
Genome-wide inventory and functional characterization of cannabinoid oxidocyclases in Cannabis sativa
Affiliations: Haixia Institute of Science and Technology, State Key Laboratory of Agricultural and Forestry Biosecurity, Fujian Provincial Key Laboratory of Haixia Plant Systems Biology, Institute of Applied Ecology, Instrumental Analysis Center, Fujian Agriculture and Forestry University, Fuzhou 350002, China; State Key Laboratory of Plant Environmental Resilience, College of Life Science, Zhejiang University, Hangzhou, Zhejiang 310058, China; Department of Plant Physiology, Swammerdam Institute for Life Sciences, University of Amsterdam, Science Park 904, Amsterdam 1098 XH, The Netherlands; Marine and Agricultural Biotechnology Laboratory, Institute of Oceanography, Minjiang University, Fuzhou 350108, China; Shenzhen Institutes of Advanced Technology, Chinese Academy of Science, Shenzhen 518055, China; Key Laboratory of Synthetic Biology, Center for Excellence in Molecular Plant Science, Institute of Plant Physiology and Ecology, Chinese Academy of Sciences, Shanghai 200032, China; Department of Biological Sciences, Brock University, St. Catharines, ON L2S 3A1, Canada; Laboratory of Southern Subtropical Plant Diversity, Fairy Lake Botanical Garden, Shenzhen & Chinese Academy of Sciences, Shenzhen 518004, China; State Key Laboratory of Agricultural Genomics, BGI-Shenzhen, Shenzhen 518083, China; Institute of Soil Water and Environmental Science, Volcani Institute, 68 HaMaccabim Road, PO Box 15159, Rishon LeZion 7505101, Israel
License: © The Author(s) 2026. Published by Oxford University Press on behalf of the Nanjing Agricultural University. CC BY 4.0 This is an Open Access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted reuse, distribution, and reproduction in any medium, provided the original work is properly cited.
Article links: DOI: 10.1093/hr/uhag023 | PMC: PMC13143376
Relevance: Moderate: mentioned 3+ times in text
Full text: PDF (1.0 MB)
Cannabis sativa is distinguished by its production of cannabinoids that exhibit diverse therapeutic effects, including antiemetic, antispasticity, anxiolytic, and antiepileptic activities. Over 120 phytocannabinoids have been identified, with Δ9-tetrahydrocannabinol (THC) and cannabidiol (CBD) being the most abundant. THC is the principal psychoactive constituent responsible for the euphoric effects of cannabis, while CBD is nonpsychoactive and counteracts several adverse effects of THC. Understanding the biosynthesis of these compounds is critical due to their pharmacological potential.
Cannabinoid oxidocyclases (CBSs), a group of berberine bridge enzyme (BBE)-like proteins, catalyze the committed step in cannabinoid biosynthesis [ref. 1]. CBSs contain a flavin adenine dinucleotide (FAD)-binding domain and a substrate-binding BBE-like domain, which mediate the conversion of the substrate CBGA into acidic cannabinoids, including Δ9-tetrahydrocannabinolic acid (THCA), cannabidiolic acid (CBDA), and cannabichromenic acid (CBCA). These acidic cannabinoids are subsequently converted into their neutral forms (THC, CBD, and CBC) through non-enzymatic decarboxylation when exposed to heat [ref. 2]. The major cannabinoid synthases (THCAS, CBDAS, CBCAS) have been extensively characterized. Heterologous expression of THCAS in tobacco leaves produced THCA and CBCA. Expression of THCAS and CBDAS in Pichia pastoris yielded mixtures of THCA, CBDA, and CBCA and additional unknown compounds, while CBCAS expressed in P. pastoris catalyzes the specific formation of CBCA [ref. 3]. Protein crystallization and site-directed mutagenesis of THCAS revealed critical residues near substrate-binding pocket and FAD-binding site [ref. 1]. Nevertheless, additional CBSs contributing to cannabinoid diversity remain largely unexplored.
