Rhodopsin-Containing Putative Encephalic Photoreceptors in House Sparrow, Passer domesticus. Gen. Comp. Endocrinol., 2011. [DOI | PubMed]
- Y. Nakane, T. Yoshimura. Deep Brain Photoreceptors and a Seasonal Signal Transduction Cascade in Birds. Cell Tissue Res., 2010. [DOI | PubMed]
- S.W. Kang. Central Nervous System Associated with Light Perception and Physiological Responses of Birds. Front. Physiol., 2021. [DOI | PubMed]
- W.I. Davies, M. Turton, S.N. Peirson, B.K. Follett, S. Halford, J.M. Garcia-Fernandez, P.J. Sharp, M.W. Hankins, R.G. Foster. Vertebrate Ancient Opsin Photopigment Spectra and the Avian Photoperiodic Response. Biol. Lett., 2012. [DOI | PubMed]
- M. Menaker, H. Keatts. Extraretinal Light Perception in the Sparrow. II. Photoperiodic Stimulation of Testis Growth. Proc. Natl. Acad. Sci. USA, 1968. [DOI | PubMed]
- Y. Nakane, K. Ikegami, H. Ono, N. Yamamoto, S. Yoshida, K. Hirunagi, S. Ebihara, Y. Kubo, T. Yoshimura. A Mammalian Neural Tissue Opsin (Opsin 5) Is a Deep Brain Photoreceptor in Birds. Proc. Natl. Acad. Sci. USA, 2010. [DOI | PubMed]
- D. Pan, Z. Wang, Y. Chen, J. Cao. Melanopsin-Mediated Optical Entrainment Regulates Circadian Rhythms in Vertebrates. Commun. Biol., 2023. [DOI | PubMed]
- M.W. Hankins, W.I. Davies, R.G. Foster. The Evolution of Non-Visual Photopigments in the Central Nervous System of Vertebrates. Evolution of Visual and Non-Visual Pigments, 2014
- R. Foster, B. Follett, J. Lythgoe. Rhodopsin-like Sensitivity of Extra-Retinal Photoreceptors Mediating the Photoperiodic Response in Quail. Nature, 1985. [DOI | PubMed]
- H. Hartwig, T. Van Veen. Spectral Characteristics of Visible Radiation Penetrating into the Brain and Stimulating Extraretinal Photoreceptors: Transmission Recordings in Vertebrates. J. Comp. Physiol., 1979. [DOI]
- S. Yasuo, M. Watanabe, N. Okabayashi, S. Ebihara, T. Yoshimura. Circadian Clock Genes and Photoperiodism: Comprehensive Analysis of Clock Gene Expression in the Mediobasal Hypothalamus, the Suprachiasmatic Nucleus, and the Pineal Gland of Japanese Quail under Various Light Schedules. Endocrinology, 2003. [DOI | PubMed]
- B. Follett, T. Nicholls. Influences of Thyroidectomy and Thyroxine Replacement on Photoperiodically Controlled Reproduction in Quail. J. Endocrinol., 1985. [DOI | PubMed]
- T. Yoshimura, S. Yasuo, M. Watanabe, M. Iigo, T. Yamamura, K. Hirunagi, S. Ebihara. Light-Induced Hormone Conversion of T4 to T3 Regulates Photoperiodic Response of Gonads in Birds. Nature, 2003. [DOI | PubMed]
- V.M. Cassone, T. Yoshimura. Circannual Cycles and Photoperiodism. Sturkie’s Avian Physiology, 2022
- J. Xu, L.E. Jarocha, T. Zollitsch, M. Konowalczyk, K.B. Henbest, S. Richert, M.J. Golesworthy, J. Schmidt, V. Déjean, D.J. Sowood. Magnetic Sensitivity of Cryptochrome 4 from a Migratory Songbird. Nature, 2021. [DOI | PubMed]
- N. Nader, G.P. Chrousos, T. Kino. Interactions of the Circadian CLOCK System and the HPA Axis. Trends Endocrinol. Metab., 2010. [DOI | PubMed]
- N. Cermakian, T. Lange, D. Golombek, D. Sarkar, A. Nakao, S. Shibata, G. Mazzoccoli. Crosstalk between the Circadian Clock Circuitry and the Immune System. Chronobiol. Int., 2013. [DOI | PubMed]
- M. Markowska, P.M. Majewski, K. Skwarło-Sońta. Avian Biological Clock–Immune System Relationship. Dev. Comp. Immunol., 2017. [DOI | PubMed]
- V. Jerigova, M. Zeman, M. Okuliarova. Circadian Disruption and Consequences on Innate Immunity and Inflammatory Response. Int. J. Mol. Sci., 2022. [DOI | PubMed]
- I. Mishra, D. Singh, V. Kumar. Temporal Expression of C-Fos and Genes Coding for Neuropeptides and Enzymes of Amino Acid and Amine Neurotransmitter Biosynthesis in Retina, Pineal and Hypothalamus of a Migratory Songbird: Evidence for Circadian Rhythm-Dependent Seasonal Responses. Neuroscience, 2018. [DOI | PubMed]
- A. Natesan, L. Geetha, M. Zatz. Rhythm and Soul in the Avian Pineal. Cell Tissue Res., 2002. [DOI | PubMed]
- S. Ma, Z. Wang, J. Cao, Y. Dong, Y. Chen. BMAL1 but Not CLOCK Is Associated with Monochromatic Green Light-Induced Circadian Rhythm of Melatonin in Chick Pinealocytes. Endocr. Connect., 2019. [DOI | PubMed]
- M.C. Beker, B. Caglayan, A.B. Caglayan, T. Kelestemur, E. Yalcin, A. Caglayan, U. Kilic, A.T. Baykal, R.J. Reiter, E. Kilic. Interaction of Melatonin and Bmal1 in the Regulation of PI3K/AKT Pathway Components and Cellular Survival. Sci. Rep., 2019. [DOI | PubMed]
- N. Gotlieb, J. Moeller, L.J. Kriegsfeld. Circadian Control of Neuroendocrine Function: Implications for Health and Disease. Curr. Opin. Physiol., 2018. [DOI | PubMed]
- E. Rich, L. Romero. Daily and Photoperiod Variations of Basal and Stress-Induced Corticosterone Concentrations in House Sparrows (Passer domesticus). J. Comp. Physiol. B, 2001. [PubMed]
- P. Schwabl, E. Bonaccorso, W. Goymann. Diurnal Variation in Corticosterone Release among Wild Tropical Forest Birds. Front. Zool., 2016. [DOI | PubMed]
- N.P. Huffeldt, F.R. Merkel, S. Jenni-Eiermann, W. Goymann, B. Helm. Melatonin and Corticosterone Profiles under Polar Day in a Seabird with Sexually Opposite Activity-Rhythms. Gen. Comp. Endocrinol., 2020. [DOI | PubMed]
- T. Greives, M. Eshleman, H. Galante, E. Elderbrock, C. Deimel, M. Hau. Early Nighttime Testosterone Peaks Are Correlated with GnRH-Induced Testosterone in a Diurnal Songbird. Gen. Comp. Endocrinol., 2021. [DOI | PubMed]
- J.L. Graham, K.B. Needham, E.M. Bertucci, A.A. Pearson, C.M. Bauer, T.J. Greives. Onset of Daily Activity in a Female Songbird Is Related to Peak-Induced Estradiol Levels. Integr. Comp. Biol., 2019. [DOI | PubMed]
- E.K. Elderbrock, M. Hau, T.J. Greives. Sex Steroids Modulate Circadian Behavioral Rhythms in Captive Animals, but Does This Matter in the Wild?. Horm. Behav., 2021. [DOI | PubMed]
- F. Gwinner. Testosterone Induces” Splitting” of Circadian Locomotor Activity Rhythms in Birds. Science, 1974. [DOI | PubMed]
- W. Quay. Histological Structure and Cytology of the Pineal Organ in Birds and Mammals. Prog. Brain Res., 1965. [PubMed]
- Z. Fejér, P. Röhlich, Á. Szél, C. Dávid, A. Zádori, M. João Manzano, B. Vígh. Comparative Ultrastructure and Cytochemistry of the Avian Pineal Organ. Microsc. Res. Tech., 2001. [DOI | PubMed]
- C. Haldar, K. Bishnupuri. Comparative View of Pineal Gland Morphology of Nocturnal and Diurnal Birds of Tropical Origin. Microsc. Res. Tech., 2001. [DOI | PubMed]
- B. Przybylska-Gornowicz, B. Lewczuk, M. Prusik, M. Nowicki. Post-Hatching Development of the Turkey Pineal Organ: Histological and Immunohistochemical Studies. Neuroendocrinol. Lett., 2005. [PubMed]
- M. Prusik, B. Lewczuk, M. Nowicki, B. Przybylska-Gornowicz. Histology and Ultrastructure of the Pineal Organ in the Domestic Goose. Histol. Histopathol., 2006. [PubMed]
- T. Sato, K. Wake. Innervation of the Avian Pineal Organ: A Comparative Study. Cell Tissue Res., 1983. [DOI | PubMed]
- H.-W. Korf, N.H. Zimmerman, A. Oksche. Intrinsic Neurons and Neural Connections of the Pineal Organ of the House Sparrow, Passer domesticus, as Revealed by Anterograde and Retrograde Transport of Horseradish Peroxidase. Cell Tissue Res., 1982. [DOI | PubMed]
- V.J. Csernus. The Avian Pineal Gland. Chronobiol. Int., 2006. [DOI | PubMed]
- T. Okano, T. Yoshizawa, Y. Fukada. Pinopsin Is a Chicken Pineal Photoreceptive Molecule. Nature, 1994. [DOI | PubMed]
- L.M. Robertson, J.S. Takahashi. Circadian Clock in Cell Culture: II. In Vitro Photic Entrainment of Melatonin Oscillation from Dissociated Chick Pineal Cells. J. Neurosci., 1988. [DOI | PubMed]
- J.S. Takahashi, N. Murakami, S.S. Nikaido, B.L. Pratt, L.M. Robertson. The Avian Pineal, a Vertebrate Model System of the Circadian Oscillator: Cellular Regulation of Circadian Rhythms by Light, Second Messengers, and Macromolecular Synthesis. Proceedings of the 1988 Laurentian Hormone Conference, 1989
- M. Prusik. Developmental Morphology of the Turkey Pineal Gland in Histological Images and 3D Models. Micron, 2022. [DOI | PubMed]
- I. Olah, B. Glick. Lymphopineal Tissue in the Chicken. Dev. Comp. Immunol., 1984. [DOI | PubMed]
- O. Doi, A. Ohno, T. Kaneda, A. Iwasawa, T. Nakamura. Immunohistochemical Demonstration and Daily Rhythms of Melatonin in the Chicken Pineal Gland. Jpn. Poult. Sci., 1995. [DOI]
- C. Timothy, N. Birdsall. The Biological Effects and Clinical Uses of the Pineal Hormone Melatonin. Altern. Med. Rev., 1996
- J. Barrenetxe, P. Delagrange, J. Martinez. Physiological and Metabolic Functions of Melatonin. J. Physiol. Biochem., 2004. [DOI | PubMed]
- N. Ziółkowska, M. Prusik, B. Lewczuk, B. Przybylska-Gornowicz. Regulation of Melatonin Secretion in the Pineal Organ of the Domestic Duck–an in Vitro Study. Pol. J. Vet. Sci., 2015. [PubMed]
- V.O. Sinkalu, J.O. Ayo, A.A. Abimbola, J.E. Ibrahim. Effects of Melatonin on Cloacal Temperature and Erythrocyte Osmotic Fragility in Layer Hens during the Hot-Dry Season. J. Appl. Anim. Res., 2015. [DOI]
- G. Şener. Karanlığın Hormonu: Melatonin. Marmara Pharm. J., 2010. [DOI]
- G.A. Bubenik. Gastrointestinal Melatonin: Localization, Function, and Clinical Relevance. Dig. Dis. Sci., 2002. [DOI | PubMed]
- V.M. Cassone, D.S. Brooks, T.A. Kelm. Comparative Distribution of 2 [125I] Iodomelatonin Binding in the Brains of Diurnal Birds: Outgroup Analysis with Turtles. Brain Behav. Evol., 1995. [DOI | PubMed]
- P.P. Lee, S.F. Pang. Identification and Characterization of Melatonin Binding Sites in the Gastrointestinal Tract of Ducks. Life Sci., 1992. [DOI | PubMed]
