
Cannabinoid Receptor Type 1 Availability in Individuals with a History of Childhood Trauma: A Positron Emission Tomography Study
Abstract
Early life adversity has a lasting impact on the endocannabinoid (eCB) system based on animal models. However, the impact of early life adversity such as childhood trauma (CT) on the eCB system has not been thoroughly studied. We assessed the availability of cannabinoid receptor type 1 (CB1R) in individuals with CT compared to healthy controls without CT (HCs). Cannabinoid receptor type 1 (CB1R) availability was compared in adults with CT (N = 22) and age- and sex-matched HCs (n = 22), using positron emission tomography (PET) imaging with the CB1R-specific radiotracer [11C]OMAR. Using linear models, the effect of the group was assessed on global and trauma-relevant brain regions (amygdala, hippocampus, and frontal cortex). Compared to HCs, lower CB1R availability was observed in CT globally (difference= −11.36%, F(1,42) = 4.35, p = 0.04), in amygdala (−13.70%, F(1,84) = 6.66, p = 0.01), and in hippocampus (−14.50%, F(1,84) = 6.59, p = 0.01), but not in frontal cortex (−8.08%, F(1,84) = 2.17, p = 0.14). There were no effects of a diagnosis of posttraumatic stress disorder, major depressive disorder, nicotine dependence, or the use of antidepressant medication. This preliminary result of lower CB1R availability in adults with CT compared with HCs suggests eCB dysregulation associated with CT. Future studies should replicate and extend this finding and examine the potential effects of various trauma features on the eCB system.
Affiliations: Yale University School of Medicine; Yale School of Medicine; Yaly university; Yale university; Yale University
License: CC BY 4.0 This work is licensed under a Creative Commons Attribution 4.0 International License, which allows reusers to distribute, remix, adapt, and build upon the material in any medium or format, so long as attribution is given to the creator. The license allows for commercial use.
Article links: DOI: 10.21203/rs.3.rs-6536815/v1 | PubMed: 40502783 | PMC: PMC12154151
Relevance: Moderate: mentioned 3+ times in text
Full text: PDF (1.1 MB)
Introduction
Childhood trauma (CT), defined as exposure to significant traumatic events before the age of 18, has profound impacts on mental health [ref. 1]. Individuals who experience CT are at higher risk of developing several psychiatric disorders, including depression and post-traumatic stress disorder (PTSD) [ref. 2, ref. 3]. CT exposure has been reported to lead to structural and functional alterations in the nervous system, resulting in long-lasting consequences [ref. 4–ref. 7]. CT typically imposes threats to survival, body integrity, or sense of self, which explains the significant alterations in the ‘threat-detection and response circuit’, particularly the hippocampus, amygdala, and prefrontal cortex (PFC), in individuals with a history of CT [ref. 7].
The endocannabinoid (eCB) system, which has a central role in stress response [ref. 8], is impacted by various forms of trauma and chronic stress [ref. 9–ref. 13]. Trauma-induced reductions in cannabinoid receptor type 1 (CB1R), the most abundant G-protein coupled receptor in the brain, have been reported in animal models of early life adversity, such as maternal deprivation and social isolation [ref. 14, ref. 15]; these findings persist into adulthood [ref. 16]. Similarly, animal models of chronic stress have reported lower levels of CB1Rs in various brain regions, including the hippocampus [ref. 9–ref. 13].
Despite this evidence from animal studies, the exact impact of CT on the eCB system in humans remains unclear. Human studies on the impact of trauma on the eCB system have predominantly focused on peripheral eCB levels and reported lower peripheral levels of 2-arachidonoyl glycerol (2-AG) and Anandamide (AEA) in those with trauma-related psychiatric disorders [ref. 17–ref. 19]. However, they often did not specifically address CT. In contrast, a few studies that investigated peripheral eCBs in individuals with CT reported higher hair levels of 1-arachidonoylglycerol (1-AG) [ref. 20], and higher plasma levels of an eCB-related compound, oleoylethanolamide (OEA) [ref. 21]. While these studies provide valuable evidence on the unique long-lasting impact of CT on the eCB system, peripheral eCB levels can originate from various sources [ref. 22], and are not correlated with central eCBs [ref. 23]. The impact of CT on the central eCB system in humans has not been reported.
