
Exploring cannabinoid modulation on autophagy mechanisms in Alzheimer’s disease: a review
Abstract
Alzheimer’s disease (AD) is a neurodegenerative disorder characterized by the accumulation of toxic protein aggregates in the brain, leading to brain cell death and cognitive impairment. Central to AD pathogenesis is the autophagy pathway, a crucial cellular self-digestion process. Cannabinoids, the fundamental phytochemical compounds derived from the Cannabis sativa plant, have been demonstrated to exhibit neuroprotective qualities when used as a treatment at microdoses. However, the impact of multi-cannabinoid treatments on autophagy induction and subsequent cell survival in AD in vitro models remains uncertain. This review seeks to explore the potential of a multi-cannabinoid treatment strategy in enhancing neuronal cell survival through autophagy activation within an AD in vitro model. The proposed approach involves a combination of cannabinoids in their potential to upregulate autophagy mechanisms, potentially supporting neuronal cell resilience. By unravelling the mechanistic link between autophagy, cannabinoid treatment, and neuronal viability, this review aims to elucidate how cannabinoids influence neuronal function and survival at a cellular and molecular level. By offering insights into the exploitation of the endocannabinoid system, this review contributes to the development of novel cannabinoid-based treatment avenues for AD. This pursuit aligns with the broader objective of addressing the debilitating effects of AD on the quality of life for those affected.
Article type: Review Article
Keywords: Alzheimer’s disease, amyloid precursor protein, amyloid-β (Aβ), autophagy, cannabinoid-based therapeutics, Cannabis sativa
Affiliations: Department of Basic Medical Sciences, School of Biomedical Sciences, Faculty of Health Sciences, University of the Free State, Bloemfontein, South Africa; Department of Pharmacology, School of Clinical Medicine, Faculty of Health Sciences, University of the Free State, Bloemfontein, South Africa; University of the Free State – Technology Innovation Agency Pharmacology Platform (AMITD), Bloemfontein, South Africa
License: Copyright © 2026 Ntsapi, Weyers, Chinheya, Jim and Matsabisa. CC BY 4.0 This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
Article links: DOI: 10.3389/fphar.2025.1748368 | PubMed: 41601969 | PMC: PMC12832750
Relevance: Moderate: mentioned 3+ times in text
Full text: PDF (3.6 MB)
Introduction
Neurodegeneration involves the progressive decline of neurons and is a hallmark of disorders such as Alzheimer’s disease (AD), Parkinson’s disease (PD), and Amyotrophic Lateral Sclerosis (ref. Mistretta et al., 2023; ref. Agnello and Ciaccio, 2022). AD, the most prevalent neurodegenerative disorder (ref. Świetlik et al., 2022), is characterized by the accumulation of amyloid-β (Aβ) plaques and hyperphosphorylated tau, leading to neuronal dysfunction, inflammation, and oxidative stress in the central nervous system (ref. Halawani et al., 2010; ref. Rohn and Head, 2009). AD is broadly classified based on its pathophysiology as either sporadic (late-onset) or familial (early-onset). Sporadic AD, accounting for approximately 95% of cases, is a complex disorder influenced by multiple factors that remain incompletely understood. These include genetic factors, such as the Apolipoprotein E allele, as well as environmental and lifestyle influences, and the interplay of epigenetic and genetic processes (ref. Tailor et al., 2019; ref. Ibanez et al., 2021). Familial AD, which affects fewer than 5% of patients, results from mutations in specific genes that alter the amyloid precursor protein (APP) or presenilin-1 and -2 (PS1/PS2) (ref. Xiao et al., 2021; ref. Lanoiselée et al., 2017). Both sporadic and familial forms of AD share these pathological features, though the underlying causes differ (ref. Tailor et al., 2019; ref. Ibanez et al., 2021; ref. Lanoiselée et al., 2017; ref. d‘Errico and Meyer-Luehmann, 2020). The amyloid cascade hypothesis suggests that impaired clearance of Aβ triggers downstream tau pathology and neuronal toxicity, as demonstrated in cellular and animal models (ref. Hardy and Higgins, 1992; ref. Alawode et al., 2022). Despite strong preclinical evidence, translating these findings into effective clinical therapies has been challenging, often due to safety concerns and limitations of conventional drug approaches.
Despite the strong preclinical evidence linking Aβ accumulation and tau pathology to AD progression, translating this robust preclinical and biomarker science into clinical benefit has proven challenging. Logistical difficulties, safety concerns, and limited tolerable dosages have hindered the success of secretase-inhibiting therapies, and several clinical trials have been prematurely halted due to adverse effects or other pharmaceutical limitations (ref. Haass and Selkoe, 2022; ref. Ricciarelli and Fedele, 2017). These challenges underscore the need for alternative therapeutic strategies, such as cannabinoids, which have shown potential in modulating neuroinflammation, oxidative stress, and Aβ- and tau-mediated neuronal toxicity, positioning them as promising multi-target agents in AD management (ref. d‘Errico and Meyer-Luehmann, 2020; ref. DeTure and Dickson, 2019; ref. Halawani et al., 2010; ref. Rohn and Head, 2009; ref. Yankner et al., 1989).
APP processing and Aβ peptide formation
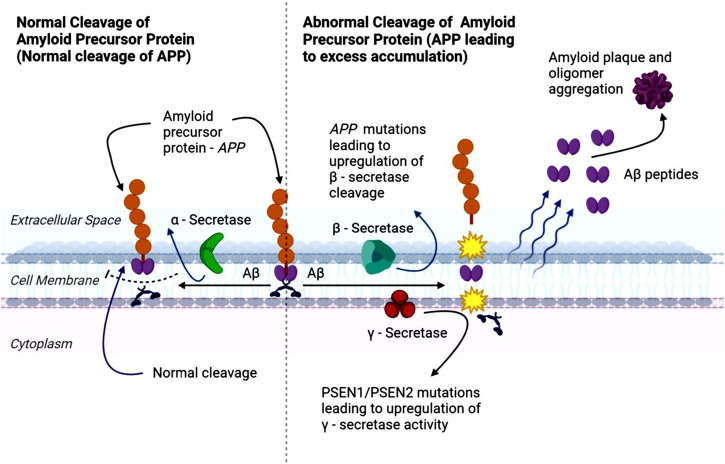
APP is a widely spread transmembrane protein (ref. Morley et al., 2019) involved in multiple physiological processes, including wound healing, calcium regulation in neurons, transport of molecules across cell membranes, and the formation and maintenance of synapses, which are critical for neuronal communication (ref. Morley et al., 2019; ref. O’Brien and Wong, 2011; ref. Bergström et al., 2016). Importantly, abnormal accumulation of APP’s proteolytic product, the Aβ peptide, is a defining feature of AD (ref. O’Brien and Wong, 2011; ref. Bergström et al., 2016). Aβ peptides are produced through the sequential cleavage of APP by β-secretase (BACE) and γ-secretase enzymes. This processing produces N-terminal fragments, such as soluble amyloid precursor protein-alpha (sAPPα) and soluble amyloid precursor protein-beta (sAPPβ), Aβ peptides, and C-terminal fragments (ref. O’Brien and Wong, 2011).
Aβ peptides are hydrophobic and prone to aggregate, forming oligomers and protofibrils that eventually deposit as Aβ plaque in the brain (ref. Morley et al., 2019). APP can undergo either non-amyloidogenic or amyloidogenic processing, with the latter favored in AD (ref. Fong et al., 2018; ref. Zhao et al., 2020). In the amyloidogenic pathway (Figure 1), β-secretase mediates the initial cleavage of APP, producing sAPPβ and a carboxy-terminal fragment β (CTFβ or C99) (ref. Roberds et al., 2001; ref. Hampel et al., 2021). C99 fragments are subsequently cleaved by γ-secretase, generating Aβ and carboxy-terminal fragment γ (CTFγ) or APP intracellular domain (AICD). Excessive APP cleavage drives the accumulation of Aβ fragments, which aggregate into oligomers and fibrils, culminating in plaque formation, and contributing to the synaptic dysfunction, inflammation, and neurotoxicity characteristic of AD (ref. Roberds et al., 2001; ref. Hampel et al., 2021; ref. Takahashi et al., 2017).
Approved drugs for AD treatment and their mechanism of action
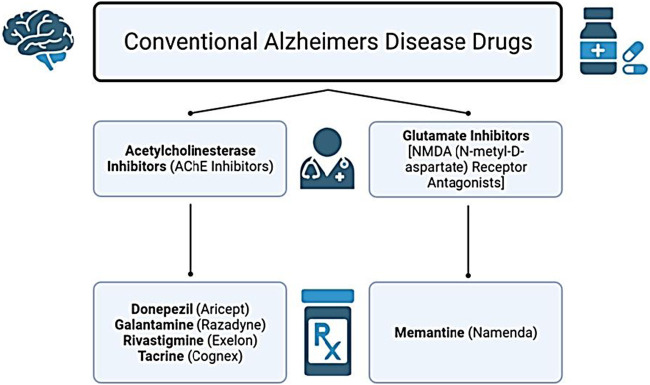
The growing global prevalence of AD underscores the urgent need for effective therapeutic strategies. Although scientific and medical advances have improved understanding of AD, no treatment currently exists that can halt or reverse disease progression. The disorder remains non-preventable, incurable, and challenging to diagnose with high accuracy (ref. d‘Errico and Meyer-Luehmann, 2020; ref. Hardy and Higgins, 1992). To date, two major classes of medications are approved for AD: acetylcholinesterase inhibitors and N-methyl-D-aspartate (NMDA) receptor antagonists (ref. Alawode et al., 2022; ref. Haass and Selkoe, 2022). In the United States, the Food and Drug Administration (FDA) has approved drugs that either modestly slow clinical decline or offer temporary symptomatic relief. These include the acetylcholinesterase inhibitors – Rivastigmine, Donepezil, Galantamine, and Tacrine – alongside Memantine (MEM), the primary NMDA receptor antagonist (glutamate inhibitor) (ref. Reisberg et al., 2003). These conventional drug classes are summarized in Figure 2.
Acetylcholinesterase in AD treatment
Neurodegeneration in AD is driven in part by the accumulation of Aβ aggregates and hyperphosphorylated tau, leading to progressive neuronal loss and deterioration of cognitive, language, and memory functions (ref. Sharma, 2019). Acetylcholine (ACh), a key neurotransmitter within cholinergic neurons of the central and autonomic nervous systems, plays an essential role in learning, memory, and synaptic plasticity (ref. Radu et al., 2017; ref. Picciotto et al., 2012). These systems include the well-known neuromuscular junctions, each exhibiting distinct nicotinic and muscarinic receptors (ref. Picciotto et al., 2012). In the brain, all five subtypes of muscarinic receptors (M1–M5) are present, with the M1 receptor subtype being particularly important for higher-order cognitive processing (ref. Tizabi et al., 2023; ref. Wright et al., 2009).
Under normal conditions, ACh is rapidly degraded in the synaptic cleft by acetylcholinesterase (AChE), which terminates cholinergic signaling. Reductions in ACh levels can arise from decreased synthesis or release, increased AChE activity or expression, or altered cholinergic receptor function (ref. Kuo et al., 2007). In AD, dysfunction of the cholinergic system is well-documented: both the concentration and functional activity of ACh are significantly reduced, impairing cognitive processes (ref. GBD, 2019, 2022). AChE, predominantly located at neuromuscular and neuronal synapses, hydrolyses into acetic acid and choline (ref. Trang and Khandhar, 2023). Its upregulation accelerates ACh breakdown, contributing to synaptic failure and cognitive impairment (ref. Iliyasu et al., 2023).
Notably, AChE also enhances Aβ aggregation by binding to Aβ and inducing fibril formation, generating a neurotoxic Aβ-AChE complex that disrupts normal synaptic function, particularly in the hippocampus (ref. Picciotto et al., 2012; ref. Dinama et al., 2010). Elevated AChE around amyloid plaques and neurofibrillary tangles is a characteristic feature of AD, although the mechanistic significance remains unclear (ref. García-Ayllón et al., 2011).
AChE inhibitors remain the primary pharmacological strategy for symptom management (ref. McGleenon et al., 1999). These inhibitors provide two main therapeutic benefits: (i) enhancement of synaptic ACh levels, improving cognitive function through strengthened cholinergic signaling; and (ii) potential long-term neuroprotective effects, including slower cortical atrophy and delayed clinical progression of dementia (ref. Moss, 2020).
Given the role of ACh across sensory, cognitive, and sleep-wake systems (ref. H Ferreira-Vieira et al., 2016), disruptions in cholinergic neurotransmission amplify AD-related deficits. More specifically, Aβ and tau pathology impairs synaptic integrity within the cholinergic pathways, reducing ACh availability and further compromising neuronal communication (ref. H Ferreira-Vieira et al., 2016; ref. Machado et al., 2020). Early cholinergic lesions occur presynaptically and later extend to post-synaptic nicotinic and muscarinic receptors, particularly M1, reinforcing the rationale for targeting the cholinergic system (ref. Klaassens et al., 2019). Moreover, impaired cholinergic function may intensify downstream AD pathology, including tau phosphorylation, neuroinflammation, and neuronal injury (ref. Chen et al., 2022), further accelerating cognitive decline in AD.
Memantine in AD treatment
The glutamatergic hypothesis of AD proposes that dysregulated glutamate signaling contributes to synaptic dysfunction and neurodegeneration. Glutamate, the major excitatory neurotransmitter in the brain, activates NMDA receptors essential for learning, memory; however, chronic overstimulation leads to excitotoxicity and neuronal injury (ref. Świetlik et al., 2022). MEM, an antagonist of extra-synaptic NMDA receptors, is an established therapeutic option and frequently administered alongside AChE inhibitors. Compared with other non-competitive NMDA receptor antagonists, MEM demonstrates superior tolerability and a more favorable pharmacological profile (ref. Lipton and Chen, 2005).