- L. Dishon, N. Avital-Cohen, D. Malamud, R. Heiblum, S. Druyan, T. Porter, M. Gumułka, I. Rozenboim. In-Ovo Monochromatic Green Light Photostimulation Enhances Embryonic Somatotropic Axis Activity. Poult. Sci., 2017. [DOI | PubMed]
- Y. Xu, Y. Tang, Y. Cheng, W. Yang, J. Liu, B. Guo, G. Luo, H. Zhu. Effects of Different Monochromatic Light on Growth Performance and Liver Circadian Rhythm of Yangzhou Geese. Poult. Sci., 2025. [DOI | PubMed]
- L. Yue, X. Qin, X. Liu, Z. Wang, Y. Dong, Y. Chen, J. Cao. Melatonin Receptor Mel1b-and Mel1c-mediated Green Light Induced the Secretion of Growth Hormone in Anterior Pituitary of Chicks. Photochem. Photobiol., 2019. [DOI | PubMed]
- X. Liu, L. Wang, Z. Wang, Y. Dong, Y. Chen, J. Cao. Mel1b and Mel1c Melatonin Receptors Mediate Green Light-Induced Secretion of Growth Hormone in Chick Adenohypophysis Cells via the AC/PKA and ERK1/2 Signalling Pathways. J. Photochem. Photobiol. B Biol., 2021. [DOI]
- R. Lazăr, C. Solcan, C. Creţu, M. Lazăr, C. Muntean, P. Boişteanu. Characterization of the Relations between Morphology and Physiological Status of the Pineal Gland in Connection with the Somatic Development Level in Turkeys Reared in Romania. Arq. Bras. Med. Veterinária Zootec., 2015. [DOI]
- W. Clark, H. Classen. The Effects of Continuously or Diurnally Fed Melatonin on Broiler Performance and Health. Poult. Sci., 1995. [DOI | PubMed]
- M. Zeman, P. Výboh, M. Jurani, D. Lamosova, L. Kostal, B. Bilcik, P. Blazicek, E. Juraniova. Effects of Exogenous Melatonin on Some Endocrine, Behavioural and Metabolic Parameters in Japanese Quail Coturnix coturnix japonica. Comp. Biochem. Physiol. Part A Physiol., 1993. [DOI | PubMed]
- M. Zeman, J. Buyse, D. Lamosova, I. Herichova, E. Decuypere. Role of Melatonin in the Control of Growth and Growth Hormone Secretion in Poultry. Domest. Anim. Endocrinol., 1999. [DOI | PubMed]
- S. Harvey, E. Decuypere, V. Darras, L. Berghman. Differential Effects of T4 and T3 on TRH and GRF-Induced GH Secretion in the Domestic Fowl. Reprod. Nutr. Dev., 1991. [DOI | PubMed]
- L. Horodincu, C. Solcan. Influence of Different Light Spectra on Melatonin Synthesis by the Pineal Gland and Influence on the Immune System in Chickens. Animals, 2023. [DOI | PubMed]
- L. Zheng, Y. Ma, L. Gu, D. Yuan, M. Shi, X. Guo, X. Zhan. Growth Performance, Antioxidant Status, and Nonspecific Immunity in Broilers under Different Lighting Regimens. J. Appl. Poult. Res., 2013. [DOI]
- G.A. Bubenik. Localization, Physiological Significance and Possible Clinical Implication of Gastrointestinal Melatonin. Neurosignals, 2001. [DOI]
- H. Ahmed, G. Essawy, H. Salem, M. Abdel Daim. Melatonin Has a Strong Antioxidant Activity and Improves Liver and Kidney Functions in Broiler Chicks. Egypt. J. Basic Appl. Physiol., 2005
- N. Sahin, M. Onderci, K. Sahin, M. Gursu, M. Smith. Ascorbic Acid and Melatonin Reduce Heat-Induced Performance Inhibition and Oxidative Stress in Japanese Quails. Br. Poult. Sci., 2004. [DOI | PubMed]
- Y. Ohta, M. Kongo, E. Sasaki, K. Nishida, I. Ishiguro. Therapeutic Effect of Melatonin on Carbon Tetrachloride-induced Acute Liver Injury in Rats. J. Pineal Res., 2000. [DOI | PubMed]
- Z. Kankova, A. Drozdova, V. Hodova, M. Zeman. Effect of Blue and Red Monochromatic Light during Incubation on the Early Post-Embryonic Development of Immune Responses in Broiler Chicken. Br. Poult. Sci., 2022. [DOI | PubMed]
- R. Brandstätter, U. Abraham. Hypothalamic Circadian Organization in Birds. I. Anatomy, Functional Morphology, and Terminology of the Suprachiasmatic Region. Chronobiol. Int., 2003. [DOI | PubMed]
- C.A. Pearson, M. Placzek. Development of the Medial Hypothalamus: Forming a Functional Hypothalamic-Neurohypophyseal Interface. Curr. Top. Dev. Biol., 2013. [PubMed]
- Y. Machluf, A. Gutnick, G. Levkowitz. Development of the Zebrafish Hypothalamus. Ann. N. Y. Acad. Sci., 2011. [DOI | PubMed]
- L. Zhang, F. Chen, J. Cao, Y. Dong, Z. Wang, Y. Chen. Melatonin Modulates Monochromatic Light-Induced Melatonin Receptor Expression in the Hypothalamus of Chicks. Acta Histochem., 2017. [DOI | PubMed]
- J.E. Levine. An Introduction to Neuroendocrine Systems. Handbook of Neuroendocrinology, 2012
- J. Schulkin. Evolutionary Conservation of Glucocorticoids and Corticotropin Releasing Hormone: Behavioral and Physiological Adaptations. Brain Res., 2011. [DOI | PubMed]
- K. Ikegami, T. Yoshimura. The Hypothalamic–Pituitary–Thyroid Axis and Biological Rhythms: The Discovery of TSH’s Unexpected Role Using Animal Models. Best Pract. Res. Clin. Endocrinol. Metab., 2017. [DOI | PubMed]
- R.G. Clark, L.C. Robinson. Up and down the Growth Hormone Cascade. Cytokine Growth Factor Rev., 1996. [DOI | PubMed]
- M.D. Gahete, R.M. Luque, J.P. Castaño. Models of GH Deficiency in Animal Studies. Best Pract. Res. Clin. Endocrinol. Metab., 2016. [DOI | PubMed]
- M.L. Vance. Growth-Hormone-Releasing Hormone. Clin. Chem., 1990. [DOI | PubMed]
- G.S. Tannenbaum, J.-C. Painson, M. Lapointe, W. Gurd, G.F. McCarthy. Interplay of Somatostatin and Growth Hormone-Releasing Hormone in Genesis of Episodic Growth Hormone Secretion. Metabolism, 1990. [DOI | PubMed]
- C. Tuggle, A. Trenkle. Control of Growth Hormone Synthesis. Domest. Anim. Endocrinol., 1996. [DOI | PubMed]
- A.A. Toogood, S. Harvey, M.O. Thorner, B.D. Gaylinn. Cloning of the Chicken Pituitary Receptor for Growth Hormone-Releasing Hormone. Endocrinology, 2006. [DOI | PubMed]
- S. Harvey, C. Gineste, B. Gaylinn. Growth Hormone (GH)-Releasing Activity of Chicken GH-Releasing Hormone (GHRH) in Chickens. Gen. Comp. Endocrinol., 2014. [DOI | PubMed]
- Y. Wang, J. Li, C.Y. Wang, A.H.Y. Kwok, F.C. Leung. Identification of the Endogenous Ligands for Chicken Growth Hormone-Releasing Hormone (GHRH) Receptor: Evidence for a Separate Gene Encoding GHRH in Submammalian Vertebrates. Endocrinology, 2007. [DOI | PubMed]
- J. Woelfle, D.J. Chia, M.B. Massart-Schlesinger, P. Moyano, P. Rotwein. Molecular Physiology, Pathology, and Regulation of the Growth Hormone/Insulin-like Growth Factor-I System. Pediatr. Nephrol., 2005. [DOI | PubMed]
- E. Dewil, V.M. Darras, G.S.G. Spencer, T.J. Lauterio, E. Decuypere. The Regulation of GH-Dependent Hormones and Enzymes after Feed Restriction in Dwarf and Control Chickens. Life Sci., 1999. [DOI | PubMed]
- C.E. Stewart, P. Rotwein. Growth, Differentiation, and Survival: Multiple Physiological Functions for Insulin-like Growth Factors. Physiol. Rev., 1996. [DOI | PubMed]
- D. Armstrong, C. McKay, D. Morrell, C. Goddard. Insulin-like Growth Factor-I Binding Proteins in Serum from the Domestic Fowl. J. Endocrinol., 1989. [DOI | PubMed]
- B. Lu, W. Liang, C. Liang, Z. Yu, X. Xie, Z. Chen. Effect of Heat Stress on Expression of Main Reproductive Hormone in Hypothalamic-Pituitary-Gonadal Axis of Wenchang Chicks. Braz. J. Poult. Sci., 2021. [DOI]
- A.E. Calogero, R. Bernardini, A.N. Margioris, G. Bagdy, W.T. Gallucci, P.J. Munson, L. Tamarkin, T.P. Tomai, L. Brady, P.W. Gold. Effects of Serotonergic Agonists and Antagonists on Corticotropin-Releasing Hormone Secretion by Explanted Rat Hypothalami. Peptides, 1989. [DOI | PubMed]
- H. Hayashi, K. Imai, K. Imai. Characterization of Chicken ACTH and α-MSH: The Primary Sequence of Chicken ACTH Is More Similar to Xenopus ACTH than to Other Avian ACTH. Gen. Comp. Endocrinol., 1991. [DOI | PubMed]
- M.H. Whitnall. Regulation of the Hypothalamic Corticotropin-Releasing Hormone Neurosecretory System. Prog. Neurobiol., 1993. [DOI | PubMed]
- J. Yu, L. Xie, A. Abou-Samra. Molecular Cloning of a Type A Chicken Corticotropin-Releasing Factor Receptor with High Affinity for Urotensin I. Endocrinology, 1996. [DOI | PubMed]
- E. Webster, D. Lewis, D. Torpy, E. Zachman, K. Rice, G. Chrousos. In Vivo and in Vitro Characterization of Antalarmin, a Nonpeptide Corticotropin-Releasing Hormone (CRH) Receptor Antagonist: Suppression of Pituitary ACTH Release and Peripheral Inflammation. Endocrinology, 1996. [DOI | PubMed]
- M.A. Latour, S. Laiche, J. Thompson, A. Pond, E.D. Peebles. Continuous Infusion of Adrenocorticotropin Elevates Circulating Lipoprotein Cholesterol and Corticosterone Concentrations in Chickens. Poult. Sci., 1996. [DOI | PubMed]
- B. De Groef, K. Geris, J. Manzano, J. Bernal, R. Millar, A.-B. Abou-Samra, T. Porter, A. Iwasawa, E. Kühn, V. Darras. Involvement of Thyrotropin-Releasing Hormone Receptor, Somatostatin Receptor Subtype 2 and Corticotropin-Releasing Hormone Receptor Type 1 in the Control of Chicken Thyrotropin Secretion. Mol. Cell. Endocrinol., 2003. [DOI | PubMed]
- B. De Groef, S.V. Grommen, I. Mertens, L. Schoofs, E.R. Kühn, V.M. Darras. Cloning and Tissue Distribution of the Chicken Type 2 Corticotropin-Releasing Hormone Receptor. Gen. Comp. Endocrinol., 2004. [DOI | PubMed]
- T. Taylor, F. Wondisford, T. Blaine, B. Weintraub. The Paraventricular Nucleus of the Hypothalamus Has a Major Role in Thyroid Hormone Feedback Regulation of Thyrotropin Synthesis and Secretion. Endocrinology, 1990. [DOI | PubMed]
- M.W. Szkudlinski, V. Fremont, C. Ronin, B.D. Weintraub. Thyroid-Stimulating Hormone and Thyroid-Stimulating Hormone Receptor Structure-Function Relationships. Physiol. Rev., 2002. [DOI | PubMed]
- Y. Sun, X. Lu, M. Gershengorn. G-Protein-Coupled Receptor Signaling in Neuroendocrine Systems. Thyrotropin-Releasing Hormone Receptors—Similarities and Differences. J. Mol. Endocrinol., 2003. [DOI | PubMed]
