
Engineering Saccharomyces cerevisiae for medical applications
Abstract
Background:
During the last decades, the advancements in synthetic biology opened the doors for a profusion of cost-effective, fast, and ecologically friendly medical applications priorly unimaginable. Following the trend, the genetic engineering of the baker’s yeast, Saccharomyces cerevisiae, propelled its status from an instrumental ally in the food industry to a therapy and prophylaxis aid.
Main text:
In this review, we scrutinize the main applications of engineered S. cerevisiae in the medical field focusing on its use as a cell factory for pharmaceuticals and vaccines, a biosensor for diagnostic and biomimetic assays, and as a live biotherapeutic product for the smart in situ treatment of intestinal ailments. An extensive view of these fields’ academic and commercial developments as well as main hindrances is presented.
Conclusion:
Although the field still faces challenges, the development of yeast-based medical applications is often considered a success story. The rapid advances in synthetic biology strongly support the case for a future where engineered yeasts play an important role in medicine.
Article type: Review Article
Keywords: Yeast, Cell factory, Pharmaceutical, Natural product, Vaccine, Biosensor, Live biotherapeutic product, Therapeutic microbe, Genetic engineering, Synthetic biology
Affiliations: https://ror.org/0234wmv40grid.7384.80000 0004 0467 6972Chair of Biochemistry of Microorganisms, Faculty of Life Sciences: Food, Nutrition and Health, University of Bayreuth, 95326 Kulmbach, Germany; https://ror.org/055s37c97grid.418398.f0000 0001 0143 807XDepartment of Microbial Pathogenicity Mechanisms, Leibniz Institute for Natural Product Research and Infection Biology, 07745 Jena, Germany; https://ror.org/05qpz1x62grid.9613.d0000 0001 1939 2794Cluster of Excellence Balance of the Microverse, Friedrich Schiller University Jena, 07743 Jena, Germany
License: © The Author(s) 2024 CC BY 4.0 Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
Article links: DOI: 10.1186/s12934-024-02625-5 | PubMed: 39789534 | PMC: PMC11720383
Relevance: Moderate: mentioned 3+ times in text
Full text: PDF (1.5 MB)
Introduction
The use of Saccharomyces cerevisiae, for the generation of value-added products is deeply rooted in the history of human society, being traced back to thousands of years ago [ref. 1–ref. 3]. Its thorough application in the production of basic goods such as wine, beer, and bread has granted S. cerevisiae the Generally Regarded As Safe (GRAS) classification by the U.S. Food and Drug Administration (FDA). Nevertheless, this millenary knowledge of yeast manipulation was only recently broadened from the traditional manufacture of goods for direct human consumption to emerging biotechnological operations. Pioneering studies on S. cerevisiae’s transformation and recombination, followed by the genome sequencing of the reference isolate S288c by Goffeau et al., 1996, paved the way for a plethora of complex genetic engineering techniques developed in the last 50 years [ref. 4–ref. 6]. In particular, the emergence of Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)-Cas systems after 2012, and its easiness of application in S. cerevisiae, permitted the exploration of metabolic engineering endeavors previously unimaginable [ref. 7–ref. 9]. Together with discoveries in yeast physiology and biochemistry, this knowledge allowed for the easy rational manipulation of the yeast genome, that propelled its status from a useful natural resource to a versatile technological platform.
The medical and pharmaceutical fields, where S. cerevisiae already played a traditional role as a probiotic and a model organism for eukaryotic cells, are no exception [ref. 10]. The establishment of molecular biology as we know it, transformed yeast into a resourceful health ally. From cell-based factories for the production of pharmaceutical goods or immunogens, to sensing platforms for the diagnostics of diseases and pathogens or detection of therapeutic molecules, and even live biotherapeutic products (LBPs), yeasts allow for fair-priced, fast, and reliable therapies and therapy-aiding devices (Fig. 1). In this review, we outline therapeutic endeavors using genetically engineered S. cerevisiae, covering different applications and future perspectives. Medical applications of so-called non-conventional yeast are covered elsewhere and are not included in this review [ref. 11–ref. 14].
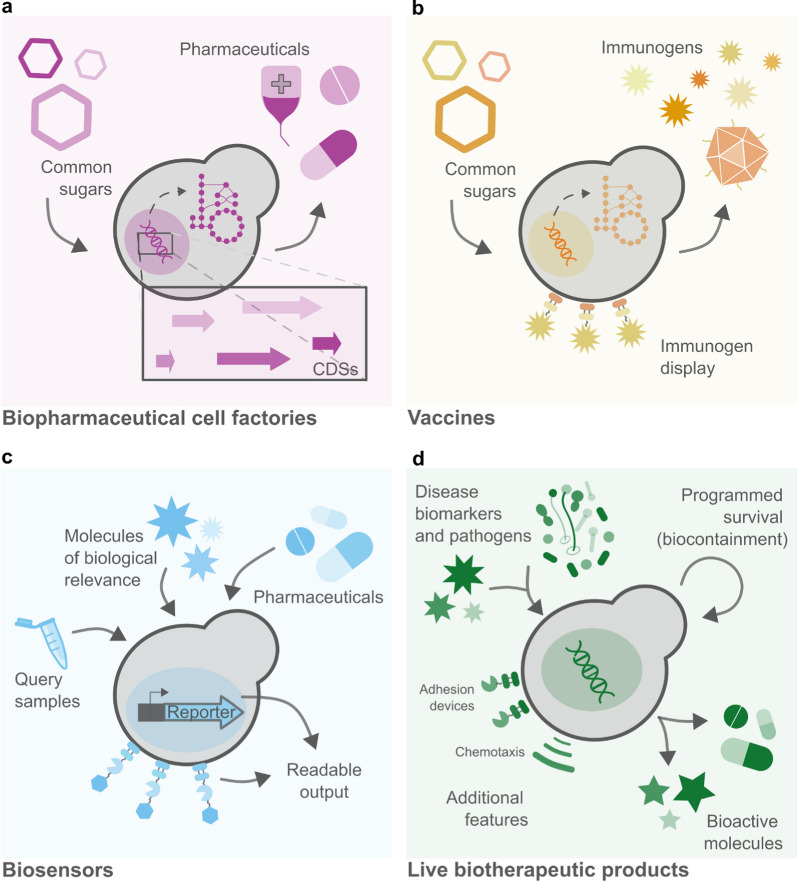
Pharmaceutical cell factories
Pharmaceutical cell factories are genetically engineered microorganisms that produce recombinant proteins of therapeutic relevance, also called biopharmaceuticals (Fig. 1a). They stand out as an impactful alternative to classical manufacturing methods and have the potential to make several therapies more accessible. Engineered cells can alleviate supply chain bottlenecks by eliminating complex cultivation/farming and transport steps that are commonly dependent on external factors—such as climate and politics. They can simplify or even eliminate handling and extraction steps by removing the need to process complex tissues. Additionally, they often align with ecological goals, utilizing substrates that range from simple sugars to waste products. Finally, pharmaceutical cell factories can improve economic aspects of production—many times reflecting on the final price for the customer—by augmenting yields, and/or reducing production costs.
The first successful example of a genetically engineered microorganism for the production of a medicament was the synthesis of recombinant insulin, in 1978. Based on the most recent advances in molecular biology at the time, Genentech researchers were able to clone genes involved in insulin production in Escherichia coli, leading to the synthesis of insulin precursors trapped in inclusion bodies [ref. 15, ref. 16]. Given S. cerevisiae’s secretory system, which shares many resemblances to higher eukaryotes’—including humans—it didn’t take long for researchers at the Novo Research Institute (currently Novo Nordisk Foundation) to turn to this organism in order to improve the production. Taking advantage of the mating factor α1 leader sequence, they could direct the proinsulin-containing fusion protein to the cell’s secretory pathway, obtaining insulin precursors directly in the culture medium [ref. 17, ref. 18]. To this day S. cerevisiae remains the preferred microorganism for insulin production—the main workhorse of an industry with an estimatedket size of approximately USD 20 billion [ref. 16, ref. 19].
The success of yeast-based insulin synthesis opened the doors for the production of a wide range of therapy agents in S. cerevisiae. As single-cell eukaryotes, they represent a good compromise between simple and cost-effective cultivation methods and handling, and a highly conserved post-translational modification (PTM) machinery. In this section, we scrutinize the state of the art of this growing field, addressing challenges and breakthroughs. Yeast cell factories for the production of antibodies were thoroughly revised elsewhere and therefore excluded from this review [ref. 20–ref. 23]. Immunogens-producing strains will be covered in the following chapter, given their close relation with the production of vaccines.
Dozens of different recombinant compounds with therapeutic activity have been successfully synthesized in S. cerevisiae to this day—Table 1 gathers many examples. Similarly to insulin, the first works towards yeast-based biopharmaceuticals relied on the expression of a single exogenous gene provided in a plasmid for protein production. Important examples of commercially relevant compounds manufactured in these terms are the human hormone glucagon, as well as the anticoagulant hirudin, naturally found in leech [ref. 24–ref. 26]. It didn’t take long for researchers to move to more complex synthesis systems, in which multi-step enzymatic pathways would be transferred to the producing host. The expression of four genes from Erwinia sp. for lycopene and β-carotene synthesis in 1994 was one of the first multi-genic pathways imported into S. cerevisiae [ref. 27]. The uprising of genomic engineering techniques permitted even more complex synthesis efforts. S. cerevisiae’s high recombination rate, which allowed for the efficient insertion of repair fragments when subjected to homologous recombination, was a key feature for the rapid development of genomically engineered biopharmaceutical cell factories. For instance, human steroids of therapeutic relevance, such as progesterone and hydrocortisone were efficiently synthesized in yeast at the beginning of the 2000s through a series of genomic integrations and deletions [ref. 28, ref. 29]. Both plant and mammalian enzymes were employed in this work, constituting one of the first examples of new-to-nature synthetic pathways for biopharmaceutical production [ref. 29].
Table 1: Molecules with therapeutic application produced in S. cerevisiae
| Compound class | Compound | Therapeutic application | Original source | References |
|---|---|---|---|---|
| Proteins | ||||
| Cytokine | Sargramostim | Hematopoietic growth factor | A variety of cells | [ref. 30] |
| Plasma protein | Coagulation factor XIII | Treatment of clotting disorder | Human plasma | [ref. 31] |
| Human albumin | Plasma expander | Human plasma | [ref. 32, ref. 33] | |
| Transferrin | Treatment of anemia | Human plasma | [ref. 34] | |
| Protease inhibitor | Aprotinin | Antifibrinolytic | Mammalian parotid glands | [ref. 35, ref. 36] |
| Pre-elafin | Treatment for lung diseases | Human lungs | [ref. 37] | |
| Other molecules | ||||
| Alkaloid | Ajmalicine | Hypotensive, antiarrhythmic | Catharanthus roseus | [ref. 38] |
| Berberine | Broad, antimicrobial, anticancer | Berberis sp., Hydrastis canadensis, Coptis sp. | [ref. 39, ref. 40] | |
| d-lysergic acid | Antipsychotic, anti-depressive, anxiolytic | Claviceps purpurea(rye ergot fungus) | [ref. 41] | |
| Hyoscyamine and scopolamine | Treatment of neuromuscular disorders | Duboisia sp.(corkwood tree) | [ref. 42, ref. 43] | |
| Noscapine | Antitussive, anticancer | Papaver somniferum(opium poppies) | [ref. 44] | |
| Opioids | Analgesic, sedative, antitussive, antispasmodic | Papaver somniferum(opium poppies) | [ref. 45–ref. 50] | |
| Psilocybin | Antipsychotic, anti-depressive, anxiolytic | Psilocybe sp.(magic mushrooms) | [ref. 51] | |
| Rauwolscine and analogs | Broad | Rauwolfia sp. | [ref. 52] | |
| Sanguinarine and analogs | Antimicrobial, presumed anti-cancer | Sanguinaria canadensis(bloodroot plant) | [ref. 38, ref. 45, ref. 53] | |
| Serpentine, Alstonine and analogs | Broad | Catharanthus roseus | [ref. 54] | |
| Strictosidine and hydroxystrictosidine | Broad | Catharanthus roseus | [ref. 55–ref. 57] | |
| Tropine and analogs | Treatment of neuromuscular disorders | Duboisia sp.(corkwood tree) | [ref. 58, ref. 59] | |
| Vinblastine | Anti-cancer | Catharanthus roseus | [ref. 60, ref. 61] | |
| Xanthine-based compounds | Broad | A variety of plants | [ref. 62] | |
| Hormone | Epidermal growth factor (EGF) | Treatment for diabetic foot ulcers | A variety of mammalian tissues | [ref. 63] |
| Glucagon | Metabolic regulator | Mammalian α-cells of the islets of Langerhans | [ref. 24] | |
| Insulin | Metabolic regulator | Mammalian beta cells of the pancreatic islets | [ref. 17, ref. 18] | |
| Insulin-like growth factor I (IGF-I) | Treatment of IGF-I-deficient patients | Mammalian liver | [ref. 64] | |
| Melatonin | Circadian cycle regulator | Mammalian pineal gland | [ref. 65] | |
| Platelet-derived growth factor (PDGF) | Growth factor | Mammalian platelet and muscle cells | [ref. 66, ref. 67] | |
| Somatotropin (human growth hormone) | Treatment of Somatotropin deficiency | Mammalian pituitary gland | [ref. 68, ref. 69] | |
| Non-ribosomal peptide | Penicillin | Antibiotic | Penicillium sp. | [ref. 70] |
| Phenylpropanoid | Anthocyanins | Broad | A variety of plants | [ref. 71] |
| Breviscapine | Treatment of vascular diseases | Erigeron breviscapus | [ref. 72] | |
| Dihydrochalcones | Broad | A variety of herbal and fructiferous plants | [ref. 73] | |
| Flavonoids (e.g. naringenin, fisetin, kaempferol) | Broad | A variety of herbal and fructiferous plants | [ref. 74–ref. 80] | |
| Icariin | Anticancer | Epimendium sp.(barrenwort) | [ref. 81] | |
| P-coumaric acid | Broad | A variety of herbal and fructiferous plants | [ref. 82, ref. 83] | |
| Salidroside | Broad | Rhodiola sp.(golden root) | [ref. 84] | |
| Shikimic acid | Anti-viral | Illicium sp.(e.g. star anise) | [ref. 85] | |
| Stilbenoids | Broad | A variety of fructiferous plants | [ref. 86–ref. 88] | |
| Steroid | Hydrocortisone | Anti-inflammatory, contraceptive | Absidia orchidis | [ref. 29, ref. 89] |
| Pregnenolone and progesterone | Anti-inflammatory, contraceptive | Human adrenal glands | [ref. 28] | |
| Terpenoid | Artemisinin | Antimalarial | Artemisia annua L(sweet wormwood) | [ref. 90–ref. 92] |
| Betulinic acid | Anticancer, antiviral | A variety of plants | [ref. 93] | |
| Cannabinoids | Analgesic, anti-emetic | Cannabis sativa(cannabis) | 113 | |
| Carnosinic acid | Antioxidant | Rosmarinus officinalis L. (rosemary) and Salvia officinalis (sage) | [ref. 94, ref. 95] | |
| Carotenoids (e.g. lycopene, astaxanthin) | Antioxidant | A variety of fructiferous plants | [ref. 27, ref. 96–ref. 98] | |
| Diterpenes (e.g. sclareol, abienol) | Broad | A variety of plants | [ref. 99] | |
| Ginsenosides | Broad | Panax ginseng(ginseng) | [ref. 100–ref. 102] | |
| Monoterpenes (e.g. limonene, geraniol) | Broad | A variety of plants | [ref. 103–ref. 107] | |
| Saponins | Vaccine adjuvant | Bupleurum falcatum | [ref. 108, ref. 109] | |
| Taxadiene | Anticancer | Taxus brevifolia(Pacific yew) | [ref. 110] | |
| Tissue plasminogen activator | Hirudin | Anticoagulant | Salivary glands of Hirudo medicinalis(medicinal leech) | [ref. 25, ref. 26] |
In 2006, Ro et al. reported the production and secretion of the antimalarial drug precursor artemisinic acid in S. cerevisiae [ref. 90]. With the potential to bring cost-effective treatment to hundreds of millions of people infected with malaria each year, the strain was optimized to yield impressive 25 g/L of artemisinic acid with high purity [ref. 91]. Considering the easiness of retrieving the pharmaceutical directly from the yeast culture medium and that its classical extraction from wild-type Artemisia annua plants yields roughly 1.4 g/m2 of cultivated area, the yeast-based production was highly celebrated as a cost-effective and efficient alternative [ref. 111–ref. 113]. In 2014, the pharmaceutical giant Sanofi started a production line of yeast-produced artemisinin (“semi-synthetic” artemisinin—SSA) alongside its classical synthesis via extraction from A. annua. Nevertheless, the SSA faced greatket resistance—mostly due to the drop in prices of the naturally derived product and the withdrawal of important supporting grants for the project [ref. 114, ref. 115].