MEM preferentially binds to open NMDA receptor-operated calcium channels, attenuating pathological calcium influx while preserving physiological synaptic transmission (ref. Yiannopou et al., 2020). Through this mechanism, MEM reduces the harmful effects of sustained glutamate elevation and reduces excitotoxic neuronal loss (ref. Folch et al., 2018). Despite these mechanisms, MEM only provides modest symptomatic benefit, may cause side effects, and does not alter disease progression (ref. Reisberg et al., 2003; ref. Alhazmi and Albratty, 2022). This therapeutic limitation highlights the need for innovative interventions capable of targeting underlying pathophysiological processes rather than solely managing symptoms.
Failure of Aβ-based therapeutics
Aβ peptides accumulate in the brain long before clinical symptoms emerge (ref. Hector and Brouillette, 2021; ref. Porsteinsson et al., 2021). Their aggregation disrupts synaptic function, alters membrane dynamics, and induces structural neuronal damage, including dystrophy, synaptic loss, and impaired neurotransmission (ref. Takahashi et al., 2017; ref. Hashimoto et al., 2003; ref. Zempel et al., 2010). Aβ accumulation also contributes to glutamatergic dysfunction and excitotoxicity (ref. Catania et al., 2019; ref. Gong et al., 2018; ref. de Rojas et al., 2021), which further compromises cholinergic signaling and reduces ACh levels (ref. Pars et al., 2013; ref. Wang and Reddy, 2017). Current FDA-approved therapies, including acetylcholinesterase inhibitors and NMDA receptor antagonists, address these downstream neurotransmitter disturbances but only offer limited symptomatic relief and do not meaningfully modify disease trajectory (ref. Pars et al., 2013; ref. Wang and Reddy, 2017; ref. Ruangritchankul et al., 2021).
Although Aβ has long been a key therapeutic target, many Aβ-directed strategies have failed in clinical trials (ref. Cummings J. L. et al., 2022; ref. Cummings J. et al., 2022; ref. Schott et al., 2019; ref. Anderson et al., 2017). These failures may reflect the complexity and multifactorial nature of AD. Interventions are often initiated after extensive Aβ pathology has already developed, reducing the likelihood of therapeutic success (ref. Anderson et al., 2017). Additionally, targeting Aβ alone is insufficient, as AD progression is shaped by interconnected processes such as neuroinflammation, oxidative stress, autophagy dysfunction, synaptic loss, and dysregulated APP processing (ref. Catania et al., 2019; ref. Gong et al., 2018; ref. de Rojas et al., 2021). Collectively, these factors emphasize the need for multi-targeted or combination therapeutic strategies capable of addressing AD’s broad pathological landscape.
Given the substantial time and financial investment required for new drug development, drug repurposing has emerged as a practical approach to accelerating therapeutic discovery. Repurposed drugs bypass early safety and pharmacokinetic stages and offer a more efficient and lower-risk path toward identifying agents with disease-modifying potential. Ultimately, the clinical imperative remains the development of therapies that can meaningfully slow or reverse AD progression, while improving patient quality of life.
Autophagy dysregulation in AD pathogenesis
Autophagy is a key cellular pathway responsible for removing damaged proteins and organelles through lysosomal processing (ref. Galluzzi et al., 2017). This process is essential for neuronal function and survival, as neurons are post-mitotic and cannot dilute toxic aggregates through cell division. Moreover, the adult brain has limited regenerative capacity, in part, due to inhibitory glial signaling and a restrictive extracellular environment, making efficient autophagy flux critical for maintaining neuronal homeostasis (ref. Evans and Holzbaur, 2020; ref. Suzuki et al., 2017; ref. Fawcett, 2020).
Autophagic is a dynamic, lysosome-driven process that supports not only intracellular quality control but also key neuronal activities, including axonal maintenance, synaptic function, neuronal connectivity, and stem cell development (ref. Stavoe and Holzbaur, 2019; ref. Valencia et al., 2021; ref. B et al., 2014). Impaired autophagy flux is strongly implicated in neurodegenerative diseases, particularly AD, where defective clearance of protein aggregates contributes to synaptic dysfunction, neuronal injury, and cell death (ref. Zhang et al., 2022; ref. Park et al., 2020). Understanding how autophagy becomes dysregulated in AD is therefore central to identifying new therapeutic targets.
Three primary autophagy pathways operate in mammalian cells: microautophagy, chaperone-mediated autophagy (CMA), and macroautophagy (ref. Galluzzi et al., 2017). Microautophagy involves direct lysosomal membrane invagination, while CMA selectively delivers misfolded proteins to lysosomes via chaperone recognition (ref. Tekirdag and Cuervo, 2018). Macroautophagy, the best-characterized pathway, sequesters cytoplasmic components within double-membrane autophagosomes that subsequently fuse with lysosomes for degradation (ref. Gomes et al., 2017). This pathway is particularly important for clearing large structures, such as protein aggregates and dysfunctional mitochondria (ref. Evans and Holzbaur, 2020; ref. Bloemberg and Quadrilatero, 2019).
Although all three pathways contribute to proteostasis, dysfunction of macroautophagy has been most strongly associated with AD pathology (ref. Ntsapi et al., 2018; ref. Klionsky et al., 2021). As a result, therapeutic strategies aimed at restoring autophagic flux have gained substantial interest as potential interventions to mitigate neurodegeneration and promote neuronal resilience in AD.
Mammalian target of rapamycin complex 1 (mTORC1) – dependent autophagy pathway
Autophagy is tightly coordinated by a conserved network of autophagy-related genes (ATGs) (ref. Levine and Kroemer, 2019). A central upstream regulator of this process is the mammalian target of rapamycin (mTOR), a serine-threonine protein kinase that forms two major complexes: mTOR complex 1 (mTORC1) and mTOR complex 2 (ref. Saxton and Sabatini, 2017). Among these, mTORC1 is the primary metabolic sensor, integrating inputs related to nutrient availability, growth factors, energy status, chronic stress, oxidative stress, and glucocorticoids (ref. Li and hua, 2019; ref. Rabanal-Ruiz et al., 2017).
mTORC1 negatively regulates autophagy; when activated, it inhibits the initiation of autophagosome formation. Hyperactivation of mTORC1, a hallmark of aging, suppresses autophagy flux, resulting in the progressive accumulation of damaged proteins and organelles (ref. Nixon and Yang, 2012). Age-related increases in mTORC1 signalling, combined with general cellular decline, diminish autophagy efficiency (ref. Zinecker and Simon, 2022; ref. Zhang and Cuervo, 2008; ref. Ntsapi and Loos, 2016). As clearance capacity falters, toxic aggregates and dysfunctional organelles accumulate, driving cellular stress and increasing susceptibility to age-associated pathologies, including neurodegenerative diseases.
An overview of the autophagy process
Upstream regulators such as phosphoinositide 3-kinase (PI3K) and protein kinase B (Akt/PKB) primarily through the PI3K/Akt/mTOR pathway (ref. Rabanal-Ruiz et al., 2017). Inhibition of PI3K, Akt, or mTOR promotes autophagy and enhances Aβ clearance in AD models (ref. Nilsson and Saido, 2014; ref. Nilsson et al., 2013; ref. Nilsson et al., 2014; ref. Nixon, 2017; ref. Nixon, 2007). Reactive oxygen species (ROS) also stimulate autophagy by suppressing Akt/mTORC1 signalling (ref. Kaushal et al., 2019; ref. Kma and Baruah, 2022).
Under nutrient-rich conditions, active mTORC1 inhibits autophagy initiation by phosphorylating the Unc-51-like autophagy activating kinase 1 (ULK1) complex (ref. Rabanal-Ruiz et al., 2017). Upon mTORC1 inactivation, the ULK1 complex triggers formation of the phagophore, the precursor membrane that expands to form the autophagosome. The phagophore elongates through two ubiquitin-like conjugating systems: the ATG12-ATG5-ATG16L complex and microtubule-associated protein light chain 3 (LC3I, MAP1LC3B) (ref. Galluzzi et al., 2017; ref. Yang and Klionsky, 2010). LC3 is converted from its cytosolic (LC3-I) to its lipidated, membrane-bound form (LC3-II), which stabilizes the autophagosomal membrane and facilitates fusion with lysosomes (ref. Wang et al., 2013) (Figure 3). Autophagosomes sequester cytoplasmic cargo, including APP, Aβ, and tau-containing aggregates, relevant to AD pathology (ref. Lucà et al., 2021; ref. Yu and Goncharova, 2022; ref. Harrison et al., 2009).
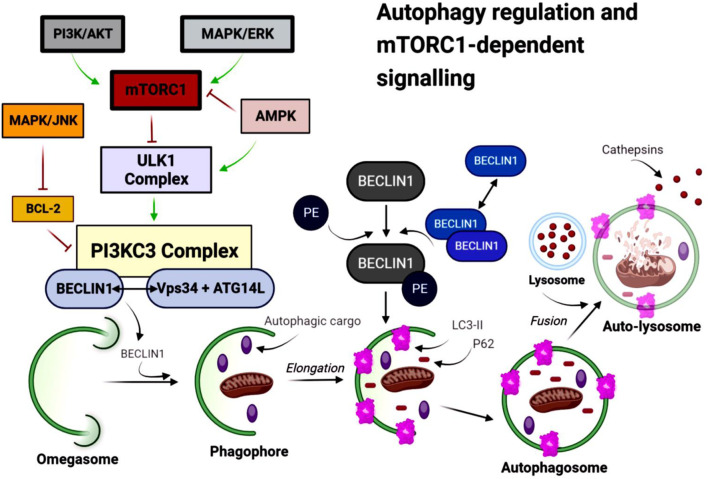
Mature autophagosomes fuse with lysosomes to form an autophagolysosome (autolysosome), where cargo is degraded by lysosomal hydrolases and the resulting metabolites are recycled (ref. Suzuki et al., 2017; ref. Yang and Klionsky, 2010). LC3-II serves as a marker of autophagosome formation, whereas decreased levels of sequestosome 1 (p62/SQSTM1) indicate efficient autophagic flux (ref. B et al., 2014).
Beyond mTORC1, AMP-activated protein kinase (AMPK) is a key metabolic regulator that promotes autophagy under low-energy conditions (ref. Li and Chen, 2019; ref. Wang S. et al., 2022; ref. Tamargo-Gómez and Mariño, 2018). AMPK both inhibits mTORC1 and directly activates ULK1 (ref. Wang S. et al., 2022; ref. Garcia and Shaw, 2017). It also regulates the class III phosphatidylinositol 3-phosphate (class III PI3P) complex – comprising vacuolar protein sorting 34 (Vps34), Beclin-1 (BECN1), and Atg14L – which drives autophagosome nucleation (ref. Suzuki et al., 2017; ref. Menon and Dhamija, 2018; ref. McKnight and Zhenyu, 2013).
mTORC2 also contributes to autophagy regulation by phosphorylating Akt (protein kinase B, PKB), which in turn enhances mTORC1 signaling (ref. Gao et al., 2014). Full activation of mTORC1additionally requires the small GTPase Rheb, which is controlled by the tuberous sclerosis complex subunit 2 (TSC1/2) (ref. Lemmon and Schlessinger, 2010; ref. Khan et al., 2013). Akt suppresses TSC1/2, thereby releasing Rheb and promoting mTORC1 activation (ref. Ntsapi et al., 2018; ref. Zou et al., 2020; ref. Lucà et al., 2021). Dysregulation of both mTORC1 and mTORC2 has been implicated in AD (ref. Yu and Goncharova, 2022; ref. Boutouja et al., 2019), with reduced levels of each complex observed in human AD brain tissue (ref. Lee et al., 2017). Pharmacological modulation of mTORC1 restores autophagic balance and demonstrates therapeutic potential in transgenic AD models (ref. Lee et al., 2017; ref. Harrison et al., 2009; ref. Carro et al., 2006).
Impaired autophagy and protein aggregation in AD
In AD, the accumulation of Aβ compromises autophagy at later stages of the pathway, contributing to neuronal dysfunction and accelerated neuronal death in vitro and in vivo, as reported by Ntsapi and Loos (ref. Ntsapi and Loos, 2021) and Lee et al. (ref. Lee et al., 2022). Aβ accumulation is the central hallmark of AD and exerts neurotoxic effects. Dysregulated or suboptimal autophagy activity is therefore considered a key contributor to the onset and progression of AD (ref. Cai et al., 2015). Whether autophagy is protective or harmful depends on the context, including the type of stress, the duration and intensity of autophagy induction, the efficiency of lysosomal clearance, and the overall “autophagic flux” (ref. Jaeger and Wyss-Coray, 2009). Deleterious overactivation or prolonged induction may lead to autophagic cell death, occurring when autophagy persists excessively, causing the degradation of cytoplasm, organelles, or essential components, leading to energy failure and neuronal death. In contrast, neuroprotective autophagy typically occurs under normal or moderately stressed conditions, acting as a quality control mechanism that removes toxic proteins, maintains metabolic and organelle homeostasis, reduces neurotoxicity, and supports neuronal health (ref. Cui and Zhao, 2025; ref. Son et al., 2012).
Increased mTORC1 activity, associated with aging, further suppresses autophagy and promotes the buildup of damaged proteins and organelles (ref. Nixon and Yang, 2012; ref. Nixon and Yang, 2011). Age-related decline in autophagic efficiency is thus thought to facilitate the pathological processes underlying AD (ref. Aman et al., 2021).
Autophagy is the primary intracellular mechanism involved in modulating APP processing and Aβ production, with substantial amounts of APP and its cleavage products found within autophagic vacuoles (AVs) (ref. Nixon et al., 2005). AVs refer broadly to vesicular structures engaged in the autophagy pathway, including the pre-autophagosomal structures, autophagosomes, lysosomes, or autophagolysosomes (ref. Nixon, 2017; ref. Boland et al., 2008). These vesicles contain immunoreactive Aβ and APP-related cleavage products in vivo AD models (ref. Nixon, 2007), highlighting their central role in Aβ generation and clearance.