- S. Sam, L.A. Frohman. Normal Physiology of Hypothalamic Pituitary Regulation. Endocrinol. Metab. Clin. N. Am., 2008. [DOI]
- B. De Groef, S. Grommen, V. Darras. The Chicken Embryo as a Model for Developmental Endocrinology: Development of the Thyrotropic, Corticotropic, and Somatotropic Axes. Mol. Cell. Endocrinol., 2008. [DOI | PubMed]
- E.A. Nillni. Regulation of the Hypothalamic Thyrotropin Releasing Hormone (TRH) Neuron by Neuronal and Peripheral Inputs. Front. Neuroendocrinol., 2010. [DOI | PubMed]
- M. Grossmann, B.D. Weintraub, M.W. Szkudlinski. Novel Insights into the Molecular Mechanisms of Human Thyrotropin Action: Structural, Physiological, and Therapeutic Implications for the Glycoprotein Hormone Family. Endocr. Rev., 1997. [DOI | PubMed]
- E. Decuypere, P. Van As, S. Van der Geyten, V. Darras. Thyroid Hormone Availability and Activity in Avian Species: A Review. Domest. Anim. Endocrinol., 2005. [DOI | PubMed]
- E. Decuypere, J. Buyse. Endocrine Control of Postnatal Growth in Poultry. J. Poult. Sci., 2005. [DOI]
- P.R. Larsen, A.M. Zavacki. Role of the Iodothyronine Deiodinases in the Physiology and Pathophysiology of Thyroid Hormone Action. Eur. Thyroid J., 2013. [DOI]
- R. Mullur, Y.-Y. Liu, G.A. Brent. Thyroid Hormone Regulation of Metabolism. Physiol. Rev., 2014. [DOI | PubMed]
- S. Velleman. Muscle Development in the Embryo and Hatchling. Poult. Sci., 2007. [DOI | PubMed]
- J. Bangsbo, P. Gollnick, T. Graham, B. Saltin. Substrates for Muscle Glycogen Synthesis in Recovery from Intense Exercise in Man. J. Physiol., 1991. [DOI | PubMed]
- N. Oksbjerg, F. Gondret, M. Vestergaard. Basic Principles of Muscle Development and Growth in Meat-Producing Mammals as Affected by the Insulin-like Growth Factor (IGF) System. Domest. Anim. Endocrinol., 2004. [DOI | PubMed]
- F.M. Hansen-Smith, D. Picou, M. Golden. Muscle Satellite Cells in Malnourished and Nutritionally Rehabilitated Children. J. Neurol. Sci., 1979. [DOI | PubMed]
- I. Jirmanová, S. Thesleff. Ultrastructural Study of Experimental Muscle Degeneration and Regeneration in the Adult Rat. Z. Zellforsch. Mikrosk. Anat., 1972. [DOI | PubMed]
- J.K. McGeachie, M.D. Grounds. Initiation and Duration of Muscle Precursor Replication after Mild and Severe Injury to Skeletal Muscle of Mice: An Autoradiographic Study. Cell Tissue Res., 1987. [DOI | PubMed]
- W.D. Phillips, M.R. Bennett. Elimination of Distributed Synaptic Acetylcholine Receptor Clusters on Developing Avian Fast-Twitch Muscle Fibres Accompanies Loss of Polyneuronal Innervation. J. Neurocytol., 1987. [DOI | PubMed]
- A. d’Albis, M. Lenfant-Guyot, C. Janmot, C. Chanoine, J. Weinman, C.L. Gallien. Regulation by Thyroid Hormones of Terminal Differentiation in the Skeletal Dorsal Muscle: I. Neonate Mouse. Dev. Biol., 1987. [DOI | PubMed]
- R. Bischoff. Regeneration of Single Skeletal Muscle Fibers in Vitro. Anat. Rec., 1975. [DOI | PubMed]
- H. Yin, F. Price, M.A. Rudnicki. Satellite Cells and the Muscle Stem Cell Niche. Physiol. Rev., 2013. [DOI | PubMed]
- S. Kuang, K. Kuroda, F. Le Grand, M.A. Rudnicki. Asymmetric Self-Renewal and Commitment of Satellite Stem Cells in Muscle. Cell, 2007. [DOI | PubMed]
- N. Motohashi, A. Asakura. Muscle Satellite Cell Heterogeneity and Self-Renewal. Front. Cell Dev. Biol., 2014. [DOI | PubMed]
- D. Cornelison, B.J. Wold. Single-Cell Analysis of Regulatory Gene Expression in Quiescent and Activated Mouse Skeletal Muscle Satellite Cells. Dev. Biol., 1997. [DOI | PubMed]
- R. Cooper, S. Tajbakhsh, V. Mouly, G. Cossu, M. Buckingham, G. Butler-Browne. In Vivo Satellite Cell Activation via Myf5 and MyoD in Regenerating Mouse Skeletal Muscle. J. Cell Sci., 1999. [DOI | PubMed]
- K.-L. Mak, R.Q. To, Y. Kong, S.F. Konieczny. The MRF4 Activation Domain Is Required to Induce Muscle-Specific Gene Expression. Mol. Cell. Biol., 1992. [PubMed]
- L.A. Sabourin, M.A. Rudnicki. The Molecular Regulation of Myogenesis. Clin. Genet., 2000. [DOI | PubMed]
- R. Perry, M.A. Rudnick. Molecular Mechanisms Regulating Myogenic Determination and Differentiation. Front. Biosci., 2000. [DOI | PubMed]
- L. Lehka, M.J. Rędowicz. Mechanisms Regulating Myoblast Fusion: A Multilevel Interplay. Semin. Cell Dev. Biol., 2020. [DOI | PubMed]
- D.J. Glass. Skeletal Muscle Hypertrophy and Atrophy Signaling Pathways. Int. J. Biochem. Cell Biol., 2005. [DOI | PubMed]
- J.H. Smith. Relation of Body Size to Muscle Cell Size and Number in the Chicken. Poult. Sci., 1963. [DOI]
- R.E. Allen, R.A. Merkel, R.B. Young. Cellular Aspect of Muscle Growth: Myogenic Cell Proliferation. J. Anim. Sci., 1979. [DOI | PubMed]
- O. Halevy, A. Geyra, M. Barak, Z. Uni, D. Sklan. Early Posthatch Starvation Decreases Satellite Cell Proliferation and Skeletal Muscle Growth in Chicks. J. Nutr., 2000. [DOI | PubMed]
- D. Powell, S. Velleman, A. Cowieson, M. Singh, W. Muir. Influence of Chick Hatch Time and Access to Feed on Broiler Muscle Development. Poult. Sci., 2016. [DOI | PubMed]
- S. Torrey, M. Mohammadigheisar, M.N. Dos Santos, D. Rothschild, L.C. Dawson, Z. Liu, E.G. Kiarie, A.M. Edwards, I. Mandell, N. Karrow. In Pursuit of a Better Broiler: Growth, Efficiency, and Mortality of 16 Strains of Broiler Chickens. Poult. Sci., 2021. [DOI | PubMed]
- H. Weintraub. The MyoD Family and Myogenesis: Redundancy, Networks, and Thresholds. Cell, 1993. [DOI | PubMed]
- Z. Yablonka-Reuveni, M.A. Rudnicki, A.J. Rivera, M. Primig, J.E. Anderson, P. Natanson. The Transition from Proliferation to Differentiation Is Delayed in Satellite Cells from Mice Lacking MyoD. Dev. Biol., 1999. [DOI | PubMed]
- Z. Yablonka–Reuveni, B.M. Paterson. MyoD and Myogenin Expression Patterns in Cultures of Fetal and Adult Chicken Myoblasts. J. Histochem. Cytochem., 2001. [DOI | PubMed]
- S. Velleman, K. Nestor, C. Coy, I. Harford, N. Anthony. Effect of Posthatch Feed Restriction on Broiler Breast Muscle Development and Muscle Transcriptional Regulatory Factor Gene and Heparan Sulfate Proteoglycan Expression. Int. J. Poult. Sci., 2010. [DOI]
- S. Velleman, C. Coy, D. Emmerson. Effect of the Timing of Posthatch Feed Restrictions on Broiler Breast Muscle Development and Muscle Transcriptional Regulatory Factor Gene Expression. Poult. Sci., 2014. [DOI | PubMed]
- J. Averous, J.-C. Gabillard, I. Seiliez, D. Dardevet. Leucine Limitation Regulates Myf5 and myoD Expression and Inhibits Myoblast Differentiation. Exp. Cell Res., 2012. [DOI | PubMed]
- D. Powell, D. McFarland, A. Cowieson, W. Muir, S. Velleman. The Effect of Nutritional Status on Myogenic Gene Expression of Satellite Cells Derived from Different Muscle Types. Poult. Sci., 2014. [DOI | PubMed]
- C. Rehfeldt, M. Te Pas, K. Wimmers, J. Brameld, P. Nissen, C. Berri, L. Valente, D. Power, B. Picard, N. Stickland. Advances in Research on the Prenatal Development of Skeletal Muscle in Animals in Relation to the Quality of Muscle-Based Food. I. Regulation of Myogenesis and Environmental Impact. Animal, 2011. [DOI | PubMed]
- M.J. Duclos. Regulation of Chicken Muscle Growth by Insulin-like Growth Factors a. Ann. N. Y. Acad. Sci., 1998. [DOI | PubMed]
- M.J. Duclos, B. Chevalier, H. Rémignon, F. Ricard, C. Goddard, J. Simon. Divergent Selection for High or Low Growth Rate Modifies the Response of Muscle Cells to Serum or Insulin-like Growth Factor-I in Vitro. Growth Regul., 1996. [PubMed]
- M. Kumegawa, E. Ikeda, S. Hosoda, T. Takuma. In Vitro Effects of Thyroxine and Insulin on Myoblasts from Chick Embryo Skeletal Muscle. Dev. Biol., 1980. [DOI | PubMed]
- N. Suthama, K. Hayashi, M. Toyomizu, Y. Tomita. Effect of Dietary Thyroxine on Growth and Muscle Protein Metabolism in Broiler Chickens. Poult. Sci., 1989. [DOI | PubMed]
- J. He, M. Cao, F. Gao, J. Wang, K. Hayashi. Dietary Thyroid Hormone Improves Growth and Muscle Protein Accumulation of Black-Boned Chickens. Br. Poult. Sci., 2006. [DOI | PubMed]
- O. El-Husseiny, S. Hashish, S. Arafa, A. Madian. Response of Poultry Performance to Environmental Light Colour. Egypt. Poult. Sci. J., 2000
- I. Rozenboim, I. Biran, Y. Chaiseha, S. Yahav, A. Rosenstrauch, D. Sklan, O. Halevy. The Effect of a Green and Blue Monochromatic Light Combination on Broiler Growth and Development. Poult. Sci., 2004. [DOI | PubMed]
- A.F. Abdelazeem. Productive and Physiological Response of Broiler Chickens Exposed to Different Colored Light-Emitting Diode and Reared under Different Stocking Densities. Egypt. Poult. Sci. J., 2019. [DOI]
- E.S. Soliman, R.A. Hassan. Impact of Lighting Color and Duration on Productive Performance and Newcastle Disease Vaccination Efficiency in Broiler Chickens. Vet. World, 2019. [DOI | PubMed]
- D. Senaratna, T. Samarakone, W. Gunawardena. Red Color Light at Different Intensities Affects the Performance, Behavioral Activities and Welfare of Broilers. Asian-Australas. J. Anim. Sci., 2015. [DOI | PubMed]
- N. Jiang, Z. Wang, J. Cao, Y. Dong, Y. Chen. Role of Monochromatic Light on Daily Variation of Clock Gene Expression in the Pineal Gland of Chick. J. Photochem. Photobiol. B Biol., 2016. [DOI | PubMed]
- N.M. Sherwood, S.L. Krueckl, J.E. McRory. The Origin and Function of the Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP)/Glucagon Superfamily. Endocr. Rev., 2000. [PubMed]
- I. Bossis, T.E. Porter. Identification of the Somatostatin Receptor Subtypes Involved in Regulation of Growth Hormone Secretion in Chickens. Mol. Cell. Endocrinol., 2001. [DOI | PubMed]