The yeast-based production of a series of therapeutically relevant benzylisoquinoline (BIA) and monoterpene indole (MIA) alkaloids has also been the subject of an exciting scientific race. Commonly extracted from plants, the complexity of the metabolic network leading to their production translates into low concentrations in planta—with instances as low as 0.0005% of dry weight [ref. 116–ref. 118]. At the same time, their chemical synthesis is often compromised due to their molecular intricacy (e.g. the presence of one or more chiral centers) and high costs [ref. 116, ref. 119, ref. 120]. Yeast-based biopharmaceutical factories have, therefore, been extensively utilized for the production of opiates—a class of BIAs accounting for a variety of potent pain relievers naturally extracted from the opium poppy (Papaver somniferum) [ref. 45–ref. 50]. Even though the production was primarily achieved via supplementation with precursors, the de novo synthesis of opiates was recently made possible due to the discovery of a key enzyme that epimerizes the (S)-benzylisoquinoline scaffold to the (R)-enantiomer [ref. 46–ref. 49, ref. 121–ref. 123]. Alongside the race for BIAs production, S. cerevisiae genome was heavily modified for MIAs production. In 2022, Zhang et al. published their remarkable effort to produce the anticancer drug vinblastine in S. cerevisiae. The final strain carried nothing less than 56 genetic edits and displayed a 1000-fold increase in the production of the intermediate product strictosidine [ref. 56, ref. 60]. In 2023, Bradley et al. reported the production of anxiolytic drugs serpentine and alstonine [ref. 54]. They took a step further, demonstrating the synthesis of new-to-nature halogenated MIAs, devising their chassis strain as a platform for the exploration of new therapies [ref. 54].
The last couple of decades have also seen a rise in research on the therapeutic potential of once marginalized drugs due to their illicit recreational use. The review of these drugs’ legal status and growing support for clinical trials has, in many cases, showcased their effectiveness in treating a wide range of ailments. Following this wave of acceptance, researchers have worked toward their yeast-based synthesis, aiming for cost-effective, safe, and controlled production means. A great example is the biosynthesis of cannabinoids in S. cerevisiae, reported by Luo et al. in 2019 [ref. 124]. Through the application of a series of enzymes from different hosts, authors were able to produce Δ9-tetrahydrocannabinolic acid (THCA) and cannabidiolic acid (CBDA)—as well as a variety of unnatural analogs with therapeutic potential from galactose. The final THCA yields (8 mg/L) are still humble compared to yields of common cannabinoids such as THC obtained from plants. Industrial cultivation of Cannabis sativa reaches THC yields of over 100 g/m2. However, the large-scale fermentation of cannabinoids independent of Cannabis cultivation could prove valuable in countries with strict ban on the plant-derived products [ref. 124, ref. 125]. In 2020, Milne et al. published the S. cerevisiae-based production of psilocybin, the active ingredient of “magic mushrooms” [ref. 51]. Yields of approximately 627 mg/L were obtained—an impressive result when compared to the 267 mg/L observed in Aspergillus nidulans but still lagging behind yields obtained with E. coli (1.16 g/L) [ref. 126–ref. 128]. They also highlighted the production of intermediates with therapeutic value, as well as the new-to-nature analog N-acetyl-4-hydroxytryptamine. More recently, in 2022, Wong et al. showcased the complete biosynthesis of d-lysergic acid in yeast [ref. 41]. Commonly associated with psychedelic recreational drugs, d-lysergic acid is the main precursor for marketed ergot alkaloids, used for the treatment of neurological disorders. Given that both chemical synthesis and natural extraction from the fungus Claviceps purpurea face purity issues, the yeast-based synthesis of d-lysergic acid stands out as a prominent production alternative despite the current maximal yield of only 1.7 mg/L in bioreactors.
Even though the list of successfully synthesized pharmaceuticals in S. cerevisiae is long (Table 1), the number of commercialized instances remains sparse (Table 2). Several aspects can negatively interfere with the transition from academic achievements to new pharmaceutical supply chains. One of these aspects is yield. The yeast-based production of therapeutics is indubitably groundbreaking, nevertheless, in many cases, final yields remain relatively low and the overall production costs might be outcompeted by alternative natural sources such as medicinal plants. This is also true for the heterologous production of secondary metabolites from bacterial or fungal origin, for which prokaryotic organisms (e.g. E. coli, Streptomyces spp.) or filamentous fungi (e.g. Aspergillus spp.) are often preferred as hosts [ref. 129, ref. 130]. Regarding the production of therapeutically relevant proteins, S. cerevisiae holds decisive advantage over bacterial systems due to its capacity to perform PTMs that resemble higher eukaryotes’. These PTMs – in particular glycosylation – generally exert a positive impact on eukaryotic protein stability and activity, improving the production and purification processes and increasing human tolerance. Still, the remaining differences between mammalian and S. cerevisiae’s PTMs are a cause of concern. S. cerevisiae’s high-mannose N-glycosylations, often compromise protein stability in vivo, by negatively impacting their half-life and bioactivity [ref. 131]. Moreover, mannose N-glycosylations increase the allergenicity of the recombinant proteins, thus encouraging researchers to develop “humanized” versions of yeast proteins through different glycoengineering approaches [ref. 132]. As early as 1992, Nagasu et al., described how the deletion of OCH1, coding for the mannosyltransferase Och1p—a key enzyme for the transfer of the first α-1,6-mannose to the outer chain of proteins—can efficiently prevent S. cerevisiae hypermannosylation [ref. 133]. Later, in 2017, Kim et al. reported the elimination of S. cerevisiae’s mannosylphosphates via the knockout of genes MN1, MN4, and MN14 that could further improve the production of human-compatible proteins [ref. 134]. Besides these gene deletions, the overexpression of endoglycosidases or the expression of sialyltransferases were also investigated as potential strategies for the humanization of recombinant proteins in S. cerevisiae [ref. 135–ref. 138]. Nevertheless, the use of unconventional yeast species (e.g. Komagataella phaffii (Pichia pastoris) and Ogataea angusta (Hansenula polymorpha), better suited to produce human-like PTMs is often a preferred strategy to circumvent the issue [ref. 139–ref. 141]. Finally, another challenging aspect is the high competitivity of the pharmaceutical field. The influence of pre-established commercial players in global markets often makes the introduction of new manufacturing technologies difficult, especially when it comes to already marketed products. Nevertheless, as the field advances and new highly efficient engineered yeasts are developed, the prospect for the commercialization of fermented pharmaceuticals increases.
Table 2: Commercialized recombinant molecules with therapeutic activity produced in S. cerevisiae
| Compound class | Compound | Trade name | Manufacturer | Therapeutic application |
|---|---|---|---|---|
| Proteins | ||||
| Cytokine | Sargramostim | LEUKINE | Partner Therapeutics | Treatment of acute myelocytic leukemia |
| Plasma protein | Coagulation factor XIII | Tretten | Novo Nordisk | Anticoagulant, treatment of factor XIII deficiency |
| Human albumin | Recombumin | Sartorius | Stabilization and delivery of drugs and vaccines | |
| Other molecules | ||||
| Hormone | Insulin and analogs | NovoLog, Levemir, Tresiba | Novo Nordisk | Metabolic regulator. Diabetes mellitus treatment |
| Somatropin (recombinant somatotropin) | Somatropin Biopartners (Declage, LB-03002, valtropin) | LG Life Sciences | Treatment of growth hormone deficiency | |
| Glucagon and analogs | Victoza, Glucagen, REGRANEX | Novo Nordisk, Raritan | Metabolic regulator. Treatment of type 2 diabetes and hypoglycemia | |
| Platelet-derived growth factor (PDGF) | Augment, GEM 21STM, Regranex | BioMimetic, Ortho-McNeil, Luitpold | Growth factor. Treatment for a series of periodontal issues | |
| Tissue plasminogenactivator | Hirudin analogs | Refludan, Revasc, Iprivask | Bayer HealthCare, Canyon Pharmaceuticals | Anticoagulant |
Vaccines
Vaccines are preparations that induce an immune response against threatening agents—e.g. viruses, microorganisms, or cancer cells—in a prophylactic or therapeutic manner. While the first generation of vaccines consisted of inactivated or attenuated forms of the disease-causing agents, new-generation vaccines, also called subunit vaccines, contain only specific antigenic parts of the pathogen – accounting for their higher safety levels. Due to its robustness as a cell factory, S. cerevisiae plays a paramount part in the success story of subunit vaccines as one of the main organisms used for the production of recombinant proteins from pathogenic origin (Fig. 1b). In this section, we explore the yeast application in vaccinology both indirectly in the production of purified immunogenic proteins or directly, as a whole-cell vaccine preparation, detailing breakthroughs and addressing future challenges. Table 3 gathers examples of all applications.
Table 3: S. cerevisiae-based immunogens
| Immunogen class | Target class | Target | Immunogen | References |
|---|---|---|---|---|
| Purified protein | Bacteria | Bacillus anthracis (anthrax) | Protective antigen of Bacillus anthracis | [ref. 142] |
| Protozoan | Plasmodium falciparum | C-terminal fragment of major merozoite surface antigen | [ref. 143] | |
| Plasmodium vivax | Truncated circumsporozoite protein | [ref. 144] | ||
| Virus | Dengue virus | Dengue envelope domain III | [ref. 145] | |
| Newcastle disease virus | Hemagglutinin-neuraminidase | [ref. 146] | ||
| SARS-CoV-2 virus | SARS-CoV-2 spike protein receptor-binding domain (RBD) antigen | [ref. 147] | ||
| Virus-like particle (VLP) | Protozoan | Plasmodium falciparum | RTS,S/AS01 fusion antigen | [ref. 148] |
| Virus | Enterovirus 71 | Polyprotein P1 and protease 3CD from enterovirus 71 | [ref. 149] | |
| Enterovirus 71 + coxsackievirus A16 | Chimeric version of the polyprotein P1 from enterovirus 71 and the sp70 epitope of coxsackievirus A16 + protease 3CD from enterovirus 71 | [ref. 150] | ||
| Hepatitis B virus (HBV) | Surface antigens of HBV | [ref. 151, ref. 152] | ||
| Hepatitis E virus (HEV) | HEV genotype 3 and rat HEV capsid proteins | [ref. 153] | ||
| Human immunodeficiency virus (HIV) | HIV type 1 Gag protein | [ref. 154] | ||
| Human papillomavirus (HPV) | HPV16 L1 protein | [ref. 155, ref. 156] | ||
| Parvovirus B19 | Viral proteins 1 and 2 | [ref. 157] | ||
| Rotavirus | Rotavirus structural proteins VP2, VP6 and VP7 | [ref. 158] | ||
| Whole-cell | Bacteria | Mycobacterium tuberculosis | Production of a fusion of the four Rv1738, Rv2032, Rv3130, and Rv3841 proteins | [ref. 159] |
| Cancer cells | Carcinoma | Production of human carcinoembryonic antigen | [ref. 160] | |
| Carcinoma | Production of brachyury | [ref. 161] | ||
| Pancreatic cancer | Production of four different RAS proteins | [ref. 162] | ||
| Fungi | Coccidioides sp. | Live or heat-killed S. cerevisiae | [ref. 163] | |
| Aspergillus sp. | Heat-killed S. cerevisiae | [ref. 164] | ||
| Candida albicans | Heat-killed S. cerevisiae | [ref. 165] | ||
| C. albicans | Membrane display of enolase 1 from C. albicans | [ref. 166] | ||
| Virus | Hepatitis C virus (HCV) | Production of NS3-Core fusion protein | [ref. 167] | |
| SARS-CoV-2 | Membrane display of spike protein RBD | [ref. 168] |
Yeasts as cell factories for immunogens
Production of purified proteins
As exemplified in the first chapter of this review, S. cerevisiae is a very popular chassis for the production of proteins with therapeutic value and therefore a natural choice for the synthesis of immunogenic proteins. Indeed, free immunogenic proteins from viruses, bacteria, and protozoan have been recombinantly produced in this yeast for their potential application in vaccines [ref. 143–ref. 145, ref. 155]. A recent example was prompted by the 2019 outbreak of the SARS-CoV-2 coronavirus and the following pandemic. Based on the recombinant production of the receptor-binding domain of the virus’ spike protein in S. cerevisiae, a preparation called CORBEVAX in India or IndoVac in Indonesia successfully passed phase I-II clinical trials and was authorized for emergency use following a phase III superiority study [ref. 147]. Even though the combat of the COVID-19 pandemic was marked by the success of mRNA-based therapies—which are rapidly developed and potentially cope better with rapidly mutating viruses—recombinant protein vaccines, such as CORBEVAX, generally face less public hesitancy, adding to the vaccination effort.
Apart from pathogen-targeting vaccines, tumor-targeting vaccines rarely rely on recombinant proteins. For this application chemically synthesized short peptides of 8 to 12 amino acids (AAs) or long peptides of 25–35 AAs often lead to better immunogenic responses [ref. 169]. The only reported case of recombinant proteins produced in S. cerevisiae for cancer immunotherapies is that of the soluble New York esophageal squamous cell carcinoma-1 (NY-ESO-1) protein, a cancer/testis antigen only expressed in malignant or germ cells, nevertheless, the preparation failed to develop into a commercialized vaccine [ref. 170].
Production of virus-like particles
Another class of yeast-based factories for immunogens focuses on virus-like particles’ (VLPs) production. VLPs utilize the intrinsic property of viral proteins to spontaneously assemble in multimers to form the capsid, a natural vehicle of viral genetic information [ref. 171]. By producing recombinant fusion proteins containing domains of these capsid proteins, researchers can engineer empty particles resembling viruses but incapable of replication. VLPs are very promising in the field of vaccinology, especially in the fight against viruses, as they display hundreds of viral epitopes on one particle—a decisive advantage over vaccines based on free recombinant proteins.
Indeed, the first vaccines using recombinant proteins implemented worldwide were based on VLPs that spontaneously assembled into nanoparticles. They contained the surface antigen of hepatitis B virus (HBsAg) produced in yeast, aiming the prevention of hepatitis B infections [ref. 151]. These vaccines quickly replaced the previously developed serum-derived hepatitis B vaccines, preparations that were cumbersome to produce and raised safety concerns. Engerix-B from GlaxoSmithKline and Recombivax-HB from Merck were the first commercially available vaccines against hepatitis B using yeast-VLPs, quickly followed by improved VLPs-based vaccines, either adjuvanted (Heplisav-B from Dynavax GmbH, Fendrix from GlaxoSmithKline, HBVaxPro from MSD VACCINS) or containing the additional recombinant pre-S1 and pre-S2 proteins’ antigens (PreHevBrio from VBI vaccine) [ref. 172]. Since these pioneering vaccines, numerous studies reported the assembly of VLPs carrying various S. cerevisiae-produced proteins for the fight against human pathogenic viruses like hepatitis B virus (HBV), hepatitis E virus (HEV), human immunodeficiency virus (HIV), human papillomavirus (HPV), enterovirus 71, Coxsackievirus A16, parvovirus, and rotavirus [ref. 142, ref. 150, ref. 152–ref. 154, ref. 156–ref. 158]. From this long list, only Gardasil9 from Merck, a VLP-based vaccine produced in yeast to prevent HPV infection, was commercialized to this moment [ref. 173].
When it comes to non-viral infections, the only VLPs-based vaccine utilizing S. cerevisiae recombinant proteins currently on the market is RTS,S/AS01 (commercialized as Mosquirix by GlaxoSmithKline), a preparation aiming to prevent malaria, caused by the parasite Plasmodium falciparum [ref. 174]. The vaccine is recommended for children in regions of moderate to high malaria prevalence by the World Health Organization (WHO). It is based on the combination of a fusion protein called RTS containing two domains of the pre-erythrocytic circumsporozoite protein from P. falciparum (called “R” and “T”) fused to the surface antigen of the HBV (called “S”) and the surface viral antigen alone (S) that spontaneously assemble to form a VLP after purification. This approach combines the high immunogenicity of the HBV—leading to a strong immune response and therefore minimizing the need for adjuvants—and the high specificity against P. falciparum.
Besides VLPs, another class of structures increasingly used in vaccine preparation are mesoporous silica nanoparticles (MSNs) [ref. 175, ref. 176]. Contrary to VLPs, these particles are synthesized chemically and serve as carriers for recombinantly produced immunogenic proteins. Their surface can be functionalized to enhance immunogenicity and their size can also be tuned to tailor particular needs. Although the utilization of MSNs to carry S. cerevisiae-produced proteins has not been reported so far, the functionalization of MSN with glycans of yeast origin allowed for the creation of S. cerevisiae-like particles, an important step in the spread of this technology [ref. 177].
Whole yeasts as vaccines
Even though S. cerevisiae turned into a key player in vaccinology by making the production of recombinant immunogenic proteins affordable, purified protein—as well as mRNA—vaccines still face major challenges: their instability and need for cautious storage. This is mainly due to the fragile nature of proteins and mRNA—either free or in the form of VLPs. To alleviate this issue, the utilization of S. cerevisiae whole cell as a carrier for immunogens was explored. The ability of yeast cells to cope with a wide range of temperatures and environmental conditions—thriving between 20 and 37 °C—allows their convenient storage at room temperature. On top of that, S. cerevisiae cells have adjuvant-like activities and are able to elicit cellular immune responses through antigen representation by dendritic cells (DCs) [ref. 178–ref. 180]. Additionally, the faculty of S. cerevisiae to survive the harsh environmental conditions of the intestinal tract opens up the possibility of administrating the preparation orally, which could facilitate vaccination campaigns [ref. 181]. Finally, once the genetic engineering of the yeast strains has been performed, the propagation of cells is cheap and straightforward, allowing even small facilities to generate vaccines.