Inhibition of mTOR has been shown to induce autophagy and reduce BACE expression in the APP/PS1 transgenic mouse model of AD (ref. Nilsson et al., 2014; ref. Nilsson et al., 2014; ref. Rahman et al., 2020). Conversely, the autophagy inhibitor, 3-methyladenine (3-MA), enhances γ-secretase activity and increases Aβ production (ref. Park et al., 2020), highlighting the importance of maintaining normal autophagy activity for APP clearance. Loss of key ATGs leads to neurodegeneration driven by the accumulation of ubiquitinated aggregates (ref. Karsli-Uzunbas et al., 2014; ref. Kim et al., 2015; ref. Jogalekar et al., 2021; ref. Towers and Thorburn, 2016), whereas enhanced autophagy flux reverses protein buildup and supports neuronal and organismal health (ref. Suresh et al., 2020).
Excessive activation of BACE and γ-secretase accelerates AD progression by increasing APP processing and Aβ formation (ref. Sinha et al., 1999; ref. Vassar et al., 1999), further underscoring the regulatory role of autophagy in APP metabolism (ref. Rahman et al., 2020; ref. Kuang et al., 2020). Pharmacological induction of autophagy in animal models of AD promotes early degradation of APP and reduces (ref. Tian et al., 2011). Ntsapi and Loos (ref. Ntsapi and Loos, 2021) reported that the persistent Aβ accumulation compromises autophagy at later stages, leading to neuronal dysfunction and accelerated cell death in vitro. Indeed, dysregulated or insufficient autophagy is considered a major contributor to AD development and progression (ref. Ntsapi et al., 2018; ref. Nixon et al., 2005; ref. Suresh et al., 2020).
Autophagy is also central to the intracellular processing of APP and Aβ, as AVs contain high levels of APP and its cleavage products (ref. Nilsson et al., 2014; ref. Nixon, 2007; ref. Bordi et al., 2016). AVs in AD models contain immunoreactive Aβ and APP precursors (ref. Ntsapi and Loos, 2016; ref. Lemmon and Schlessinger, 2010), and the relatively stable levels of APP within AVs indicate that the cleavage of APP can also occur inside these vesicles (ref. Nixon, 2007). This suggests that AVs may serve as sites for abnormal APP processing (ref. Yu et al., 2004). Consistent with these findings, inhibition of mTOR induces autophagy and reduces BACE expression in the APP/PS1 transgenic mouse model of AD (ref. Nilsson et al., 2013; ref. Nilsson et al., 2014; ref. Nixon et al., 2005). In contrast, the autophagy inhibitor 3-MA enhances γ-secretase activity and increases Aβ production (ref. Cai et al., 2015; ref. Yu et al., 2004). Taken together, maintaining functional autophagy is essential for efficient and sustained clearance of APP and Aβ-related substrates in neuronal cells.
Modulation of autophagy in AD
There is substantial evidence supporting the therapeutic potential of targeting autophagy in the treatment of AD (ref. Zhao et al., 2020; ref. Aman et al., 2021; ref. Li et al., 2022). In AD, impaired autophagy and protein aggregation reinforce one another in a vicious cycle that accelerates neurodegeneration, highlighting the need to identify safe and effective autophagy modulators. Recent research has explored cannabinoid-based therapeutics, which have shown promise in inducing autophagy and promoting cell death in vivo and in vitro in the context of cancer (ref. Lastres-Becker et al., 2005; ref. García et al., 2011). For example, Salazar and colleagues demonstrated that activation of the autophagy pathway is required for the antitumor effects of cannabinoids (ref. Salazar et al., 2009). Additional work in MDA-MB-231 breast cancer cells indicated that cannabinoid-induced cell death and autophagy can occur independently of cannabinoid receptor activation. More recently, cannabidiol was shown to induce autophagy in an in vitro model of PD (ref. Kang et al., 2021).
Collectively, these studies suggest that cannabinoid-based treatment approaches may hold therapeutic potential for a range of human diseases, including AD. However, further research is needed to identify cannabinoid-derived modulators that are both safe and effective, and to clarify the mechanisms through which cannabinoid-based autophagy may contribute to AD treatment. Continued advances in understanding pharmacokinetics and neuronal effects of cannabinoid-based therapeutics have made this an increasingly prominent area of biomedical research.
The endocannabinoid system
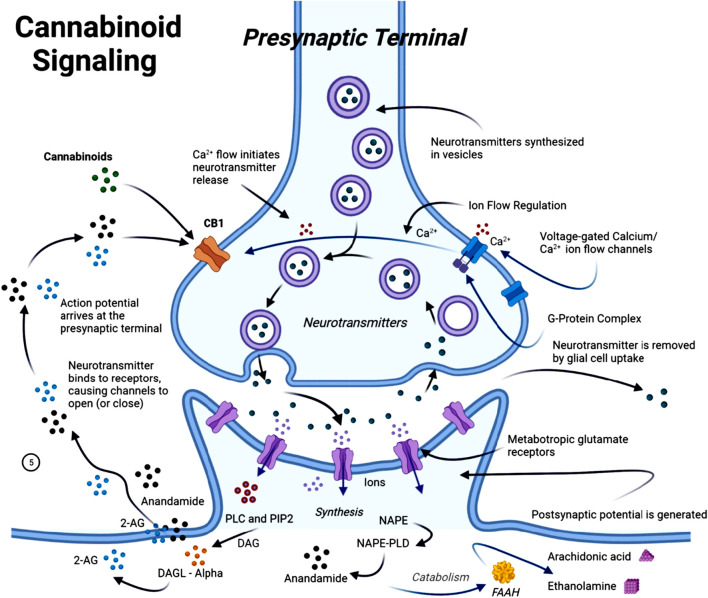
The endocannabinoid system (ECS) is a complex cell-signalling system in humans and other animals (ref. Yoder and Watson, 2017). It regulates a wide range of physiological and cognitive processes, including motor function (ref. Young and Denovan-Wright, 2022; ref. Basavarajappa et al., 2017), synaptic plasticity (ref. Piette et al., 2020; ref. Heifets and Castillo, 2009), neuroinflammation (ref. Burstein and Zurier, 2009; ref. Marchalant et al., 2009), and neural cell fate (ref. Guzmán et al., 2002; ref. Guzmán, 2003). The medical benefits of the Cannabis sativa plant date back to ancient times, with the earliest documented references appearing in the “Pen-ts’ao ching”, regarded as the world’s oldest pharmacopeia (China, 2737 BC) (ref. Pantoja-Ruiz et al., 2022; ref. Zuardi, 1999). In recent years, research interest has increasingly shifted toward targeting the ECS and investigating the use of phytocannabinoids, naturally occurring compounds that are found in the Cannabis sativa plant, as a potential therapeutic agent. This interest is largely driven by growing evidence of ECS dysfunction during the progression of AD (ref. Núñez et al., 2004; ref. Westlake et al., 1994; ref. Benito et al., 2008; ref. Russo, 2018). The ECS plays a central role in maintaining homeostasis across several physiological functions, including cognition, anxiety regulation, pain perception, neurogenesis, immune signalling, inflammation, and synaptic responsiveness and plasticity (ref. Chen et al., 2012). The ECS consists of two primary G-protein-coupled receptors, cannabinoid receptors 1 and 2 (CB1R and CB2R), as well as endocannabinoids such as anandamide (AEA) and 2-arachidonoylglycerol. It also includes metabolic enzymes such as fatty acid amide hydrolase (FAAH) and monoglyceride lipase (MAGL), which are responsible for the synthesis and degradation of endocannabinoids (ref. Alexander, 2016; ref. Aizpurua-Olaizola et al., 2017) (Figure 4).
The effects of cannabinoids on autophagy activity
Within the nervous system, the ECS is composed of naturally occurring lipid signalling pathways involved in numerous neurophysiological processes (ref. Schuster et al., 2015). This system consists of endocannabinoid receptors, primarily CB1, predominantly expressed in the CNS, and CB2, mainly expressed peripherally in the microcirculation, along with metabolic enzymes that regulate the synthesis and degradation of endocannabinoids (ref. Vrechi et al., 2021; ref. Wang Z. et al., 2022). Through these components, the ECS plays a central role in maintaining neuronal function and homeostasis (ref. Ghosh et al., 2021).
Autophagy is an evolutionarily conserved cellular degradation process that maintains cellular homeostasis and responds to proapoptotic stimuli and programmed cell death. It relies on the formation of specialized structures, including phagosomes, autophagosomes, and autophagolysosomes (ref. Padman et al., 2019). Dysregulated autophagy has been implicated in a range of human diseases, including neurodegenerative disorders, and is closely linked to inflammatory processes (ref. Ghosh et al., 2021; ref. Chung et al., 2017). Autophagy can interact with apoptosis pathways, either promoting cell survival by downregulating apoptosis or contributing to cell death when co-activated with apoptotic mechanisms (ref. Salazar et al., 2009; ref. Lu et al., 2019).
Cannabinoids, particularly cannabidiol (CBD and tetrahydrocannabinol (THC), exhibit neuroprotective and anti-inflammatory effects, partly through autophagy modulation. One possible mechanism by which cannabinoids exert anti-inflammatory and neuroprotective effects is through the regulation of mitochondrial function through autophagy (ref. Lu et al., 2019; ref. Zhou et al., 2011; ref. Ji et al., 2021). They can regulate mitochondrial function and activate peroxisome proliferator-activated receptor (PPAR)γ, restoring autophagy, modulating apoptosis, and enhancing mitochondrial β-oxidation and biosynthesis (ref. O’Sullivan, 2016; ref. Ni et al., 2022; ref. Ribeiro et al., 2012). Activation of PPAR by cannabinoids has been associated with reduced inflammation in models of acute and chronic neuronal injury (ref. Kapadia et al., 2008).
In animal studies, chronic administration of THC has been shown to influence apoptosis and immune responses in animal models, promoting cell survival and reducing inflammatory damage (ref. Molina et al., 2014).
Notably, cannabinoids modulate key autophagy-regulatory pathways. CBD inhibits phosphorylation of PI3K, Akt, and mTORC1 in human cholangiocarcinoma cells, increasing LC3B-II and decreasing p62 expression–classical markers of enhanced autophagic flux (ref. Pongking et al., 2024). Although this study used a cancer model, it demonstrates that cannabinoids can shift PI3K/Akt/mTOR signalling toward autophagy, rather than proliferation. In neuronal models, CBD induces autophagy via Akt and extracellular signal-regulated kinase (ERK) modulation, independently of mTOR (ref. Vrechi et al., 2021). Using human SH-SY5Y neuroblastoma cells, ref. Vrechi et al. (2021) reported that 10 µM CBD increased autophagic flux (LC3-II, autophagosome/lysosome formation), reduced Akt phosphorylation, and activated ERK1/2, without altering classical mTORC1 downstream targets, indicating an mTOR-independent, ULK1-dependent mechanism. CBD also enhances neuronal health and longevity via autophagy. In hippocampal neurons, SH-SY5Y cells, and in vivo (Caenorhabditis elegans), CBD promoted autophagic flux, improved neuronal integrity, these effects were dependent on core ATGs and the sirtuin pathway, suggesting a multi-pathway mechanism converging on proteostasis and neuroprotection (ref. Wang Z. et al., 2022).
Collectively, these findings indicate that cannabinoids can modulate autophagy through both PI3K/Akt/mTOR-dependent and independent mechanisms, enhancing neuronal resilience, maintaining proteostasis, and potentially counteracting neurodegenerative processes.
Cannabinoid-induced autophagy: receptor-dependent and independent pathways
Cannabinoid-induced autophagy operates through both receptor-dependent and receptor-independent mechanisms, varying with cell type and signalling context. In human SH-SY5Y neuroblastoma cells and murine astrocytes, CBD-induced autophagic flux was markedly reduced by antagonists of CB1, CB2, and transient receptor potential vanilloid 1 (TRPV1) – a Ca2+-permeable ion channel that drives autophagy via Ca2+/AMPK and ERK pathways (ref. Karki et al., 2015). This indicates that activation of these receptors contributes to CBD’s autophagy-modulating effects. CBD also modulated signalling pathways by suppressing Akt phosphorylation and activating ERK1/2, with autophagy induction dependent on ULK1, but independent of classical mTORC1 downstream targets (ref. Vrechi et al., 2021). These observations suggest a receptor-dependent initiation step that feeds into a non-classical, mTOR-independent autophagy mechanism in neuronal cells.
Conversely, there is also evidence that supports receptor-independent activation of autophagy by cannabinoids. In cancer cell models, THC and related cannabinoids induce autophagy and cell death through endoplasmic reticulum (ER) stress, ceramide accumulation, and phosphorylation of eukaryotic initiation factor 2α, leading to inhibition of Akt/mTORC1 signalling and autophagy induction independent of cannabinoid receptor activation (ref. Salazar et al., 2009). Cannabinoids can also trigger autophagy through non-receptor mechanisms such as ROS generation, ER stress, and activation of transient receptor potential channels, a family of calcium-permeable ion channels involved in Ca2+-dependent autophagy signalling (ref. Oakes et al., 2019). These findings support a broader ligand–receptor-independent network through which cannabinoids can modulate autophagy. Taken together, current evidence supports a unified model in which cannabinoids induce autophagy through both receptor-dependent and receptor-independent pathways. In receptor-expressing cells, cannabinoids activate canonical targets (CB1/CB2/TRPV1), triggering downstream signalling (e.g., ERK1/2 activation, Akt inhibition) that engages ULK1 and initiates autophagy, often independent of mTORC1 regulation. In other cellular contexts – particularly stressed, non-neuronal, or transformed cells – cannabinoids can bypass surface receptors, promoting ER stress, ceramide accumulation, or ROS generation that converge on autophagic induction via suppression of survival pathways or activation of stress-response signalling. These two complementary mechanisms help reconcile divergent findings across models and demonstrate that cannabinoid-induced autophagy is context-dependent and driven by partially overlapping pathways. In neurodegenerative settings, such flexibility may be beneficial, as non-canonical pathways could maintain pro-autophagic activity even when receptor expression is altered (e.g., CB1 downregulation), thereby supporting proteostasis and cellular clearance.