CB1Rs are principal components of the eCB system that can be measured in vivo using PET imaging and the CB1R specific radiotracers, such as [11C]OMAR [ref. 24–ref. 27]. In this study, for the first time, we aimed to compare CB1R availability in adult humans with and without CT histories. Consistent with evidence from animal models [ref. 14, ref. 15], we hypothesized to observe a lower CB1R availability in individuals with a history of CT versus healthy controls with no history of CT (HC), both at a whole-brain level, and in three specific brain regions that express high levels of CB1R and are largely affected by CT, i.e., amygdala, hippocampus, and frontal cortex [ref. 28].
Methods
Participants:
Adult men and women (age ≥ 18) with CT histories (n=22) comprised a CT group, and age- and sex-matched healthy individuals without CT histories served as the HC group (n=22). All individuals received detailed information about the study and signed written informed consent. Inclusion in the CT group was determined based on exposure to a major traumatic event based on Criterion A of the DSM-5 criteria for PTSD, assessed by the Clinician-Administered PTSD Scale for DSM-5 (CAPS-5) or Structured Clinical Interview for DSM-5 Disorders (SCID-5), and exposure to traumatic events for the first time before the age of 18, assessed by the Trauma Questionnaire (TQ) and Childhood Trauma Questionnaire (CTQ) [ref. 29–ref. 32]. Nicotine dependence was assessed using the Fagerstrom Test for Nicotine Dependence (FTND) [ref. 33]. Exclusion criteria included severe mental illness (e.g., schizophrenia), current substance use disorder (excluding nicotine), recent cannabis use, use of anticoagulants, and any medical condition that could interfere with the study, as determined by the primary investigator. The study protocol was approved by the institutional review boards of Yale School of Medicine and the VA Connecticut Health Care System. Additional HCs were included from our group’s previously published research investigating CB1R availability in individuals with other psychiatric disorders [ref. 34, ref. 35].
Radiochemistry and Imaging:
Eligible participants underwent structural Magnetic Resonance Imaging (MRI) using a Siemens 3T system (Siemens Medical Solutions, Malvern, Pennsylvania). Standard T1-weighted anatomical sequences were obtained to exclude anatomical abnormalities and to co-register with PET scan images. [11C]OMAR, a specific CB1R tracer, was used to measure CB1R availability. [11C]OMAR was synthesized with high molar activity according to a previously published protocol adapted to the TRACERlab FXC Pro automated synthesis module (GE Healthcare, Milwaukee, WI) [ref. 36]. Dynamic PET scans were acquired using a High Resolution Research Tomograph (HRRT, Siemens Medical Systems, Knoxville, TN) in 3D mode. Imaging started with a six-minute transmission scan used for attenuation correction. Subsequently, participants were scanned for 120 minutes following an intravenous bolus injection of 12.8 (4.16) mCi [11C]OMAR over a period of one minute. To account for motion, an optical system (Polaris Vicra, Northern Digital Incorporated, Waterloo, Ontario, Canada) was positioned behind the PET scanner to track the position of an infrared reflective tool mounted on the participant’s head. An arterial line was placed prior to the scan with arterial blood sampling acquired throughout the imaging session. Blood samples collected at 5, 15, 30, 60, 90, and 120 minutes post-injection were analyzed using column-switching high-performance liquid chromatography (HPLC) to measure the unmetabolized parent tracer fraction for calculation of the metabolite-corrected arterial plasma input function [ref. 37, ref. 38].
Image Processing:
Consistent with previous work, dynamic list mode data were reconstructed and corrected for attenuation, normalization, scatter, randoms, dead time, and motion using the ordered subset-expectation maximization (OSEM) algorithm [ref. 39]. Subsequently, PET images were registered to the participant’s MR anatomical image and then to an MR template image [ref. 40]. Anatomical Automatic Labeling (AAL) was used to define thirteen regions of interest (ROIs), namely amygdala, caudate, cerebellum, anterior cingulate cortex, posterior cingulate cortex, frontal, hippocampus, insula, occipital, parietal, putamen, temporal, and thalamus [ref. 41]. Partial volume correction was also applied using a method similar to previous studies [ref. 40, ref. 42]. Time-activity curves for each ROI were extracted, and multilinear analysis-1 (t* = 30 min) was used to estimate [11C]OMAR VT in each region [ref. 43].