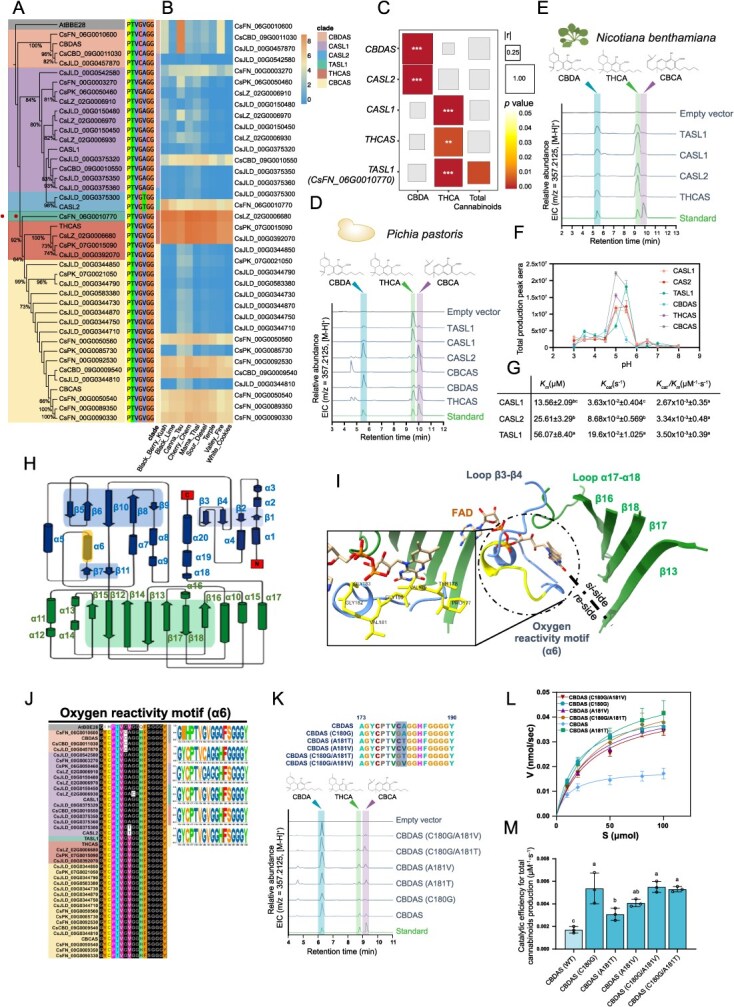
To address this, we performed a genome-wide survey of five cannabis genomes and identified 36 putatively functional CsCBS genes. These included 8 from CSFN, 4 from CSPK, 3 from CSCBD, and 17 from CSJLD, plus five previously reported genes (Fig. 1A). Phylogenetic analysis grouped these sequences into five clades, largely corresponding to functionally characterized CBSs, except one clade harboring CBDAS-like genes (CBDAS-like 1, CASL1, and CBDAS-like 2 CASL2) of unknown function [ref. 4]. A distinct gene, CsFN_06G0010770, that diverged from other CBSs, was named THCAS-like 1 (TASL1). Transcriptome profiling of mature glandular trichomes across nine varieties revealed high expression of CBDAS, THCAS, and CBCAS, as well as detectable expression of CASL1, CASL2, and TASL1 (Fig. 1B) [ref. 2]. Expression correlation analyses across 21 female flower varieties showed that CBDAS and THCAS expression levels correlated with CBDA and THCA accumulation, respectively. Interestingly, TASL1 and CASL1 expression correlated with THCA production, whereas CASL2 correlated with CBDA (Fig. 1C and Fig. S1), suggesting these enzymes might function differently in planta. Heterologous expression of CASL1, CASL2, and TASL1 in P. pastoris and Nicotiana benthamiana confirmed their enzymatic activity, while revealing distinct product preferences (Fig. 1D and E). For the N. benthamiana assays, Agrobacterium tumefaciens GV3101 harboring pEAQ-CASL1/CASL2/TASL1 was cultured and used for agroinfiltration (10 mM MES, 10 mM MgCl2, 100 μM acetosyringone to OD600 = 0.6), the CBGA substrate was infiltrated into the same leaf areas at 4 days post-agroinfiltration, and leaf tissues were harvested for metabolic analysis 1 day later. CASL1 and TASL1 converted CBGA to THCA as the major product in both heterologous systems, whereas CASL2 exhibited host-dependent activity, producing CBDA and CBCA as the predominant products in P. pastoris, but primarily THCA in N. benthamiana. All three enzymes displayed a pH optimum ~5.5, consistent with apoplastic conditions reported for CBS activity (Fig. 1F) [ref. 5]. Kinetic analyses indicated that TASL1 and CASL2 exhibited comparable catalytic efficiencies (kcat/Km), both of which were modestly higher than that of CASL1 (Fig. 1G). The Km value of TASL1 (56.07 on average) was much higher than CASL1 and CASL2 (13.56 and 25.61 on average, respectively), indicating a lower affinity between TASL1 and CBGA, while the kcat value of TASL1 was about 5.4 and 2.2 times higher than that of CASL1 and CASL2, respectively, indicating a greater turnover efficiency of TASL1. Collectively, these results indicate that CASL1, CASL2, and TASL1 likely contribute differently to cannabinoid biosynthesis and overall cannabinoid yield.