ECS alterations in AD: implications for therapeutic strategies
The ECS has demonstrated neuroprotective properties against excitotoxicity, inflammation, and oxidative stress – pathological processes central to AD and targets of many current AD therapies (ref. Xiong and Lim, 2021; ref. van der Stelt et al., 2006; ref. Trezza and Campolongo, 2013). Early evidence for a role of the ECS’s in AD was based on the high density of CB1 receptors in the hippocampus and cerebral cortex (ref. Riedel and Davies, 2005), as well as an increased expression of the endocannabinoid-metabolizing enzyme, FAAH, on plaque-associated astrocytes (ref. Benito et al., 2003).
AD is associated with ECS alterations, though it remains debated whether ECS upregulation serves to counteract neuronal hyperactivity and neuroinflammation or whether it contributes to symptoms such as memory loss (ref. O’Sullivan, 2016; ref. Ni et al., 2022). Consistently, studies have observed increased CB2R receptor expression in the brain tissue of individuals with AD and in AD-relevant animal models. This upregulation reflects immunomodulatory responses to pathogenic events. For example, clinical studies report elevated CB2R expression in microglia near hippocampal plaques (ref. Ji et al., 2021), with a positive correlation between CB2R expression and Aβ42 levels and plaque burden (ref. Ribeiro et al., 2012). Similarly, rats treated with Aβ exhibit increased CB2R levels (ref. Monteiro et al., 2020; ref. Esposito et al., 2006). Activation of microglial CB2R promotes Aβ clearance in human AD tissue sections (ref. Tolón et al., 2009), and prevents Aβ-induced microglia activation in vitro and in vivo (ref. Ramírez et al., 2005).
In contrast, reductions in CB1R receptor expression have been observed in areas of microglial activation in both humans and AD-relevant rodent models (ref. Ramírez et al., 2005), and decreases in CB1R and the endocannabinoid AEA were reported in Aβ-treated rats (ref. Esposito et al., 2007a). Moreover, cortical downregulation of AEA in AD post-mortem tissue correlates inversely with Aβ42 levels and cognitive deficits (ref. Jung et al., 2012).
The ECS holds therapeutic potential due to its broad neuromodulatory effects, which in some cases mimic the results of FDA-approved drugs or provide superior symptom relief. For example, the synthetic endocannabinoid HU-211 (Dexanabinol), acts as a stereoselective NMDA receptor inhibitor, similar to MEM, reducing calcium influx and limiting excitotoxicity (ref. Gowran et al., 2011; ref. Nadler et al., 1993). Endocannabinoid-mediated neuroprotection against excitotoxicity is a well-established and can occur through multiple mechanisms (ref. Kasatkina et al., 2021; ref. Xu et al., 2017; ref. Aguado et al., 2007; ref. Wu et al., 2019). While the AD-ECS relationship is complex and requires further investigation, these findings suggest that key ECS components are critically involved in AD pathology and represent promising therapeutic targets.
The effects of phytocannabinoids on Aβ-related pathology in AD
Cannabis sativa (commonly known as hemp or marijuana) is a multipurpose plant belonging to the family Cannabaceae, widely used for medicinal, recreational, and industrial purposes (ref. Dowling et al., 2021). The plant contains more than 500 bioactive compounds, including over 100 phytocannabinoids, with diverse pharmacological properties (ref. Gülck and Møller, 2020; ref. Caprioglio et al., 2022). These compounds have attracted significant interest for their potential therapeutic effects in neurodegenerative diseases such as AD.
Among them, THC and CBD are the most extensively studied for their neuroprotective and disease-modifying potential (ref. Elsohly and Slade, 2005; ref. Reekie et al., 2017). Although they share the same molecular formula (C21H30O2) and a terpenophenolic backbone, their distinct functional groups result in different receptor interactions and pharmacokinetic profiles. THC contains a cyclic ether (closed-ring) structure and acts primarily as a partial agonist at CB1 receptors within the CNS. In contrast, CBD contains free hydroxyl groups (open-chain), shows low direct affinity for CB1 receptors, and instead modulates the ECS indirectly through allosteric and non-canonical pathways (ref. Pertwee, 2008).
Both CBD (non-intoxicating) and THC modulate the ECS and exhibit neuroprotective, anti-inflammatory, and antioxidant properties. Emerging evidence suggests that these phytocannabinoids exert therapeutic-like effects on AD-related pathology, including Aβ accumulation and tau hyperphosphorylation (ref. Coles et al., 2022; ref. Barber et al., 2023; ref. Libro et al., 2016). Collectively, these findings highlight the therapeutic promise of cannabinoids as modulators of key neurodegenerative pathways and support further investigation into their molecular mechanisms and translational potential for AD management.
CBD’s neuroprotective potential in AD
Preclinical evidence increasingly supports the therapeutic potential of CBD in AD through multiple molecular and cellular mechanisms.
In vitro, CBD demonstrates several neuroprotective properties directly relevant to AD pathology. It inhibits tau hyperphosphorylation and reduces APP expression in APP-transfected human neuroblastoma cells. CBD also downregulates BACE1, PS1, and PS2 – key genes involved in the enzymatic generation of Aβ (ref. Libro et al., 2016). Additionally, CBD protects against Aβ-induced cytotoxicity in inducible human neuron-like cell models (ref. Schubert et al., 2019), while hemp CBD-rich hemp seed oil (∼80% CBD) significantly reduces Aβ42 + Cu(II)-induced oxidative stress (ref. Raja et al., 2020). Beyond its antioxidant effects, CBD has been shown to restore mitochondrial function (ref. Raja et al., 2020; ref. Esposito et al., 2007b), a central feature of AD pathogenesis. Recent preclinical findings further reinforce these mechanisms, showing that CBD decreases phosphorylated tau and Aβ aggregation, reduces their axonal spread between cortical and hippocampal neurons, and promotes microglial polarization toward a neuroprotective M2 phenotype (ref. Raïch et al., 2025). This study additionally showed that CBD reduces ROS formation, enhances neuronal viability, and partially restores neurite formation disrupted by Aβ and Tau, highlighting CBD’s role in supporting neuronal structural integrity.
In vivo studies further support these neuroprotective effects. Short-term CBD treatment (7 days) attenuates Aβ-evoked neuroinflammation (ref. Martín-Moreno et al., 2011), whereas chronic administration of 20 mg/kg CBD prevents learning and memory impairments in Aβ pharmacological mouse models (ref. Coles et al., 2022; ref. Cheng et al., 2014a). Dose-dependent effects have also been observed in APP/PS1 transgenic mice, where chronic CBD treatment (5–50 mg/kg) not only reverses established cognitive deficits and, at 20 mg/kg, also prevents their onset (ref. Coles et al., 2022; ref. Cheng et al., 2014b; ref. Cao et al., 2014). These results align with emerging evidence showing that chronic daily CBD administration (10 mg/kg for 28 days) significantly improves both short- and long-term spatial memory in 5xFAD mice, underscoring CBD’s capacity to ameliorate behavioral and cognitive impairments in AD models (ref. Raïch et al., 2025). Additional evidence from a sporadic AD model demonstrates that CBD restores object recognition, spatial memory, and social behavior, reduces hippocampal pro-inflammatory gene expression, mitigates increases in Aβ and p-Tau, and exerts its cognitive and anti-inflammatory effects predominantly through CB1 receptor activation (ref. Toledano and Akirav, 2025).
Although clinical data remain limited, expanding translational interest has triggered several ongoing clinical trials investigating CBD-based therapies for AD (e.g., NCT04436081, 2019-002106-52, ACTRN12621001364864). Notably, emerging evidence from a 26-week randomized controlled trial of low-dose THC-CBD extract reported significantly improved MMSE scores and a favorable safety profile compared with placebo (ref. Raïch et al., 2025).
Collectively, the preclinical evidence highlights CBD’s multimodal actions, including anti-inflammatory, antioxidant, mitochondrial-restorative, and anti-amyloidogenic, underscoring its promise as a disease-modifying candidate for AD intervention.
THC’s neuroprotective potential in AD
THC is the principal psychoactive phytocannabinoid in Cannabis sativa. Structurally, it is a diterpenoid featuring a 6a,7,8,10a-tetrahydro-6H-benzo [c]chromene core substituted by a hydroxyl group at position 1, two methyl groups at position 6, one methyl group at position 9, and a pentyl group at position 3 (ref. Tyrakis et al., 2024). Pharmacologically, THC acts primarily as a partial agonist at CB1 and CB2 receptors, producing neuroactive, analgesic, and psychotropic effects (ref. Lee et al., 2024; ref. Zou and Kumar, 2018; ref. Paronis et al., 2012). Its actions extend beyond canonical ECS to include modulation of neurotransmission, oxidative stress, and amyloid processing—mechanisms highly relevant to AD pathophysiology.
Preclinical evidence indicates that THC targets multiple AD-related pathways. In vitro, THC attenuates pathological processes in neuro-2a APPSwe cells (ref. Marsh et al., 2024). A notable finding is its competitive inhibition of AChE, preventing the breakdown of ACh, a neurotransmitter significantly depleted in AD (ref. Sam and Bordoni, 2023; ref. Puopolo et al., 2022). By enhancing cholinergic signalling, THC may provide symptomatic benefits similar to conventional AChE inhibitors but without the hepatotoxicity associated with agents such as Tacrine (Cognex). THC has also been shown to inhibit Aβ aggregation, thereby potentially reducing amyloid plaque formation and associated neurotoxicity (ref. Eubanks et al., 2006; ref. Calabrese and Rubio-Casillas, 2018).
Recent preclinical findings further reinforce THC’s mechanistic relevance in AD models. A 2024 study using APP/PS1 mice demonstrated that low-dose THC combined with CBD modulates hippocampal glutamate dynamics, reducing extracellular glutamate and hippocampal hyperexcitability – a key contributor to excitotoxic injury in AD (ref. Marsh et al., 2024). Complementing these in vivo findings, the in vitro experiments using cannabis extracts standardized for THC demonstrated neuroprotection against Aβ-induced cytotoxicity, improving neuronal viability and reducing amyloid-mediated oxidative damage.
In clinical contexts, THC-based formulations have primarily been used for appetite stimulation, antiemetic purposes, and the management of behavioral and psychological symptoms of dementia, including agitation and weight loss (223–226). Such interactions are dose-dependent and often biphasic, meaning that therapeutic outcomes depend critically on dose ratios, treatment duration, and biological context (232–234).
Although clinical evidence remains limited, research interest in THC-based interventions for AD is increasing. Two ongoing clinical trials (NCT04516057, NCT02792257) are specifically assessing THC-containing treatments for agitation in AD patients. These studies aim to determine the optimal dosing strategies, safety profiles, and whether THC – alone or in combination with CBD – may offer symptomatic relief or even disease-modifying potential. These investigations are supported by preclinical and narrative review evidence, which highlights THC’s multilayered neuroprotective effects, including anti-inflammatory, antioxidant, and anti-amyloidogenic actions (ref. Tyrakis et al., 2024), positioning it as a promising candidate for modulating AD-related neurodegeneration.
Multi-cannabinoid approaches in AD pathophysiology
Although clinical evidence remains limited, research interest in cannabinoid-based interventions for AD has increased substantially in recent years. Two ongoing trials (NCT04516057, NCT02792257) are evaluating THC-containing interventions for agitation in AD patients, reflecting a broader shift toward assessing cannabinoids for both behavioral and potential neuroprotective effects. Emerging evidence increasingly supports multi-cannabinoid formulations, particularly THC and CBD combinations, which produce effects not observed with either compound alone.
The first robust clinical evidence for such combinations was provided by ref. Cury et al. (2025), who conducted a 26-week, randomized, double-blind, placebo-controlled Phase-2 trial of a low-dose balanced THC: CBD extract (0.350 mg THC +0.245 mg CBD daily) in older adults with AD-associated dementia. The THC: CBD group showed significant improvement in Mini-Mental State Examination (MMSE) scores versus placebo, without increased adverse events (ref. Cury et al., 2025). Although secondary outcomes were unchanged, the study demonstrates that chronic low-dose THC: CBD can be safely administered and provides preliminary evidence of symptomatic cognitive benefit, warranting larger, biomarker-rich trials.
Mechanistic is provided by ref. Sánchez-Fernández et al. (2024), who examined hippocampal glutamate dynamics in APP/PS1 mice chronically treated with non-psychoactive doses of THC + CBD. The combination, but not either cannabinoid alone, reduced extracellular glutamate levels and lowered hippocampal excitability, two alterations closely linked to excitotoxic injury and cognitive decline in AD. Notably, these effects occurred without overt changes in synaptic structure or canonical synaptic plasticity markers, suggesting that THC + CBD may act through modulation of glutamate uptake, clearance, or network-level excitability rather than direct synaptic remodelling (ref. Sánchez-Fernández et al., 2024). These results support a mechanistic rationale for combination therapy, with THC and CBD appearing to produce emergent effects that cannot be inferred from single-approach studies. This highlights the importance of including neurochemical and electrophysiological biomarkers, such as magnetic resonance spectroscopy (glutamate), EEG markers of excitability, and fluid synaptic injury markers, when evaluating cannabinoid interventions in translational studies.
Further preclinical evidence from ref. Arnanz et al. (2024) in the 5xFAD mouse model, an aggressive amyloidogenic AD model, showed that chronic low-dose treatment with a THC: CBD combination improved spatial memory, whereas the single agents did not. Interestingly, all cannabinoid-treated groups, including the combination, exhibited increased Aβ42 cortical levels, highlighting complex effects on amyloid processing. Cognitive improvements occurred without significant changes in classical inflammatory markers, underscoring that multi-cannabinoid therapy may exert functional benefits via mechanisms distinct from classical anti-inflammatory pathways. These findings indicate that while combination therapy offers synergistic behavioural benefits, careful longitudinal assessment of amyloid and other disease-modifying endpoints is essential.
Collectively, these studies highlight that multi-cannabinoid regimens exert synergistic effects across cognitive, excitotoxic, and network-level outcomes. THC and CBD engage complementary mechanisms, including CB1/CB2 modulation, glutamate regulation, and antioxidant activity, while divergent effects on amyloid pathology underscore the need for mechanistically informed, biomarker-guided clinical trials. Overall, the evidence provides a strong rationale for advancing multi-cannabinoid therapeutics in AD, with clear priorities for dose optimization, biomarker integration, and longitudinal evaluation to determine both symptomatic and disease-modifying potential in future translational and clinical studies.