Statistical Analysis:
All outcomes were assessed for normality using normal probability plots and Kolmogorov-Smirnov tests. Similar to previous studies [ref. 19, ref. 34, ref. 44], a global composite of CB1R availability was calculated by averaging [11C]OMAR VT across all 13 study regions and served as a primary outcome and was compared between groups (CT vs HC) using a linear model. Additionally, three ROIs (amygdala, frontal cortex, and hippocampus) were selected as primary outcomes, given the evidence on the heavy impact of CT on these regions [ref. 7]. These were analyzed using a linear mixed model (LMM) with group as a between-subjects factor and region as a within-subjects factor. The interaction was modeled, and the correlation between observations measured repeatedly within each subject was modeled using structured variance-covariance matrices. The best-fitting model was selected according to information criteria, and residual plots confirmed model fit. Least-square (LS) means were estimated and compared post-hoc. The rest of the ten ROIs were considered as exploratory outcomes and were analyzed using similar LMMs. Previous studies have suggested that age, sex, and body mass index (BMI) relate to CB1R availability in some ROIs [ref. 40]; therefore, these were assessed in each model. However, their inclusion did not alter the model results and had no significant effect on the outcomes, and thus, they were dropped from the model for parsimony. Potential effects of clinical factors within CT (current psychiatric diagnosis, medication use, number of trauma events, nicotine dependence) on global CB1R were assessed using similar linear models as described above. All primary outcomes were tested using a two-sided alpha=0.05 significance threshold. For exploratory analyses testing for group differences in secondary regions and for the effects of clinical factors within the CT group, Type I error was corrected using the less conservative step-down (SD) Sidak adjustment [ref. 45]. All analyses were conducted using SAS, version 9.4 (SAS Institute Inc., Cary, NC, USA).
Results
Baseline characteristics.
Age, sex, BMI, and the dose and mass of the tracer injected during the PET imaging were similar between groups (Table 1). Of the 22 participants with CT, 8 met criteria for current posttraumatic stress disorder (PTSD), and 7 met criteria for current major depressive disorder (MDD, 6 of whom had both MDD and PTSD) (based on SCID-5). Six individuals with CT were taking antidepressant medications.
Table 1:: The baseline characteristics of participants.
| CT (N=22) | HC (N=22) | p vlaue1 | |
|---|---|---|---|
| # of female participants; n(%) | 4 (18.2%) | 4 (18.2%) | 1.00 |
| Age; mean(SD) | 42.5 (10.1) | 39.4 (7.93) | 0.251 |
| BMI; mean(SD) | 29.8 (6.22) | 27.6 (5.14) | 0.214 |
| Tracer dose; mCi; mean(SD) | 12.8 (4.16) | 13.8 (4.00) | 0.553 |
| Tracer mass; μg/kg; mean(SD) | 0.03 (0.02) | 0.03 (0.02) | 0.682 |
| Current PTSD; n (%) | 8 (36.4%) | – | |
| # of traumatic events; n(%) | 19.0 (33.4) | – | |
| Nicotine dependence; n (%) | 2 (9.09%) | – | – |
| FTND Score2; mean (SD) | 4.5 (0.5) | – | – |
| Current MDD; n(%) | 7 (38.9%) | ||
| Current antidepressant use; n (%) | 6 (27.3%) | – | – |
| Current benzodiazepine use; n (%) | 1 (4.5%) | – | – |
Abbreviations: BMI, Body Mass Index; FTND, Fagerstrom Test for Nicotine Dependence; MDD, Major Depressive Disorder; PTSD, Post-Traumatic Stress Disorder.
1 T test (continuous) and Chi-square test (categorical).
2 In those with nicotine dependence.
CB1R availability in the CT versus HC groups.
Lower composite CB1R availability was observed in the CT versus HC groups (difference −11.36%, F(1,42)=4.35; p = 0.04, d = 0.63). In the LMM examining the pre-selected 3 PTSD-relevant ROIs, the group-by-region interactions was significant (F(2,84)=4.49, p=0.014) driving by lower CB1R availability among CT participants in the amygdala (−13.70%, F(1,84)=6.66, p=0.01, d=0.72) and hippocampus (−14.50%, F(1,84)=6.59, p=0.01, d=0.78), but not in the frontal region (−8.08%, F(1,84)=2.17, p=0.14, d=0.47) (Figure1). In the exploratory analysis of other ROIs, no statistically significant differences were observed (Table 2).