- F. Meng, G. Huang, S. Gao, J. Li, Z. Yan, Y. Wang. Identification of the Receptors for Somatostatin (SST) and Cortistatin (CST) in Chickens and Investigation of the Roles of cSST28, cSST14, and cCST14 in Inhibiting cGHRH1–27NH2-Induced Growth Hormone Secretion in Cultured Chicken Pituitary Cells. Mol. Cell. Endocrinol., 2014. [DOI | PubMed]
- J.R. Florini, D.Z. Ewton, S.A. Coolican. Growth Hormone and the Insulin-like Growth Factor System in Myogenesis. Endocr. Rev., 1996. [PubMed]
- Y. Kajimoto, P. Rotwein. Structure and Expression of a Chicken Insulin-like Growth Factor I Precursor. Mol. Endocrinol., 1989. [DOI | PubMed]
- A.P. D’Costa, D.M. Prevette, L.J. Houenou, S. Wang, K. Zackenfels, H. Rohrer, J. Zapf, P. Caroni, R.W. Oppenheim. Mechanisms of Insulin-like Growth Factor Regulation of Programmed Cell Death of Developing Avian Motoneurons. J. Neurobiol., 1998. [DOI | PubMed]
- M.J. Duclos, C. Goddard. Insulin-like Growth Factor Receptors in Chicken Liver Membranes: Binding Properties, Specificity, Developmental Pattern and Evidence for a Single Receptor Type. J. Endocrinol., 1990. [DOI | PubMed]
- R.C. Baxter. Insulin-like Growth Factor (IGF)-Binding Proteins: Interactions with IGFs and Intrinsic Bioactivities. Am. J. Physiol.-Endocrinol. Metab., 2000. [DOI | PubMed]
- S.V. Allander, E. Ehrenborg, H. Luthman, D.R. Powell. Conservation of IGFBP Structure during Evolution: Cloning of Chicken Insulin-like Growth Factor Binding Protein-5. Prog. Growth Factor Res., 1995. [DOI | PubMed]
- S.V. Allander, M. Coleman, H. Luthman, D.R. Powell. Chicken Insulin-like Growth Factor Binding Protein (IGFBP)-5: Conservation of IGFBP-5 Structure and Expression during Evolution. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol., 1997. [DOI | PubMed]
- T.J. Schoen, C.A. Bondy, J. Zhou, R. Dhawan, K. Mazuruk, D.R. Arnold, I.R. Rodriguez, R.J. Waldbillig, D.C. Beebe, G.J. Chader. Differential Temporal and Spatial Expression of Insulin-like Growth Factor Binding Protein-2 in Developing Chick Ocular Tissues. Investig. Ophthalmol. Vis. Sci., 1995. [PubMed]
- K. Kelley, K. Schmidt, L. Berg, K. Sak, M. Galima, C. Gillespie, L. Balogh, A. Hawayek, J. Reyes, M. Jamison. Beyond Carrier Proteins Comparative Endocrinology of the Insulin-like Growth Factor-Binding Protein. J. Endocrinol., 2002. [DOI | PubMed]
- D.O. Daza, G. Sundström, C.A. Bergqvist, C. Duan, D. Larhammar. Evolution of the Insulin-like Growth Factor Binding Protein (IGFBP) Family. Endocrinology, 2011. [DOI | PubMed]
- H.-B. Wang, H. Li, Q.-G. Wang, X.-Y. Zhang, S.-Z. Wang, Y.-X. Wang, X.-P. Wang. Profiling of Chicken Adipose Tissue Gene Expression by Genome Array. BMC Genom., 2007. [DOI]
- Y. Yamanaka, E.M. Wilson, R.G. Rosenfeld, Y. Oh. Inhibition of Insulin Receptor Activation by Insulin-like Growth Factor Binding Proteins. J. Biol. Chem., 1997. [DOI | PubMed]
- J.W. Kim. The Endocrine Regulation of Chicken Growth. Asian-Australas. J. Anim. Sci., 2010. [DOI]
- R. Baxter. Insulin-like Growth Factor (IGF) Binding Proteins: The Role of Serum IGFBPs in Regulating IGF Availability. Acta Paediatr., 1991. [DOI]
- K.W. Frommer, K. Reichenmiller, B.S. Schutt, A. Hoeflich, M.B. Ranke, G. Dodt, M.W. Elmlinger. IGF-Independent Effects of IGFBP-2 on the Human Breast Cancer Cell Line Hs578T. J. Mol. Endocrinol., 2006. [DOI | PubMed]
- B. Schutt, M. Langkamp, U. Rauschnabel, M. Ranke, M. Elmlinger. Integrin-Mediated Action of Insulin-like Growth Factor Binding Protein-2 in Tumor Cells. J. Mol. Endocrinol., 2004. [DOI | PubMed]
- S. Mohan, Y. Nakao, Y. Honda, E. Landale, U. Leser, C. Dony, K. Lang, D.J. Baylink. Studies on the Mechanisms by Which Insulin-like Growth Factor (IGF) Binding Protein-4 (IGFBP-4) and IGFBP-5 Modulate IGF Actions in Bone Cells (∗). J. Biol. Chem., 1995. [DOI | PubMed]
- R. Hashimoto, M. Ono, H. Fujiwara, N. Higashihashi, M. Yoshida, T. Enjoh-Kimura, K. Sakano. Binding Sites and Binding Properties of Binary and Ternary Complexes of Insulin-like Growth Factor-II (IGF-II), IGF-Binding Protein-3, and Acid-Labile Subunit. J. Biol. Chem., 1997. [DOI | PubMed]
- J.W. Kim, Y.R. Boisclair. Growth Hormone Signaling in the Regulation of Acid Labile Subunit. Asian-Australas. J. Anim. Sci., 2008. [DOI]
- H. Kim, Y. Fu, H.J. Hong, S.-G. Lee, D.S. Lee, H.M. Kim. Structural Basis for Assembly and Disassembly of the IGF/IGFBP/ALS Ternary Complex. Nat. Commun., 2022. [DOI | PubMed]
- A. Suwanichkul, Y.R. Boisclair, R.C. Olney, S.K. Durham, D.R. Powell. Conservation of a Growth Hormone-Responsive Promoter Element in the Human and Mouse Acid-Labile Subunit Genes. Endocrinology, 2000. [DOI | PubMed]
- R.C. Baxter, J. Dai. Purification and Characterization of the Acid-Labile Subunit of Rat Serum Insulin-like Growth Factor Binding Protein Complex. Endocrinology, 1994. [DOI | PubMed]
- G.T. Ooi, F.J. Cohen, L.Y.-H. Tseng, M.M. Rechler, Y.R. Boisclair. Growth Hormone Stimulates Transcription of the Gene Encoding the Acid-Labile Subunit (ALS) of the Circulating Insulin-like Growth Factor-Binding Protein Complex and ALS Promoter Activity in Rat Liver. Mol. Endocrinol., 1997. [DOI | PubMed]
- R.C. Bunn, J.L. Fowlkes. Insulin-like Growth Factor Binding Protein Proteolysis. Trends Endocrinol. Metab., 2003. [DOI | PubMed]
- S.K. Durham, B.L. Riggs, S.A. Harris, C.A. Conover. Alterations in Insulin-like Growth Factor (IGF)-Dependent IGF-Binding Protein-4 Proteolysis in Transformed Osteoblastic Cells. Endocrinology, 1995. [DOI | PubMed]
- R.A. Frost, C.H. Lang. Differential Effects of Insulin-like Growth Factor I (IGF-I) and IGF-Binding Protein-1 on Protein Metabolism in Human Skeletal Muscle Cells. Endocrinology, 1999. [DOI | PubMed]
- M.C. Fisher, C. Meyer, G. Garber, C.N. Dealy. Role of IGFBP2, IGF-I and IGF-II in Regulating Long Bone Growth. Bone, 2005. [DOI | PubMed]
- D.Z. Ewton, S.A. Coolican, S. Mohan, S.D. Chernausek, J.R. Florini. Modulation of Insulin-like Growth Factor Actions in L6A1 Myoblasts by Insulin-like Growth Factor Binding Protein (IGFBP)-4 and IGFBP-5: A Dual Role for IGFBP-5. J. Cell. Physiol., 1998. [DOI | PubMed]
- F.J. Ballard, R.J. Johnson, P.C. Owens, G.L. Francis, F.M. Upton, J.P. McMurtry, J.C. Wallace. Chicken Insulin-like Growth Factor-I: Amino Acid Sequence, Radioimmunoassay, and Plasma Levels between Strains and during Growth. Gen. Comp. Endocrinol., 1990. [DOI | PubMed]
- T.E. Porter, L.E. Ellestad, A. Fay, J.L. Stewart, I. Bossis. Identification of the Chicken Growth Hormone-Releasing Hormone Receptor (GHRH-R) mRNA and Gene: Regulation of Anterior Pituitary GHRH-R mRNA Levels by Homologous and Heterologous Hormones. Endocrinology, 2006. [DOI | PubMed]
- E.M. Kajkowski, L.A. Price, M.H. Pausch, K.H. Young, B.A. Ozenberger. Investigation of Growth Hormone Releasing Hormone Receptor Structure and Activity Using Yeast Expression Technologies. J. Recept. Signal Transduct., 1997. [DOI]
- C. Wang, Y. Wang, J. Li, F. Leung. Expression Profiles of Growth Hormone-Releasing Hormone and Growth Hormone-Releasing Hormone Receptor during Chicken Embryonic Pituitary Development. Poult. Sci., 2006. [DOI | PubMed]
- Y. Wang, J. Li, C.Y. Wang, A.Y. Kwok, X. Zhang, F. Leung. Characterization of the Receptors for Chicken GHRH and GHRH-Related Peptides: Identification of a Novel Receptor for GHRH and the Receptor for GHRH-LP (PRP). Domest. Anim. Endocrinol., 2010. [DOI | PubMed]
- K.E. Mayo, T.L. Miller, V. DeAlmeida, J. Zheng, P.A. Godfrey. The Growth-Hormone-Releasing Hormone Receptor: Signal Transduction, Gene Expression, and Physiological Function in Growth Regulation. Ann. N. Y. Acad. Sci., 1996. [DOI | PubMed]
- R.S. Struthers, W.W. Vale, C. Arias, P.E. Sawchenko, M.R. Montminy. Somatotroph Hypoplasia and Dwarfism in Transgenic Mice Expressing a Non-Phosphorylatable CREB Mutant. Nature, 1991. [DOI | PubMed]
- L.M. Bilezikjian, W.W. Vale. Stimulation of Adenosine 3′, 5′-Monophosphate Production by Growth Hormone-Releasing Factor and Its Inhibition by Somatostatin in Anterior Pituitary Cells in Vitro. Endocrinology, 1983. [DOI | PubMed]
- L.N. Møller, C.E. Stidsen, B. Hartmann, J.J. Holst. Somatostatin Receptors. Biochim. Biophys. Acta (BBA)-Biomembr., 2003. [DOI]
- T. Reisine, H. Kong, K. Raynor, H. Yano, J. Takeda, K. Yasuda, G.I. Bell. Splice Variant of the Somatostatin Receptor 2 Subtype, Somatostatin Receptor 2B, Couples to Adenylyl Cyclase. Mol. Pharmacol., 1993. [DOI | PubMed]
- M. Cakir, D. Dworakowska, A. Grossman. Somatostatin Receptor Biology in Neuroendocrine and Pituitary Tumours: Part 1—Molecular Pathways. J. Cell. Mol. Med., 2010. [DOI | PubMed]
- X. Qin, X. Liu, X. Yan, M. Long, Z. Wang, Y. Dong, Y. Chen, J. Cao. Melatonin Mediates Monochromatic Light-Induced Expression of Somatostatin in the Hypothalamus and Pituitary of Chicks. Poult. Sci., 2021. [DOI | PubMed]
- F. Dehkhoda, C.M. Lee, J. Medina, A.J. Brooks. The Growth Hormone Receptor: Mechanism of Receptor Activation, Cell Signaling, and Physiological Aspects. Front. Endocrinol., 2018. [DOI | PubMed]
- A.J. Brooks, J.W. Wooh, K.A. Tunny, M.J. Waters. Growth Hormone Receptor; Mechanism of Action. Int. J. Biochem. Cell Biol., 2008. [DOI | PubMed]
- H. Lan, W. Li, Z. Fu, Y. Yang, T. Wu, Y. Liu, H. Zhang, H. Cui, Y. Li, P. Hong. Differential Intracellular Signalling Properties of the Growth Hormone Receptor Induced by the Activation of an Anti-GHR Antibody. Mol. Cell. Endocrinol., 2014. [DOI | PubMed]
- K. Hull, A. Thiagarajah, S. Harvey. Cellular Localization of Growth Hormone Receptors/Binding Proteins in Immune Tissues. Cell Tissue Res., 1996. [DOI | PubMed]