To this moment, however, only one antiviral vaccine based on whole S. cerevisiae cells entered phase II clinical trials: GI-5005 from Globeimmune, against hepatitis C virus (HCV) [ref. 167]. In this design, a fusion of the nonstructural protein 3 (NS3) and core viral proteins is constitutively produced to stimulate specific immunogenic responses to the virus. Another highlight is the use of protein-display strategies on S. cerevisiae to engineer whole-cell vaccines against SARS-CoV-2 [ref. 168]. In a study published by Gao et al., 2021, the full-length receptor binding domain (RBD) of the spike protein of SARS-CoV-2 was expressed on the surface of S. cerevisiae. Mice vaccinated orally with this preparation produced significant humoral mucosal and cellular immune responses. Given the high efficiency of other vaccines—the commercialization of this specific whole-cell vaccine is unrealistic, nevertheless, the knowledge gained from such projects can pave the way for the rapid development of other vaccines in the future.
When it comes to whole-cell vaccines against microorganisms, a similar approach as for GI-5005 was used for the development of GI-19007—a yeast-based preparation against the bacterium Mycobacterium tuberculosis—again developed by Globeimmune [ref. 159]. M. tuberculosis is the main causative agent of tuberculosis, the leading cause of death by infectious disease worldwide over the past decades [ref. 182]. In light of the controversies regarding the current attenuated bacteria-based vaccine—commonly known as the Bacillus Calmette-Guérin (BCG) vaccine—regarding its variable effectivity in treatment, GI-19007 could represent a good complementary therapy, as it induced survival in animal models already bearing the disease [ref. 183]. S. cerevisiae was also engineered as a vaccine targeting pathogenic fungi. The designs take advantage of the natural effect of injections of inactivated wild-type S. cerevisiae, a measure that reduces fungal burden and increases survival rates after infection with Coccidioides, Aspergillus, and Candida albicans [ref. 163–ref. 165]. In 2013, Shibasaki et al. improved the anti-candidiasis effect by engineering the membrane display of the antigen protein Eno1 (glycolytic enzyme enolase 1) from C. albicans on the surface of S. cerevisiae. The delivery of the cells as either nasal or oral vaccines increased survival rates by 60% in mice infected by C. albicans [ref. 166].
Finally, the utilization of whole S. cerevisiae cells as vaccines targeting cancer cells was also reported. In a study published by Bernstein et al., a modified yeast strain expressing the tumor-associated carcinoembryonic antigen on its membrane induced the activation and maturation of DCs in vitro and elicited immune and antitumor responses [ref. 160]. Later, Heery et al. described the development of GI-6301, a heat-killed S. cerevisiae strain producing recombinant brachyury, a transcription factor (TF) that plays a role in the development of sarcoma [ref. 161]. Following a promising phase I clinical trial, patients were enrolled in a placebo-controlled phase II study, which unfortunately failed to demonstrate a substantial therapeutic effect [ref. 184]. More recently, the therapeutic potential of S. cerevisiae strains engineered to produce mutant versions of rat sarcoma virus (RAS) proteins—called G-4000—was investigated [ref. 185]. The strategy takes advantage of the key role of RAS proteins in cell growth and survival. Recombinant mutant RAS proteins used as immunogens activate DCs and generate cell cytotoxicity against target cells expressing cancer antigens, especially pancreatic cancer. Nevertheless, a placebo-controlled phase II trial testing of G-400 therapeutic efficacy in pancreatic cancer was not successful [ref. 162].
As seen throughout this chapter, S. cerevisiae is an essential protagonist of vaccinology, and several commercial vaccines using this yeast as a cell factory are currently on the market (Table 4). Nevertheless, common drawbacks of yeast-based protein production such as their inability to add complex (e.g. human) glycosylation patterns to proteins post-translationally also apply [ref. 186]. Whole-cell vaccines circumvent stability issues of immunogens by producing and displaying proteins in situ, however, the field is still in its infancy and no commercial vaccine based on this technology is yet available. As with most medical novelties, overcoming public hesitancy is likely a challenge lying close ahead of this technology. In this regard, the development of a variety of S. cerevisiae whole-cell vaccines targeting animal diseases might provide valuable insights into their efficiency and potential associated risks once marketed [ref. 187–ref. 189].
Table 4: Commercially available vaccines using proteins recombinantly produced in S. cerevisiae
| Immunogen | Target class | Target | Tradename | Manufacturer |
|---|---|---|---|---|
| Proteins | Virus | SARS-CoV-2 virus | CORBEVAX | Biological E |
| IndoVac | Bio Farma | |||
| VLPs | Virus | Hepatitis B virus (HBV) | Recombivax HB | Merk |
| Engerix B | GlaxoSmithKline Biologicals | |||
| PreHevbri | VBI Vaccines | |||
| Heplisav0B | Dynavax Technologies Corporation | |||
| HBVaxPro | Merk Sharp & Dohme | |||
| Human papillomavirus (HPV) | Gardasil9 | Merk Sharp & Dohme | ||
| Protozoan | Plasmodium falciparum | Mosquirix | GlaxoSmithKline Biologicals |
Biosensors
Yeast-based biosensors are another class of genetically engineered microorganisms extensively explored as healthcare allies. These are whole cells equipped with a sensing and a reporter module that allow for the low-cost and accurate identification of target molecules or environmental cues (Fig. 1c). While the sensing module executes the recognition of the target, the reporter converts the sensed cue to a qualitative or quantitative signal. Yeasts are particularly amenable to sensor building. They can rely on different sensing systems, spanning from (1) TFs-based detectors—grounded on the repression or activation of promoters; (2) G-protein-coupled receptors (GPCRs)—a large class of membrane proteins highly conserved among eukaryotes, including humans; (3) protein-based affinity molecules, such as antibodies and aptamers, engineered to be displayed on the cell membrane; and (4) RNA-based biosensors, designated riboswitches, that regulate transcription, translation or mRNA decay (Fig. 2). On top of that, a large variety of reporter signals can be emitted by yeast biosensors—such as fluorescent, luminescent, colorimetric, and electrochemical—allowing them to be coupled with common clinical analysis equipment (if any) for results interpretation. Due to their capacity of long-period storage in lyophilized active form, yeasts also constitute a hot target for the development of point-of-care (POC) devices. Common medical applications of yeast-based biosensors are cost-effective diagnostic tests and biomimetic models—mainly used for drug improvement and screening of therapy targets. In addition to these, many yeast-based biosensors focus on the detection of harmful molecules in the environment. Even though such applications have arguably an indirect effect on human health, they were not included in this review—further information on this topic can be found elsewhere [ref. 190]. Table 5 gathers some of the main examples of yeast-based biosensors applied in the medical field developed so far.
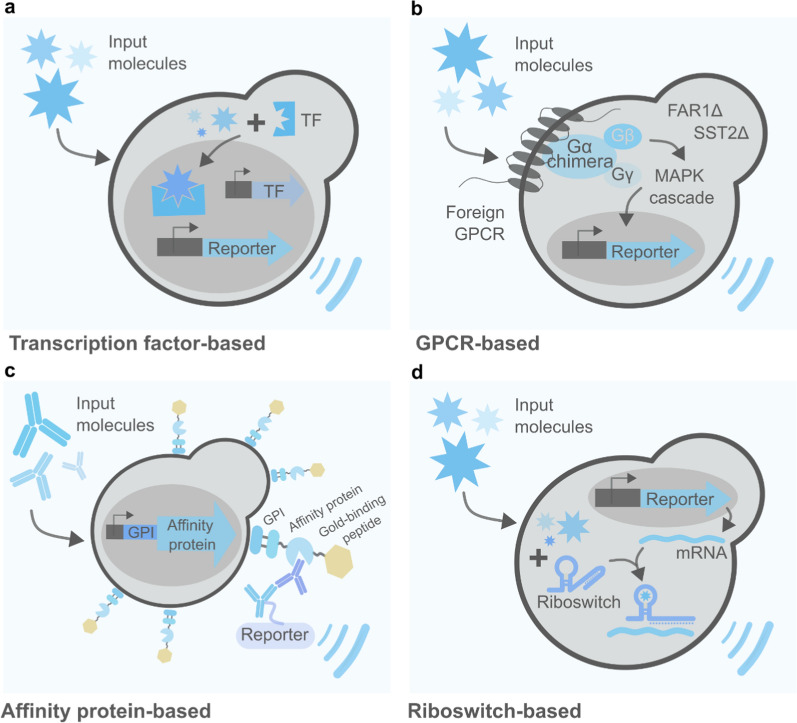
Table 5: S. cerevisiae-based biosensors for medical applications
| Sensor type | Target | Reporter | Application | References |
|---|---|---|---|---|
| Affinity molecules display | A variety of IgG-type antibodies (e.g. Human Albumin) | Biochemical | Immunoassays and affinity purification | [ref. 191, ref. 192] |
| Anti-hepatitis C Virus (HCV) core antibody | Electrochemical and fluorescent | POC detection of HVC | [ref. 193] | |
| Dengue virus nonstructural protein 1 (NS1) | Fluorescent, spectroscopic | Detection of Dengue virus | [ref. 194, ref. 195] | |
| Entamoeba histolytica cyst antigens | Fluorescent, spectroscopic, electrochemical | Detection of Entamoeba histolytica | [ref. 196–ref. 198] | |
| Invasive non-typhoid Salmonella (iNTS) antigen | Electrochemical and fluorescent | POC detection of iNTS | [ref. 199] | |
| SARS-CoV-2 spike protein | Raman signals | Detection of SARS-CoV-2 | [ref. 200] | |
| Soluble cancer protein biomarkers | Spectroscopic | Detection of cancer biomarkers | [ref. 201] | |
| G-protein-coupled receptor (GPCR) | Angiotensin (ang II) | Fluorescent | Screening for treatments of ang II receptor AGTR1-related ailments | [ref. 202] |
| Bitter tastants | Fluorescence | Development of biomimetic bitter taste-sensing systems | [ref. 203] | |
| Cannabinoids | Fluorescent, colorimetric and luminescent | Detection of endocannabinoid disease biomarkers and quality control for therapeutics manufacturing | [ref. 204, ref. 205] | |
| Cystatin C | Fluorescent | POC detection of chronic kidneydisease | [ref. 206] | |
| Fungal pathogens’ mating peptides | Colorimetric | POC detection of a variety of pathogenic fungi | [ref. 207] | |
| Light | Fluorescent | Screening of mutations associated with retinal disease | [ref. 208, ref. 209] | |
| Melatonin | Biochemical | Screening for treatments of melatonin receptor Mel1a-related ailments | [ref. 210] | |
| Neurotensin | Fluorescent | Screening for treatments of neurotensin receptor NTSR1-related ailments | [ref. 211] | |
| Odorants | Luminescent | Development of biomimetic odor-sensing systems | [ref. 212–ref. 214] | |
| Opioids | Fluorescent | Screening of new drugs that target the human opioid receptors | [ref. 215] | |
| Peptides of the calcitonin family | Biochemical | Screening for treatments of calcitonin-like peptide and adrenomedullin receptors-related ailments | [ref. 216] | |
| Serotonin | Fluorescent | Screening for optimized production, receptor characterization | [ref. 217–ref. 220] | |
| Somatostatin | Fluorescent | Screening for new drugs for acromegaly treatment | [ref. 221] | |
| Volatile organic ligands | Biochemical | Development of biomimetic systems for Caenorhabditis elegans chemoreceptors | [ref. 222] | |
| Riboswitch | Theophylline | Fluorescent | Detection of theophylline and screening of improved enzymes | [ref. 223–ref. 225] |
| Thiamine | Fluorescent | Detection of thiamine and screening of improved enzymes | [ref. 226] | |
| Transcription factor | Aromatic AAs (AAAs) | Fluorescent | Screening for optimized AAAs and muconic acid production | [ref. 227–ref. 229] |
| Differences in redox cofactors’ ratios | Fluorescent | Oxidative stress activation of live biotherapeutic products (LBPs) | [ref. 230, ref. 231] | |
| Flavonoids (e.g. Naringenin, genistein) | Fluorescent | Screening for optimized flavonoid production | [ref. 232–ref. 234] | |
| Genotoxins | Biochemical, Fluorescent | Screening of procarcinogens and other DNA-damaging compounds | [ref. 235–ref. 239] | |
| Isoprenoids (e.g. Isopentenyl pyrophosphate) | Fluorescent | Screening for optimized isoprenoid production | [ref. 240] | |
| l-arginine | Fluorescent | Screening for optimized L-arginine production | [ref. 232] | |
| Malonic acid | Fluorescent | Screening for optimized malonic acid production | [ref. 232] | |
| Malonyl-CoA | Fluorescent | Screening for optimized malonyl-CoA production | [ref. 241, ref. 242] | |
| Phenolic acids (e.g. Protocatechuic acid) | Fluorescent | Screening for optimized protocatechuic acid production | [ref. 232] |
Transcription factor-based biosensors
Exploiting TFs for the precise induction or repression of genetic pathways is a hallmark of synthetic biology. Historically, native and synthetic inducible promoters have been greatly explored for the fine-tuning of genetic pathways, allowing the incorporation of Boolean logic functions into pathway designs. The efficiency and specificity with which TFs can respond to extracellular ligands make them naturally suitable for the development of biosensors. The increasing comprehension of TFs engineering in eukaryotes, as well as the successful examples of imported prokaryotic TFs in yeast allowed for a crescent of yeast TF-based biosensors—also in the medical field (Fig. 2a).
Most of the early work on yeast-based medically relevant TF biosensors focused on the detection of DNA-damaging compounds. For that, two main sensing systems based on S. cerevisiae’s native stress-responsive promoters coupled with either fluorescent or biochemical reporters were used to generate numerous screening platforms [ref. 235–ref. 238]. Biosensors based on promoters pRAD54 and pRNR2 rely on their native function, inducing DNA repair and synthesis genes, while pHUG1 promoter-based systems take advantage of its induction by hydroxyurea, ultraviolet, and gamma radiation for the genotoxins-dependent activation of reporters. In 2016, Bui et al. improved a pRAD54-based system for the detection of both carcinogens and procarcinogens, such as polycyclic aromatic hydrocarbons and aflatoxin B1 [ref. 239]. Their design is based on the expression of human cytochrome P450 monooxygenases (CYPs), which metabolize and activate procarcinogens into genotoxins, that can ultimately induce reporters under pRAD54. Another great example of TF-based biosensors in the medical field is the work towards a S. cerevisiae-based redox biosensor, for the in vivo monitoring of oxidative stress – a biomarker for colitis. The biosensor was first established by Zhang et al., 2016, who created hybrid versions of yeast’s natural promoters with multiple copies of the redox-responsive TF Yap1p [ref. 231]. In 2021, Dacquay et al. improved the system’s dynamic range and detection limit, by employing synthetic minimal promoters in their design [ref. 230]. In another example of TF-based yeast biosensor, Skjoedt et al. developed a general approach to transplant prokaryotic transcriptional activators of the LysR-type superfamily into S. cerevisiae [ref. 232]. Setting the core of the yeast CYC1 promoter as their starting point, they identified a series of optimizations allowing the development of efficient biosensors for molecules of therapeutic interest such as cis,cis-muconic acid, naringenin, protocatechuic acid, l-arginine and malonic acid.
GPCR-based biosensors
Many therapeutically relevant yeast-based biosensors rely, however, on another class of transcription-based system: GPCRs. These are integral proteins with seven transmembrane α-helical domains, associated with a trimeric guanine nucleotide-binding protein (G protein). They constitute the largest group of membrane receptors in eukaryotes (the human genome, for instance, encodes 831 GPCRs) and are implicated in the sensing of a variety of extracellular signals—including hormones, nutrients, light, neurotransmitters, and olfactory molecules [ref. 243]. Due to their fundamental role in the maintenance of a number of basic body functions, GPCRs stand out as important therapy targets as well as subjects to basic and applied medical research. Yeast’s indigenous GPCR responsible for pheromone sensing, has long been explored to leverage yeast’s and higher eukaryotes’ signaling via the hybridization of their mating pathway with exogenous GPCRs—the first successful example being the expression of the human β2-adrenergic receptor (hβ-AR) in S. cerevisiae [ref. 244]. Together with the introduction of reporter genes under the control of natural pheromone-responsive promoters, the heterologous expression of GPCRs transforms the yeast mating pathway into a powerful biosensing platform (Fig. 2b).
Indeed, the first yeast GPCR-based biosensors for medical applications relied mainly on the substitution of the yeast STE2/3 gene—coding for the yeast pheromones GPCR receptors—by an exogenous GPCR, alongside the expression of a reporter gene under responsive promoters—mainly pFUS1 and pFIG1. Examples are biosensors for melatonin and bitter taste recognition which constitute important biomimetic platforms for drug development [ref. 203, ref. 210]. The last decade, however, has seen a rise in yeast GPCR-based biosensors of evermore complexity and effectiveness. The substitution of the yeast native G protein’s Gα subunit by chimeric versions was proven to enhance the system performance in many instances [ref. 202, ref. 208, ref. 217, ref. 221]. Such chimeric Gα subunits typically bear five C-terminal AAs corresponding to the heterologous GPCR counterpart, enhancing the communication between native and exogenous parts [ref. 220]. Additionally, the deletion or adjustment in expression levels of a series of genes related to mating function was also found to positively influence coupling. In 2019, Shaw et al. took a refactoring approach to establish a comprehensive cell model, heavily modified to retain only core components for GPCR biosensing [ref. 245]. Finally, the replacement of the fungal sterol ergosterol by cholesterol, humanizing the yeast’s membrane, was also proven to improve the functionality of yeast-based biosensors, such as in the opioid-sensing yeast platform reported by Bean et al., 2022 [ref. 215, ref. 246–ref. 248].