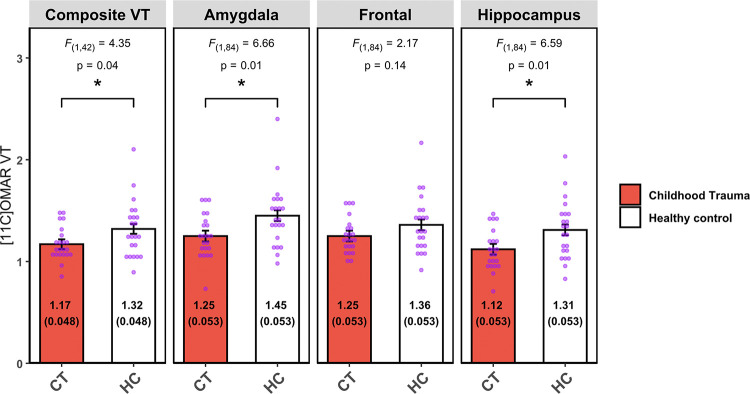
Table 2:: Estimated LS Means of [11C]OMAR VT and LMM-Derived Statistics for Secondary Regions.
| LS mean (SE) | Contrast % | F | PSD Sidak | ||
|---|---|---|---|---|---|
| CT | HC | ||||
| Caudate | 0.94 (0.047) | 1.11 (0.047) | −15.3% | F(1,42) = 6.48 | 0.14 |
| ACC | 1.32 (0.053) | 1.49 (0.053) | −11.4% | F(1,42) = 5.02 | 0.22 |
| PCC | 0.99 (0.050) | 1.10 (0.050) | −10.0% | F(1,42) = 2.15 | 0.32 |
| Cerebellum | 1.16 (0.045) | 1.27 (0.045) | −8.6% | F(1,42) = 3.41 | 0.32 |
| Insula | 1.30 (0.056) | 1.46 (0.056) | −10.9% | F(1,42) = 4.04 | 0.31 |
| Occipital | 1.17 (0.050) | 1.30 (0.050) | −10.0% | F(1,42) = 3.49 | 0.32 |
| Parietal | 1.21 (0.052) | 1.30 (0.052) | −6.9% | F(1,42) = 1.69 | 0.32 |
| Putamen | 1.34 (0.056) | 1.53 (0.056) | −12.4% | F(1,42) = 5.55 | 0.19 |
| Temporal | 1.25 (0.051) | 1.38 (0.051) | −9.4% | F(1,42) = 3.68 | 0.32 |
| Thalamus | 0.94 (0.040) | 1.04 (0.040) | −9.6% | F(1,42) = 2.84 | 0.32 |
Abbreviations: ACC, Anterior Cingulate Cortex; PCC; Posterior Cingulate Cortex
The effect of clinical factors on global CB1R availability among CT
There was no difference in CB1R availability based on meeting criteria for PTSD (F(1,20)=4.01, pSD Sidak=0.27) or MDD (F(1,20)=4.11, pSD Sidak=0.27). On average, CT participants reported encountering 19.0 ± 33.4 traumatic events in their lifetime, and event frequency was not associated with CB1R availability (Spearman r = −0.05, pSD Sidak = 0.84). No differences were observed between participants who were or were not taking antidepressant medications (F(1,20)=2.63, pSD Sidak=0.32). While there was no significant difference in CB1R availability between tobacco users and non-users (F(1,20)=0.51, pSD Sidak=0.71), this result should be interpreted with caution, as only two CT participants reported smoking.
Discussion
Whole-brain CB1R availability is lower in adults with a history of CT compared to controls with a reduction in specific regions, including the hippocampus and amygdala. To the best of our knowledge, this is the first report on the in vivo CB1R availability in adult humans with CT. Our main finding aligns with most preclinical studies that have reported lower CB1R immunoreactivity, expression levels, or binding following early life stressors in various brain regions, especially the hippocampus, with findings persisting into adulthood [ref. 14, ref. 46–ref. 48]. However, not all preclinical studies have reported such findings [ref. 16, ref. 49].
Both animal and human studies have demonstrated that CB1R expression gradually increases during early childhood, reaches its maximum in adolescence, and then drops to stable levels in early adulthood [ref. 50]. Thus, exposure to CT, when the eCB system is still immature, could disrupt the normal development of the main elements of the eCB system, such as CB1R. In addition, the eCB system and the hypothalamic-pituitary-adrenal (HPA) axis interact closely in the initiation and termination of stress responses [ref. 51]. CT has been shown to dysregulate the coupling of the glucocorticoid and eCB systems, leading to impaired regulation of stress responses by glucocorticoids [ref. 52]. Chronic stress typically induces downregulation of presynaptic CB1Rs, which is likely due to the activation of genomic glucocorticoid receptors and could be blocked by glucocorticoid receptor antagonists [ref. 12, ref. 13]. While the precise mechanisms for glucocorticoid receptor activation leading to downregulation of CB1Rs are not fully understood, two mechanisms have been proposed [ref. 8]: 1) direct negative regulation of the CB1R gene by glucocorticoids [ref. 53], or 2) glucocorticoid-mediated recruitment of eCB signaling, resulting in agonist-induced receptor desensitization, as observed with sustained 2-AG signaling elevation [ref. 54].