- M.A. Lawlor, D.R. Alessi. PKB/Akt: A Key Mediator of Cell Proliferation, Survival and Insulin Responses?. J. Cell Sci., 2001. [DOI | PubMed]
- D.C. Fingar, J. Blenis. Target of Rapamycin (TOR): An Integrator of Nutrient and Growth Factor Signals and Coordinator of Cell Growth and Cell Cycle Progression. Oncogene, 2004. [DOI | PubMed]
- C.J. Potter, L.G. Pedraza, T. Xu. Akt Regulates Growth by Directly Phosphorylating Tsc2. Nat. Cell Biol., 2002. [DOI | PubMed]
- K. Inoki, Y. Li, T. Zhu, J. Wu, K.-L. Guan. TSC2 Is Phosphorylated and Inhibited by Akt and Suppresses mTOR Signalling. Nat. Cell Biol., 2002. [DOI | PubMed]
- D.-H. Kim, D.D. Sarbassov, S.M. Ali, J.E. King, R.R. Latek, H. Erdjument-Bromage, P. Tempst, D.M. Sabatini. mTOR Interacts with Raptor to Form a Nutrient-Sensitive Complex That Signals to the Cell Growth Machinery. Cell, 2002. [DOI | PubMed]
- S.M. Schieke, D. Phillips, J.P. McCoy, A.M. Aponte, R.-F. Shen, R.S. Balaban, T. Finkel. The Mammalian Target of Rapamycin (mTOR) Pathway Regulates Mitochondrial Oxygen Consumption and Oxidative Capacity. J. Biol. Chem., 2006. [DOI | PubMed]
- J.O. Lipton, M. Sahin. The Neurology of mTOR. Neuron, 2014. [DOI | PubMed]
- T.R. Peterson, S.S. Sengupta, T.E. Harris, A.E. Carmack, S.A. Kang, E. Balderas, D.A. Guertin, K.L. Madden, A.E. Carpenter, B.N. Finck. mTOR Complex 1 Regulates Lipin 1 Localization to Control the SREBP Pathway. Cell, 2011. [DOI | PubMed]
- C. Chauvin, V. Koka, A. Nouschi, V. Mieulet, C. Hoareau-Aveilla, A. Dreazen, N. Cagnard, W. Carpentier, T. Kiss, O. Meyuhas. Ribosomal Protein S6 Kinase Activity Controls the Ribosome Biogenesis Transcriptional Program. Oncogene, 2014. [DOI | PubMed]
- S. Sultana, M.R. Hassan, H.S. Choe, K.S. Ryu. The Effect of Monochromatic and Mixed LED Light Colour on the Behaviour and Fear Responses of Broiler Chicken. Avian Biol. Res., 2013. [DOI]
- O. Halevy, Y. Piestun, I. Rozenboim, Z. Yablonka-Reuveni. In Ovo Exposure to Monochromatic Green Light Promotes Skeletal Muscle Cell Proliferation and Affects Myofiber Growth in Posthatch Chicks. Am. J. Physiol.-Regul. Integr. Comp. Physiol., 2006. [DOI | PubMed]
- X. Bai, Y. Wang, Z. Wang, J. Cao, Y. Dong, Y. Chen. In Ovo Exposure to Monochromatic Lights Affect Posthatch Muscle Growth and Satellite Cell Proliferation of Chicks: Role of IGF-1. Growth Factors, 2016. [DOI | PubMed]
- L. Zhang, H. Zhang, J. Wang, S. Wu, X. Qiao, H. Yue, J. Yao, G. Qi. Stimulation with Monochromatic Green Light during Incubation Alters Satellite Cell Mitotic Activity and Gene Expression in Relation to Embryonic and Posthatch Muscle Growth of Broiler Chickens. Animal, 2014. [DOI | PubMed]
- I. Helva, M. Aksit, S. Yalcin. Effects of Monochromatic Light on Growth Performance, Welfare and Hormone Levels in Broiler Chickens. Eur. Poult. Sci., 2019. [DOI]
- M. Arowolo, J. He, S. He, T. Adebowale. The Implication of Lighting Programmes in Intensive Broiler Production System. World’s Poult. Sci. J., 2019. [DOI]
- K. Schwean-Lardner, B.I. Fancher, H.L. Classen. Impact of Daylength on Behavioural Output in Commercial Broilers. Appl. Anim. Behav. Sci., 2012. [DOI]
- H. Olanrewaju, J. Thaxton, W. Dozier, J. Purswell, W. Roush, S. Branton. A Review of Lighting Programs for Broiler Production. Int. J. Poult. Sci., 2006. [DOI]
- C. Magee, H. Olanrewaju, J. Campbell, J. Purswell. Effect of Photoperiod on Live Performance in Broiler Chicks from Placement to 14-Days-of-Age. J. Appl. Poult. Res., 2022. [DOI]
- Y. Yang, W. Cong, J. Liu, M. Zhao, P. Xu, W. Han, D. Wang, R. Zhao. Constant Light in Early Life Induces Fear-Related Behavior in Chickens with Suppressed Melatonin Secretion and Disrupted Hippocampal Expression of Clock-and BDNF-Associated Genes. J. Anim. Sci. Biotechnol., 2022. [DOI | PubMed]
- G.G. Ashabranner, M. Czarick, B.D. Fairchild. Evaluating the Effect of Daylength (24, 20, and 18 Hours) during Brooding on Broiler Performance and Physiological Responses to Light Environment. J. Appl. Poult. Res., 2025. [DOI]
- S. Jiang, Y. Fu, H. Cheng. Daylight Exposure and Circadian Clocks in Broilers: Part I—Photoperiod Effect on Broiler Behavior, Skeletal Health, and Fear Response. Poult. Sci., 2023. [DOI | PubMed]
- D.E. Fidan, A. Nazligü, M.K. Türkyilmaz, S.U. Aypak, F.S. Kilimci, S. Karaarslan, M. Kaya. Effect of photoperiod length and light intensity on some welfare criteria, carcass, and meat quality characteristics in broilers. R. Bras. Zootec, 2017. [DOI]
- I. Dunn, N. Ciccone, N. Joseph. Endocrinology and Genetics of the Hypothalamic-Pituitary-Gonadal Axis. Biology of Breeding Poultry, 2009
- F. Robinson, R. Etches. Ovarian Steroidogenesis during Foillicular Maturation in the Domestic Fowl (Gallus domesticus). Biol. Reprod., 1986. [DOI | PubMed]
- T. Ubuka, G.E. Bentley, K. Tsutsui. Neuroendocrine Regulation of Gonadotropin Secretion in Seasonally Breeding Birds. Front. Neurosci., 2013. [DOI | PubMed]
- P. Rangel, C. Gutierrez. Reproduction in Hens: Is Testosterone Necessary for the Ovulatory Process?. Gen. Comp. Endocrinol., 2014. [DOI | PubMed]
- O. Terada, K. Shimada, N. Saito. Effect of Oestradiol Replacement in Ovariectomized Chickens on Pituitary LH Concentrations and Concentrations of mRNAs Encoding LH β and α Subunits. Reproduction, 1997. [DOI]
- M.M. Bain, Y. Nys, I.C. Dunn. Increasing Persistency in Lay and Stabilising Egg Quality in Longer Laying Cycles. What Are the Challenges?. Br. Poult. Sci., 2016. [DOI | PubMed]
- M.A. Ottinger, M.R. Bakst. Endocrinology of the Avian Reproductive System. J. Avian Med. Surg., 1995
- T. Ubuka, K. Ukena, P.J. Sharp, G.E. Bentley, K. Tsutsui. Gonadotropin-Inhibitory Hormone Inhibits Gonadal Development and Maintenance by Decreasing Gonadotropin Synthesis and Release in Male Quail. Endocrinology, 2006. [DOI | PubMed]
- V.S. Chowdhury, K. Yamamoto, T. Ubuka, G.E. Bentley, A. Hattori, K. Tsutsui. Melatonin Stimulates the Release of Gonadotropin-Inhibitory Hormone by the Avian Hypothalamus. Endocrinology, 2010. [DOI | PubMed]
- T. Ubuka, G.E. Bentley, K. Ukena, J.C. Wingfield, K. Tsutsui. Melatonin Induces the Expression of Gonadotropin-Inhibitory Hormone in the Avian Brain. Proc. Natl. Acad. Sci. USA, 2005. [DOI | PubMed]
- R. Jozsa, A. Mess, R. Gladwell, F. Cunningham, P. Sharp. The Stimulatory Action of the Glutamate Agonist, N-Methyl-Aspartate, on Luteinizing Hormone Release in the Cockerel with Immunocytochemical Observations on Its Mode of Action. Neuroendocrinology: Retrospect and Perspectives, 1997
- R. Teruyama, M. Beck. Double Immunocytochemistry of Vasoactive Intestinal Peptide and cGnRH-I in Male Quail: Photoperiodic Effects. Cell Tissue Res., 2001. [DOI | PubMed]
- A.M. Contijoch, S. Malamed, D.K. Sarkar, J.-P. Advis. β-Endorphin Regulation of LHRH Release at the Median Eminence Level: Immunocytochemical and Physiological Evidence in Hens. Neuroendocrinology, 1993. [DOI | PubMed]
- T. Boswell, I. Dunn, S. Corr. Hypothalamic Neuropeptide Y mRNA Is Increased after Feed Restriction in Growing Broilers. Poult. Sci., 1999. [DOI | PubMed]
- N. Ciccone, I. Dunn, P. Sharp. Increased Food Intake Stimulates GnRH-I, Glycoprotein Hormone α-Subunit and Follistatin mRNAs, and Ovarian Follicular Numbers in Laying Broiler Breeder Hens. Domest. Anim. Endocrinol., 2007. [DOI | PubMed]
- J. Advis, A. Contijoch. The Median Eminence as a Site for Neuroendocrine Control of Reproduction in Hens. Poult. Sci., 1993. [DOI | PubMed]
- R. Bhatt, O. Youngren, S. Kang, M. El Halawani. Dopamine Infusion into the Third Ventricle Increases Gene Expression of Hypothalamic Vasoactive Intestinal Peptide and Pituitary Prolactin and Luteinizing Hormone β Subunit in the Turkey. Gen. Comp. Endocrinol., 2003. [DOI | PubMed]
- A. Thayananuphat, S. Kang, T. Bakken, J. Millam, M. El Halawani. Rhythmic Dependent Light Induction of Gonadotrophin-releasing hormone-I Expression and Activation of Dopaminergic Neurones within the Premammillary Nucleus of the Turkey Hypothalamus. J. Neuroendocrinol., 2007. [DOI | PubMed]
- G.S. Fraley, W.J. Kuenzel. Immunocytochemical and Histochemical Analyses of Gonadotrophin Releasing Hormone, Tyrosine Hydroxylase, and Cytochrome Oxidase Reactivity within the Hypothalamus of Chicks Showing Early Sexual Maturation. Histochemistry, 1993. [DOI | PubMed]
- A.M. Contijoch, C. Gonzalez, H.N. Singh, S. Malamed, S. Troncoso, J.-P. Advis. Dopaminergic Regulation of Luteinizing Hormone-Releasing Hormone Release at the Median Eminence Level: Immunocytochemical and Physiological Evidence in Hens. Neuroendocrinology, 1992. [DOI | PubMed]
- S.L. Meddle, B.K. Follett. Photoperiodically Driven Changes in Fos Expression within the Basal Tuberal Hypothalamus and Median Eminence of Japanese Quail. J. Neurosci., 1997. [DOI | PubMed]
- T. Yamamura, S. Yasuo, K. Hirunagi, S. Ebihara, T. Yoshimura. T3 Implantation Mimics Photoperiodically Reduced Encasement of Nerve Terminals by Glial Processes in the Median Eminence of Japanese Quail. Cell Tissue Res., 2006. [DOI | PubMed]
- J. Proudman, F. Vandesande, L. Berghman. Immunohistochemical Evidence That Follicle-Stimulating Hormone and Luteinizing Hormone Reside in Separate Cells in the Chicken Pituitary. Biol. Reprod., 1999. [DOI | PubMed]
- I.C. Dunn, J.R. Millam. Gonadotropin Releasing Hormone: Forms and Functions in Birds. Avian Poult. Biol. Rev., 1998