One of the first works to demonstrate the potential of yeast-based GPCR biosensors for POC diagnostics was that of Ostrov et al., 2017 [ref. 207]. Using an optimized chassis strain, the authors substituted S. cerevisiae’s pheromone sensing GPCR for that of ten common human and plant fungal pathogens. The engineered pheromone sensing systems were coupled with a lycopene-producing pathway, and developed into a dipstick assay, achieving a highly sensitive and specific reagent-free sensor for the surveillance of pathogens in different substrates. Another highlight is the work of Adeniran et al., 2018, who developed a POC assay for Cystatin C (a biomarker for chronic kidney disease) detection in urine [ref. 206]. This was achieved by direct evolution of the yeast native GPCR and coupling with green fluorescent protein (GFP), demonstrating a generalizable approach to sensing different peptides via substrate walking. In 2022, both Miettinen et al. and Shaw et al. revealed the parallel effort toward the establishment of biosensors for cannabinoids, aiming to detect known or unknown cannabinoid receptor agonists or to optimize microbially manufactured cannabinoid therapeutics [ref. 204, ref. 205]. While the first group focused on the development of various POC devices—including a portable luminometric biosensor based on mobile phone reading, the second group concentrated on the development of a high-throughput plate reader assay for the screening of microbially fermented THC.
Affinity protein-based biosensors
The robustness of yeast surface display techniques also accounts for relevant biosensors in the medical field. These devices are commonly based on glycosylphosphatidylinositol (GPI) carrier proteins – such as agglutinin, and flocculin, that anchor affinity proteins to the yeast’s cell wall. Instead of relying on the transcription of reporter genes, these protein-displaying cells are often coupled with either immunofluorescent or electrochemical assays as reporter systems (Fig. 2c) [ref. 249, ref. 250]. Affinity molecules-based yeast biosensors are particularly successful in the fast and inexpensive POC detection of pathogens. In 2015, Venkatesh et al. developed a dual affinity yeast, engineered to display both single-chain variable fragment (scFv) antibodies and gold-binding peptide on their surfaces, for the electrochemical impedance spectroscopy-based detection of antigens [ref. 199]. Detection of invasive non-typhoid Salmonella (iNTS) antigen served as a proof of concept of the system’s applicability. In 2016, Aronoff-Spencer et al. made use of a similar strategy for the detection of HCV core antibodies [ref. 193]. S. cerevisiae cells were engineered to display HCV core protein and gold-binding peptide repeats, which permitted the smartphone-based sensing of HCV core antibodies via a potentiostat. The development of nanoyeast-scFvs also gained attention in the past years. These are nanosized scFv-displaying yeast cell wall pieces, obtained via the mechanical fragmentation of genetically engineered whole-cells [ref. 251]. Nanoyeast-scFvs were successfully developed for the detection of a series of medically relevant proteins, such as cancer biomarkers, and human pathogen-specific antigens (e.g. Entamoeba histolytica, dengue virus, and SARS-CoV-2) [ref. 194–ref. 198, ref. 200, ref. 201].
Riboswitch-based biosensors
Finally, another class of yeast-based biosensors makes use of riboswitches for the specific sensing of ligands. These are hairpin-shaped mRNA aptamers composed of two distinct domains, originally discovered in bacteria [ref. 252, ref. 253]. While a ligand-binding domain interacts with specific target molecules, an antisense domain (expression platform), undergoes conformational changes upon ligand binding that elicit the regulation of downstream coding sequences by affecting the translation of target mRNAs. Coupling of the antisense domain with the mRNA of reporter proteins (e.g. GFP), converts riboswitches into robust biosensing platforms (Fig. 2d). In 2005, Bayer and Smolke engineered the first synthetic riboswitches in S. cerevisiae, named antiswitches [ref. 224]. For that, the RNA of interest was cloned between two hammerhead ribozymes under the control of Pol II promoters, supporting the creation of noncoding RNAs free of interfering flanking sequences. By applying an aptamer that binds the bronchodilator theophylline to their design, the authors were able to regulate GFP expression based on theophylline concentrations. The yeast-based theophylline biosensor was further improved in a series of designs—e.g. the implementation of Boolean logic gates, and the use of a FACS-based screening approach [ref. 223, ref. 254, ref. 255]. A robust theophylline biosensing platform is of great medical importance for the monitoring of administering concentrations, given that this molecule presents a narrow therapeutic index [ref. 256]. Another great example of yeast riboswitch-based biosensors for medical applications is the thiamine pyrophosphate (TPP) riboswitch. It is involved in the regulation of thiamine (vitamin B1) biosynthesis in different eukaryotic organisms (e.g. algae, plants, and filamentous fungi), and stands out as the only native eukaryotic riboswitch identified so far [ref. 257–ref. 259]. In 2018, Donovan et al. described a TPP riboswitch in the yeast Candida parapsilosis and its functional heterologous expression in S. cerevisiae [ref. 226]. By linking thiamine concentrations with a purple fluorescent protein (yEmRFP), the authors obtained an S. cerevisiae whole-cell biosensor for thiamine.
Even though yeast-based biosensors represent effective alternatives for the low-cost and precise detection of many cues of medical relevance, their application remains limited. As with many technological novelties, a main hindrance to the broader use of these devices is the existence of other well-established methods. Medical institutions often prefer keeping to expensive but highly accurate diagnostic techniques, such as mass spectrometry even if these methods come with longer waiting times. Another great challenge is safety, especially when it comes to POC systems. Unlike cell factories, in which synthesized molecules (often subjected to further purification steps) comprise the final product, yeast-based biosensing devices are composed of live cells (except for Nanoyeast-scFvs). As genetically modified organisms (GMOs), the proper containment or selective removal of cells to avoid environmental contamination is imperative. Additionally, the amenability of yeast cells to isolation and culturing techniques raises concerns about intellectual property protection. To overcome these major obstacles, the biocontainment of yeast-based biosensors via genetically embedded safeguard systems has been explored [ref. 260–ref. 264]. Given the particular importance of those systems in the design and implementation of LBPs, more information can be found in the following section.
Live biotherapeutic products
Aside from the indirect application of yeast cells in medicine—e.g. for the in-vitro production of pharmaceuticals or the POC diagnosis of diseases—the direct application of engineered microorganisms is an emerging therapeutic modality. Known as LBPs, these are live microorganisms genetically engineered for the prevention or treatment of diseases and metabolic disbalances in the human body. The in situ activity provided by LBPs holds many advantages over systemic therapy approaches. The necessary concentration of effector molecules to achieve a certain therapeutic effect is significantly reduced, leading to fewer side effects. Microorganisms can also be engineered to produce more than one effector molecule, achieving combinatory therapy. On top of that, LBP-based therapies often contribute to the maintenance of microbiota homeostasis—a relevant aspect of human health, that is constantly disturbed by systemic treatments. Finally, LBPs can also be engineered to work in a rational manner, delivering treatment only when specific disease cues are sensed, instead of permanently producing effectors.
To achieve effective treatment, LBPs often comprise different operating modules that can either act synergistically or execute separate functions. Common modules are (1) effector elements, that enable the production and secretion of specific therapeutic molecules; (2) sensing elements, that act as small diagnostic devices, sensing disease states and adjusting the function of other modules accordingly; (3) biocontainment elements, that act as safeguard systems, preventing the LBP escape to the external environment; and (4) supplementary modules, such as motility or attachment devices, that enable or enhance specific LBP designs. As speculated by Claesen and Fischbach, the next generation of LBPs will potentially include several such modules, enabling the performance of diagnosis tasks and their rational translation into appropriate treatments as well as self-elimination from the human host once treatment is completed [ref. 265]. Alternatively, consortia of engineered microorganisms armed with cooperating modules can also be employed and extend the therapeutic possibilities. To this moment, however, most examples rely on single or double modules—mainly sensing and effector modules– as proof of concept for the effective treatment of specific target diseases.
Most LBPs are designed to treat diseases and metabolic disbalances in the gut due to their easiness of administration via the oral route. For this reason, probiotic microorganisms are often the preferred chassis, given their historical application in the treatment of intestinal ailments. When it comes to yeast-based LBPs, Saccharomyces boulardii (Sb)—short for Saccharomyces cerevisiae var. boulardii, commonly sold under the trade names Ultra Levure and Florastor (Biocodex)—is often used. While classified as an S. cerevisiae strain, sharing more than 99% identity with the S288c reference genome, Sb is better adapted to the gastrointestinal tract, being resistant to high temperatures, bile acids, and low pH [ref. 266–ref. 268]. In 2016 Liu et al. set the basis for the genomic engineering of Sb [ref. 269]. They developed auxotrophic Sb strains and enabled the production and secretion of human lysozyme, showcasing that therapeutic molecules can be synthesized in Sb. Durmusoglu et al. 2021, explored the strain properties further for its application as an LBP chassis [ref. 270]. The authors characterized several synthetic parts commonly used for gene expression in S. cerevisiae in Sb, and demonstrated the strains’ capacity for in vivo construction of biosynthetic pathways into plasmids, via the assembly of a multigenic β-carotene pathway from linearized parts. Most importantly, the authors evaluated Sb’s colonization patterns in the mouse gut in varied contexts, demonstrating that the production of heterologous proteins (i.e. β-carotene) by engineered Sb is possible inside the host.
The advancements in the understanding of Sb’s genetic engineering and colonization patterns, as well as the success of LBP’s applications in other probiotic species—mostly E. coli Nissle 1917—opened the doors for the development of yeast-based LBPs in the last years [ref. 271–ref. 273]. In 2020, Chen et al. described the development of an Sb-based immunotherapy against Clostridioides difficile infection (CDI), as an alternative treatment for the growing number of C. difficile antibiotic-resistant strains, a major cause of recurring CDI [ref. 274, ref. 275]. The authors engineered an effector module in Sb, to constitutively secrete a tetravalent antibody, designated as ABAB, targeting C. difficile major virulence factors—toxins TcdA and TcdB. Both prophylactic and remediating effects of the LBP were demonstrated in vivo in challenges with C. difficile spores. Another great example is the work of Scott et al., 2021, in which S. cerevisiae was engineered for the smart self-tunable treatment of inflammatory bowel disease (IBD) [ref. 276]. The design takes advantage of the production of extracellular adenosine triphosphate (eATP) in the gastrointestinal tract of affected individuals, as a disease-associated signal, to drive the production of an effector module through a biosensing element. To enable sensing, the P2Y2 GPCR, which detects both eATP and extracellular uridine triphosphate (eUTP) was coupled with S. cerevisiae’s pheromone sensing pathway and engineered for increased sensitivity to eATP. An effector module assists the conversion of pro-inflammatory eATP into immunosuppressive adenosine via the production of a potato (Solanum tuberosum) apyrase in an eATP-dependent manner. Ultimately, the engineered cells significantly reduced intestinal inflammation and colitis-associated fibrosis and dysbiosis in mice. More recently, Hedin et al., 2023, have developed an Sb-based LBP with antiobesity effects [ref. 277]. The authors engineered an effector module for the constitutive production and secretion of Exendin-4, an agonist of the glucagon-like peptide-1 (GLP-1) receptor. When combined with exposure to cold (8 °C), the authors observed a 25% suppression in appetite, as well as a fourfold loss in weight of mice administered the engineered strains. Table 6 showcases the most relevant yeast-based LBPs developed so far.
Table 6: S. cerevisiae-based live biotherapeutic products
| Therapeutic application | S. cerevisiae strain | Effector module | Biosensing module | Biocontainment module | References |
|---|---|---|---|---|---|
| Anti Clostridioides difficile infection (CDI) | BY4741 and S. cerevisiae var. Boulardii (Sb) MYA-796 | Production and secretion of a tetra-specific antibody targeting C. difficile toxins | – | – | [ref. 274] |
| Antilisterial | Sb CNCM I-745 | Production and secretion of leucocin C | – | – | [ref. 278] |
| Anti-obesity | Sb MYA-796 | Production and secretion of exendin-4 | – | – | [ref. 277] |
| Antitumoral | Sb MYA-797 | Production and secretion of immune checkpoint inhibitors | – | – | [ref. 279] |
| Broad | BY4741 | – | GPCR-based sensing of serotonin | – | [ref. 218] |
| Sb MYA-796 | Production of β-carotene | – | – | [ref. 270] | |
| Sb MYA-796 | Production and secretion of human lysozyme | – | – | [ref. 269, ref. 280] | |
| Sb MYA-796 | – | Different transcription factor-based sensors (i.g. galactose, copper, and IPTG) | – | [ref. 281] | |
| Sb MYA-796 | – | – | Auxotrophy to thiamine combined with cold-induced growth defect | [ref. 282] | |
| S288C and Sb CNCM I-745 | – | Transcription factor-based sensing of redox imbalance | – | [ref. 230] | |
| Delivery of immunogenic proteins | Sb CNCM I-745 | Production and secretion of Ovalbumin | – | – | [ref. 283] |
| Prebiotics production | Sb MYA-796 | Production and secretion of neoagarooligosaccharides by an endo-type β-agarase | – | – | [ref. 284] |
| Treatment of inflammatory bowel disease (IBD) | BS016 | Production and secretion of an apyrase with ATPase activity | eATP sensing via the expression of human GPCR P2Y2 | – | [ref. 276] |
| BY4741 | Production and secretion of lactic acid | – | – | [ref. 285] | |
| Sb Enterol | Production and secretion of lactic acid | – | – | [ref. 286, ref. 287] | |
| Sb MYA-796 | Production and secretion of atrial natriuretic peptide (ANP) | – | – | [ref. 288] |
Even though the development of LBPs is rather recent, they stand out as a promising alternative in the prevention and treatment of a wide range of diseases. This is especially true for therapy regimes that demand the direct delivery of molecules with short half-lives to the target tissue, as well as treatment of pathogenic microorganisms prone to develop antibiotic resistance. However, transitioning this technology from academic research to commercial products faces obstacles. A major concern is the lack of understanding of yeast colonization patterns in different individuals (with different microbiota). While there’s a growing number of characterization assays of both laboratory and probiotic S. cerevisiae in what concerns mice and rat strains, our knowledge of their residence time and inter/intra niche interactions in the human gut remains sparse—even for commercialized wild-type probiotic strains [ref. 270, ref. 289, ref. 290]. Another great obstacle is the development of robust biocontainment systems. Numerous highly operational safeguard systems exist for S. cerevisiae industrial applications, however, these are often dependent on triggering molecules or environmental cues that could lead to cross-feeding in the context of LBPs [ref. 260–ref. 263]. Hedin et al., 2023 have set the basis for the development of biocontainment modules for yeast-based LBPs. Their concept is based on an auxotrophy to the vitamin thiamine coupled with a cold-sensitive strategy based on the knockout of gene BTS1 coding for the yeast geranylgeranyl diphosphate synthase [ref. 282]. Yet, an LBP-suitable system that complies with the NIH escape frequency guidelines (under 1 in 108 escapees per colony forming unit) and isn’t prone to cross-feeding remains necessary [ref. 291]. Finally, public acceptance of genetically engineered LBPs stands out as an important challenge ahead. Similar to transgenic crop plants, which still face low consumer adoption rates more than two decades after their introduction, the commercialization of LBPs is likely to encounter resistance [ref. 292]. The establishment of a defined and specific regulatory framework for such endeavors is primordial to push forward the application of both wild-type and genetically engineered LBPs [ref. 293]. As part of this effort, Rouanet et al. 2020 have published a comprehensive roadmap for their safety assessment [ref. 294].
Concluding remarks and future perspectives
Synthetic biology enabled the development of a variety of medically relevant yeast strains. Whether used as cell factories for the production of pharmaceuticals and vaccines or applied as whole living cells as biosensors and live therapeutics, the positive impact of genetically engineered yeasts on human health is outstanding. Nevertheless, their commercialization still faces challenges.
When it comes to the heterologous production of glycoproteins of high eukaryotic origin, their so-called “humanization” via efficient post-translational processing remains a major obstacle. Indeed, the market share of microbially produced biotherapeutics shrinks by the year, giving room to the expanding trade of mammalian-based therapeutics. While before the 2000’s microbial biopharmaceuticals added to more than half of the total production of new active ingredients, in 2022 this number dropped to approximately 28% [ref. 295, ref. 296]. The tendency is driven by major improvements in genetic engineering and production strategies (e.g. media composition and cell line development) in mammalian cells, but not only. The production of full-length monoclonal antibodies currently dominates the global market of biopharmaceuticals, and due to their high complexity, mammalian cells are preferred [ref. 295, ref. 296]. This trend is, however, not applicable to all therapeutics. Microorganisms, and notably yeast, remain major production platforms for hormones, growth factors, interleukins, and vaccines [ref. 295].