Considerable research has focused on the potential role of epigenetic programming in the delayed effects of CT on the eCB system. These studies generally indicate that CT has a persistent impact on gene expression and behavior through epigenetic mechanisms, with the HPA axis and glucocorticoid receptors being among the most consistently affected systems. A systematic review reported that CT, both in animal models and human studies, results in increased methylation and lower expression of glucocorticoid receptors [ref. 55]. This implies glucocorticoid resistance and dysregulation of the HPA axis response to stress in adulthood [ref. 55, ref. 56]. Other studies indicate that CT impairs the coupling of the glucocorticoid and eCB systems in the hippocampus, resulting in impaired stress regulation later in life [ref. 52]. Moreover, genetic studies have found significant interactions between CT, particularly sexual abuse, and eCB-related gene polymorphisms in amygdala-related stress habituation and symptoms of cannabis use disorder [ref. 57, ref. 58]. Our findings support the potential specific impact of CT on the eCB system, which may differ from the effects of trauma exposure when it occurs during adulthood.
Future directions: Further studies are required to replicate these findings and investigate the clinical implications of low CB1R availability in individuals with a history of CT. The availability of cannabinoids and the recent development of eCB modulators make it possible now to target the eCB system in recovery from CT [ref. 59]. Moreover, given the critical development of the eCB system in adolescence, future studies should investigate the differential impact of trauma exposure during early childhood versus adolescence. The effects of other factors, such as the cumulative effects of major traumatic events and chronic stress, or the types of trauma, are also important to investigate.
Limitations
Study limitations warrant consideration. First, the sample size was small, but within the range of other published exploratory PET imaging studies on the eCB system [ref. 19, ref. 34, ref. 44]. Nevertheless, these preliminary findings warrant further research with larger sample sizes and more granular information on CT to investigate the impact of various trauma-related factors. Second, the small sample size did not allow for the investigation of sex differences. Sex differences in the eCB system and stress responses have been reported [ref. 60]. Future research should address potential sex differences in CB1R availability among individuals with CT histories. Third, not all HC participants completed questionnaires related to childhood trauma, and the extent of childhood trauma was not fully assessed in all cases. However, any history of CT in a subset of the HCs would likely attenuate rather than inflate the main findings. Lastly, we did not investigate the clinical impact of lower CB1R availability in individuals with CT histories. Further studies are needed to examine how low CB1R availability in individuals with CT histories may relate to stress responses or clinical symptoms.
Conclusions
This exploratory study showed lower CB1R availability in individuals with a history of CT, raising the intriguing possibility of targeting the eCB in the treatment of CT consequences on mental health.