Potential cannabinoid interactions with FDA-approved AD therapies
Multi-cannabinoid interventions in AD raise important considerations regarding interactions with FDA-approved therapies, including AChEIs and MEM. Recent in vitro studies demonstrate that CBD, THC, and other cannabinoids moderately inhibit AChE and butyrylcholinesterase (BChE), a related cholinergic enzyme that hydrolyzes acetylcholine and other choline esters. In AD, BChE activity increases in later stages, and its inhibition can help sustain cholinergic signaling, complementing AChE-targeted therapies (ref. Puopolo et al., 2022; ref. Patil et al., 2023). While these interactions suggest potential synergy, high doses could theoretically amplify cholinergic effects and increase adverse events.
Interactions with MEM are more complex: both cannabinoids and MEM modulate glutamatergic signaling, with preclinical data indicating potential for synergistic neuroprotection as well as receptor-level antagonism depending on dose and context (ref. Marszalek-Grabska et al., 2021). Pharmacokinetic interactions are also notable, as CBD and THC influence cytochrome P450 enzymes (CYP3A4, CYP2C19, CYP2D6), which metabolize many AChEIs and memantine, highlighting the need for dose optimization and careful monitoring in older adults with polypharmacy (ref. Balachandran et al., 2021; ref. Beers et al., 2021).
Future studies should prioritize preclinical combination models alongside early-phase clinical trials to comprehensively evaluate safety, pharmacokinetics, pharmacodynamics, and efficacy. This includes preclinical experiments assessing cognitive, synaptic, and biomarker outcomes, as well as phase 1-2 trials examining cannabinoids in combination with AChEIs or MEM. Such research is essential to determine whether multi-cannabinoid therapy can safely and effectively complement existing AD treatments, optimizing symptomatic relief while minimizing potential adverse interactions.
Conclusion
The therapeutic potential of cannabinoid-based interventions for AD is well-supported; however, the inherent chemical complexity of Cannabis sativa continues to pose significant translational challenges. Precisely defined formulations, as well as careful control of dose, ratio, and route of administration routes, remain essential to achieving consistent therapeutic outcomes. Emerging evidence, including a recent case report (ref. Ruver-Martins et al., 2022), suggests that cannabinoid microdosing may offer a potential strategy for reducing AD-related symptoms while minimizing adverse effects. Yet, these preliminary findings require rigorous validation through larger, well-controlled clinical studies. Pharmacokinetic data further indicate that multi-cannabinoid formulations, particularly those combining THC and CBD, add additional minor cannabinoids, may provide enhanced therapeutic efficacy and improved safety profiles compared to monotherapy (ref. Zuardi et al., 2012). However, it is important to note that evidence on cannabinoid-induced autophagy in AD remains limited and heterogeneous. Most studies are preclinical, with variations in cannabinoid type, dose, and measurement of autophagy markers, and quantitative data in disease-relevant models are sparse, highlighting a critical knowledge gap. Although both in vitro and in vivo studies increasingly support the potential of such combination therapies, the mechanistic interplay between multi-cannabinoid therapeutics, autophagy modulation, and neuronal survival remains insufficiently understood. Therefore, there is an urgent need for comprehensive, mechanistically grounded research to define the optimal THC: CBD ratio and broader cannabinoid profiles capable of effectively modulating autophagy, mitigating amyloidogenic and inflammatory pathways, and ultimately providing robust neuroprotection in AD-relevant experimental systems. Such work will be critical for translating cannabinoid-based treatments from promising preclinical leads into safe, targeted, and effective clinical interventions for AD.
References
- L. Agnello, M. Ciaccio. Neurodegenerative diseases: from molecular basis to therapy.. Int. J. Mol. Sci., 2022. [DOI | PubMed]
- T. Aguado, E. Romero, K. Monory, J. Palazuelos, M. Sendtner, G. Marsicano. The CB1 cannabinoid receptor mediates excitotoxicity-induced neural progenitor proliferation and neurogenesis.. J. Biol. Chem., 2007. [DOI | PubMed]
- O. Aizpurua-Olaizola, I. Elezgarai, I. Rico-Barrio, I. Zarandona, N. Etxebarria, A. Usobiaga. Targeting the endocannabinoid system: future therapeutic strategies.. Drug Discov. Today, 2017. [DOI | PubMed]
- D. O. T. Alawode, N. C. Fox, H. Zetterberg, A. J. Heslegrave. Alzheimer’s disease biomarkers revisited from the amyloid Cascade hypothesis standpoint.. Front. Neurosci., 2022. [DOI | PubMed]
- S. P. H. Alexander. Therapeutic potential of cannabis-related drugs.. Prog. Neuropsychopharmacol. Biol. Psychiatry, 2016. [DOI | PubMed]
- H. A. Alhazmi, M. Albratty. An update on the novel and approved drugs for Alzheimer disease.. Saudi Pharm. J. SPJ, 2022. [DOI | PubMed]
- Y. Aman, T. Schmauck-Medina, M. Hansen, R. I. Morimoto, A. K. Simon, I. Bjedov. Autophagy in healthy aging and disease.. Nat. Aging, 2021. [DOI | PubMed]
- R. M. Anderson, C. Hadjichrysanthou, S. Evans, M. M. Wong. Why do so many clinical trials of therapies for Alzheimer’s disease fail?. Lancet, 2017. [DOI | PubMed]
- M. A. Arnanz, S. Ruiz de Martín Esteban, A. M. Martínez Relimpio, N. Rimmerman, N. Tweezer Zaks, M. T. Grande. Effects of chronic, low-dose cannabinoids, cannabidiol, Delta-9-Tetrahydrocannabinol and a combination of both, on amyloid pathology in the 5xFAD mouse model of alzheimer’s disease.. Cannabis Cannabinoid Res., 2024. [DOI | PubMed]
- L. B, T. A du, H. Jh. Defining and measuring autophagosome Flux—concept and reality.. Autophagy, 2014. [DOI | PubMed]
- P. Balachandran, M. Elsohly, K. P. Hill. Cannabidiol interactions with medications, illicit substances, and alcohol: a comprehensive review.. J. Gen. Intern Med., 2021. [DOI | PubMed]
- K. Barber, P. Mendonca, K. F. A. Soliman. The neuroprotective effects and therapeutic potential of the chalcone cardamonin for alzheimer’s disease.. Brain Sci., 2023. [DOI | PubMed]
- B. S. Basavarajappa, M. Shivakumar, V. Joshi, S. Subbanna. Endocannabinoid system in neurodegenerative disorders.. J. Neurochem., 2017. [DOI | PubMed]
- J. L. Beers, D. Fu, K. D. Jackson. Cytochrome P450–Catalyzed metabolism of cannabidiol to the active metabolite 7-Hydroxy-Cannabidiol.. Drug Metab. Dispos., 2021. [DOI | PubMed]
- C. Benito, E. Núñez, R. M. Tolón, E. J. Carrier, A. Rábano, C. J. Hillard. Cannabinoid CB2 receptors and fatty acid amide hydrolase are selectively overexpressed in neuritic plaque-associated glia in Alzheimer’s disease brains.. J. Neurosci. Off. J. Soc. Neurosci., 2003. [DOI | PubMed]
- C. Benito, R. M. Tolón, M. R. Pazos, E. Núñez, A. I. Castillo, J. Romero. Cannabinoid CB2 receptors in human brain inflammation.. Br. J. Pharmacol., 2008. [DOI | PubMed]
- P. Bergström, L. Agholme, F. H. Nazir, T. M. Satir, J. Toombs, H. Wellington. Amyloid precursor protein expression and processing are differentially regulated during cortical neuron differentiation.. Sci. Rep., 2016. [DOI | PubMed]
- D. Bloemberg, J. Quadrilatero. Autophagy, apoptosis, and mitochondria: molecular integration and physiological relevance in skeletal muscle.. Am. J. Physiol. – Cell Physiol., 2019. [DOI | PubMed]
- B. Boland, A. Kumar, S. Lee, F. M. Platt, J. Wegiel, W. H. Yu. Autophagy induction and autophagosome clearance in neurons: relationship to autophagic pathology in Alzheimer’s disease.. J. Neurosci. Off. J. Soc. Neurosci., 2008. [DOI | PubMed]
- M. Bordi, M. J. Berg, P. S. Mohan, C. M. Peterhoff, M. J. Alldred, S. Che. Autophagy flux in CA1 neurons of Alzheimer hippocampus: increased induction overburdens failing lysosomes to propel neuritic dystrophy.. Autophagy, 2016. [DOI | PubMed]
- F. Boutouja, C. M. Stiehm, H. W. Platta. mTOR: a cellular regulator interface in health and disease.. Cells, 2019. [DOI | PubMed]
- S. H. Burstein, R. B. Zurier. Cannabinoids, endocannabinoids, and related analogs in inflammation.. AAPS J., 2009. [DOI | PubMed]
- Z. Cai, Y. Zhou, Z. Liu, Z. Ke, B. Zhao. Autophagy dysfunction upregulates beta-amyloid peptides via enhancing the activity of γ-secretase complex.. Neuropsychiatr. Dis. Treat., 2015. [DOI | PubMed]
- E. J. Calabrese, A. Rubio-Casillas. Biphasic effects of THC in memory and cognition.. Eur. J. Clin. Invest, 2018. [DOI | PubMed]
- C. Cao, Y. Li, H. Liu, G. Bai, J. Mayl, X. Lin. The potential therapeutic effects of THC on Alzheimer’s disease.. J. Alzheimers Dis. Jad., 2014. [DOI | PubMed]
- D. Caprioglio, H. I. M. Amin, O. Taglialatela-Scafati, E. Muñoz, G. M. AppendinoPhytocannabinoids. A misleading name but a promising opportunity for biomedical research.. Biomolecules, 2022. [DOI | PubMed]
- E. Carro, J. L. Trejo, C. Spuch, D. Bohl, J. M. Heard, I. Torres-Aleman. Blockade of the insulin-like growth factor I receptor in the choroid plexus originates alzheimer’s-like neuropathology in rodents: new cues into the human disease?. Neurobiol. Aging, 2006. [DOI | PubMed]
- M. Catania, G. Giaccone, M. Salmona, F. Tagliavini, G. Di Fede. Dreaming of a new world where alzheimer’s is a treatable disorder.. Front. Aging Neurosci., 2019. [DOI | PubMed]
- R. Chen, J. Zhang, Y. Wu, D. Wang, G. Feng, Y. P. Tang. Monoacylglycerol lipase is a new therapeutic target for Alzheimer’s disease.. Cell Rep., 2012. [DOI | PubMed]
- Z. R. Chen, J. B. Huang, S. L. Yang, F. F. Hong. Role of cholinergic signaling in Alzheimer’s disease.. Molecules, 2022. [DOI | PubMed]
- D. Cheng, J. K. Low, W. Logge, B. Garner, T. Karl. Chronic cannabidiol treatment improves social and object recognition in double transgenic APPswe/PS1ΔE9 mice.. Psychopharmacol. Berl., 2014a. [DOI | PubMed]
- D. Cheng, A. S. Spiro, A. M. Jenner, B. Garner, T. Karl. Long-term cannabidiol treatment prevents the development of social recognition memory deficits in Alzheimer’s disease transgenic mice.. J. Alzheimers Dis. Jad., 2014b. [DOI | PubMed]
- K. W. Chung, K. M. Kim, Y. J. Choi, H. J. An, B. Lee, D. H. Kim. The critical role played by endotoxin-induced liver autophagy in the maintenance of lipid metabolism during sepsis.. Autophagy, 2017. [DOI | PubMed]
- M. Coles, G. Z. Steiner-Lim, T. Karl. Therapeutic properties of multi-cannabinoid treatment strategies for Alzheimer’s disease.. Front. Neurosci., 2022. [DOI | PubMed]
- J. Cui, C. Zhao. Autophagy and apoptosis in Alzheimer’s disease-associated neurons.. Discov. Neurosci., 2025. [DOI]
- J. L. Cummings, D. P. Goldman, N. R. Simmons-Stern, E. Ponton. The costs of developing treatments for Alzheimer’s disease: a retrospective exploration.. Alzheimers Dement., 2022. [DOI | PubMed]
- J. Cummings, G. Lee, P. Nahed, MEZN Kambar, K. Zhong, J. Fonseca. Alzheimer’s disease drug development pipeline: 2022.. Alzheimers Dement. Transl. Res. Clin. Interv., 2022. [DOI | PubMed]
- R. de M. Cury, T. da Silva, F. Cezar-dos-Santos, Y. R. C. Fakih, K. A. R. Narvaez, M. C. Gouvea. A randomized clinical trial of low-dose cannabis extract in Alzheimer’s disease.. J. Alzheimer’s Dis.,, 2025. [DOI | PubMed]
- I. de Rojas, S. Moreno-Grau, N. Tesi, B. Grenier-Boley, V. Andrade, I. E. Jansen. Common variants in Alzheimer’s disease and risk stratification by polygenic risk scores.. Nat. Commun., 2021. [DOI | PubMed]