For whole-cell applications, on the other hand, the baker’s yeast combination of GRAS status, robust growth, and ease of genetic engineering is often unmatched. When it comes to whole-cell biosensors, the ease of transfer of mammalian GPCRs to S. cerevisiae makes them a first-rate platform in the study of human GPCR-related diseases and the development of diagnostic assays for GPCR-binding molecules, allowing for quick high-throughput screening methods [ref. 297]. As for LBP approaches, while other probiotic chassis such as the bacteria E. coli Nissle 1917 and Lactococcus lactis have been successfully engineered for the treatment of a series of ailments, safety concerns of the first and the difficulty of engineering of the latter also make the case for S. cerevisiae-based LBPs [ref. 298]. The lack of trustworthy pharmacokinetics and risk assessment techniques in most regulatory agencies remain main hindrances to the commercialization of these endeavors, nevertheless, S. cerevisiae will most likely stand out as a key player in the LBP market in the decades to come.
References
- J Legras, D Merdinoglu, J Cornuet, F Karst. Bread, beer and wine: Saccharomyces cerevisiae diversity reflects human history. Mol Ecol, 2007. [DOI | PubMed]
- D Cavalieri, PE McGovern, DL Hartl, R Mortimer, M Polsinelli. Evidence for S. cerevisiae fermentation in ancient wine. J Mol Evol, 2003. [DOI | PubMed]
- PE McGovern, J Zhang, J Tang, Z Zhang, GR Hall, RA Moreau. Fermented beverages of pre- and proto-historic China. Proc Natl Acad Sci USA, 2004. [DOI | PubMed]
- JW Szostak, TL Orr-Weaver, RJ Rothstein, FW Stahl. The double-strand-break repair model for recombination. Cell, 1983. [DOI | PubMed]
- TL Orr-Weaver, JW Szostak, RJ Rothstein. Yeast transformation: a model system for the study of recombination. Proc Natl Acad Sci USA, 1981. [DOI | PubMed]
- A Goffeau, BG Barrell, H Bussey, RW Davis, B Dujon, H Feldmann. Life with 6000 genes. Science, 1996. [DOI | PubMed]
- LA Gilbert, MH Larson, L Morsut, Z Liu, GA Brar, SE Torres. CRISPR-mediated modular RNA-guided regulation of transcription in eukaryotes. Cell, 2013. [DOI | PubMed]
- JE Dicarlo, JE Norville, P Mali, X Rios, J Aach, GM Church. Genome engineering in Saccharomyces cerevisiae using CRISPR-Cas systems. Nucl Acids Res, 2013. [DOI | PubMed]
- M Jinek, K Chylinski, I Fonfara, M Hauer, JA Doudna, E Charpentier. A Programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity. Science, 2012. [DOI | PubMed]
- WH Mager, J Winderickx. Yeast as a model for medical and medicinal research. Trends Pharmacol Sci, 2005. [DOI | PubMed]
- KR Love, NC Dalvie, JC Love. The yeast stands alone: the future of protein biologic production. Curr Opin Biotechnol, 2018. [DOI | PubMed]
- P Patra, M Das, P Kundu, A Ghosh. Recent advances in systems and synthetic biology approaches for developingel cell-factories in non-conventional yeasts. Biotechnol Adv, 2021. [DOI | PubMed]
- A Adeniran, M Sherer, KEJ Tyo. Yeast-based biosensors: design and applications. FEMS Yeast Res, 2015. [PubMed]
- S Rebello, A Abraham, A Madhavan, R Sindhu, P Binod, AK Babu. Non-conventional yeast cell factories for sustainable bioprocesses. FEMS Microbiol Lett, 2018. [DOI]
- DV Goeddel, DG Kleid, F Bolivar, HL Heyneker, DG Yansura, R Crea. Expression in Escherichia coli of chemically synthesized genes for human insulin. Proc Natl Acad Sci USA, 1979. [DOI | PubMed]
- NA Baeshen, MN Baeshen, A Sheikh, RS Bora, MMM Ahmed, HAI Ramadan. Cell factories for insulin production. Microb Cell Fact, 2014. [DOI | PubMed]
- L Thim, MT Hansen, AR Sørensen. Secretion of human insulin by a transformed yeast cell. S Lett, 1987
- L Thim, MT Hansen, K Norris, I Hoegh, E Boel, J Forstrom. Secretion and processing of insulin precursors in yeast. Proc Natl Acad Sci USA, 1986. [DOI | PubMed]
- D Beran, EAM Gale, JS Yudkin. The insulin market reaches 100. Diabetologia, 2022. [DOI | PubMed]
- O Spadiut, S Capone, F Krainer, A Glieder, C Herwig. Microbials for the production of monoclonal antibodies and antibody fragments. Trends Biotechnol, 2014. [DOI | PubMed]
- YJ Lee, KJ Jeong. Challenges to production of antibodies in bacteria and yeast. J Biosci Bioeng, 2015. [DOI | PubMed]
- PK Das, A Sahoo, VD Veeranki. Recombinant monoclonal antibody production in yeasts: challenges and considerations. Int J Biol Macromol, 2024. [DOI | PubMed]
- KJ Jeong, SH g, N Velmurugan. Recombinant antibodies: Engineering and production in yeast and bacterial hosts. Biotechnol J, 2011. [DOI | PubMed]
- AJ Moody, F Norris, K Norris, MT Hansen, L Thim. The secretion of glucagon by transformed yeast strains. S Lett, 1987
- N Riehl-Bellon, D Carvallo, M Acker, A Van Dorsselaer, M quet, G Loison. Purification and biochemical characterization of recombinant hirudin produced by Saccharomyces cerevisiae. Biochem, 1989. [DOI | PubMed]
- T Achstetter, M Nguyen-Juilleret, A Findeli, M Merkamm, Y Lemoine. A new signal peptide useful for secretion of heterologous proteins from yeast and its application for synthesis of hirudin. Gene, 1992. [DOI | PubMed]
- S Yamano, T Ishii, M Nakagawa, H Ikenaga, N Misawa. Metabolic engineering for production of β -carotene and lycopene in Saccharomyces cerevisiae. Biosci Biotechnol Biochem, 1994. [DOI | PubMed]
- C Duport, R Spagnoli, E Degryse, D Pompon. Self-sufficient biosynthesis of pregnenolone and progesterone in engineered yeast. Nat Biotechnol, 1998. [DOI | PubMed]
- FM Szczebara, C Chandelier, C Villeret, A Masurel, S Bourot, C Duport. Total biosynthesis of hydrocortisone from a simple carbon source in yeast. Nat Biotechnol, 2003. [DOI | PubMed]
- G Damiani, TS McCormick, LO Leal, MA Ghannoum. Recombinant human granulocyte macrophage-colony stimulating factor expressed in yeast (sargramostim): a potential ally to combat serious infections. Clin Immunol, 2020. [DOI | PubMed]
- P Jagadeeswaran, P Haas. Synthesis of human coagulation factor XIII in yeast. Gene, 1990. [DOI | PubMed]
- D Sleep, GP Belfield, AR Goodey. The secretion of human serum albumin from the yeast Saccharomyces cerevisiae using five different leader sequences. Nat Biotechnol, 1990. [DOI]
- M Kálmán, I Cserpän, G Bajszár, A Dobi, É Horváth, C Pázmán. Synthesis of a gene for human serum albumin and its expression in Saccharomyces cerevisiae. Nucl Acids Res, 1990. [DOI | PubMed]
- CJ Finnis, T Payne, J Hay, N Dodsworth, D Wilkinson, P Morton. High-level production of animal-free recombinant transferrin from Saccharomyces cerevisiae. Microb Cell Fact, 2010. [DOI | PubMed]
- A Meta, H Nakatake, T Imamura, C Nozaki, K Sugimura. High-yield production and characterization of biologically active recombinantotinin expressed in Saccharomyces cerevisiae. Protein Expr Purif, 2009. [DOI | PubMed]
- K Norris, F Norris, SE Bjørn, I Diers, LC Peterson. Aprotinin andotinin analogues expressed in yeast. Biol Chem Hoppe Seyler, 1990. [PubMed]
- Y Bourbonnais, C Larouche, GM Tremblay. Production of full-length human pre-elafin, an elastase specific inhibitor, from yeast requires the absence of a functional Yapsin 1 (Yps1p) endoprotease. Protein Expr Purif, 2000. [DOI | PubMed]
- T Liu, Y Gou, B Zhang, R Gao, C Dong, M Qi. Construction of ajmalicine and sanguinarine de novo biosynthetic pathways using stable integration sites in yeast. Biotechnol Bioeng, 2022. [DOI | PubMed]
- S Galanie, CD Smolke. Optimization of yeast-based production of medicinal protoberberine alkaloids. Microb Cell Fact, 2015. [DOI | PubMed]
- J Han, S Li. De novo biosynthesis of berberine and halogenated benzylisoquinoline alkaloids in Saccharomyces cerevisiae. Commun Chem, 2023. [DOI | PubMed]
- G Wong, LR Lim, YQ Tan, MK Go, DJ Bell, PS Freemont. Reconstituting the complete biosynthesis of D-lysergic acid in yeast. Nat Commun, 2022. [DOI | PubMed]
- P Srinivasan, CD Smolke. Biosynthesis of medicinal tropane alkaloids in yeast. Nature, 2020. [DOI | PubMed]
- P Srinivasan, CD Smolke. Engineering cellular metabolite transport for biosynthesis of computationally predicted tropane alkaloid derivatives in yeast. Proc Natl Acad Sci USA, 2021. [DOI | PubMed]
- Y Li, CD Smolke. Engineering biosynthesis of the anticancer alkaloid noscapine in yeast. Nat Commun, 2016. [DOI | PubMed]
- KM Hawkins, CD Smolke. Production of benzylisoquinoline alkaloids in Saccharomyces cerevisiae. Nat Chem Biol, 2008. [DOI | PubMed]
- S Galanie, K Thodey, IJ Trenchard, M Filsinger Interrante, CD Smolke. Complete biosynthesis of opioids in yeast. Science, 2015. [DOI | PubMed]
- K Thodey, S Galanie, CD Smolke. A microbial biomanufacturing platform for natural and semisynthetic opioids. Nat Chem Biol, 2014. [DOI | PubMed]
- E Fossati, L Narcross, A Ekins, JP Falgueyret, VJJ Tin. Synthesis of morphinan alkaloids in Saccharomyces cerevisiae. PLoS ONE, 2015. [DOI | PubMed]
- WC DeLoache, ZN Russ, L Narcross, AM Gonzales, VJJ tin, JE Dueber. An enzyme-coupled biosensor enables (S)-reticuline production in yeast from glucose. Nat Chem Biol, 2015. [DOI | PubMed]
- OK Jamil, A Cravens, JT Payne, CY Kim, CD Smolke. Biosynthesis of tetrahydropapaverine and semisynthesis of papaverine in yeast. Proc Natl Acad Sci USA, 2022. [DOI | PubMed]
- N Milne, P Thomsen, N Knudsen, P Rubaszka, M Kristensen, I Borodina. Metabolic engineering of Saccharomyces cerevisiae for the de novo production of psilocybin and related tryptamine derivatives. Metab Eng, 2020. [DOI | PubMed]
- SA Bradley, FG Hansson, BJ Lehka, D Rago, P Pinho, H Peng. Yeast platforms for production and screening of bioactive derivatives of Rauwolscine. ACS Synth Biol, 2024. [DOI | PubMed]
- E Fossati, A Ekins, L Narcross, Y Zhu, JP Falgueyret, GAW Beaudoin. Reconstitution of a 10-gene pathway for synthesis of the plant alkaloid dihydrosanguinarine in Saccharomyces cerevisiae. Nat Commun, 2014. [DOI | PubMed]
- SA Bradley, BJ Lehka, FG Hansson, KB Adhikari, D Rago, P Rubaszka. Biosynthesis of natural and halogenated plant monoterpene indole alkaloids in yeast. Nat Chem Biol, 2023. [DOI | PubMed]
- AM Ehrenworth, S Sarria, P Peralta-Yahya. Pterin-dependent mono-oxidation for the microbial synthesis of a modified monoterpene indole alkaloid. ACS Synth Biol, 2015. [DOI | PubMed]
- S Brown, M Clastre, V Courdavault, SE O’Connor. De novo production of the plant-derived alkaloid strictosidine in yeast. Proc Natl Acad Sci USA, 2015. [DOI | PubMed]
- A Campbell, P Bauchart, ND Gold, Y Zhu, V De Luca, VJJ tin. Engineering of a nepetalactol-producing platform strain of Saccharomyces cerevisiae for the production of plant seco-iridoids. ACS Synth Biol, 2016. [DOI | PubMed]
- P Srinivasan, CD Smolke. Engineering a microbial biosynthesis platform for de novo production of tropane alkaloids. Nat Commun, 2019. [DOI | PubMed]
- Y Ping, X Li, W You, G Li, M Yang, W Wei. De novo production of the plant-derived tropine and pseudotropine in yeast. ACS Synth Biol, 2019. [DOI | PubMed]
- J Zhang, LG Hansen, O Gudich, K Viehrig, LMM Lassen, L Schrübbers. A microbial supply chain for production of the anti-cancer drug vinblastine. Nature, 2022. [DOI | PubMed]
- Y Qu, MLAE Easson, J Froese, R Simionescu, T Hudlicky, V DeLuca. Completion of the seven-step pathway from tabersonine to the anticancer drug precursor vindoline and its assembly in yeast. Proc Natl Acad Sci USA, 2015. [DOI | PubMed]
- M McKeague, YH Wang, A Cravens, MN Win, CD Smolke. Engineering a microbial platform for de novo biosynthesis of diverse methylxanthines. Metab Eng, 2016. [DOI]
- C George-Nascimento, A Gyenes, SM Halloran, J Merryweather, P Valenzuela, KS Steimer. Characterization of recombinant human epidermal growth factor produced in yeast. Biochem, 1988. [DOI | PubMed]
- ML Bayne, J Applebaum, GG Chicchi, NS Hayes, BG Green, MA Cascieri. Expression, purification and characterization of recombinant human insulin-like growth factor I in yeast. Gene, 1988. [DOI | PubMed]
- SM Germann, SA Baallal Jacobsen, K Schneider, SJ Harrison, NB Jensen, X Chen. Glucose-based microbial production of the hormone melatonin in yeast Saccharomyces cerevisiae. Biotechnol J, 2016. [DOI | PubMed]
- M Calderón-Cacia, P Tekamp-Olson, J Allen, C George-Nascimento. Incomplete process of recombinant human Platelet-derived growth factor produced in yeast and its effect on the biological activity. Biochem Biophys Res Commun, 1992. [DOI | PubMed]
- Y Wang, L Xue, Y Li, Y Zhu, B Yang, X Wang. High-level secretory production of recombinant human platelet-derived growth factor-BB by Saccharomyces cerevisiae under the non-selective conditions. Appl Biochem Microbiol, 2009. [DOI]
- T Tomoko, I Shigenori, G Hideyuki, K Kazumi, O Eiko, I Morio. Expression of a synthetic human growth hormone gene in yeast. Gene, 1985. [DOI | PubMed]
- V Peterkova, I Arslanoglu, E Bolshova-Zubkovskaya, T Romer, D Zdravkovic, J Kratzsch. A randomized, double-blind study to assess the efficacy and safety of valtropin, a biosimilar growth hormone, in children with growth hormone deficiency. Horm Res Paediatr, 2007. [DOI]
- AR Awan, BA Blount, DJ Bell, WM Shaw, JCH Ho, RM McKiernan. Biosynthesis of the antibiotic nonribosomal peptide penicillin in baker’s yeast. Nat Commun, 2017. [DOI | PubMed]
- M Eichenberger, A Hansson, D Fischer, L Dürr, M Naesby. De novo biosynthesis of anthocyanins in Saccharomyces cerevisiae. FEMS Yeast Res, 2018. [DOI | PubMed]
- X Liu, J Cheng, G Zhang, W Ding, L Duan, J Yang. Engineering yeast for the production of breviscapine by genomic analysis and synthetic biology approaches. Nat Commun, 2018. [DOI | PubMed]
- M Eichenberger, BJ Lehka, C Folly, D Fischer, S tens, E Simón. Metabolic engineering of Saccharomyces cerevisiae for de novo production of dihydrochalcones with known antioxidant, antidiabetic, and sweet tasting properties. Metab Eng, 2017. [DOI | PubMed]
- F Koopman, J Beekwilder, B Crimi, A Van Houwelingen, RD Hall, D Bosch. De novo production of the flavonoid naringenin in engineered Saccharomyces cerevisiae. Microb Cell Fact, 2012. [DOI | PubMed]
- J Mao, MT Mohedano, J Fu, X Li, Q Liu, J Nielsen. Fine-tuning of p-coumaric acid synthesis to increase (2S)-naringenin production in yeast. Metab Eng, 2023. [DOI | PubMed]
- X Lyu, KR Ng, JL Lee, R k, WN Chen. Enhancement of naringenin biosynthesis from tyrosine by metabolic engineering of Saccharomyces cerevisiae. J Agric Food Chem, 2017. [DOI | PubMed]