- J.A. Proudman, C.G. Scanes, S.A. Johannsen, L.R. Berghman, M.J. Camp. Comparison of the Ability of the Three Endogenous GnRHs to Stimulate Release of Follicle-Stimulating Hormone and Luteinizing Hormone in Chickens. Domest. Anim. Endocrinol., 2006. [DOI | PubMed]
- P. Sharp, R. Talbot, G. Main, I. Dunn, H. Fraser, N. Huskisson. Physiological Roles of Chicken LHRH-I and-II in the Control of Gonadotrophin Release in the Domestic Chicken. J. Endocrinol., 1990. [DOI | PubMed]
- N.T. Joseph, K. Morgan, R. Sellar, D. McBride, R.P. Millar, I.C. Dunn. The Chicken Type III GnRH Receptor Homologue Is Predominantly Expressed in the Pituitary, and Exhibits Similar Ligand Selectivity to the Type I Receptor. J. Endocrinol., 2009. [DOI | PubMed]
- A. Heding, M. Vrecl, J. Bogerd, A. McGregor, R. Sellar, P.L. Taylor, K.A. Eidne. Gonadotropin-Releasing Hormone Receptors with Intracellular Carboxyl-Terminal Tails Undergo Acute Desensitization of Total Inositol Phosphate Production and Exhibit Accelerated Internalization Kinetics. J. Biol. Chem., 1998. [DOI | PubMed]
- A. Pawson, A. Katz, Y. Sun, J. Lopes, N. Illing, R. Millar, J. Davidson. Contrasting Internalization Kinetics of Human and Chicken Gonadotropin-Releasing Hormone Receptors Mediated by C-Terminal Tail. J. Endocrinol., 1998. [DOI | PubMed]
- K.K. Arora, Z. Cheng, K.J. Catt. Dependence of Agonist Activation on an Aromatic Moiety in the DPLIY Motif of the Gonadotropin-Releasing Hormone Receptor. Mol. Endocrinol., 1996. [PubMed]
- K.K. Arora, Z. Cheng, K.J. Catt. Mutations of the Conserved DRS Motif in the Second Intracellular Loop of the Gonadotropin-Releasing Hormone Receptor Affect Expression, Activation, and Internalization. Mol. Endocrinol., 1997. [DOI | PubMed]
- A.J. Pawson, S.R. Maudsley, J. Lopes, A.A. Katz, Y.-M. Sun, J.S. Davidson, R.P. Millar. Multiple Determinants for Rapid Agonist-Induced Internalization of a Nonmammalian Gonadotropin-Releasing Hormone Receptor: A Putative Palmitoylation Site and Threonine Doublet within the Carboxyl-Terminal Tail Are Critical. Endocrinology, 2003. [DOI | PubMed]
- C.A. McArdle, J. Franklin, L. Green, J. Hislop. Signalling, Cycling and Desensitisation of Gonadotrophin-Releasing Hormone Receptors. J. Endocrinol., 2002. [DOI | PubMed]
- C.A. McArdle, J.S. Davidson, G.B. Willars. The Tail of the Gonadotrophin-Releasing Hormone Receptor: Desensitization at, and Distal to, G Protein-Coupled Receptors. Mol. Cell. Endocrinol., 1999. [DOI | PubMed]
- F. Ruf, M.Y. Fink, S.C. Sealfon. Structure of the GnRH Receptor-Stimulated Signaling Network: Insights from Genomics. Front. Neuroendocrinol., 2003. [DOI | PubMed]
- Y.-M. Sun, C.A. Flanagan, N. Illing, T.R. Ott, R. Sellar, B.J. Fromme, J. Hapgood, P. Sharp, S.C. Sealfon, R.P. Millar. A Chicken Gonadotropin-Releasing Hormone Receptor That Confers Agonist Activity to Mammalian Antagonists: Identification of D-Lys6 in the Ligand and Extracellular Loop Two of the Receptor as Determinants. J. Biol. Chem., 2001. [DOI | PubMed]
- M. Shimizu, G.Y. Bédécarrats. Activation of the Chicken Gonadotropin-Inhibitory Hormone Receptor Reduces Gonadotropin Releasing Hormone Receptor Signaling. Gen. Comp. Endocrinol., 2010. [DOI | PubMed]
- S. You, J. Bridgham, D. Foster, A. Johnson. Characterization of the Chicken Follicle-Stimulating Hormone Receptor (cFSH-R) Complementary Deoxyribonucleic Acid, and Expression of cFSH-R Messenger Ribonucleic Acid in the Ovary. Biol. Reprod., 1996. [DOI | PubMed]
- A. Hernandez, J. Bahr. Role of FSH and Epidermal Growth Factor (EGF) in the Initiation of Steroidogenesis in Granulosa Cells Associated with Follicular Selection in Chicken Ovaries. Reproduction, 2003. [DOI | PubMed]
- A.J. Davis, C.F. Brooks, P.A. Johnson. Activin A and Gonadotropin Regulation of Follicle-Stimulating Hormone and Luteinizing Hormone Receptor Messenger RNA in Avian Granulosa Cells. Biol. Reprod., 2001. [DOI | PubMed]
- A. Johnson, J. Bridgham, D. Woods. Cellular Mechanisms and Modulation of Activin A-and Transforming Growth Factor β-Mediated Differentiation in Cultured Hen Granulosa Cells. Biol. Reprod., 2004. [DOI | PubMed]
- A. Johnson, J. Bridgham. Regulation of Steroidogenic Acute Regulatory Protein and Luteinizing Hormone Receptor Messenger Ribonucleic Acid in Hen Granulosa Cells. Endocrinology, 2001. [DOI | PubMed]
- A. Johnson, J. Bridgham, B. Wagner. Characterization of a Chicken Luteinizing Hormone Receptor (cLH-R) Complementary Deoxyribonucleic Acid, and Expression of cLH-R Messenger Ribonucleic Acid in the Ovary. Biol. Reprod., 1996. [DOI | PubMed]
- V.S. Chowdhury, T. Ubuka, K. Tsutsui. Melatonin Stimulates the Synthesis and Release of Gonadotropin-Inhibitory Hormone in Birds. Gen. Comp. Endocrinol., 2013. [DOI | PubMed]
- S.R. Maddineni, O.M. Ocon-Grove, S.M. Krzysik-Walker, G.L. Hendricks, R. Ramachandran. Gonadotropin-Inhibitory Hormone (GnIH) Receptor Gene Is Expressed in the Chicken Ovary: Potential Role of GnIH in Follicular Maturation. Reproduction, 2008. [DOI | PubMed]
- G.Y. Bédécarrats, H. McFarlane, S.R. Maddineni, R. Ramachandran. Gonadotropin-Inhibitory Hormone Receptor Signaling and Its Impact on Reproduction in Chickens. Gen. Comp. Endocrinol., 2009. [DOI | PubMed]
- S. Han, Y. Wang, L. Liu, D. Li, Z. Liu, X. Shen, H. Xu, X. Zhao, Q. Zhu, H. Yin. Influence of Three Lighting Regimes during Ten Weeks Growth Phase on Laying Performance, Plasma Levels- and Tissue Specific Gene Expression- of Reproductive Hormones in Pengxian Yellow Pullets. PLoS ONE, 2017. [DOI | PubMed]
- P. Lewis, N. Tyler, R. Gous, I. Dunn, P. Sharp. Photoperiodic Response Curves for Plasma LH Concentrations and Age at First Egg in Female Broiler Breeders. Anim. Reprod. Sci., 2008. [DOI | PubMed]
- M. Liu, L. Tan, X. Li, H. Li, Y. Zhang, X. Zi, C. Ge, K. Wang. Analysis of Reproductive Hormones and Transcriptomics in the Hypothalamus and Pituitary under Different Photoperiods. Poult. Sci., 2025. [DOI | PubMed]
- N. Mobarkey, N. Avital, R. Heiblum, I. Rozenboim. The Role of Retinal and Extra-Retinal Photostimulation in Reproductive Activity in Broiler Breeder Hens. Domest. Anim. Endocrinol., 2010. [DOI | PubMed]
- N. Gongruttananun. Influence of Red Light on Reproductive Performance, Eggshell Ultrastructure, and Eye Morphology in Thai-Native Hens. Poult. Sci., 2011. [DOI | PubMed]
- S.F. Pang, C. Ralph, D.P. Reilly. Melatonin in the Chicken Brain: Its Origin, Diurnal Variation, and Regional Distribution. Gen. Comp. Endocrinol., 1974. [DOI | PubMed]
- N. Mobarkey, N. Avital, R. Heiblum, I. Rozenboim. The Effect of Parachlorophenylalanine and Active Immunization Against Vasoactive Intestinal Peptide on Reproductive Activities of Broiler Breeder Hens Photostimulated with Green Light. Biol. Reprod., 2013. [DOI | PubMed]
- D. Foss, L. Carew, E. Arnold. Physiological Development of Cockerels as Influenced by Selected Wavelengths of Environmental Light. Poult. Sci., 1972. [DOI | PubMed]
- V.M. Darras, C.H. Verhoelst, G.E. Reyns, E.R. Kühn, S.V. der Geyten. Thyroid Hormone Deiodination in Birds. Thyroid, 2006. [DOI | PubMed]
- M. Ritchie, A.A. Pilny. The Anatomy and Physiology of the Avian Endocrine System. Vet. Clin. N. Am. Exot. Anim. Pract., 2008. [DOI]
- K.I. Abdel-Fattah, S. Bobek, M. Pietras, A. Sechman, J. Niezgoda. Hypometabolic Effect of 3, 3′, 5′-Triiodothyronine in Chickens: Interaction with Hypermetabolic Effect of 3, 5, 3′-Triiodothyronine. Gen. Comp. Endocrinol., 1990. [DOI | PubMed]
- E. Kühn, M. Herremans, E. Dewil, A. Vanderpooten, P. Rudas, T. Bartha, G. Verheyen, L. Berghman, E. Decuypere. Thyrotropin-Releasing Hormone (TRH) Is Not Thyrotropic but Somatotropic in Fed and Starved Adult Chickens. Reprod. Nutr. Dev., 1991. [DOI | PubMed]
- B. De Groef, S. Van der Geyten, V.M. Darras, E.R. Kühn. Role of Corticotropin-Releasing Hormone as a Thyrotropin-Releasing Factor in Non-Mammalian Vertebrates. Gen. Comp. Endocrinol., 2006. [DOI | PubMed]
- K.L. Geris, G. Meeussen, E.R. Kühn, V.M. Darras. Distribution of Somatostatin in the Brain and of Somatostatin and Thyrotropin-Releasing Hormone in Peripheral Tissues of the Chicken. Brain Res., 2000. [DOI | PubMed]
- R. Goel, R. Raju, J. Maharudraiah, G.S.S. Kumar, K. Ghosh, A. Kumar, T.P. Lakshmi, J. Sharma, R. Sharma, L. Balakrishnan. A Signaling Network of Thyroid-Stimulating Hormone. J. Proteom. Bioinform., 2011. [DOI]
- K. Kowalik, A. Sechman. Role of Thyroid Hormones in Birds in the Light of Recent Knowledge. Rocz. Nauk. Zool., 2020
- S.J. Richardson. Cell and Molecular Biology of Transthyretin and Thyroid Hormones. Int. Rev. Cytol., 2007. [PubMed]
- A. Moura Neto, D.E. Zantut-Wittmann. Abnormalities of Thyroid Hormone Metabolism during Systemic Illness: The Low T3 Syndrome in Different Clinical Settings. Int. J. Endocrinol., 2016. [DOI | PubMed]
- G. Reyns, K. Venken, G.M. De Escobar, E. Kühn, V. Darras. Dynamics and Regulation of Intracellular Thyroid Hormone Concentrations in Embryonic Chicken Liver, Kidney, Brain, and Blood. Gen. Comp. Endocrinol., 2003. [DOI | PubMed]
- S. Van der Geyten, J.P. Sanders, E. Kaptein, V.M. Darras, E.R. Kühn, J.L. Leonard, T.J. Visser. Expression of Chicken Hepatic Type I and Type III Iodothyronine Deiodinases during Embryonic Development. Endocrinology, 1997. [DOI | PubMed]
- S. Van der Geyten, I. Van den Eynde, I.B. Segers, E.R. Kühn, V.M. Darras. Differential Expression of Iodothyronine Deiodinases in Chicken Tissues during the Last Week of Embryonic Development. Gen. Comp. Endocrinol., 2002. [DOI | PubMed]
- C.B. Harvey, G.R. Williams. Mechanism of Thyroid Hormone Action. Thyroid, 2002. [DOI | PubMed]