References
- ETC Lippard, CB Nemeroff. The Devastating Clinical Consequences of Child Abuse and Neglect: Increased Disease Vulnerability and Poor Treatment Response in Mood Disorders.. Am J Psychiatry, 2020. [PubMed]
- J Sun, F Patel, R Rose-Jacobs, DA Frank, MM Black, M Chilton. Mothers’ adverse childhood experiences and their young children’s development.. American journal of preventive medicine, 2017. [PubMed]
- KT Brady, SE Back. Childhood trauma, posttraumatic stress disorder, and alcohol dependence.. Alcohol research: current reviews, 2012. [PubMed]
- C Anacker, KJ O’Donnell, MJ Meaney. Early life adversity and the epigenetic programming of hypothalamic-pituitary-adrenal function.. Dialogues in clinical neuroscience, 2014. [PubMed]
- A Danese, TE Moffitt, H Harrington, BJ Milne, G Polanczyk, CM Pariante. Adverse childhood experiences and adult risk factors for age-related disease: depression, inflammation, and clustering of metabolic risk markers.. Archives of pediatrics & adolescent medicine, 2009. [PubMed]
- A Danese, BS McEwen. Adverse childhood experiences, allostasis, allostatic load, and age-related disease.. Physiology & behavior, 2012. [PubMed]
- MH Teicher, JA Samson, CM Anderson, K Ohashi. The effects of childhood maltreatment on brain structure, function and connectivity.. Nat Rev Neurosci, 2016. [PubMed]
- M Morena, S Patel, JS Bains, MN Hill. Neurobiological Interactions Between Stress and the Endocannabinoid System.. Neuropsychopharmacology, 2016. [PubMed]
- MN Hill, S Patel, EJ Carrier, DJ Rademacher, BK Ormerod, CJ Hillard. Downregulation of endocannabinoid signaling in the hippocampus following chronic unpredictable stress.. Neuropsychopharmacology, 2005. [PubMed]
- MN Hill, EJ Carrier, RJ McLaughlin, AC Morrish, SE Meier, CJ Hillard. Regional alterations in the endocannabinoid system in an animal model of depression: effects of concurrent antidepressant treatment.. J Neurochem, 2008. [PubMed]
- W Hu, M Zhang, B Czéh, W Zhang, G Flügge. Chronic restraint stress impairs endocannabinoid mediated suppression of GABAergic signaling in the hippocampus of adult male rats.. Brain Res Bull, 2011. [PubMed]
- JI Wamsteeker, JB Kuzmiski, JS Bains. Repeated stress impairs endocannabinoid signaling in the paraventricular nucleus of the hypothalamus.. J Neurosci, 2010. [PubMed]
- S Rossi, V De Chiara, A Musella, H Kusayanagi, G Mataluni, G Bernardi. Chronic psychoemotional stress impairs cannabinoid-receptor-mediated control of GABA transmission in the striatum.. J Neurosci, 2008. [PubMed]
- J Suárez, R Llorente, SY Romero-Zerbo, B Mateos, FJ Bermúdez-Silva, FR de Fonseca. Early maternal deprivation induces gender-dependent changes on the expression of hippocampal CB(1) and CB(2) cannabinoid receptors of neonatal rats.. Hippocampus, 2009. [PubMed]
- S Alteba, N Korem, I Akirav. Cannabinoids reverse the effects of early stress on neurocognitive performance in adulthood.. Learning & Memory, 2016. [PubMed]
- SA Robinson, RE Loiacono, A Christopoulos, PM Sexton, DT Malone. The effect of social isolation on rat brain expression of genes associated with endocannabinoid signaling.. Brain Res, 2010. [PubMed]
- MN Hill, LM Bierer, I Makotkine, JA Golier, S Galea, BS McEwen. Reductions in circulating endocannabinoid levels in individuals with post-traumatic stress disorder following exposure to the World Trade Center attacks.. Psychoneuroendocrinology, 2013. [PubMed]
- MN Hill, GE Miller, WS Ho, BB Gorzalka, CJ Hillard. Serum endocannabinoid content is altered in females with depressive disorders: a preliminary report.. Pharmacopsychiatry, 2008. [PubMed]
- A Neumeister, MD Normandin, RH Pietrzak, D Piomelli, M-Q Zheng, A Gujarro-Anton. Elevated brain cannabinoid CB1 receptor availability in post-traumatic stress disorder: a positron emission tomography study.. Molecular psychiatry, 2013. [PubMed]
- AM Koenig, W Gao, M Umlauft, K Schury, F Reister, C Kirschbaum. Altered hair endocannabinoid levels in mothers with childhood maltreatment and their newborns.. Biol Psychol, 2018. [PubMed]