- S Isogai, N Okahashi, R Asama, T Nakamura, T Hasunuma, F Matsuda. Synthetic production of prenylated naringenins in yeast using promiscuous microbial prenyltransferases. Metab Eng Commun, 2021. [DOI | PubMed]
- A Rodriguez, T Strucko, SG Stahlhut, M Kristensen, DK Svenssen, J Forster. Metabolic engineering of yeast for fermentative production of flavonoids. Bioresour Technol, 2017. [DOI | PubMed]
- E Trantas, N Panopoulos, F Ververidis. Metabolic engineering of the complete pathway leading to heterologous biosynthesis of various flavonoids and stilbenoids in Saccharomyces cerevisiae. Metab Eng, 2009. [DOI | PubMed]
- W Xu, M Liu, H Li, J Chen, J Zhou. De novo synthesis of chrysin in Saccharomyces cerevisiae. J Agric Food Chem, 2024. [DOI | PubMed]
- T An, G Lin, Y Liu, L Qin, Y Xu, X Feng. De novo biosynthesis of anticarcinogenic icariin in engineered yeast. Metab Eng, 2023. [DOI | PubMed]
- Q Liu, T Yu, X Li, Y Chen, K Campbell, J Nielsen. Rewiring carbon metabolism in yeast for high level production of aromatic chemicals. Nat Commun, 2019. [DOI | PubMed]
- A Rodriguez, KR Kildegaard, M Li, I Borodina, J Nielsen. Establishment of a yeast platform strain for production of p-coumaric acid through metabolic engineering of aromatic amino acid biosynthesis. Metab Eng, 2015. [DOI | PubMed]
- J Jiang, H Yin, S Wang, Y Zhuang, S Liu, T Liu. Metabolic engineering of Saccharomyces cerevisiae for high-level production of salidroside from glucose. J Agric Food Chem, 2018. [DOI | PubMed]
- M Suástegui, W Guo, X Feng, Z Shao. Investigating strain dependency in the production of aromatic compounds in Saccharomyces cerevisiae. Biotechnol Bioeng, 2016. [DOI | PubMed]
- SY Shin, NS Han, YC Park, MD Kim, JH Seo. Production of resveratrol from p-coumaric acid in recombinant Saccharomyces cerevisiae expressing 4-coumarate:coenzyme A ligase and stilbene synthase genes. Enzyme Microb Technol, 2011. [DOI | PubMed]
- L Meng, M Diao, Q Wang, L Peng, J Li, N Xie. Efficient biosynthesis of resveratrol via combining phenylalanine and tyrosine pathways in Saccharomyces cerevisiae. Microb Cell Fact, 2023. [DOI | PubMed]
- M Li, K Schneider, M Kristensen, I Borodina, J Nielsen. Engineering yeast for high-level production of stilbenoid antioxidants. Sci Rep, 2016. [DOI | PubMed]
- J Chen, F Fan, G Qu, J Tang, Y Xi, C Bi. Identification of Absidia orchidis steroid 11β-hydroxylation system and its application in engineering Saccharomyces cerevisiae for one-step biotransformation to produce hydrocortisone. Metab Eng, 2020. [DOI | PubMed]
- DK Ro, EM Paradise, M Ouellet, KJ Fisher, KL Newman, JM Ndungu. Production of the antimalarial drug precursor artemisinic acid in engineered yeast. Nature, 2006. [DOI | PubMed]
- PJ Westfall, DJ Pitera, JR Lenihan, D Eng, FX Woolard, R Regentin. Production of amorphadiene in yeast, and its conversion to dihydroartemisinic acid, precursor to the antimalarial agent artemisinin. Proc Natl Acad Sci USA, 2012. [DOI | PubMed]
- DK Ro, M Ouellet, EM Paradise, H Burd, D Eng, CJ Paddon. Induction of multiple pleiotropic drug resistance genes in yeast engineered to produce an increased level of anti-malarial drug precursor, artemisinic acid. BMC Biotechnol, 2008. [DOI | PubMed]
- M Tang, X Xu, Y Liu, J Li, G Du, X Lv. Combinatorial metabolic engineering for improving betulinic acid biosynthesis in Saccharomyces cerevisiae. ACS Synth Biol, 2024. [DOI | PubMed]
- C Ignea, A Athanasakoglou, A Andreadelli, M Apostolaki, M Iakovides, EG Stephanou. Overcoming the plasticity of plant specialized metabolism for selective diterpene production in yeast. Sci Rep, 2017. [DOI | PubMed]
- U Scheler, W Brandt, A Porzel, K Rothe, D Manzano, D Božić. Elucidation of the biosynthesis of carnosic acid and its reconstitution in yeast. Nat Commun, 2016. [DOI | PubMed]
- W Xie, X Lv, L Ye, P Zhou, H Yu. Construction of lycopene-overproducing Saccharomyces cerevisiae by combining directed evolution and metabolic engineering. Metab Eng, 2015. [DOI | PubMed]
- Y Chen, W Xiao, Y Wang, H Liu, X Li, Y Yuan. Lycopene overproduction in Saccharomyces cerevisiae through combining pathway engineering with host engineering. Microb Cell Fact, 2016. [DOI | PubMed]
- J Jin, Y Wang, M Yao, X Gu, B Li, H Liu. Astaxanthin overproduction in yeast by strain engineering and new gene target uncovering. Biotechnol Biofuels, 2018. [DOI | PubMed]
- C Ignea, FA Trikka, AK Nikolaidis, P Georgantea, E Ioannou, S Loupassaki. Efficient diterpene production in yeast by engineering Erg20p into a geranylgeranyl diphosphate synthase. Metab Eng, 2015. [DOI | PubMed]
- Z Dai, B Wang, Y Liu, M Shi, D Wang, X Zhang. Producing aglycons of ginsenosides in bakers’ yeast. Sci Rep, 2014. [DOI | PubMed]
- P Wang, W Wei, W Ye, X Li, W Zhao, C Yang. Synthesizing ginsenoside Rh2 in Saccharomyces cerevisiae cell factory at high-efficiency. Cell Discov, 2019. [DOI | PubMed]
- P Wang, Y Wei, Y Fan, Q Liu, W Wei, C Yang. Production of bioactive ginsenosides Rh2 and Rg3 by metabolically engineered yeasts. Metab Eng, 2015. [DOI | PubMed]
- C Ignea, MH Raadam, MS Motawia, AM Makris, CE Vickers, SC Kampranis. Orthogonal monoterpenoid biosynthesis in yeast constructed on an isomeric substrate. Nat Commun, 2019. [DOI | PubMed]
- C Ignea, M Pontini, ME Maffei, AM Makris, SC Kampranis. Engineering monoterpene production in yeast Using a synthetic dominant negative geranyl diphosphate synthase. ACS Synth Biol, 2014. [DOI | PubMed]
- B Peng, LK Nielsen, SC Kampranis, CE Vickers. Engineered protein degradation of farnesyl pyrophosphate synthase is an effective regulatory mechanism to increase monoterpene production in Saccharomyces cerevisiae. Metab Eng, 2018. [DOI | PubMed]
- GZ Jiang, MD Yao, Y Wang, L Zhou, TQ Song, H Liu. Manipulation of GES and ERG20 for geraniol overproduction in Saccharomyces cerevisiae. Metab Eng, 2017. [DOI | PubMed]
- C Zhang, M Li, GR Zhao, W Lu. Alpha-terpineol production from an engineered Saccharomyces cerevisiae cell factory. Microb Cell Fact, 2019. [DOI | PubMed]
- T Moses, J Pollier, L Almagro, D Buyst, M Van Montagu, MA Pedreño. Combinatorial biosynthesis of sapogenins and saponins in Saccharomyces cerevisiae using a C-16α hydroxylase from Bupleurum falcatum. Proc Natl Acad Sci USA, 2014. [DOI | PubMed]
- Y Liu, X Zhao, F Gan, X Chen, K Deng, SA Crowe. Complete biosynthesis of QS-21 in engineered yeast. Nature, 2024. [DOI | PubMed]
- MZ Ding, HF Yan, LF Li, F Zhai, LQ Shang, Z Yin. Biosynthesis of taxadiene in Saccharomyces cerevisiae: selection of geranylgeranyl diphosphate synthase directed by a computer-aided docking strategy. PLoS ONE, 2014. [DOI | PubMed]
- BJ Lee, M Weyers, RK Haynes, F Van Der Kooy. Discovery of artemisinin in Artemisia annua, its current production, and relevance to sub-Saharan Africa. S Afr J Bot, 2023. [DOI]
- L Zhao, Y Zhu, H Jia, Y Han, X Zheng, M Wang. From plant to yeast—advances in biosynthesis of artemisinin. Molecules, 2022. [DOI | PubMed]
- Y Li, Y Yang, L Li, K Tang, X Hao, G Kai. Advanced metabolic engineering strategies for increasing artemisinin yield in Artemisia annua L. Hortic Res., 2024. [DOI]
- M Peplow. Synthetic biology’s first malaria drug meets market resistance. Nature, 2016. [DOI | PubMed]
- M Peplow. Malaria drug made in yeast causes market ferment. Nature, 2013. [DOI | PubMed]
- L Cao, D Teo, Y Wang, Q Ye, C Liu, C Ding. Advancements in microbial cell engineering for benzylisoquinoline alkaloid production. ACS Synth Biol, 2024. [DOI | PubMed]
- V Salim, SA Jarecki, M Vick, R Miller. Advances in metabolic engineering of plant monoterpene indole alkaloids. Biology, 2023. [DOI | PubMed]
- F Ghirga, A Bonamore, L Calisti, I D’Acquarica, M Mori, B Botta. Green routes for the production of enantiopure benzylisoquinoline alkaloids. Int J Mol Sci, 2017. [DOI | PubMed]
- GAW Beaudoin, PJ Facchini. Benzylisoquinoline alkaloid biosynthesis in opium poppy. Planta, 2014. [DOI | PubMed]
- G Pasquali, DD Porto, AG Fett-Neto. Metabolic engineering of cell cultures versus whole plant complexity in production of bioactive monoterpene indole alkaloids: Recent progress related to old dilemma. J Biosci Bioeng, 2006. [DOI | PubMed]
- T Winzer, M Kern, AJ King, TR Larson, RI Teodor, SL Donninger. Morphinan biosynthesis in opium poppy requires a P450-oxidoreductase fusion protein. Science, 2015. [DOI | PubMed]
- SC Farrow, JM Hagel, GAW Beaudoin, DC Burns, PJ Facchini. Stereochemical inversion of (S)-reticuline by a cytochrome P450 fusion in opium poppy. Nat Chem Biol, 2015. [DOI | PubMed]
- IJ Trenchard, MS Siddiqui, K Thodey, CD Smolke. De novo production of the key branch point benzylisoquinoline alkaloid reticuline in yeast. Metab Eng, 2015. [DOI | PubMed]
- X Luo, MA Reiter, L d’Espaux, J Wong, CM Denby, A Lechner. Complete biosynthesis of cannabinoids and their unnatural analogues in yeast. Nature, 2019. [DOI | PubMed]
- R Backer, T Schwinghamer, P Rosenbaum, V McCarty, S Eichhorn Bilodeau, D Lyu. Closing the yield gap for cannabis: a meta-analysis of factors determining cannabis yield. Front Plant Sci, 2019. [DOI]
- AM Adams, NA Kaplan, Z Wei, JD Brinton, CS Monnier, AL Enacopol. In vivo production of psilocybin in E. coli. Metab Eng, 2019. [DOI | PubMed]
- S evska, S Weiser, Y Huang, J Lin, S Hoefgen, K Jojić. Optimized psilocybin production in tryptophan catabolism-repressed fungi. Microb Biotechnol, 2024. [DOI | PubMed]
- S Hoefgen, J Lin, J Fricke, MC Stroe, DJ Mattern, JE Kufs. Facile assembly and fluorescence-based screening method for heterologous expression of biosynthetic pathways in fungi. Metab Eng, 2018. [DOI | PubMed]
- SE Unkles, V Valiante, DJ Mattern, AA Brakhage. Synthetic biology tools for bioprospecting of natural products in eukaryotes. Chem Biol, 2014. [DOI | PubMed]
- J Liu, X Wang, G Dai, Y Zhang, X Bian. Microbial chassis engineering drives heterologous production of complex secondary metabolites. Biotechnol Adv, 2022. [DOI | PubMed]
- S Wildt, TU Gerngross. The humanization of N-glycosylation pathways in yeast. Nat Rev Microbiol, 2005. [DOI | PubMed]
- A Al-Ghouleh, R Johal, IK Sharquie, M Emara, H Harrington, F Shakib. The glycosylation pattern of common allergens: the recognition and uptake of Der p 1 by epithelial and dendritic cells Is carbohydrate dependent. PLoS ONE, 2012. [DOI]
- T Nagasu, Y Shimma, Y Nakanishi, J Kuromitsu, K Iwama, K Nakayama. Isolation of new temperature-sensitive mutants of Saccharomyces cerevisiae deficient in mannose outer chain elongation. Yeast, 1992. [DOI | PubMed]
- YH Kim, JY Kang, JY Gil, SY Kim, KK Shin, HA Kang. Abolishment of N-glycan mannosylphosphorylation in glyco-engineered Saccharomyces cerevisiae by double disruption of MNN4 and MNN14 genes. Appl Microbiol Biotechnol, 2017. [DOI | PubMed]
- MA Piirainen, H Salminen, AD Frey. Production of galactosylated complex-type N-glycans in glycoengineered Saccharomyces cerevisiae. Appl Microbiol Biotechnol, 2022. [DOI | PubMed]
- S Ryckaert, V Tens, KD Vusser, R Contreras. Development of a S. cerevisiae whole cell biocatalyst for in vitro sialylation of oligosaccharides. J Biotechnol, 2005. [DOI | PubMed]
- A Shenoy, S Yalamanchili, AR Davis, AW Barb. Expression and display of glycoengineered antibodies and antibody fragments with an engineered yeast Ssrain. Antibodies, 2021. [DOI | PubMed]
- F Parsaie Nasab, M Aebi, G Bernhard, AD Frey. A combined system for engineering glycosylation efficiency and glycan structure in Saccharomyces cerevisiae. Appl Environ Microbiol, 2013. [DOI | PubMed]
- RK Bretthauer. Genetic engineering of Pichia pastoris to humanize N-glycosylation of proteins. Trends Biotechnol, 2003. [DOI | PubMed]
- MW Kim. Characterization of N-linked oligosaccharides assembled on secretory recombinant glucose oxidase and cell wall mannoproteins from the methylotrophic yeast Hansenula polymorpha. Glycobiology, 2003. [DOI | PubMed]
- Y Song, MH Choi, JN Park, MW Kim, EJ Kim, HA Kang. Engineering of the yeast Yarrowia lipolytica for the production of glycoproteins lacking the outer-chain mannose residues of N-glycans. Appl Environ Microbiol, 2007. [DOI | PubMed]
- R Hepler, R Kelly, T Mcneely, H Fan, M Losada, H George. A recombinant 63-kDa form of Bacillus anthracis protective antigen produced in the yeast Saccharomyces cerevisiae provides protection in rabbit and primate inhalational challenge models of anthrax infection. Vaccine, 2006. [DOI | PubMed]
- SP Chang, HL Gibson, CT Lee-Ng, PJ Barr, GS Hui. A carboxyl-terminal fragment of Plasmodium falciparum gp195 expressed by a recombinant baculovirus induces antibodies that completely inhibit parasite growth. J Immunol, 1992. [DOI | PubMed]
- WE Collins, RS Nussenzweig, WR Ballou, TK Ruebush, EH Nardin, JD Chulay. Immunization of Saimiri Sciureus Boliviensis with recombinant vaccines based on the circumsporozoite protein of Plasmodium Vivax. Am J Trop Med Hyg, 1989. [DOI | PubMed]
- NL Nguyen, JM Kim, JA Park, SM Park, YS g, MS Yang. Expression and purification of an immunogenic dengue virus epitope using a synthetic consensus sequence of envelope domain III and Saccharomyces cerevisiae. Protein Expr Purif, 2013. [DOI | PubMed]
- SA Khulape, HK Maity, DC Pathak, CM Mohan, S Dey. Antigenic validation of recombinant hemagglutinin-neuraminidase protein of Newcastle disease virus expressed in Saccharomyces cerevisiae. Acta Virol, 2015. [DOI | PubMed]
- PJ Hotez, R Adhikari, WH Chen, YL Chen, P Gillespie, NY Islam. From concept to delivery: a yeast-expressed recombinant protein-based COVID-19 vaccine technology suitable for global access. Expert Rev Vaccines, 2023. [DOI | PubMed]
- J Cohen, V Nussenzweig, J Vekemans, A Leach. From the circumsporozoite protein to the RTS, S/AS candidate vaccine. Hum Vaccine, 2010. [DOI]
- HY Li, JF Han, CF Qin, R Chen. Virus-like particles for enterovirus 71 produced from Saccharomyces cerevisiae potently elicits protective immune responses in mice. Vaccine, 2013. [DOI | PubMed]
- H Zhao, HY Li, JF Han, YQ Deng, SY Zhu, XF Li. Novel recombinant chimeric virus-like particle is immunogenic and protective against both enterovirus 71 and coxsackievirus A16 in mice. Sci Rep, 2015. [DOI | PubMed]