- N.M. Bourgeois, S.L. Van Herck, P. Vancamp, J. Delbaere, C. Zevenbergen, S. Kersseboom, V.M. Darras, T.J. Visser. Characterization of Chicken Thyroid Hormone Transporters. Endocrinology, 2016. [DOI | PubMed]
- D.D. Newmeyer. The Nuclear Pore Complex and Nucleocytoplasmic Transport. Curr. Opin. Cell Biol., 1993. [DOI | PubMed]
- M.L. Andersson, B. Vennström. Chicken Thyroid Hormone Receptor α Requires the N-Terminal Amino Acids for Exclusive Nuclear Localization. FEBS Lett., 1997. [DOI | PubMed]
- G.A. Brent. Mechanisms of Thyroid Hormone Action. J. Clin. Investig., 2012. [DOI | PubMed]
- A.N. Hollenberg. The Role of the Thyrotropin-Releasing Hormone (TRH) Neuron as a Metabolic Sensor. Thyroid, 2008. [DOI | PubMed]
- E. Gorska, K. Popko, A. Stelmaszczyk-Emmel, O. Ciepiela, A. Kucharska, M. Wasik. Leptin Receptors. Eur. J. Med. Res., 2010. [DOI | PubMed]
- P. Joseph-Bravo, L. Jaimes-Hoy, J.-L. Charli. Advances in TRH Signaling. Rev. Endocr. Metab. Disord., 2016. [DOI | PubMed]
- C. Fekete, R.M. Lechan. Central Regulation of Hypothalamic-Pituitary-Thyroid Axis under Physiological and Pathophysiological Conditions. Endocr. Rev., 2014. [DOI | PubMed]
- X. Li, Z. Li, Y. Deng, J. Zhang, J. Li, Y. Wang. Characterization of a Novel Thyrotropin-Releasing Hormone Receptor, TRHR3, in Chickens. Poult. Sci., 2020. [DOI | PubMed]
- L.E. Ericson, M. Nilsson. Structural and Functional Aspects of the Thyroid Follicular Epithelium. Toxicol. Lett., 1992. [DOI | PubMed]
- C.E. Citterio, H.M. Targovnik, P. Arvan. The Role of Thyroglobulin in Thyroid Hormonogenesis. Nat. Rev. Endocrinol., 2019. [DOI | PubMed]
- L.-Å. Nilsson, G. Wick, J. Kite. Demonstration of Thyroglobulin in the Thyroid Glands of Obese Strain and Normal White Leghorn Chicken Embryos. Clin. Exp. Immunol., 1972. [PubMed]
- J. Jaroszewski, R.S. Sundick, N.R. Rose. Effects of Antiserum Containing Thyroglobulin Antibody on the Chicken Thyroid Gland. Clin. Immunol. Immunopathol., 1978. [DOI | PubMed]
- V.E. Avvedimento, D. Tramontano, M.V. Ursini, A. Monticelli, R. Di Lauro. The Level of Thyroglobulin mRNA Is Regulated by TSH Both in Vitro and in Vivo. Biochem. Biophys. Res. Commun., 1984. [DOI | PubMed]
- B. Friedrichs, C. Tepel, T. Reinheckel, J. Deussing, K. von Figura, V. Herzog, C. Peters, P. Saftig, K. Brix. Thyroid Functions of Mouse Cathepsins B, K, and L. J. Clin. Investig., 2003. [DOI | PubMed]
- B. Rousset, C. Dupuy, F. Miot, J. Dumont. Chapter 2 Thyroid Hormone Synthesis and Secretion. Endotext, 2015
- C. Di Cosmo, X.-H. Liao, A.M. Dumitrescu, N.J. Philp, R.E. Weiss, S. Refetoff. Mice Deficient in MCT8 Reveal a Mechanism Regulating Thyroid Hormone Secretion. J. Clin. Investig., 2010. [DOI | PubMed]
- J. Apriletti, N. Eberhardt, K. Latham, J. Baxter. Affinity Chromatography of Thyroid Hormone Receptors. Biospecific Elution from Support Matrices, Characterization of the Partially Purified Receptor. J. Biol. Chem., 1981. [DOI | PubMed]
- R.M. Evans. The Steroid and Thyroid Hormone Receptor Superfamily. Science, 1988. [DOI | PubMed]
- R.L. Wagner, J.W. Apriletti, M.E. McGrath, B.L. West, J.D. Baxter, R.J. Fletterick. A Structural Role for Hormone in the Thyroid Hormone Receptor. Nature, 1995. [DOI | PubMed]
- G.A. Brent, J.W. Harney, Y. Chen, R.L. Warne, D.D. Moore, P.R. Larsen. Mutations of the Rat Growth Hormone Promoter Which Increase and Decrease Response to Thyroid Hormone Define a Consensus Thyroid Hormone Response Element. Mol. Endocrinol., 1989. [DOI | PubMed]
- L.G. Hudson, J.B. Santon, C.K. Glass, G.N. Gill. Ligand-Activated Thyroid Hormone and Retinoic Acid Receptors Inhibit Growth Factor Receptor Promoter Expression. Cell, 1990. [DOI | PubMed]
- M.A. Lazar, T.J. Berrodin, H.P. Harding. Differential DNA Binding by Monomeric, Homodimeric, and Potentially Heteromeric Forms of the Thyroid Hormone Receptor. Mol. Cell. Biol., 1991. [PubMed]
- J.D. Fondell, H. Ge, R.G. Roeder. Ligand Induction of a Transcriptionally Active Thyroid Hormone Receptor Coactivator Complex. Proc. Natl. Acad. Sci. USA, 1996. [DOI | PubMed]
- A.C. Bianco, B.W. Kim. Deiodinases: Implications of the Local Control of Thyroid Hormone Action. J. Clin. Investig., 2006. [DOI | PubMed]
- F.A. Mcnabb, D.B. King. Thyroid Hormone Effects on Growth, Development, and Metabolism. The Endocrinology of Growth, Development, and Metabolism in Vertebrates, 1993
- J. May. Effect of Fasting on T3 and T4 Concentrations in Chicken Serum. Gen. Comp. Endocrinol., 1978. [DOI | PubMed]
- H. Klandorf, S. Harvey. Food Intake Regulation of Circulating Thyroid Hormones in Domestic Fowl. Gen. Comp. Endocrinol., 1985. [DOI | PubMed]
- S. Yahav. Effect of Early-Age Thermal Conditioning and Food Restriction on Performance and Thermotolerance of Male Broiler Chickens. Br. Poult. Sci., 1999. [DOI | PubMed]
- B. Gancedo, A.L. Alonso-Gómez, N. de Pedro, M.J. Delgado, M. Alonso-Bedate. Daily Changes in Thyroid Activity in the Frog Rana Perezi: Variation with Season. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol., 1996. [DOI]
- D. Hazlerigg, A. Loudon. New Insights into Ancient Seasonal Life Timers. Curr. Biol., 2008. [DOI | PubMed]
- R.A. Hut. Photoperiodism: Shall EYA Compare Thee to a Summer’s Day?. Curr. Biol., 2011. [DOI | PubMed]
- N. Nakao, H. Ono, T. Yamamura, T. Anraku, T. Takagi, K. Higashi, S. Yasuo, Y. Katou, S. Kageyama, Y. Uno. Thyrotrophin in the Pars Tuberalis Triggers Photoperiodic Response. Nature, 2008. [DOI | PubMed]
- J. Osol, D. Foss, L. Carew. Effects of Light Environment and Pinealectomy on Growth and Thyroid Function in the Broiler Cockerel. Poult. Sci., 1980. [DOI]
- Y. Liu, Z. Wang, J. Cao, Y. Dong, T. Gao, Y. Chen. Effects of Thyroid Hormone on Monochromatic Light Combinations Mediate Skeletal Muscle Fiber Pattern in Broilers. Poult. Sci., 2024. [DOI | PubMed]
- S. Yahav, A. Straschnow, I. Plavnik, S. Hurwitz. Effects of Diurnally Cycling versus Constant Temperatures on Chicken Growth and Food Intake. Br. Poult. Sci., 1996. [DOI | PubMed]
- W.E. Visser, K.A. Heemstra, S.M. Swagemakers, Z. Ozgur, E.P. Corssmit, J. Burggraaf, W.F. van Ijcken, P.J. van der Spek, J.W. Smit, T.J. Visser. Physiological Thyroid Hormone Levels Regulate Numerous Skeletal Muscle Transcripts. J. Clin. Endocrinol. Metab., 2009. [DOI | PubMed]
- C. Luongo, M. Dentice, D. Salvatore. Deiodinases and Their Intricate Role in Thyroid Hormone Homeostasis. Nat. Rev. Endocrinol., 2019. [DOI | PubMed]
- F.F. Bloise, A. Cordeiro, T.M. Ortiga-Carvalho. Role of Thyroid Hormone in Skeletal Muscle Physiology. J. Endocrinol., 2018. [DOI | PubMed]
- S. Sagliocchi, A.G. Cicatiello, E. Di Cicco, R. Ambrosio, C. Miro, D. Di Girolamo, A. Nappi, G. Mancino, M.A. De Stefano, C. Luongo. The Thyroid Hormone Activating Enzyme, Type 2 Deiodinase, Induces Myogenic Differentiation by Regulating Mitochondrial Metabolism and Reducing Oxidative Stress. Redox Biol., 2019. [DOI | PubMed]
- A.G. Cicatiello, S. Sagliocchi, A. Nappi, E. Di Cicco, C. Miro, M. Murolo, M. Stornaiuolo, M. Dentice. Thyroid Hormone Regulates Glutamine Metabolism and Anaplerotic Fluxes by Inducing Mitochondrial Glutamate Aminotransferase GPT2. Cell Rep., 2022. [DOI | PubMed]
- I. Villanueva, C. Alva-Sánchez, J. Pacheco-Rosado. The Role of Thyroid Hormones as Inductors of Oxidative Stress and Neurodegeneration. Oxidative Med. Cell. Longev., 2013. [DOI]
- M. Costilla, R. Macri Delbono, A. Klecha, G.A. Cremaschi, M.L. Barreiro Arcos. Oxidative Stress Produced by Hyperthyroidism Status Induces the Antioxidant Enzyme Transcription through the Activation of the Nrf-2 Factor in Lymphoid Tissues of Balb/c Mice. Oxidative Med. Cell. Longev., 2019. [DOI]
- J.W. Dransfield. The Lymphatic System of the Domestic Fowl. Vet. J., 1945. [DOI]
- S.L. Freedman. The Innervation of the Suprarenal Gland of the Fowl (Gallus domesticus). Cells Tissues Organs, 1968. [DOI]
- K. Unsicker. Fine Structure and Innervation of the Avian Adrenal Gland: II. Cholinergic Innervation of Adrenal Chromaffin Cells. Z. Zellforsch. Mikrosk. Anat., 1973. [DOI | PubMed]
- K. Unsicker. Fine Structure and Innervation of the Avian Adrenal Gland: III. Non-Cholinergic Nerve Fibers. Z. Zellforsch. Mikrosk. Anat., 1973. [DOI | PubMed]
- K. Unsicker. Fine Structure and Innervation of the Avian Adrenal Gland: V. Innervation of Interrenal Cells. Z. Zellforsch. Mikrosk. Anat., 1973. [DOI | PubMed]
- U. Moawad, M. Randa. Histocytological and Histochemical Features of the Adrenal Gland of Adult Egyptian Native Breeds of Chicken (Gallus gallus domesticus). Beni-Suef Univ. J. Basic Appl. Sci., 2017. [DOI]
- T. Kot, V. Prokopenko. Peculiarities of the Morphology of the Adrenal Glands of Chickens. Наукoві Гoризoнти, 2020
- T. Kot, S. Tkachuk, S. Usenko, V. Prokopenko. Adrenal Gland of Poultry: Anatomy, Microscopy, Morphometry, and Histochemistry. J. World’s Poult. Res., 2023. [DOI]
- S. Sarkar, M.N. Islam, N. Adhikary, B. Paul, N. Bhowmik. Morphological and Histological Studies on the Adrenal Gland in Male and Female Chicken (Gallus domesticus). Int. J. Biol. Pharm. Res., 2014
- I.M. Elzoghby. Light and Electron Microscope Studies of the Adrenal Glands of the Egyptian Geese (Alopochen aegyptiacus). Lucr. Științifice Med. Vet. Univ. Științe Agric. Med. Vet., 2010
- Y. Ohmori, Y. Okada, T. Watanabe. Immunohistochemical Localization of Serotonin, Galanin, Cholecystokinin, and Methionine-Enkephalin in Adrenal Medullary Cells of the Chicken. Tissue Cell, 1997. [DOI | PubMed]