- C Schaefer, F Enning, JK Mueller, JM Bumb, C Rohleder, TM Odorfer. Fatty acid ethanolamide levels are altered in borderline personality and complex posttraumatic stress disorders.. Eur Arch Psychiatry Clin Neurosci, 2014. [PubMed]
- CJ Hillard. Circulating Endocannabinoids: From Whence Do They Come and Where are They Going?. Neuropsychopharmacology, 2018. [PubMed]
- A Minichino, M Senior, N Brondino, SH Zhang, BR Godwlewska, PWJ Burnet. Measuring Disturbance of the Endocannabinoid System in Psychosis: A Systematic Review and Meta-analysis.. JAMA Psychiatry, 2019. [PubMed]
- DF Wong, H Kuwabara, AG Horti, V Raymont, J Brasic, M Guevara. Quantification of cerebral cannabinoid receptors subtype 1 (CB1) in healthy subjects and schizophrenia by the novel PET radioligand [11C]OMAR.. Neuroimage, 2010. [PubMed]
- MD Normandin, MQ Zheng, KS Lin, NS Mason, SF Lin, J Ropchan. Imaging the cannabinoid CB1 receptor in humans with [11C]OMAR: assessment of kinetic analysis methods, test-retest reproducibility, and gender differences.. Journal of cerebral blood flow and metabolism : official journal of the International Society of Cerebral Blood Flow and Metabolism, 2015. [PubMed]
- T Tsujikawa, SS Zoghbi, J Hong, SR Donohue, KJ Jenko, RL Gladding. In vitro and in vivo evaluation of (11)C-SD5024, a novel PET radioligand for human brain imaging of cannabinoid CB1 receptors.. Neuroimage, 2014. [PubMed]
- A Neumeister, MD Normandin, RH Pietrzak, D Piomelli, MQ Zheng, A Gujarro-Anton. Elevated brain cannabinoid CB1 receptor availability in post-traumatic stress disorder: a positron emission tomography study.. Mol Psychiatry, 2013. [PubMed]
- NG Harnett, AM Goodman, DC Knight. PTSD-related neuroimaging abnormalities in brain function, structure, and biochemistry.. Experimental neurology, 2020. [PubMed]
- FW Weathers, MJ Bovin, DJ Lee, DM Sloan, PP Schnurr, DG Kaloupek. The Clinician-Administered PTSD Scale for DSM–5 (CAPS-5): Development and initial psychometric evaluation in military veterans.. Psychological assessment, 2018. [PubMed]
- MB First, JBW Williams, RS Karg, RL Spitzer. Structured clinical interview for DSM-5 disorders: SCID-5-CV research version., 2015
- DP Bernstein, L Fink, L Handelsman, J Foote. Childhood trauma questionnaire.. Assessment of family violence: A handbook for researchers and practitioners, 1998
- J Davidson, R Smith. Traumatic experiences in psychiatric outpatients.. Journal of traumatic stress, 1990
- TF Heatherton, LT Kozlowski, RC Frecker, KO Fagerstrom. The Fagerström test for nicotine dependence: a revision of the Fagerstrom Tolerance Questionnaire.. British journal of addiction, 1991. [PubMed]
- DC D’Souza, JA Cortes-Briones, M Ranganathan, H Thurnauer, G Creatura, T Surti. Rapid Changes in CB1 Receptor Availability in Cannabis Dependent Males after Abstinence from Cannabis.. Biol Psychiatry Cogn Neurosci Neuroimaging, 2016
- A Neumeister, MD Normandin, JW Murrough, S Henry, CR Bailey, DA Luckenbaugh. Positron emission tomography shows elevated cannabinoid cb 1 receptor binding in men with alcohol dependence.. Alcoholism: Clinical and Experimental Research, 2012. [PubMed]
- AG Horti, H Fan, H Kuwabara, J Hilton, HT Ravert, DP Holt. 11C-JHU75528: a radiotracer for PET imaging of CB1 cannabinoid receptors.. Journal of Nuclear Medicine, 2006. [PubMed]
- MD Normandin, M-Q Zheng, K-S Lin, NS Mason, S-F Lin, J Ropchan. Imaging the cannabinoid cb1 receptor in humans with [11c] omar: assessment of kinetic analysis methods, test–retest reproducibility, and gender differences.. Journal of Cerebral Blood Flow & Metabolism, 2015. [PubMed]
- J Hilton, F Yokoi, RF Dannals, HT Ravert, Z Szabo, DF Wong. Column-switching HPLC for the analysis of plasma in PET imaging studies.. Nucl Med Biol, 2000. [PubMed]
- 39.Design of a motion-compensation OSEM list-mode algorithm for resolution-recovery reconstruction for the HRRT. Proceedings of the Nuclear Science Symposium Conference Record, 2003 IEEE2003. IEEE.