- P Valenzuela, A Medina, WJ Rutter, G Ammerer, BD Hall. Synthesis and assembly of hepatitis B virus surface antigen particles in yeast. Nature, 1982. [DOI | PubMed]
- N Hadiji-Abbes, M tin, W Benzina, H Karray-Hakim, C Gergely, A Gargouri. Extraction and purification of hepatitis B virus-like M particles from a recombinant Saccharomyces cerevisiae strain using alumina powder. J Virol Methods, 2013. [DOI | PubMed]
- M Simanavicius, PL Tamosiunas, R Petraityte-Burneikiene, R Johne, RG Ulrich, A Zvirbliene. Generation in yeast and antigenic characterization of hepatitis E virus capsid protein virus-like particles. Appl Microbiol Biotechnol, 2018. [DOI | PubMed]
- N Tomo, T Goto, Y Morikawa. Trans-packaging of human immunodeficiency virus type 1 genome into Gag virus-like particles in Saccharomyces cerevisiae. Microb Cell Fact, 2013. [DOI | PubMed]
- MA Park, HJ Kim, HJ Kim. Optimum conditions for production and purification of human papillomavirus type 16 L1 protein from Saccharomyces cerevisiae. Protein Expr Purif, 2008. [DOI | PubMed]
- SN Kim, HS Jeong, SN Park, HJ Kim. Purification and immunogenicity study of human papillomavirus type 16 L1 protein in Saccharomyces cerevisiae. J Virol Methods, 2007. [DOI | PubMed]
- RR Penkert, NS Young, SL Surman, RE Sealy, J Rosch, PR Dormitzer. Saccharomyces cerevisiae -derived virus-like particle parvovirus B19 vaccine elicits binding and neutralizing antibodies in a mouse model for sickle cell disease. Vaccine, 2017. [DOI | PubMed]
- WA Rodríguez-Limas, KE Tyo, J Nielsen, OT Ramírez, LA Palomares. Molecular and process design for rotavirus-like particle production in Saccharomyces cerevisiae. Microb Cell Fact, 2011. [DOI | PubMed]
- TH King, CA Shanley, Z Guo, D Bellgrau, T Rodell, S Furney. GI-19007, Saccharomyces cerevisiae-based therapeutic vaccine against tuberculosis. Clin Vaccine Immunol, 2017. [DOI | PubMed]
- MB Bernstein, M Chakraborty, EK Wansley, Z Guo, A Franzusoff, S Mostböck. Recombinant Saccharomyces cerevisiae (yeast-CEA) as a potent activator of murine dendritic cells. Vaccine, 2008. [DOI | PubMed]
- CR Heery, BH Singh, M Rauckhorst, JL té, RN Donahue, I Grenga. Phase I trial of a yeast-based therapeutic cancer vaccine (GI-6301) targeting the transcription factor brachyury. Cancer Immunol Res, 2015. [DOI | PubMed]
- P Muscarella, T Bekaii-Saab, K McIntyre, A Rosemurgy, SB Ross, DA Richards. A phase 2 randomized placebo-controlled adjuvant trial of GI-4000, a recombinant yeast expressing mutated RAS proteins in patients with resected pancreas cancer. J Pancreat Cancer, 2021. [DOI | PubMed]
- J Capilla, KV Clemons, M Liu, HB Levine, DA Stevens. Saccharomyces cerevisiae as a vaccine against coccidioidomycosis. Vaccine, 2009. [DOI | PubMed]
- M Liu, J Capilla, ME Johansen, D Alvarado, M tinez, V Chen. Saccharomyces as a vaccine against systemic aspergillosis: ‘the friend of man’ a friend again?. J Med Microbiol, 2011. [DOI | PubMed]
- M Liu, KV Clemons, ME Johansen, M tinez, V Chen, DA Stevens. Saccharomyces as a vaccine against systemic candidiasis. Immunol Invest, 2012. [DOI | PubMed]
- S Shibasaki, W Aoki, T Nomura, A Miyoshi, S Tafuku, T Sewaki. An oral vaccine against candidiasis generated by a yeast molecular display system. Pathog Dis, 2013. [DOI | PubMed]
- AA Haller, GM Lauer, TH King, C Kemmler, V Fiolkoski, Y Lu. Whole recombinant yeast-based immunotherapy induces potent T cell responses targeting HCV NS3 and Core proteins. Vaccine, 2007. [DOI | PubMed]
- T Gao, Y Ren, S Li, X Lu, H Lei. Immune response induced by oral administration with a Saccharomyces cerevisiae-based SARS-CoV-2 vaccine in mice. Microb Cell Fact, 2021. [DOI | PubMed]
- AJ Stephens, NA Burgess-Brown, S Jiang. Beyond just peptide antigens: the complex world of peptide-based cancer vaccines. Front Immunol, 2021. [DOI]
- A Wadle, A Mischo, S Strahl, H Nishikawa, G Held, F Neumann. NY-ESO-1 protein glycosylated by yeast induces enhanced immune responses. Yeast, 2010. [DOI | PubMed]
- DT Le, KM Müller. In vitro assembly of virus-like particles and their applications. Life, 2021. [DOI | PubMed]
- M Mironova, MG Ghany. Hepatitis B vaccine: four decades on. Vaccines, 2024. [DOI | PubMed]
- ZQ Toh, J Kosasih, FM Russell, SM Garland, EK Mulholland, PV Licciardi. Recombinant human papillomavirus nonavalent vaccine in the prevention of cancers caused by human papillomavirus. Infect Drug Resist, 2019. [DOI | PubMed]
- Efficacy and safety of RTS, S/AS01 malaria vaccine with or without a booster dose in infants and children in Africa: final results of a phase 3, individually randomised, controlled trial. Lancet, 2015. [DOI | PubMed]
- KT Mody, A Popat, D Mahony, AS Cavallaro, C Yu, N Mitter. Mesoporous silica nanoparticles as antigen carriers and adjuvants for vaccine delivery. Nanoscale, 2013. [DOI | PubMed]
- A Yu, X Dai, Z Wang, H Chen, B Guo, L Huang. Recent advances of mesoporous silica as a platform for cancer immunotherapy. Biosensors, 2022. [DOI | PubMed]
- G Ma, F Li, X Wang, Q Li, Y Hong, Q Wei. A bionic yeast tumor vaccine using the co-loading strategy to prevent post-operative tumor recurrence. ACS Nano, 2023. [DOI | PubMed]
- AC Stubbs, KS tin, C Coeshott, SV Skaates, DR Kuritzkes, D Bellgrau. Whole recombinant yeast vaccine activates dendritic cells and elicits protective cell-mediated immunity. Nat Med, 2001. [DOI | PubMed]
- DL Williams, HA Pretus, RB McNamee, EL Jones, HE Ensley, IW Browder. Development of a water-soluble, sulfated (1 → 3)-β-d-glucan biological response modifier derived from Saccharomyces cerevisiae. Carbohydr Res, 1992. [DOI | PubMed]
- M Rios-Hernandez, NJ Dos-Santos, JL Bello-Gárciga, M Pedroso. Immunopharmacological studies of beta-1,3-glucan. Arch Med Res, 1994. [PubMed]
- KWY Kwong, Y Xin, NCY Lai, JCC Sung, KC Wu, YK Hamied. Oral vaccines: a better future of immunization. Vaccines, 2023. [DOI | PubMed]
- Global, regional, and national age-sex specific mortality for 264 causes of death, 1980–2016: a systematic analysis for the Global burden of disease study 2016. Lancet, 2017. [DOI | PubMed]
- A Ahmed, S Rakshit, V Adiga, M Dias, P Dwarkanath, G D’Souza. A century of BCG: impact on tuberculosis control and beyond. Immunol Rev, 2021. [DOI | PubMed]
- PJ DeMaria, M Bilusic, DM Park, CR Heery, RN Donahue, RA Madan. Randomized, double-blind, placebo-controlled phase II study of yeast-brachyury vaccine (GI-6301) in combination with standard-of-care radiotherapy in locally advanced, unresectable chordoma. Oncologist, 2021. [DOI | PubMed]
- A Cohn, MA Morse, B O’Neil, S Whiting, C Coeshott, J Ferraro. Whole recombinant Saccharomyces cerevisiae yeast expressing Ras mutations as treatment for patients with solid rumors Bbearing Ras mutations: results from a phase 1 trial. J Immunother, 2018. [DOI | PubMed]
- TU Gerngross. Advances in the production of human therapeutic proteins in yeasts and filamentous fungi. Nat Biotechnol, 2004. [DOI | PubMed]
- H Lei, X Lu, S Li, Y Ren. High immune efficacy against different avian influenza H5N1 viruses due to oral administration of a Saccharomyces cerevisiae-based vaccine in chickens. Sci Rep, 2021. [DOI | PubMed]
- L Wang, M Yang, S Luo, G Yang, X Lu, J Lu. Oral vaccination of recombinant Saccharomyces cerevisiae expressing ORF132 induces protective immunity against cyprinid herpesvirus-2. Vaccines, 2023. [DOI | PubMed]
- H Li, D Hua, Q Qu, H Cao, Z Feng, N Liu. Oral immunization with recombinant Saccharomyces cerevisiae expressing viral capsid protein 2 of infectious bursal disease virus induces unique specific antibodies and protective immunity. Vaccines, 2023. [DOI | PubMed]
- S Jarque, M Bittner, L Blaha, K Hilscherova. Yeast biosensors for detection of environmental pollutants: current state and limitations. Trends Biotechnol, 2016. [DOI | PubMed]
- Y Nakamura, S Shibasaki, M Ueda, A Tanaka, H Fukuda, A Kondo. Development ofel whole-cell immunoadsorbents by yeast surface display of the IgG-binding domain. Appl Microbiol Biotechnol, 2001. [DOI | PubMed]
- R Shimojyo, H Furukawa, H Fukuda, A Kondo. Preparation of yeast strains displaying IgG binding domain ZZ and enhanced green fluorescent protein forel antigen detection systems. J Biosci Bioeng, 2003. [DOI | PubMed]
- E Aronoff-Spencer, AG Venkatesh, A Sun, H Brickner, D Looney, DA Hall. Detection of hepatitis C core antibody by dual-affinity yeast chimera and smartphone-based electrochemical sensing. Biosens Bioelectron, 2016. [DOI | PubMed]
- F Farokhinejad, RE Lane, RJ Lobb, S Edwardraja, A Wuethrich, CB Howard. Generation of nanoyeast single-chain variable fragments as high-avidity biomaterials for dengue virus detection. ACS Biomater Sci Eng, 2021. [DOI | PubMed]
- F Farokhinejad, J Li, LE Hugo, CB Howard, A Wuethrich, M Trau. Detection of Dengue virus 2 with single infected mosquito resolution using yeast affinity bionanofragments and plasmonic SERS nanoboxes. Anal Chem, 2022. [DOI | PubMed]
- YS Grewal, MJA Shiddiky, LJ Spadafora, GA Cangelosi, M Trau. Nano-yeast–scFv probes on screen-printed gold electrodes for detection of Entamoeba histolytica antigens in a biological matrix. Biosens Bioelectron, 2014. [DOI | PubMed]
- Y Wang, S Rauf, YS Grewal, LJ Spadafora, MJA Shiddiky, GA Cangelosi. Duplex microfluidic SERS detection of pathogen antigens with nanoyeast single-chain variable fragments. Anal Chem, 2014. [DOI | PubMed]
- YS Grewal, MJA Shiddiky, SA Gray, KM Weigel, GA Cangelosi, M Trau. Label-free electrochemical detection of an Entamoeba histolytica antigen using cell-free yeast-scFv probes. Chem Commun, 2013. [DOI]
- AG Venkatesh, A Sun, H Brickner, D Looney, DA Hall, E Aronoff-Spencer. Yeast dual-affinity biobricks: progress towards renewable whole-cell biosensors. Biosens Bioelectron, 2015. [DOI | PubMed]
- J Li, A Wuethrich, S Edwardraja, RJ Lobb, S Puttick, S Rose. Amplification-free SARS-CoV-2 detection using nanoyeast-scFv and ultrasensitive plasmonic nanobox-integrated nanomixing microassay. Anal Chem, 2021. [DOI | PubMed]
- J Li, J Wang, YS Grewal, CB Howard, LJ Raftery, S Mahler. Multiplexed SERS detection of soluble cancer protein biomarkers with gold–silver alloy nanoboxes and nanoyeast single-chain variable fragments. Anal Chem, 2018. [DOI | PubMed]
- Y Nakamura, J Ishii, A Kondo. Construction of a yeast-based signaling biosensor for human angiotensin II type 1 receptor via functional coupling between Asn295-mutated receptor and Gpa1/Gi3 chimeric Gα. Biotechnol Bioeng, 2014. [DOI | PubMed]
- T Sugawara, K Ito, M Shiroishi, N Tokuda, H Asada, T Yurugi-Kobayashi. Fluorescence-based optimization of human bitter taste receptor expression in Saccharomyces cerevisiae. Biochem Biophys Res Commun, 2009. [DOI | PubMed]
- K Miettinen, N Leelahakorn, A Almeida, Y Zhao, LR Hansen, IE Nikolajsen. A GPCR-based yeast biosensor for biomedical, biotechnological, and point-of-use cannabinoid determination. Nat Commun, 2022. [DOI | PubMed]
- WM Shaw, Y Zhang, X Lu, AS Khalil, G Ladds, X Luo. Screening microbially produced Δ9-tetrahydrocannabinol using a yeast biosensor workflow. Nat Commun, 2022. [DOI | PubMed]
- A Adeniran, S Stainbrook, JW Bostick, KEJ Tyo. Detection of a peptide biomarker by engineered yeast receptors. ACS Synth Biol, 2018. [DOI | PubMed]
- N Ostrov, M Jimenez, S Billerbeck, J Brisbois, J Matragrano, A Ager. A modular yeast biosensor for low-cost point-of-care pathogen detection. Sci Adv, 2017. [DOI | PubMed]
- BM Scott, SK Chen, N Bhattacharyya, AY Moalim, SV Plotnikov, E Heon. Coupling of human rhodopsin to a yeast signaling pathway enables characterization of mutations associated with retinal disease. Genetics, 2019. [DOI | PubMed]
- BM Scott, LE Wybenga-Groot, CJ McGlade, E Heon, SG Peisajovich, BSW Chang. Screening of chemical libraries using a yeast model of retinal disease. SLAS Discov, 2019. [DOI | PubMed]
- T Kokkola, MA Watson, J White, S Dowell, SM Foord, JT Laitinen. Mutagenesis of human Mel1amelatonin Receptor expressed in yeast reveals domains important for receptor function. Biochem Biophys Res Commun, 1998. [DOI | PubMed]
- H Hashi, Y Nakamura, J Ishii, A Kondo. Modifying expression modes of human neurotensin receptor type 1 alters sensing capabilities for agonists in yeast signaling biosensor. Biotechnol J, 2018. [DOI]
- Y Fukutani, T Nakamura, M Yorozu, J Ishii, A Kondo, M Yohda. The N-terminal replacement of an olfactory receptor for the development of a Yeast-based biomimetic odor sensor. Biotechnol Bioeng, 2012. [DOI | PubMed]
- Y Fukutani, A Hori, S Tsukada, R Sato, J Ishii, A Kondo. Improving the odorant sensitivity of olfactory receptor-expressing yeast with accessory proteins. Anal Chem, 2015
- J Minic, M Persuy, E Godel, J Aioun, I Connerton, R Salesse. Functional expression of olfactory receptors in yeast and development of a bioassay for odorant screening. S J, 2005
- BDM Bean, CJ Mulvihill, RK Garge, DR Boutz, O Rousseau, BM Floyd. Functional expression of opioid receptors and other human GPCRs in yeast engineered to produce human sterols. Nat Commun, 2022. [DOI | PubMed]
- JJ Miret, L Rakhilina, L Silverman, B Oehlen. Functional expression of heteromeric calcitonin gene-related peptide and adrenomedullin receptors in yeast. J Biol Chem, 2002. [DOI | PubMed]
- Y Nakamura, J Ishii, A Kondo. Applications of yeast-based signaling sensor for characterization of antagonist and analysis of site-directed mutants of the human serotonin 1A receptor. Biotechnol Bioeng, 2015. [DOI | PubMed]
- B Lengger, EE Hoch-Schneider, CN Jensen, T Jakočiu̅nas, AA Petersen, TM Frimurer. Serotonin G protein-coupled receptor-based biosensing modalities in yeast. ACS Sens., 2022. [DOI | PubMed]
- AM Ehrenworth, T Claiborne, P Peralta-Yahya. Medium-throughput screen of microbially produced serotonin via a G-protein-coupled receptor-based sensor. Biochem, 2017. [DOI | PubMed]
- AJ Brown, SL Dyos, MS Whiteway, JHM White, MAEA Watson, M zioch. Functional coupling of mammalian receptors to the yeast mating pathway usingel yeast/mammalian G protein alpha-subunit chimeras. Yeast, 2000. [DOI | PubMed]
- Y Iguchi, J Ishii, H Nakayama, A Ishikura, K Izawa, T Tanaka. Control of signalling properties of human somatostatin receptor subtype-5 by additional signal sequences on its amino-terminus in yeast. J Biochem, 2010. [DOI | PubMed]