- K. Unsicker. Fine Structure and Innervation of the Avian Adrenal Gland: I. Fine Structure of Adrenal Chromaffin Cells and Ganglion Cells. Z. Zellforsch. Mikrosk. Anat., 1973. [DOI | PubMed]
- W. Holmes, S. Al-Ghawas, J. Cronshaw, K. Rohde. The Structural Organization and the Steroidogenic Responsiveness in Vitro of Adrenal Gland Tissue from the Neonatal Mallard Duck (Anas platyrhynchos). Cell Tissue Res., 1991. [DOI]
- I.C. Jones, J. Phillips. The Adrenal and Interrenal Glands. Morphological Considerations, 1986
- K.A. Humayun, M. Aoyama, S. Sugita. Morphological and Histological Studies on the Adrenal Gland of the Chicken (Gallus domesticus). J. Poult. Sci., 2012. [DOI]
- P. Dadgar Moghadam, A.A. Mohammadpour. Histomorphological and Stereological Study on the Adrenal Glands of Adult Female Guinea Fowl (Numida meleagris). Comp. Clin. Pathol., 2017. [DOI]
- S.M. El-Desoky, F.E.A. Mustafa. Morphological and Histological Studies of the Adrenal Gland in the Japanese Quail (Coturnix japonica). Microsc. Res. Tech., 2021. [DOI | PubMed]
- G. Aguilera. Regulation of Pituitary ACTH Secretion during Chronic Stress. Front. Neuroendocrinol., 1994. [DOI | PubMed]
- F. Madison, A. Jurkevich, W. Kuenzel. Sex Differences in Plasma Corticosterone Release in Undisturbed Chickens (Gallus gallus) in Response to Arginine Vasotocin and Corticotropin Releasing Hormone. Gen. Comp. Endocrinol., 2008. [DOI | PubMed]
- M. Salem, H. Norton, A. Nalbandov. A Study of ACTH and CRF in Chickens. Gen. Comp. Endocrinol., 1970. [DOI | PubMed]
- R. de Matos. Adrenal Steroid Metabolism in Birds: Anatomy, Physiology, and Clinical Considerations. Vet. Clin. N. Am. Exot. Anim. Pract., 2008. [DOI]
- H.-Y. Lin, G. Song, F. Lei, D. Li, Y. Qu. Avian Corticosteroid-Binding Globulin: Biological Function and Regulatory Mechanisms in Physiological Stress Responses. Front. Zool., 2021. [DOI | PubMed]
- G.L. Hammond. Molecular Properties of Corticosteroid Binding Globulin and the Sex-Steroid Binding Proteins. Endocr. Rev., 1990. [DOI | PubMed]
- C. Breuner, S. Lynn, G. Julian, J. Cornelius, B. Heidinger, O. Love, R. Sprague, H. Wada, B. Whitman. Plasma-Binding Globulins and Acute Stress Response. Horm. Metab. Res., 2006. [DOI | PubMed]
- M.S. Khan, W. Rosner. Investigation of the Binding Site of Human Corticosteroid-Binding Globulin by Affinity Labeling. Demonstration of a Cysteinyl Residue in the Binding Site. J. Biol. Chem., 1977. [DOI | PubMed]
- G. Defaye, M. Basset, N. Monnier, E. Chambaz. Electron Spin Resonance Study of Human Transcortin Thiol Groups and Binding Site Topography. Biochim. Biophys. Acta (BBA)-Protein Struct., 1980. [DOI]
- K. Mazancová, M. Kučka, I. Mikšík, J. Pácha. Glucocorticoid Metabolism and Na+ Transport in Chicken Intestine. J. Exp. Zool. Part A Comp. Exp. Biol., 2005. [DOI]
- M. Kučka, K. Vagnerova, P. Klusoňová, I. Mikšík, J. Pacha. Corticosterone Metabolism in Chicken Tissues: Evidence for Tissue-Specific Distribution of Steroid Dehydrogenases. Gen. Comp. Endocrinol., 2006. [DOI | PubMed]
- G. Vashchenko, S. Das, K.-M. Moon, J.C. Rogalski, M.D. Taves, K.K. Soma, F. Van Petegem, L.J. Foster, G.L. Hammond. Identification of Avian Corticosteroid-Binding Globulin (SerpinA6) Reveals the Molecular Basis of Evolutionary Adaptations in SerpinA6 Structure and Function as a Steroid-Binding Protein. J. Biol. Chem., 2016. [DOI | PubMed]
- T. Reisine, G. Rougon, J. Barbet, H.-U. Affolter. Corticotropin-Releasing Factor-Induced Adrenocorticotropin Hormone Release and Synthesis Is Blocked by Incorporation of the Inhibitor of Cyclic AMP-Dependent Protein Kinase into Anterior Pituitary Tumor Cells by Liposomes. Proc. Natl. Acad. Sci. USA, 1985. [DOI | PubMed]
- M.E. Coleman, F. DeMayo, K.C. Yin, H.M. Lee, R. Geske, C. Montgomery, R.J. Schwartz. Myogenic Vector Expression of Insulin-like Growth Factor I Stimulates Muscle Cell Differentiation and Myofiber Hypertrophy in Transgenic Mice. J. Biol. Chem., 1995. [DOI | PubMed]
- E. Harno, T. Gali Ramamoorthy, A.P. Coll, A. White. POMC: The Physiological Power of Hormone Processing. Physiol. Rev., 2018. [DOI | PubMed]
- M.V. Mikhailova, P.R. Mayeux, A. Jurkevich, W.J. Kuenzel, F. Madison, A. Periasamy, Y. Chen, L.E. Cornett. Heterooligomerization between Vasotocin and Corticotropin-Releasing Hormone (CRH) Receptors Augments CRH-Stimulated 3′, 5′-Cyclic Adenosine Monophosphate Production. Mol. Endocrinol., 2007. [DOI | PubMed]
- P.F. Hall, S.B. Koritz. Action of ACTH upon Steroidogenesis in the Chicken Adrenal Gland. Endocrinology, 1966. [DOI | PubMed]
- B. De Groef, K. Vandenborne, P. Van As, V.M. Darras, E.R. Kühn, E. Decuypere, K.L. Geris. Hypothalamic Control of the Thyroidal Axis in the Chicken: Over the Boundaries of the Classical Hormonal Axes. Domest. Anim. Endocrinol., 2005. [DOI | PubMed]
- R.D. Cone. Studies on the Physiological Functions of the Melanocortin System. Endocr. Rev., 2006. [DOI | PubMed]
- S. Takeuchi, T. Kudo, S. Takahashi. Molecular Cloning of the Chicken Melanocortin 2 (ACTH)-Receptor Gene. Biochim. Biophys. Acta (BBA)-Mol. Cell Res., 1998. [DOI]
- J.A. Sebag, P.M. Hinkle. Regulation of G Protein–Coupled Receptor Signaling: Specific Dominant-Negative Effects of Melanocortin 2 Receptor Accessory Protein 2. Sci. Signal., 2010. [DOI | PubMed]
- M.J. Agulleiro, S. Roy, E. Sánchez, S. Puchol, N. Gallo-Payet, J.M. Cerdá-Reverter. Role of Melanocortin Receptor Accessory Proteins in the Function of Zebrafish Melanocortin Receptor Type 2. Mol. Cell. Endocrinol., 2010. [DOI | PubMed]
- T.K. Barlock, D.T. Gehr, R.M. Dores. Analysis of the Pharmacological Properties of Chicken Melanocortin-2 Receptor (cMC2R) and Chicken Melanocortin-2 Accessory Protein 1 (cMRAP1). Gen. Comp. Endocrinol., 2014. [DOI | PubMed]
- L.A. Metherell, J.P. Chapple, S. Cooray, A. David, C. Becker, F. Rüschendorf, D. Naville, M. Begeot, B. Khoo, P. Nürnberg. Mutations in MRAP, Encoding a New Interacting Partner of the ACTH Receptor, Cause Familial Glucocorticoid Deficiency Type 2. Nat. Genet., 2005. [DOI | PubMed]
- S.N. Cooray, I. Almiro Do Vale, K.-Y. Leung, T.R. Webb, J.P. Chapple, M. Egertová, M.E. Cheetham, M.R. Elphick, A.J. Clark. The Melanocortin 2 Receptor Accessory Protein Exists as a Homodimer and Is Essential for the Function of the Melanocortin 2 Receptor in the Mouse Y1 Cell Line. Endocrinology, 2008. [DOI | PubMed]
- W. Pratt, Y. Morishima, M. Murphy, M. Harrell. Chaperoning of Glucocorticoid Receptors. Mol. Chaperones Health Dis., 2006
- E. Kirschke, D. Goswami, D. Southworth, P.R. Griffin, D.A. Agard. Glucocorticoid Receptor Function Regulated by Coordinated Action of the Hsp90 and Hsp70 Chaperone Cycles. Cell, 2014. [DOI | PubMed]
- J.J. Kovacs, P.J. Murphy, S. Gaillard, X. Zhao, J.-T. Wu, C.V. Nicchitta, M. Yoshida, D.O. Toft, W.B. Pratt, T.-P. Yao. HDAC6 Regulates Hsp90 Acetylation and Chaperone-Dependent Activation of Glucocorticoid Receptor. Mol. Cell, 2005. [DOI | PubMed]
- M.R. Yudt, J.A. Cidlowski. Molecular Identification and Characterization of a and b Forms of the Glucocorticoid Receptor. Mol. Endocrinol., 2001. [DOI | PubMed]
- A. Kwok, Y. Wang, C. Wang, F. Leung. Cloning of Chicken Glucocorticoid Receptor (GR) and Characterization of Its Expression in Pituitary and Extrapituitary Tissues. Poult. Sci., 2007. [DOI | PubMed]
- S. Jenkins, T. Porter. Ontogeny of the Hypothalamo–Pituitary–Adrenocortical Axis in the Chicken Embryo: A Review. Domest. Anim. Endocrinol., 2004. [DOI | PubMed]
- S. Leach, K. Suzuki. Adrenergic Signaling in Circadian Control of Immunity. Front. Immunol., 2020. [DOI | PubMed]
- L.M. Romero, M.J. Dickens, N.E. Cyr. The Reactive Scope Model—A New Model Integrating Homeostasis, Allostasis, and Stress. Horm. Behav., 2009. [DOI | PubMed]
- M. Hau, W. Goymann. Endocrine Mechanisms, Behavioral Phenotypes and Plasticity: Known Relationships and Open Questions. Front. Zool., 2015. [DOI | PubMed]
- M. Hau, S. Casagrande, J.Q. Ouyang, A.T. Baugh. Glucocorticoid-Mediated Phenotypes in Vertebrates: Multilevel Variation and Evolution. Adv. Study Behav., 2016
- M. Hesham, A. El Shereen, S. Enas. Impact of Different Light Colors in Behavior, Welfare Parameters and Growth Performance of Fayoumi Broiler Chickens Strain. J. Hell. Vet. Med. Soc., 2018. [DOI]
- G. Archer. Exposing Broiler Eggs to Green, Red and White Light during Incubation. Animal, 2017. [DOI | PubMed]
- H. Olanrewaju, J. Purswell, S. Collier, S. Branton. Effects of Color Temperatures (Kelvin) of LED Bulbs on Blood Physiological Variables of Broilers Grown to Heavy Weights. Poult. Sci., 2015. [DOI | PubMed]
- H. Olanrewaju, W. Miller, W. Maslin, S. Collier, J. Purswell, S. Branton. Influence of Photoperiod, Light Intensity and Their Interaction on Health Indices of Modern Broilers Grown to Heavy Weights. Int. J. Poult. Sci., 2015. [DOI]
- D. Xie, Z. Wang, Y. Dong, J. Cao, J. Wang, J. Chen, Y. Chen. Effects of Monochromatic Light on Immune Response of Broilers. Poult. Sci., 2008. [DOI | PubMed]
- B.R. Franco, T. Shynkaruk, T. Crowe, B. Fancher, N. French, S. Gillingham, K. Schwean-Lardner. Light Color and the Commercial Broiler: Effect on Behavior, Fear, and Stress. Poult. Sci., 2022. [DOI | PubMed]
- G.M. Mehaisen, M.G. Eshak, A.M. Elkaiaty, A.-R.M. Atta, M.M. Mashaly, A.O. Abass. Comprehensive Growth Performance, Immune Function, Plasma Biochemistry, Gene Expressions and Cell Death Morphology Responses to a Daily Corticosterone Injection Course in Broiler Chickens. PLoS ONE, 2017. [DOI | PubMed]