- M. A. DeTure, D. W. Dickson. The neuropathological diagnosis of Alzheimer’s disease.. Mol. Neurodegener., 2019. [DOI | PubMed]
- M. C. Dinamarca, J. P. Sagal, R. A. Quintanilla, J. A. Godoy, M. S. Arrázola, N. C. Inestrosa. Amyloid-β-Acetylcholinesterase complexes potentiate neurodegenerative changes induced by the Aβ peptide. Implications for the pathogenesis of Alzheimer’s disease.. Mol. Neurodegener., 2010. [DOI | PubMed]
- C. A. Dowling, R. Melzer, S. Schilling. Timing is everything: the genetics of flowering time in Cannabis sativa.. Biochem., 2021. [DOI]
- P. d‘Errico, M. Meyer-Luehmann. Mechanisms of pathogenic tau and Aβ protein spreading in alzheimer’s disease.. Front. Aging Neurosci., 2020. [DOI | PubMed]
- M. A. Elsohly, D. Slade. Chemical constituents of marijuana: the complex mixture of natural cannabinoids.. Life Sci., 2005. [DOI | PubMed]
- G. Esposito, D. De Filippis, R. Carnuccio, A. A. Izzo, T. Iuvone. The marijuana component cannabidiol inhibits beta-amyloid-induced tau protein hyperphosphorylation through wnt/beta-catenin pathway rescue in PC12 cells.. J. Mol. Med. Berl. Ger., 2006. [DOI | PubMed]
- G. Esposito, T. Iuvone, C. Savani, C. Scuderi, D. De Filippis, M. Papa. Opposing control of cannabinoid receptor stimulation on amyloid-beta-induced reactive gliosis: in vitro and in vivo evidence.. J. Pharmacol. Exp. Ther., 2007a. [DOI | PubMed]
- G. Esposito, C. Scuderi, C. Savani, L. Steardo, D. De Filippis, P. Cottone. Cannabidiol in vivo blunts beta-amyloid induced neuroinflammation by suppressing IL-1beta and iNOS expression.. Br. J. Pharmacol., 2007b. [DOI | PubMed]
- L. M. Eubanks, C. J. Rogers, A. E. Beuscher, G. F. Koob, A. J. Olson, T. J. Dickerson. A molecular link between the active component of marijuana and alzheimer’s disease pathology.. Mol. Pharm., 2006. [DOI | PubMed]
- C. S. Evans, E. L. F. Holzbaur. Quality control in neurons: mitophagy and other selective autophagy mechanisms.. J. Mol. Biol., 2020. [DOI | PubMed]
- J. W. Fawcett. The struggle to make CNS axons regenerate: why has it been so difficult?. Neurochem. Res., 2020. [DOI | PubMed]
- J. Folch, O. Busquets, M. Ettcheto, E. Sánchez-López, R. D. Castro-Torres, E. Verdaguer. Memantine for the treatment of dementia: a review on its current and future applications.. J. Alzheimers Dis., 2018. [DOI | PubMed]
- L. K. Fong, M. M. Yang, R. dos Santos Chaves, S. M. Reyna, V. F. Langness, G. Woodruff. Full-length amyloid precursor protein regulates lipoprotein metabolism and amyloid-β clearance in human astrocytes.. J. Biol. Chem., 2018. [DOI | PubMed]
- L. Galluzzi, E. H. Baehrecke, A. Ballabio, P. Boya, J. M. Bravo-San Pedro, F. Cecconi. Molecular definitions of autophagy and related processes.. EMBO J., 2017. [DOI | PubMed]
- Y. Gao, A. Moten, H. K. Lin. Akt: a new activation mechanism.. Cell Res., 2014. [DOI | PubMed]
- D. Garcia, R. J. Shaw. AMPK: mechanisms of cellular energy sensing and restoration of metabolic balance.. Mol. Cell, 2017. [DOI | PubMed]
- C. García, C. Palomo-Garo, M. García-Arencibia, J. Ramos, R. Pertwee, J. Fernández-Ruiz. Symptom-relieving and neuroprotective effects of the phytocannabinoid Δ9-THCV in animal models of Parkinson’s disease.. Br. J. Pharmacol., 2011. [DOI | PubMed]
- M. S. García-Ayllón, D. H. Small, J. Avila, J. Sáez-Valero. Revisiting the role of acetylcholinesterase in alzheimer’s disease: cross-talk with P-tau and β-Amyloid.. Front. Mol. Neurosci., 2011. [DOI | PubMed]
- Dementia forecasting collaborators. Estimation of the global prevalence of dementia in 2019 and forecasted prevalence in 2050: an analysis for the global burden of disease study 2019.. Lancet Public Health, 2022. [DOI | PubMed]
- S. Ghosh, K. Stansak, B. J. Walters. Cannabinoid signaling in auditory function and development.. Front. Mol. Neurosci., 2021. [DOI | PubMed]
- L. R. Gomes, C. F. M. Menck, A. M. Cuervo. Chaperone-mediated autophagy prevents cellular transformation by regulating MYC proteasomal degradation.. Autophagy, 2017. [DOI | PubMed]
- C. X. Gong, F. Liu, K. Iqbal. Multifactorial hypothesis and multi-targets for alzheimer’s disease.. J. Alzheimers Dis., 2018. [DOI | PubMed]
- A. Gowran, J. Noonan, V. A. Campbell. The multiplicity of action of cannabinoids: implications for treating neurodegeneration.. CNS Neurosci. Ther., 2011. [DOI | PubMed]
- T. Gülck, B. L. Møller. Phytocannabinoids: origins and biosynthesis.. Trends Plant Sci., 2020. [DOI | PubMed]
- M. Guzmán. Neurons on cannabinoids: dead or alive?. Br. J. Pharmacol., 2003. [DOI | PubMed]
- M. Guzmán, C. Sánchez, I. Galve-Roperh. Cannabinoids and cell fate.. Pharmacol. Ther., 2002. [DOI | PubMed]
- T. H Ferreira-Vieira, I. M Guimaraes, F. R Silva, F. M Ribeiro. Alzheimer’s disease: targeting the cholinergic system.. Curr. Neuropharmacol., 2016. [DOI | PubMed]
- C. Haass, D. Selkoe. If amyloid drives Alzheimer disease, why have anti-amyloid therapies not yet slowed cognitive decline?. PLOS Biol., 2022. [DOI | PubMed]
- D. Halawani, S. Tessier, D. Anzellotti, D. A. Bennett, M. Latterich, A. C. LeBlanc. Identification of Caspase-6-mediated processing of the valosin containing protein (p97) in Alzheimer’s disease: a novel link to dysfunction in ubiquitin proteasome system-mediated protein degradation.. J. Neurosci. Off. J. Soc. Neurosci., 2010. [DOI | PubMed]
- H. Hampel, R. Vassar, B. De Strooper, J. Hardy, M. Willem, N. Singh. The β-Secretase BACE1 in alzheimer’s disease.. Biol. Psychiatry, 2021. [DOI | PubMed]
- J. A. Hardy, G. A. Higgins. Alzheimer’s disease: the amyloid Cascade hypothesis.. Science, 1992. [DOI | PubMed]
- D. E. Harrison, R. Strong, Z. D. Sharp, J. F. Nelson, C. M. Astle, K. Flurkey. Rapamycin fed late in life extends lifespan in genetically heterogeneous mice.. Nature, 2009. [DOI | PubMed]
- M. Hashimoto, E. Rockenstein, L. Crews, E. Masliah. Role of protein aggregation in mitochondrial dysfunction and neurodegeneration in Alzheimer’s and Parkinson’s diseases.. Neuromolecular Med., 2003. [DOI | PubMed]
- A. Hector, J. Brouillette. Hyperactivity induced by soluble Amyloid-β oligomers in the early stages of alzheimer’s disease.. Front. Mol. Neurosci., 2021. [DOI | PubMed]
- B. D. Heifets, P. E. Castillo. Endocannabinoid signaling and long-term synaptic plasticity.. Annu. Rev. Physiol., 2009. [DOI | PubMed]
- L. Ibanez, C. Cruchaga, M. V. Fernández. Advances in genetic and molecular understanding of alzheimer’s disease.. Genes, 2021. [DOI | PubMed]
- M. O. Iliyasu, S. A. Musa, S. B. Oladele, A. I. Iliya. Amyloid-beta aggregation implicates multiple pathways in Alzheimer’s disease: understanding the mechanisms.. Front. Neurosci., 2023. [DOI | PubMed]
- P. A. Jaeger, T. Wyss-Coray. All-you-can-eat: autophagy in neurodegeneration and neuroprotection.. Mol. Neurodegener., 2009. [DOI | PubMed]
- W. Ji, T. Wan, F. Zhang, X. Zhu, S. Guo, X. Mei. Aldehyde dehydrogenase 2 protects against lipopolysaccharide-induced myocardial injury by suppressing mitophagy.. Front. Pharmacol., 2021. [DOI | PubMed]
- M. P. Jogalekar, A. Veerabathini, P. Gangadaran. Recent developments in autophagy-targeted therapies in cancer.. Exp. Biol. Med., 2021. [DOI | PubMed]
- K. M. Jung, G. Astarita, S. Yasar, V. Vasilevko, D. H. Cribbs, E. Head. An amyloid β42-dependent deficit in anandamide mobilization is associated with cognitive dysfunction in Alzheimer’s disease.. Neurobiol. Aging, 2012. [DOI | PubMed]
- S. Kang, J. Li, Z. Yao, J. Liu. Cannabidiol induces autophagy to protects neural cells from mitochondrial dysfunction by upregulating SIRT1 to inhibits NF-κB and NOTCH pathways.. Front. Cell Neurosci., 2021. [DOI | PubMed]
- R. Kapadia, J. H. Yi, R. Vemuganti. Mechanisms of anti-inflammatory and neuroprotective actions of PPAR-gamma agonists.. Front. Biosci. J. Virtual Libr., 2008. [DOI | PubMed]
- P. Karki, K. Smith, J. Johnson, M. Aschner, E. Lee. Role of transcription factor yin yang 1 in manganese-induced reduction of astrocytic glutamate transporters: putative mechanism for manganese-induced neurotoxicity.. Neurochem. Int., 2015. [DOI | PubMed]
- G. Karsli-Uzunbas, J. Y. Guo, S. Price, X. Teng, S. V. Laddha, S. Khor. Autophagy is required for glucose homeostasis and lung tumor maintenance.. Cancer Discov., 2014. [DOI | PubMed]
- L. A. Kasatkina, S. Rittchen, E. M. Sturm. Neuroprotective and immunomodulatory action of the endocannabinoid system under neuroinflammation.. Int. J. Mol. Sci., 2021. [DOI | PubMed]
- G. P. Kaushal, K. Chandrashekar, L. A. Juncos. Molecular interactions between reactive oxygen species and autophagy in kidney disease.. Int. J. Mol. Sci., 2019. [DOI | PubMed]
- K. H. Khan, T. A. Yap, L. Yan, D. Cunningham. Targeting the PI3K-AKT-mTOR signaling network in cancer.. Chin. J. Cancer, 2013. [DOI | PubMed]
- S. W. Kim, J. H. Lee, J. H. Moon, U. M. D. Nazim, Y. J. Lee, J. W. Seol. Niacin alleviates TRAIL-mediated colon cancer cell death via autophagy flux activation.. Oncotarget, 2015. [DOI | PubMed]
- B. L. Klaassens, J. M. van Gerven, E. S. Klaassen, J. van der Grond, S. A. Rombouts. Cholinergic and serotonergic modulation of resting state functional brain connectivity in Alzheimer’s disease.. Neuroimage., 2019. [DOI | PubMed]
- D. J. Klionsky, G. Petroni, R. K. Amaravadi, E. H. Baehrecke, A. Ballabio, P. Boya. Autophagy in major human diseases.. EMBO J., 2021. [DOI | PubMed]
- L. Kma, T. J. Baruah. The interplay of ROS and the PI3K/Akt pathway in autophagy regulation.. Biotechnol. Appl. Biochem., 2022. [DOI | PubMed]
- H. Kuang, C. Y. Tan, H. Z. Tian, L. H. Liu, M. W. Yang, F. F. Hong. Exploring the bi-directional relationship between autophagy and Alzheimer’s disease.. CNS Neurosci. Ther., 2020. [DOI | PubMed]
- M. F. Kuo, J. Grosch, F. Fregni, W. Paulus, M. A. Nitsche. Focusing effect of acetylcholine on neuroplasticity in the human motor cortex.. J. Neurosci., 2007. [DOI | PubMed]
- H. M. Lanoiselée, G. Nicolas, D. Wallon, A. Rovelet-Lecrux, M. Lacour, S. Rousseau. APP, PSEN1, and PSEN2 mutations in early-onset Alzheimer disease: a genetic screening study of familial and sporadic cases.. PLoS Med., 2017. [DOI | PubMed]
- I. Lastres-Becker, F. Molina-Holgado, J. A. Ramos, R. Mechoulam, J. Fernández-Ruiz. Cannabinoids provide neuroprotection against 6-hydroxydopamine toxicity in vivo and in vitro: relevance to Parkinson’s disease.. Neurobiol. Dis., 2005. [DOI | PubMed]
- H. K. Lee, B. Kwon, C. A. Lemere, S. de la Monte, K. Itamura, A. Y. Ha. mTORC2 (Rictor) in alzheimer’s disease and reversal of Amyloid-β expression-induced insulin resistance and toxicity in rat primary cortical neurons.. J. Alzheimers Dis. Jad., 2017. [DOI | PubMed]
- J. H. Lee, D. S. Yang, C. N. Goulbourne, E. Im, P. Stavrides, A. Pensalfini. Faulty autolysosome acidification in Alzheimer’s disease mouse models induces autophagic build-up of Aβ in neurons, yielding senile plaques.. Nat. Neurosci., 2022. [DOI | PubMed]
- S. Lee, Y. Lee, Y. Kim, H. Kim, H. Rhyu, K. Yoon. Beneficial effects of cannabidiol from cannabis.. Appl. Biol. Chem., 2024. [DOI]
- M. A. Lemmon, J. Schlessinger. Cell signaling by receptor tyrosine kinases.. Cell, 2010. [DOI | PubMed]
- B. Levine, G. Kroemer. Biological functions of autophagy genes: a disease perspective.. Cell, 2019. [DOI | PubMed]
- Y. Li, Y. Chen. AMPK and autophagy.. Adv. Exp. Med. Biol., 2019. [DOI | PubMed]