- M Tehseen, M Dumancic, L Briggs, J Wang, A Berna, A Anderson. Functional coupling of a nematode chemoreceptor to the yeast pheromone response pathway. PLoS ONE, 2014. [DOI | PubMed]
- MN Win, CD Smolke. A modular and extensible RNA-based gene-regulatory platform for engineering cellular function. Proc Natl Acad Sci USA, 2007. [DOI | PubMed]
- TS Bayer, CD Smolke. Programmable ligand-controlled riboregulators of eukaryotic gene expression. Nat Biotechnol, 2005. [DOI | PubMed]
- JK Michener, CD Smolke. High-throughput enzyme evolution in Saccharomyces cerevisiae using a synthetic RNA switch. Metab Eng, 2012. [DOI | PubMed]
- PD Donovan, LM Holland, L Lombardi, AY Coughlan, DG Higgins, KH Wolfe. TPP riboswitch-dependent regulation of an ancient thiamin transporter in Candida. PLoS Genet, 2018. [DOI | PubMed]
- JM Leavitt, JM Wagner, CC Tu, A Tong, Y Liu, HS Alper. biosensor-enabled directed evolution to improve muconic acid production in Saccharomyces cerevisiae. Biotechnol J, 2017. [DOI]
- JM Leavitt, A Tong, J Tong, J Pattie, HS Alper. Coordinated transcription factor and promoter engineering to establish strong expression elements in Saccharomyces cerevisiae. Biotechnol J, 2016. [DOI | PubMed]
- J Zhang, SD Petersen, T Radivojevic, A Ramirez, A Pérez-Manríquez, E Abeliuk. Combining mechanistic and machine learning models for predictive engineering and optimization of tryptophan metabolism. Nat Commun, 2020. [DOI | PubMed]
- LC Dacquay, DR McMillen. Improving the design of an oxidative stress sensing biosensor in yeast. FEMS Yeast Res, 2021. [DOI]
- J Zhang, N Sonnenschein, TPB Pihl, KR Pedersen, MK Jensen, JD Keasling. Engineering an NADPH/NADP + Redox Biosensor in Yeast. ACS Synth Biol, 2016. [DOI | PubMed]
- ML Skjoedt, T Snoek, KR Kildegaard, D Arsovska, M Eichenberger, TJ Goedecke. Engineering prokaryotic transcriptional activators as metabolite biosensors in yeast. Nat Chem Biol, 2016. [DOI | PubMed]
- D Liu, MS Sica, J Mao, LFI Chao, V Siewers. A p -coumaroyl-CoA biosensor for dynamic regulation of naringenin biosynthesis in Saccharomyces cerevisiae. ACS Synth Biol, 2022. [DOI | PubMed]
- LFI Chao, D Liu, V Siewers. A highly selective cell-based fluorescent biosensor for genistein detection. Eng Microbiol, 2023. [DOI]
- D Averbeck, S Averbeck. Induction of the genes RAD54 and RNR2 by varoius DNA damaging agents in Saccharomyces cerevisiae. Mutat Res, 1994. [DOI | PubMed]
- GM Cole, D Schild, ST Lovett, RK Mortimer. Regulation of RAD54 – and RAD52-lacZ Gene Fusions in Saccharomyces cerevisiae in Response to DNA Damage. Mol Cell Biol, 1987. [PubMed]
- MG Benton, NR Glasser, SP Palecek. The utilization of a Saccharomyces cerevisiae HUG1P-GFP promoter–reporter construct for the selective detection of DNA damage. Mutat Res, 2007. [DOI | PubMed]
- RM Walmsley, N Billinton, WD Heyer. Green fluorescent protein as a reporter for the DNA damage-induced gene RAD54 in Saccharomyces cerevisiae. Yeast, 1997. [DOI | PubMed]
- VN Bui, TTH Nguyen, CT Mai, Y Bettarel, TY Hoang, TTL Trinh. Procarcinogens—determination and evaluation by yeast-based biosensor transformed with plasmids incorporating RAD54 reporter construct and cytochrome P450 genes. PLoS ONE, 2016. [DOI | PubMed]
- HH Chou, JD Keasling. Programming adaptive control to evolve increased metabolite production. Nat Commun, 2013. [DOI | PubMed]
- S Li, T Si, M Wang, H Zhao. Development of a synthetic malonyl-CoA sensor in Saccharomyces cerevisiae for intracellular metabolite monitoring and genetic screening. ACS Synth Biol, 2015. [DOI | PubMed]
- F David, J Nielsen, V Siewers. Flux control at the malonyl-CoA node through hierarchical dynamic pathway regulation in Saccharomyces cerevisiae. ACS Synth Biol, 2016. [DOI | PubMed]
- AS Hauser, MM Attwood, M Rask-Andersen, HB Schiöth, DE Gloriam. Trends in GPCR drug discovery: new agents, targets and indications. Nat Rev Drug Discov, 2017. [DOI | PubMed]
- K King, HG Dohlman, J Thorner, MG Caron, RJ Lefkowitz. Control of yeast mating signal transduction by a mammalian β2-adrenergic receptor and Gsα subunit. Science, 1990. [DOI | PubMed]
- WM Shaw, H Yamauchi, J Mead, GOF Gowers, DJ Bell, D Öling. Engineering a model cell for rational tuning of GPCR signaling. Cell, 2019. [DOI | PubMed]
- SM Kitson, W Mullen, RJ Cogdell, RM Bill, NJ Fraser. GPCR production in ael yeast strain that makes cholesterol-like sterols. Methods, 2011. [DOI | PubMed]
- S Morioka, T Shigemori, K Hara, H Morisaka, K Kuroda, M Ueda. Effect of sterol composition on the activity of the yeast G-protein-coupled receptor Ste2. Appl Microbiol Biotechnol, 2013. [DOI | PubMed]
- B Lagane, G Gaibelet, E Meilhoc, JM Masson, L Cézanne, A Lopez. Role of sterols in modulating the human μ-opioid receptor function in Saccharomyces cerevisiae. J Biol Chem, 2000. [DOI | PubMed]
- A Kondo, M Ueda. Yeast cell-surface display—applications of molecular display. Appl Microbiol Biotechnol, 2004. [DOI | PubMed]
- N Sato, T Matsumoto, M Ueda, A Tanaka, H Fukuda, A Kondo. Long anchor using Flo1 protein enhances reactivity of cell surface-displayed glucoamylase to polymer substrates. Appl Microbiol Biotechnol, 2002. [DOI | PubMed]
- YS Grewal, MJA Shiddiky, SM Mahler, GA Cangelosi, M Trau. Nanoyeast and other cell envelope compositions for protein studies and biosensor applications. ACS Appl Mater Interfaces, 2016. [DOI | PubMed]
- AS Mironov, I Gusarov, R Rafikov, LE Lopez, K Shatalin, RA Kreneva. Sensing small molecules by nascent RNA: a mechanism to control transcription in bacteria. Cell, 2002. [DOI | PubMed]
- A Nahvi, N Sudarsan, MS Ebert, X Zou, KL Brown, RR Breaker. Genetic control by a metabolite binding mRNA. Chem Biol, 2002. [DOI | PubMed]
- MN Win, CD Smolke. Higher-order cellular information processing with synthetic RNA devices. Science, 2008. [DOI | PubMed]
- JC Liang, AL Chang, AB Kennedy, CD Smolke. A high-throughput, quantitative cell-based screen for efficient tailoring of RNA device activity. Nucl Acids Res, 2012. [DOI | PubMed]
- EF Ellis. Theophylline toxicity. J Allergy Clin Immunol, 1985. [DOI | PubMed]
- A Wachter, M Tunc-Ozdemir, BC Grove, PJ Green, DK Shintani, RR Breaker. Riboswitch control of gene expression in plants by splicing and alternative 3′ end processing of mRNAS. Plant Cell, 2007. [DOI | PubMed]
- MT Cheah, A Wachter, N Sudarsan, RR Breaker. Control of alternative RNA splicing and gene expression by eukaryotic riboswitches. Nature, 2007. [DOI | PubMed]
- D McRose, J Guo, A Monier, S Sudek, S Wilken, S Yan. Alternatives to vitamin B1 uptake revealed with discovery of riboswitches in multipleine eukaryotic lineages. ISME J, 2014. [DOI | PubMed]
- Y Cai, N Agmon, WJ Choi, A Ubide, G Stracquadanio, K Caravelli. Intrinsic biocontainment: multiplex genome safeguards combine transcriptional and recombinational control of essential yeast genes. Proc Natl Acad Sci USA, 2015. [DOI | PubMed]
- N Agmon, Z Tang, K Yang, B Sutter, S Ikushima, Y Cai. Low escape-rate genome safeguards with minimal molecular perturbation of Saccharomyces cerevisiae. Proc Natl Acad Sci USA, 2017. [DOI | PubMed]
- JI Yoo, S pälä, MA O’Malley. Engineered fluoride sensitivity enables biocontainment and selection of genetically-modified yeasts. Nat Commun, 2020. [DOI | PubMed]
- T Chang, W Ding, S Yan, Y Wang, H Zhang, Y Zhang. A robust yeast biocontainment system with two-layered regulation switch dependent on unnatural amino acid. Nat Commun, 2023. [DOI | PubMed]
- SA Hoffmann, Y Cai. Engineering stringent genetic biocontainment of yeast with a protein stability switch. Nat Commun, 2024. [DOI | PubMed]
- J Claesen, MA Fischbach. Synthetic microbes as drug delivery systems. ACS Synth Biol, 2015. [DOI | PubMed]
- J Fu, J Liu, X Wen, G Zhang, J Cai, Z Qiao. Unique probiotic properties and bioactive metabolites of Saccharomyces boulardii. Probiotics Antimicrob Proteins, 2023. [DOI | PubMed]
- I Khatri, R Tomar, K Ganesan, GS Prasad, S Subramanian. Complete genome sequence and comparative genomics of the probiotic yeast Saccharomyces boulardii. Sci Rep, 2017. [DOI | PubMed]
- LE Hudson, MB Fasken, CD McDermott, SM McBride, EG Kuiper, DB Guiliano. Functional heterologous protein expression by genetically engineered probiotic yeast Saccharomyces boulardii. PLoS ONE, 2014. [DOI | PubMed]
- JJ Liu, II Kong, GC Zhang, LN Jayakody, H Kim, PF Xia. Metabolic engineering of probiotic Saccharomyces boulardii. Appl Environ Microbiol, 2016. [DOI | PubMed]
- D Durmusoglu, IS Al’Abri, SP Collins, J Cheng, A Eroglu, CL Beisel. In situ biomanufacturing of small molecules in the mammalian gut by probiotic Saccharomyces boulardii. ACS Synth Biol, 2021. [DOI | PubMed]
- CL Ho, HQ Tan, KJ Chua, A Kang, KH Lim, KL Ling. Engineered commensal microbes for diet-mediated colorectal-cancer chemoprevention. Nat Biomed Eng, 2018. [DOI | PubMed]
- M Tscherner, TW Giessen, L key, CA Kumamoto, PA Silver. A synthetic system that senses Candida albicans and inhibits virulence factors. ACS Synth Biol, 2019. [DOI | PubMed]
- IY Hwang, E Koh, A Wong, JC ch, WE Bentley, YS Lee. Engineered probiotic Escherichia coli can eliminate and prevent Pseudomonas aeruginosa gut infection in animal models. Nat Commun, 2017. [DOI | PubMed]
- K Chen, Y Zhu, Y Zhang, T Hamza, H Yu, A Saint Fleur. A probiotic yeast-based immunotherapy against Clostridioides difficile infection. Sci Transl Med., 2020. [DOI | PubMed]
- C Wenisch, B Parschalk, M Hasenhundl, AM Hirschl, W Graninger. Comparison of vancomycin, teicoplanin, metronidazole, and fusidic acid for the treatment of Clostridium difficile—associated diarrhea. Clin Infect Dis, 1996. [DOI | PubMed]
- BM Scott, C Gutiérrez-Vázquez, LM Sanmarco, JA Da Silva Pereira, Z Li, A Plasencia. Self-tunable engineered yeast probiotics for the treatment of inflammatory bowel disease. Nat Med, 2021. [DOI | PubMed]
- KA Hedin, H Zhang, V Kruse, VE Rees, F Bäckhed, TU Greiner. Cold exposure and oral delivery of GLP-1R agonists by an engineered probiotic yeast strain have antiobesity effects in mice. ACS Synth Biol, 2023. [DOI | PubMed]
- R Li, X Wan, TM Takala, PEJ Saris. Heterologous expression of the leuconostoc bacteriocin leucocin C in probiotic yeast Saccharomyces boulardii. Probiotics Antimicrob Proteins, 2021. [DOI | PubMed]
- ON Rebeck, MJ Wallace, J Prusa, J Ning, EM Evbuomwan, S Rengarajan. A yeast-based oral therapeutic delivers immune checkpoint inhibitors to reduce intestinal tumor burden. Cell Chem Biol, 2024. [DOI | PubMed]
- J Kim, C Atkinson, MJ Miller, KH Kim, YS Jin. Microbiome engineering using probiotic yeast: Saccharomyces boulardii and the secreted human lysozyme lead to changes in the gut microbiome and metabolome of mice. Microbiol Spectr., 2023. [DOI | PubMed]
- D Durmusoglu, DJ Haller, IS Al’Abri, K Day, C Sands, A Clark. Programming probiotics: diet-responsive gene expression and colonization control in engineered S. boulardii. ACS Synth Biol, 2024. [DOI | PubMed]
- KA Hedin, V Kruse, R Vazquez-Uribe, MOA Sommer. Biocontainment strategies for in vivo applications of Saccharomyces boulardii. Front Bioeng Biotechnol, 2023. [DOI]
- G Bagherpour, H Ghasemi, B Zand, N Zarei, F Roohvand, EM Ardakani. Oral administration of recombinant Saccharomyces boulardii expressing ovalbumin-CPE fusion protein induces antibody response in mice. Front Microbiol, 2018. [DOI]
- Y Jin, S Yu, JJ Liu, EJ Yun, JW Lee, YS Jin. Production of neoagarooligosaccharides by probiotic yeast Saccharomyces cerevisiae var. boulardii engineered as a microbial cell factory. Microb Cell Fact, 2021. [DOI | PubMed]
- S Sun, X Xu, L Liang, X Wang, X Bai, L Zhu. Lactic acid-producing probiotic Saccharomyces cerevisiae attenuates ulcerative colitis via suppressing macrophage pyroptosis and modulating gut microbiota. Front Immunol, 2021. [DOI]
- BT De Carvalho, A Subotić, P Vandecruys, S Deleu, S Vermeire, JM Thevelein. Enhancing probiotic impact: engineering Saccharomyces boulardii for optimal acetic acid production and gastric passage tolerance. Appl Environ Microbiol, 2024. [DOI | PubMed]
- S Deleu, I Jacobs, JF Vazquez Castellanos, S Verstockt, B Trindade De Carvalho, A Subotić. Effect of mutant and engineered high-acetate-producing Saccharomyces cerevisiae var. boulardii strains in dextran sodium sulphate-induced colitis. Nutrients, 2024. [DOI | PubMed]
- CH Liu, JH Chang, YC Chang, KY Mou. Treatment of murine colitis by Saccharomyces boulardii secreting atrial natriuretic peptide. J Mol Med, 2020. [DOI | PubMed]
- Z Yang, Y Wu, X Liu, M Zhang, J Peng, H Wei. S. boulardii early intervention maintains gut microbiome structure and promotes gut mucosal barrier function in early-weaned rats. Nutrients, 2022. [DOI | PubMed]
- H Stier, S Bischoff. Influence of Saccharomyces boulardii CNCM I-745 on the gut-associated immune system. Clin Exp Gastroenterol, 2016. [DOI | PubMed]
- 291.National Institute of HealthNIH guidelines for research involving recombinant or synthetic nucleic acid molecules2024BethesdaU.S. Department of health and human services national institutes of healthNational Institute of Health. NIH guidelines for research involving recombinant or synthetic nucleic acid molecules. Bethesda: U.S. Department of health and human services national institutes of health; 2024.
- T Ishii, M Araki. Consumer acceptance of food crops developed by genome editing. Plant Cell Rep, 2016. [DOI | PubMed]
- M Cordaillat-Simmons, A Rouanet, B Pot. Live biotherapeutic products: the importance of a defined regulatory framework. Exp Mol Med, 2020. [DOI | PubMed]
- A Rouanet, S Bolca, A Bru, I Claes, H Cvejic, H Girgis. Live biotherapeutic products, a road map for safety assessment. Front Med, 2020. [DOI]
- LA Rettenbacher, K Arauzo-Aguilera, L Buscajoni, A Castillo-Corujo, B Ferrero-Bordera, A Kostopoulou. Microbial protein cell factories fight back?. Trends Biotechnol, 2022. [DOI | PubMed]
- G Walsh, E Walsh. Biopharmaceutical benchmarks 2022. Nat Biotechnol, 2022. [DOI | PubMed]
- VR Saca, C Burdette, TP Sakmar. GPCR biosensors to study conformational dynamics and signaling in drug discovery. Annu Rev Pharmacol Toxicol, 2024. [DOI | PubMed]
- K Gronbach, U Eberle, M Müller, TA Ölschläger, U Dobrindt, F Leithäuser. Safety of probiotic Escherichia coli strain Nissle 1917 depends on intestinal microbiota and adaptive immunity of the host. Infect Immun, 2010. [DOI | PubMed]