- R Radhakrishnan, PD Worhunsky, M-Q Zheng, S Najafzadeh, J-D Gallezot, B Planeta. Age, gender and body-mass-index relationships with in vivo CB1 receptor availability in healthy humans measured with [11C] OMAR PET.. NeuroImage, 2022. [PubMed]
- N Tzourio-Mazoyer, B Landeau, D Papathanassiou, F Crivello, O Etard, N Delcroix. Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain.. Neuroimage, 2002. [PubMed]
- G Giovacchini, A Lerner, MT Toczek, C Fraser, K Ma, JC DeMar. Brain Incorporation of 11C-Arachidonic Acid, Blood Volume, and Blood Flow in Healthy Aging: A Study With Partial-Volume Correction.. Journal of Nuclear Medicine, 2004. [PubMed]
- M Ichise, H Toyama, RB Innis, RE Carson. Strategies to improve neuroreceptor parameter estimation by linear regression analysis.. Journal of cerebral blood flow and metabolism : official journal of the International Society of Cerebral Blood Flow and Metabolism, 2002. [PubMed]
- M Ranganathan, J Cortes-Briones, R Radhakrishnan, H Thurnauer, B Planeta, P Skosnik. Reduced Brain Cannabinoid Receptor Availability in Schizophrenia.. Biol Psychiatry, 2016. [PubMed]
- BS Holland, MD Copenhaver. An improved sequentially rejective Bonferroni test procedure.. Biometrics, 1987
- A Romano-López, M Méndez-Díaz, AE Ruiz-Contreras, R Carrisoza, O Prospéro-García. Maternal separation and proclivity for ethanol intake: a potential role of the endocannabinoid system in rats.. Neuroscience, 2012. [PubMed]
- MN Hill, L Eiland, TTY Lee, CJ Hillard, BS McEwen. Early life stress alters the developmental trajectory of corticolimbic endocannabinoid signaling in male rats.. Neuropharmacology, 2019. [PubMed]
- TT Lee, MN Hill. Age of stress exposure modulates the immediate and sustained effects of repeated stress on corticolimbic cannabinoid CB1 receptor binding in male rats.. Neuroscience, 2013. [PubMed]
- EM Marco, V Echeverry-Alzate, JA López-Moreno, E Giné, S Peñasco, MP Viveros. Consequences of early life stress on the expression of endocannabinoid-related genes in the rat brain.. Behav Pharmacol, 2014. [PubMed]
- A Bara, JN Ferland, G Rompala, H Szutorisz, YL Hurd. Cannabis and synaptic reprogramming of the developing brain.. Nat Rev Neurosci, 2021. [PubMed]
- MN Hill, P Campolongo, R Yehuda, S Patel. Integrating Endocannabinoid Signaling and Cannabinoids into the Biology and Treatment of Posttraumatic Stress Disorder.. Neuropsychopharmacology, 2018. [PubMed]
- P Atsak, M Morena, C Schoenmaker, E Tabak, CA Oomen, S Jamil. Glucocorticoid-endocannabinoid uncoupling mediates fear suppression deficits after early – Life stress.. Psychoneuroendocrinology, 2018. [PubMed]
- P Mailleux, JJ Vanderhaeghen. Glucocorticoid regulation of cannabinoid receptor messenger RNA levels in the rat caudate-putamen. An in situ hybridization study.. Neurosci Lett, 1993. [PubMed]
- JE Schlosburg, JL Blankman, JZ Long, DK Nomura, B Pan, SG Kinsey. Chronic monoacylglycerol lipase blockade causes functional antagonism of the endocannabinoid system.. Nat Neurosci, 2010. [PubMed]
- G Turecki, MJ Meaney. Effects of the Social Environment and Stress on Glucocorticoid Receptor Gene Methylation: A Systematic Review.. Biol Psychiatry, 2016. [PubMed]
- T Klengel, EB Binder. Epigenetics of Stress-Related Psychiatric Disorders and Gene × Environment Interactions.. Neuron, 2015. [PubMed]
- CE Carey, A Agrawal, B Zhang, ED Conley, L Degenhardt, AC Heath. Monoacylglycerol lipase (MGLL) polymorphism rs604300 interacts with childhood adversity to predict cannabis dependence symptoms and amygdala habituation: Evidence from an endocannabinoid system-level analysis.. J Abnorm Psychol, 2015. [PubMed]
- J Lazary, N Eszlari, G Juhasz, G Bagdy. Genetically reduced FAAH activity may be a risk for the development of anxiety and depression in persons with repetitive childhood trauma.. Eur Neuropsychopharmacol, 2016. [PubMed]
- LM Mayo, CA Rabinak, MN Hill, M Heilig. Targeting the Endocannabinoid System in the Treatment of Posttraumatic Stress Disorder: A Promising Case of Preclinical-Clinical Translation?. Biol Psychiatry, 2022. [PubMed]
- A Bassir Nia, R Bender, I Harpaz-Rotem. Endocannabinoid System Alterations in Posttraumatic Stress Disorder: A Review of Developmental and Accumulative Effects of Trauma.. Chronic Stress (Thousand Oaks), 2019