- X. zhi Li, Y. X. hua. Sensors for the mTORC1 pathway regulated by amino acids.. J. Zhejiang Univ. Sci. B, 2019. [DOI | PubMed]
- T. Li, A. Li, X. Pan. Targeting impaired autophagy as a therapeutic strategy for Koolen-de vries syndrome.. Autophagy, 2022. [DOI | PubMed]
- R. Libro, F. Diomede, D. Scionti, A. Piattelli, G. Grassi, F. Pollastro. Cannabidiol modulates the expression of alzheimer’s disease-related genes in mesenchymal stem cells.. Int. J. Mol. Sci., 2016. [DOI | PubMed]
- S. A. Lipton, H. V. Chen. Paradigm shift in NMDA receptor drug development.. Expert Opin. Ther. Targets, 2005. [DOI | PubMed]
- Y. Lu, R. Li, J. Zhu, Y. Wu, D. Li, L. Dong. Fibroblast growth factor 21 facilitates peripheral nerve regeneration through suppressing oxidative damage and autophagic cell death.. J. Cell Mol. Med., 2019. [DOI | PubMed]
- R. Lucà, M. R. Assenza, F. Maiullari, L. Pieroni, S. Maiullari, G. Federici. Inhibition of the mTOR pathway and reprogramming of protein synthesis by MDM4 reduce ovarian cancer metastatic properties.. Cell Death Dis., 2021. [DOI | PubMed]
- A. Machado, D. Ferreira, M. J. Grothe, H. Eyjolfsdottir, P. M. Almqvist, L. Cavallin. The cholinergic system in subtypes of Alzheimer’s disease: an in vivo longitudinal MRI study.. Alzheimers Res. Ther., 2020. [DOI | PubMed]
- Y. Marchalant, H. M. Brothers, G. J. Norman, K. Karelina, A. C. DeVries, G. L. Wenk. Cannabinoids attenuate the effects of aging upon neuroinflammation and neurogenesis.. Neurobiol. Dis., 2009. [DOI | PubMed]
- D. T. Marsh, M. Shibuta, R. Kato, S. D. Smid. Medicinal cannabis extracts are neuroprotective against Aβ1-42-mediated toxicity in vitro .. Basic Clin. Pharmacol. Toxicol., 2024. [DOI | PubMed]
- M. Marszalek-Grabska, I. Smaga, P. Surowka, P. Grochecki, T. Slowik, M. Filip. Memantine prevents the WIN 55,212-2 evoked cross-priming of ethanol-induced conditioned place preference (CPP).. Int. J. Mol. Sci., 2021. [DOI | PubMed]
- A. M. Martín-Moreno, D. Reigada, B. G. Ramírez, R. Mechoulam, N. Innamorato, A. Cuadrado. Cannabidiol and other cannabinoids reduce microglial activation in vitro and in vivo: relevance to Alzheimer’s disease.. Mol. Pharmacol., 2011. [DOI | PubMed]
- B. M. McGleenon, K. B. Dynan, A. P. Passmore. Acetylcholinesterase inhibitors in Alzheimer’s disease.. Br. J. Clin. Pharmacol., 1999. [DOI | PubMed]
- N. C. McKnight, Y. Zhenyu. Beclin 1, an essential component and master regulator of PI3K-III in health and disease.. Curr. Pathobiol. Rep., 2013. [DOI | PubMed]
- M. B. Menon, S. Dhamija. Beclin 1 phosphorylation – at the center of autophagy regulation.. Front. Cell Dev. Biol., 2018. [DOI | PubMed]
- M. Mistretta, A. Farini, Y. Torrente, C. Villa. Multifaceted nanoparticles: emerging mechanisms and therapies in neurodegenerative diseases.. Brain, 2023. [DOI | PubMed]
- P. E. Molina, A. M. Amedee, N. J. LeCapitaine, J. Zabaleta, M. Mohan, P. J. Winsauer. Modulation of Gut-specific mechanisms by chronic Δ9-Tetrahydrocannabinol administration in male rhesus macaques infected with simian immunodeficiency virus: a systems biology analysis.. AIDS Res. Hum. Retroviruses, 2014. [DOI | PubMed]
- K. L. C. Monteiro, M. G. dos Santos Alcântara, T. M. de Aquino, E. F. da Silva-Júnior. Cannabinoid pharmacology and its therapeutic uses in Alzheimer’s disease.. Neural Regen. Res., 2020. [DOI | PubMed]
- J. E. Morley, S. A. Farr, A. D. Nguyen, F. Xu. What is the physiological function of amyloid-beta protein?. J. Nutr. Health Aging, 2019. [DOI | PubMed]
- D. E. Moss. Improving anti-neurodegenerative benefits of acetylcholinesterase inhibitors in alzheimer’s disease: are irreversible inhibitors the future?. Int. J. Mol. Sci., 2020. [DOI | PubMed]
- V. Nadler, R. Mechoulam, M. Sokolovsky. Blockade of 45Ca2+ influx through the N-methyl-D-aspartate receptor ion channel by the non-psychoactive cannabinoid HU-211.. Brain Res., 1993. [DOI | PubMed]
- J. Ni, H. Zhu, L. Lu, Z. Zhao, J. Jiang, X. You. Hydroxychloroquine induces apoptosis of myeloid-derived suppressor cells via up-regulation of CD81 contributing to alleviate lupus symptoms.. Mol. Med. Camb Mass, 2022. [DOI | PubMed]
- P. Nilsson, T. C. Saido. Dual roles for autophagy: degradation and secretion of Alzheimer’s disease Aβ peptide.. BioEssays News Rev. Mol. Cell Dev. Biol., 2014. [DOI | PubMed]
- P. Nilsson, K. Loganathan, M. Sekiguchi, Y. Matsuba, K. Hui, S. Tsubuki. Aβ secretion and plaque Formation depend on autophagy.. Cell Rep., 2013. [DOI | PubMed]
- P. Nilsson, M. Sekiguchi, T. Akagi, S. Izumi, T. Komori, K. Hui. Autophagy-related protein 7 deficiency in amyloid β (Aβ) precursor protein transgenic mice decreases Aβ in the multivesicular bodies and induces Aβ accumulation in the golgi.. Am. J. Pathol., 2014. [DOI | PubMed]
- R. A. Nixon. Autophagy, amyloidogenesis and Alzheimer disease.. J. Cell Sci., 2007. [DOI | PubMed]
- R. A. Nixon. Amyloid precursor protein and endosomal-lysosomal dysfunction in Alzheimer’s disease: inseparable partners in a multifactorial disease.. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol., 2017. [DOI | PubMed]
- R. A. Nixon, D. S. Yang. Autophagy failure in Alzheimer’s disease–locating the primary defect.. Neurobiol. Dis., 2011. [DOI | PubMed]
- R. A. Nixon, D. S. Yang. Autophagy and neuronal cell death in neurological disorders.. Cold Spring Harb. Perspect. Biol., 2012. [DOI | PubMed]
- R. A. Nixon, J. Wegiel, A. Kumar, W. H. Yu, C. Peterhoff, A. Cataldo. Extensive involvement of autophagy in Alzheimer disease: an immuno-electron microscopy study.. J. Neuropathol. Exp. Neurol., 2005. [DOI | PubMed]
- C. Ntsapi, B. Loos. Caloric restriction and the precision-control of autophagy: a strategy for delaying neurodegenerative disease progression.. Exp. Gerontol., 2016. [DOI | PubMed]
- C. M. Ntsapi, B. Loos. Neurons die with heightened but functional macro- and chaperone mediated autophagy upon increased amyloid-ß induced toxicity with region-specific protection in prolonged intermittent fasting.. Exp. Cell Res., 2021. [DOI | PubMed]
- C. Ntsapi, D. Lumkwana, C. Swart, A. du Toit, B. Loos. New insights into autophagy dysfunction related to amyloid beta toxicity and neuropathology in alzheimer’s disease.. Int. Rev. Cell Mol. Biol., 2018. [DOI | PubMed]
- E. Núñez, C. Benito, M. R. Pazos, A. Barbachano, O. Fajardo, S. González. Cannabinoid CB2 receptors are expressed by perivascular microglial cells in the human brain: an immunohistochemical study.. Synap. N. Y. N., 2004. [DOI | PubMed]
- M. Oakes, W. J. Law, R. Komuniecki. Cannabinoids stimulate the TRP channel-dependent release of both serotonin and dopamine to modulate behavior in C. elegans.. J. Neurosci., 2019. [DOI | PubMed]
- R. J. O’Brien, P. C. Wong. Amyloid precursor protein processing and alzheimer’s disease.. Annu. Rev. Neurosci., 2011. [DOI | PubMed]
- S. E. O’Sullivan. An update on PPAR activation by cannabinoids.. Br. J. Pharmacol., 2016. [DOI | PubMed]
- B. S. Padman, T. N. Nguyen, L. Uoselis, M. Skulsuppaisarn, L. K. Nguyen, M. Lazarou. LC3/GABARAPs drive ubiquitin-independent recruitment of optineurin and NDP52 to amplify mitophagy.. Nat. Commun., 2019. [DOI | PubMed]
- C. Pantoja-Ruiz, P. Restrepo-Jimenez, C. Castañeda-Cardona, A. Ferreirós, D. Rosselli. Cannabis and pain: a scoping review.. Braz J. Anesthesiol. Engl., 2022. [DOI | PubMed]
- H. Park, J. H. Kang, S. Lee. Autophagy in neurodegenerative diseases: a hunter for aggregates.. Int. J. Mol. Sci., 2020. [DOI | PubMed]
- C. A. Paronis, S. P. Nikas, V. G. Shukla, A. Makriyannis. Δ9-Tetrahydrocannabinol acts as a partial agonist/antagonist in mice.. Behav. Pharmacol., 2012. [DOI | PubMed]
- C. G. Parsons, W. Danysz, A. Dekundy, I. Pulte. Memantine and cholinesterase inhibitors: complementary mechanisms in the treatment of Alzheimer’s disease.. Neurotox. Res., 2013. [DOI | PubMed]
- N. Patil, V. Chandel, A. Rana, M. Jain, P. Kaushik. Investigation of Cannabis sativa phytochemicals as anti-alzheimer’s agents: an in silico study.. Plants, 2023. [DOI | PubMed]
- R. G. Pertwee. The diverse CB1 and CB2 receptor pharmacology of three plant cannabinoids: Δ9-tetrahydrocannabinol, cannabidiol and Δ9-tetrahydrocannabivarin.. Br. J. Pharmacol., 2008. [DOI | PubMed]
- M. R. Picciotto, M. J. Higley, Y. S. Mineur. Acetylcholine as a neuromodulator: cholinergic signaling shapes nervous system function and behavior.. Neuron, 2012. [DOI | PubMed]
- C. Piette, Y. Cui, N. Gervasi, L. Venance. Lights on endocannabinoid-mediated synaptic potentiation.. Front. Mol. Neurosci., 2020. [DOI | PubMed]
- T. Pongking, K. Intuyod, P. Thongpon, R. Thanan, C. Sitthirach, A. Chaidee. Cannabidiol suppresses proliferation and induces cell death, autophagy and senescence in human cholangiocarcinoma cells via the PI3K/AKT/mTOR pathway.. J. Tradit. Complement. Med., 2024. [DOI | PubMed]
- A. P. Porsteinsson, R. S. Isaacson, S. Knox, M. N. Sabbagh, I. Rubino. Diagnosis of early alzheimer’s disease: clinical practice in 2021.. J. Prev. Alzheimers Dis., 2021. [DOI | PubMed]
- T. Puopolo, C. Liu, H. Ma, N. P. Seeram. Inhibitory effects of cannabinoids on acetylcholinesterase and butyrylcholinesterase enzyme activities.. Med. Cannabis Cannabinoids., 2022. [DOI | PubMed]
- Y. Rabanal-Ruiz, E. G. Otten, V. I. Korolchuk. mTORC1 as the main gateway to autophagy,”. Editors. 2017. [DOI]
- B. M. Radu, A. M. M. Osculati, E. Suku, A. Banciu, G. Tsenov, F. Merigo. All muscarinic acetylcholine receptors (M1-M5) are expressed in murine brain microvascular endothelium.. Sci. Rep., 2017. [DOI | PubMed]
- M. A. Rahman, M. S. Rahman, M. D. H. Rahman, M. Rasheduzzaman, A. Mamun-Or-Rashid, M. J. Uddin. Modulatory effects of autophagy on APP processing as a potential treatment target for alzheimer’s disease.. Biomedicines, 2020. [DOI | PubMed]
- I. Raïch, J. Lillo, J. B. Rebassa, C. Griñán-Ferré, A. Bellver-Sanchis, I. Reyes-Resina. Cannabidiol as a multifaceted therapeutic agent: mitigating Alzheimer’s disease pathology and enhancing cognitive function.. Alzheimers Res. Ther., 2025. [DOI | PubMed]
- A. Raja, S. Ahmadi, F. de Costa, N. Li, K. Kerman. Attenuation of oxidative stress by cannabinoids and cannabis extracts in differentiated neuronal cells.. Pharmaceuticals, 2020. [DOI | PubMed]
- B. G. Ramírez, C. Blázquez, T. Gómez del Pulgar, M. Guzmán, M. L. de Ceballos. Prevention of Alzheimer’s disease pathology by cannabinoids: neuroprotection mediated by blockade of microglial activation.. J. Neurosci. Off. J. Soc. Neurosci., 2005. [DOI | PubMed]
- T. A. Reekie, M. P. Scott, M. Kassiou. The evolving science of phytocannabinoids.. Nat. Rev. Chem., 2017. [DOI]
- B. Reisberg, R. Doody, A. Stöffler, F. Schmitt, S. Ferris, H. J. Möbius. Memantine in moderate-to-severe Alzheimer’s disease.. N. Engl. J. Med., 2003. [DOI | PubMed]
- A. Ribeiro, V. Ferraz-de-Paula, M. L. Pinheiro, L. B. Vitoretti, D. P. Mariano-Souza, W. M. Quinteiro-Filho. Cannabidiol, a non-psychotropic plant-derived cannabinoid, decreases inflammation in a murine model of acute lung injury: role for the adenosine A(2A) receptor.. Eur. J. Pharmacol., 2012. [DOI | PubMed]
- R. Ricciarelli, E. Fedele. The amyloid Cascade hypothesis in alzheimer’s disease: it’s time to change our mind.. Curr. Neuropharmacol., 2017. [DOI | PubMed]
- G. Riedel, S. N. Davies. Cannabinoid function in learning, memory and plasticity.. Handb. Exp. Pharmacol. (, 2005. [DOI | PubMed]