To explore the basis of functional diversity among CsCBSs, we next examined structural motifs that may influence product profiles and catalytic efficiency. Overall, CsCBSs display a conserved topology comprising distinct FAD-binding and substrate-binding domains (Fig. 1H). Four β-sheets (β13, β16, β17, and β18) and the loop connecting α17 and α18 form a wide-open substrate-binding pocket located on the si-side of the FAD isoalloxazine ring (Fig. 1I). The loop connecting β3 to β4 and an α-helix (α6) located on the re-side of the FAD isoalloxazine ring. A motif containing α6 and its flanking amino acids was previously identified as an oxygen reactivity motif (ORM), which may function as an oxygen cavity that traps dioxygen and catalyze the oxidation of the flavin [ref. 5].
We next compared sequence homology across seven conserved motifs forming the active site of BBE-like enzymes in Arabidopsis (Fig. S2). Two residues within the ORM were conserved within specific CsCBS phylogenetic clades (Fig. 1J), and their variation paralleled the overall phylogenetic relationships among family members (Fig. 1A). In THCAS, FAD-mediated redox catalysis involves hydride transfer from CBGA to the oxidized FAD cofactor [ref. 1], while the ORM facilitates reoxidation of reduced FAD for subsequent catalytic cycles [ref. 5]. To assess the contribution of individual ORM residues to CBS activity, we introduced targeted substitutions in CBDAS: C180G and A181T or A181V. All variants retained CBDA as the main product (Fig. 1K) but exhibited increased catalytic efficiency (kcat/Km) relative to the wild type (Fig. 1L and M). Notably, the C180G substitution, which likely enlarges the oxygen-binding cavity, led to a 3-fold increase in catalytic efficiency, while A181V conferred greater activity than A181T (Fig. 1M), indicating that amino acid substitutions within the ORM can significantly enhance catalytic efficiency in cannabinoid biosynthesis. Molecular docking of CBDAS and its variants with CBGA as the ligand revealed that the variants display higher binding affinities for CBGA than the wild-type enzyme. Within the active pocket of the CBDAS variants, the CBGA substrate is positioned in closer proximity to the FAD cofactor and the essential active site residues TYR483 (Fig. S3), providing a structural basis for the enhanced catalytic performance.
In summary, our genome-wide analysis identified 36 putatively functional CsCBS genes, including the evolutionarily distinct TASL1. Transcriptomic and biochemical evidence indicates TASL1, CASL1, and CASL2 contribute to cannabinoid diversity, with TASL1 catalyzing CBGA conversion to THCA, CBDA, and CBCA. Structural and mutational analyses reveal that ORM residues influence catalytic efficiency, reflecting evolutionary adaptation toward optimized cannabinoid production. These findings advance our understanding of the genetic and enzymatic framework underlying cannabinoid formation and provide a basis for enzyme engineering, metabolic pathway optimization in plants, and applications in synthetic biology.
References
- Structure and function of Δ1-tetrahydrocannabinolic acid (THCA) synthase, the enzyme controlling the psychoactivity of Cannabis sativa.. J Mol Biol., 2012. [PubMed]
- Gene networks underlying cannabinoid and terpenoid accumulation in cannabis.. Plant Physiol., 2019. [PubMed]
- Origin and evolution of the cannabinoid oxidocyclase gene family.. Genome Biol Evol., 2021. [PubMed]
- Cannabidiolic-acid synthase, the chemotype-determining enzyme in the fiber-type Cannabis sativa.. FEBS Lett., 2007. [PubMed]
- Rationally engineered flavin-dependent oxidase reveals steric control of dioxygen reduction.. FEBS J., 2015. [PubMed]