- S. L. Roberds, J. Anderson, G. Basi, M. J. Bienkowski, D. G. Branstetter, K. S. Chen. BACE knockout mice are healthy despite lacking the primary β-secretase activity in brain: implications for Alzheimer’s disease therapeutics.. Hum. Mol. Genet., 2001. [DOI | PubMed]
- T. T. Rohn, E. Head. Caspases as therapeutic targets in Alzheimer’s disease: is it time to ‘cut’ to the chase?. Int. J. Clin. Exp. Pathol., 2009. [PubMed]
- S. Ruangritchankul, P. Chantharit, S. Srisuma, L. C. Gray. Adverse drug reactions of acetylcholinesterase inhibitors in older people living with dementia: a comprehensive literature review.. Ther. Clin. Risk Manag., 2021. [DOI | PubMed]
- E. B. Russo. Cannabis therapeutics and the future of neurology.. Front. Integr. Neurosci., 2018. [DOI | PubMed]
- A. C. Ruver-Martins, M. A. Bicca, F. S. de Araujo, B. H. L. de Noronha Sales Maia, F. A. Pamplona, E. G. da Silva. Cannabinoid extract in microdoses ameliorates mnemonic and nonmnemonic Alzheimer’s disease symptoms: a case report.. J. Med. Case Rep., 2022. [DOI | PubMed]
- M. Salazar, A. Carracedo, I. J. Salanueva, S. Hernández-Tiedra, M. Lorente, A. Egia. Cannabinoid action induces autophagy-mediated cell death through stimulation of ER stress in human glioma cells.. J. Clin. Invest, 2009. [DOI | PubMed]
- C. Sam, B. Bordoni. Physiology, acetylcholine,” in. StatPearls (, 2023
- N. Sánchez-Fernández, L. Gómez-Acero, A. Castañé, A. Adell, L. Campa, J. Bonaventura. A combination of Δ9-tetrahydrocannabinol and cannabidiol modulates glutamate dynamics in the hippocampus of an animal model of Alzheimer’s disease.. Neurotherapeutics, 2024. [DOI | PubMed]
- R. A. Saxton, D. M. Sabatini. mTOR signaling in growth, metabolism, and disease.. Cell, 2017. [DOI | PubMed]
- J. M. Schott, P. S. Aisen, J. L. Cummings, R. J. Howard, N. C. Fox. Unsuccessful trials of therapies for Alzheimer’s disease.. Lancet, 2019. [DOI | PubMed]
- D. Schubert, D. Kepchia, Z. Liang, R. Dargusch, J. Goldberg, P. Maher. Efficacy of cannabinoids in a pre-clinical drug-screening platform for alzheimer’s disease.. Mol. Neurobiol., 2019. [DOI | PubMed]
- C. Schuster, K. D. Gerold, K. Schober, L. Probst, K. Boerner, M. J. Kim. The autoimmunity-associated gene CLEC16A modulates thymic epithelial cell autophagy and alters T cell selection.. Immunity, 2015. [DOI | PubMed]
- K. Sharma. Cholinesterase inhibitors as Alzheimer’s therapeutics.. Mol. Med. Rep., 2019. [DOI | PubMed]
- S. Sinha, J. P. Anderson, R. Barbour, G. S. Basi, R. Caccavello, D. Davis. Purification and cloning of amyloid precursor protein beta-secretase from human brain.. Nature, 1999. [DOI | PubMed]
- J. H. Son, J. H. Shim, K. H. Kim, J. Y. Ha, J. Y. Han. Neuronal autophagy and neurodegenerative diseases.. Exp. Mol. Med., 2012. [DOI | PubMed]
- A. K. H. Stavoe, E. L. F. Holzbaur. Autophagy in neurons.. Annu. Rev. Cell Dev. Biol., 2019. [DOI | PubMed]
- S. N. Suresh, A. Chakravorty, M. Giridharan, L. Garimella, R. Manjithaya. Pharmacological tools to modulate autophagy in neurodegenerative diseases.. J. Mol. Biol., 2020. [DOI | PubMed]
- H. Suzuki, T. Osawa, Y. Fujioka, N. N. Noda. Structural biology of the core autophagy machinery.. Curr. Opin. Struct. Biol., 2017. [DOI | PubMed]
- D. Świetlik, A. Kusiak, A. Ossowska. Computational modeling of therapy with the NMDA antagonist in neurodegenerative disease: information theory in the mechanism of action of memantine.. Int. J. Environ. Res. Public Health, 2022. [DOI | PubMed]
- B. Tailor, A. Pilozzi, X. Huang. Contributing factors of neurodegeneration in alzheimer’s disease,” in. Alzheimer’s disease. Editor, 2019
- R. H. Takahashi, T. Nagao, G. K. Gouras. Plaque formation and the intraneuronal accumulation of β-amyloid in Alzheimer’s disease.. Pathol. Int., 2017. [DOI | PubMed]
- I. Tamargo-Gómez, G. Mariño. AMPK: regulation of metabolic dynamics in the context of autophagy.. Int. J. Mol. Sci., 2018. [DOI | PubMed]
- K. Tekirdag, A. M. Cuervo. Chaperone-mediated autophagy and endosomal microautophagy: joint by a chaperone.. J. Biol. Chem., 2018. [DOI | PubMed]
- Y. Tian, V. Bustos, M. Flajolet, P. Greengard. A small-molecule enhancer of autophagy decreases levels of Aβ and APP-CTF via Atg5-dependent autophagy pathway.. FASEB J., 2011. [DOI | PubMed]
- Y. Tizabi, B. Getachew, V. Tsytsarev, A. B. Csoka, R. L. Copeland, T. Heinbockel. Central nicotinic and muscarinic receptors in health and disease,” in. Acetylcholine – Recent advances and new perspectives (, 2023
- R. S. Toledano, I. Akirav. Cannabidiol prevents cognitive and social deficits in a male rat model of Alzheimer’s disease through CB1 activation and inflammation modulation.. Neuropsychopharmacology, 2025. [DOI | PubMed]
- R. M. Tolón, E. Núñez, M. R. Pazos, C. Benito, A. I. Castillo, J. A. Martínez-Orgado. The activation of cannabinoid CB2 receptors stimulates in situ and in vitro beta-amyloid removal by human macrophages.. Brain Res., 2009. [DOI | PubMed]
- C. G. Towers, A. Thorburn. Therapeutic targeting of autophagy.. EBioMedicine, 2016. [DOI | PubMed]
- A. Trang, P. B. Khandhar. Physiology, acetylcholinesterase. In: Statpearls., 2023
- V. Trezza, P. Campolongo. The endocannabinoid system as a possible target to treat both the cognitive and emotional features of post-traumatic stress disorder (PTSD).. Front. Behav. Neurosci., 2013. [DOI | PubMed]
- P. Tyrakis, C. Agridi, M. Kourti. A comprehensive exploration of the multifaceted neuroprotective role of cannabinoids in alzheimer’s disease across a decade of research.. Int. J. Mol. Sci., 2024. [DOI | PubMed]
- M. Valencia, S. R. Kim, Y. Jang, S. H. Lee. Neuronal autophagy: characteristic features and roles in neuronal pathophysiology.. Biomol. Ther., 2021. [DOI | PubMed]
- M. van der Stelt, C. Mazzola, G. Esposito, I. Matias, S. Petrosino, D. De Filippis. Endocannabinoids and beta-amyloid-induced neurotoxicity in vivo: effect of pharmacological elevation of endocannabinoid levels.. Cell Mol. Life Sci. CMLS, 2006. [DOI | PubMed]
- R. Vassar, B. D. Bennett, S. Babu-Khan, S. Kahn, E. A. Mendiaz, P. Denis. Beta-secretase cleavage of Alzheimer’s amyloid precursor protein by the transmembrane aspartic protease BACE.. Science., 1999. [DOI | PubMed]
- T. A. M. Vrechi, AHFF Leão, I. B. M. Morais, V. C. Abílio, A. W. Zuardi, J. E. C. Hallak. Cannabidiol induces autophagy via ERK1/2 activation in neural cells.. Sci. Rep., 2021. [DOI | PubMed]
- R. Wang, P. H. Reddy. Role of glutamate and NMDA receptors in alzheimer’s disease.. J. Alzheimers Dis. Jad., 2017. [DOI | PubMed]
- L. Wang, M. Chen, J. Yang, Z. Zhang. LC3 fluorescent puncta in autophagosomes or in protein aggregates can be distinguished by FRAP analysis in living cells.. Autophagy, 2013. [DOI | PubMed]
- S. Wang, H. Li, M. Yuan, H. Fan, Z. Cai. Role of AMPK in autophagy.. Front. Physiol., 2022. [DOI | PubMed]
- Z. Wang, P. Zheng, X. Chen, Y. Xie, K. Weston-Green, N. Solowij. Cannabidiol induces autophagy and improves neuronal health associated with SIRT1 mediated longevity.. GeroScience, 2022. [DOI | PubMed]
- T. M. Westlake, A. C. Howlett, T. I. Bonner, L. A. Matsuda, M. Herkenham. Cannabinoid receptor binding and messenger RNA expression in human brain: an in vitro receptor autoradiography and in situ hybridization histochemistry study of normal aged and Alzheimer’s brains.. Neuroscience, 1994. [DOI | PubMed]
- M. C. Wright, S. Potluri, X. Wang, E. Dentcheva, D. Gautam, A. Tessler. Distinct muscarinic acetylcholine receptor subtypes contribute to stability and growth, but not compensatory plasticity, of neuromuscular synapses.. J. Neurosci., 2009. [DOI | PubMed]
- M. M. Wu, X. Zhang, M. J. Asher, S. A. Thayer. Druggable targets of the endocannabinoid system: implications for the treatment of HIV-associated neurocognitive disorder.. Brain Res., 2019. [DOI | PubMed]
- X. Xiao, H. Liu, X. Liu, W. Zhang, S. Zhang, B. Jiao. APP, PSEN1, and PSEN2 variants in alzheimer’s disease: systematic Re-evaluation according to ACMG guidelines.. Front. Aging Neurosci., 2021. [DOI | PubMed]
- Y. Xiong, C. S. Lim. Understanding the modulatory effects of cannabidiol on alzheimer’s disease.. Brain Sci., 2021. [DOI | PubMed]
- C. Xu, D. J. Hermes, B. Nwanguma, I. R. Jacobs, K. Mackie, S. Mukhopadhyay. Endocannabinoids exert CB1 receptor-mediated neuroprotective effects in models of neuronal damage induced by HIV-1 Tat protein.. Mol. Cell Neurosci., 2017. [DOI | PubMed]
- Z. Yang, D. J. Klionsky. Mammalian autophagy: core molecular machinery and signaling regulation.. Curr. Opin. Cell Biol., 2010. [DOI | PubMed]
- B. A. Yankner, L. R. Dawes, S. Fisher, L. Villa-Komaroff, M. L. Oster-Granite, R. L. Neve. Neurotoxicity of a fragment of the amyloid precursor associated with Alzheimer’s disease.. Science., 1989. [DOI | PubMed]
- K. G. Yiannopoulou, S. G. Papageorgiou. Current and future treatments in alzheimer disease: an update.. J. Cent. Nerv. Syst. Dis., 2020. [DOI | PubMed]
- A. Yoder. Chapter 7 – the endocannabinoid system and chronic disease: opportunity for innovative therapies,” in. Physical activity and the aging brain. Editor, 2017
- A. P. Young, E. M. Denovan-Wright. The dynamic role of microglia and the endocannabinoid system in neuroinflammation.. Front. Pharmacol., 2022. [DOI | PubMed]
- J. J. Yu, E. A. Goncharova. mTOR signaling network in cell biology and human disease.. Int. J. Mol. Sci., 2022. [DOI | PubMed]
- W. H. Yu, A. Kumar, C. Peterhoff, L. Shapiro Kulnane, Y. Uchiyama, B. T. Lamb. Autophagic vacuoles are enriched in amyloid precursor protein-secretase activities: implications for beta-amyloid peptide over-production and localization in Alzheimer’s disease.. Int. J. Biochem. Cell Biol., 2004. [DOI | PubMed]
- H. Zempel, E. Thies, E. Mandelkow, E. M. Mandelkow. Abeta oligomers cause localized Ca(2+) elevation, missorting of endogenous tau into dendrites, tau phosphorylation, and destruction of microtubules and spines.. J. Neurosci. Off. J. Soc. Neurosci., 2010. [DOI | PubMed]
- C. Zhang, A. M. Cuervo. Restoration of chaperone-mediated autophagy in aging liver improves cellular maintenance and hepatic function.. Nat. Med., 2008. [DOI | PubMed]
- W. Zhang, C. Xu, J. Sun, H. M. Shen, J. Wang, C. Yang. Impairment of the autophagy–lysosomal pathway in Alzheimer’s diseases: pathogenic mechanisms and therapeutic potential.. Acta Pharm. Sin. B, 2022. [DOI | PubMed]
- J. Zhao, X. Liu, W. Xia, Y. Zhang, C. Wang. Targeting amyloidogenic processing of APP in alzheimer’s disease.. Front. Mol. Neurosci., 2020. [DOI | PubMed]
- R. Zhou, A. S. Yazdi, P. Menu, J. Tschopp. A role for mitochondria in NLRP3 inflammasome activation.. Nature, 2011. [DOI | PubMed]
- H. Zinecker, A. K. Simon. Autophagy takes it all – autophagy inducers target immune aging.. Dis. Model Mech., 2022. [DOI | PubMed]
- S. Zou, U. Kumar. Cannabinoid receptors and the endocannabinoid system: signaling and function in the central nervous system.. Int. J. Mol. Sci., 2018. [DOI | PubMed]
- Z. Zou, T. Tao, H. Li, X. Zhu. mTOR signaling pathway and mTOR inhibitors in cancer: progress and challenges.. Cell Biosci., 2020. [DOI | PubMed]
- A. W. Zuardi. History of cannabis as a medicine: a review.. Rev. Bras. Psiquiatr. Sao Paulo Braz, 1999. [DOI | PubMed]
- A. W. Zuardi, J. E. C. Hallak, J. A. S. Crippa. Interaction between cannabidiol (CBD) and Δ(9)-tetrahydrocannabinol (THC): influence of administration interval and dose ratio between the cannabinoids.. Psychopharmacol. Berl., 2012. [DOI | PubMed]