
Medicinal cannabis plant extract (NTI164) modifies epigenetic, ribosomal, and immune pathways in paediatric acute-onset neuropsychiatric syndrome
Abstract
Paediatric acute-onset neuropsychiatric syndrome (PANS) is a syndrome of infection-provoked abrupt-onset obsessive-compulsive disorder (OCD) or eating restriction. Based on the hypothesis that PANS is an epigenetic disorder of immune and brain function, a full-spectrum medicinal cannabinoid-rich low-THC cannabis (NTI164) was selected for its known epigenetic and immunomodulatory properties. This open-label trial of 14 children with chronic-relapsing PANS (mean age 12·1 years; range 4–17; 71 % male) investigated the safety and efficacy of 20 mg/kg/day NTI164 over 12 weeks. Clinical outcomes were assessed using gold standard tools. To define the biological effects of NTI164, blood samples were collected pre- and post-treatment for bulk and single-cell transcriptomics, proteomics, phosphoproteomics, and DNA methylation. NTI164 was well-tolerated, and 12 weeks of treatment decreased the mean Clinical Global Impression-Severity (CGI-S) score from 4·8 to 3·3 (p = 0·002). Significant improvements were observed in emotional regulation (RCADS-P, p < 0·0001), obsessive-compulsive disorder (CYBOCS-II, p = 0·0001), tics (YGTSS, p < 0·0001), attention-deficit hyperactivity disorder (Conner’s, p = 0·028), and overall quality of life (EQ-5D-Y, p = 0·011). At baseline, the multi-omic approach revealed that leucocytes from patients with PANS had dysregulated epigenetic (chromatin structure, DNA methylation, histone modifications, transcription factors), ribosomal, mRNA processing, immune, and signalling pathways. These pathways were significantly modulated by NTI164 treatment. NTI164 shows promise as a disease-modifying therapeutic for PANS. Multi-omics reveal broad epigenetic and immune dysregulation in patients, which was modified by NTI164, presenting epigenetic machinery as a therapeutic target in PANS.
Article type: Research Article
Keywords: Cannabis, Epigenetics, Immune, Multi-omics, PANS, Ribosome
Affiliations: Kids Neuroscience Centre, The Children’s Hospital at Westmead, Faculty of Medicine and Health, University of Sydney, NSW, Australia; Fenix Innovation Group Pty Ltd, Mount Waverley, VIC 3149, Australia; Khoo Teck Puat-National University Children’s Medical Institute, National University Health System, Singapore, Singapore; Department of Paediatrics, Yong Loo Lin School of Medicine, National University of Singapore, Singapore; Bioinformatics Group, Children’s Medical Research Institute, University of Sydney, Westmead, NSW, Australia; Faculty of Medicine and Health, Sydney Medical School, University of Sydney, Camperdown, NSW, Australia; Westmead Research Hub, Westmead Institute for Medical Research, Westmead, NSW, Australia; Australian Genome Research Facility Ltd, Melbourne, VIC, Australia; Australian Genome Research Facility Ltd, Westmead, NSW, Australia; Westmead Institute for Medical Research, Sydney, Australia; Biomedical Proteomics, Children’s Medical Research Institute, The University of Sydney, NSW, Australia; The Children’s Hospital at Westmead Clinical School, Faculty of Medicine and Health, University of Sydney, Sydney, NSW, Australia; Department of Neurology, Monash Children’s Hospital, Monash Health, Clayton, VIC, Australia; The University of Sydney, School of Medical Sciences and Discipline of Child and Adolescent Health, Faculty of Medicine and Health, Sydney, NSW, Australia
License: © 2026 The Authors CC BY 4.0 This is an open access article under the CC BY license (http://creativecommons.org/licenses/by/4.0/).
Article links: DOI: 10.1016/j.neurot.2025.e00828 | PubMed: 41513541 | PMC: PMC12976522
Relevance: Relevant: mentioned in keywords or abstract
Full text: PDF (1.8 MB)
Introduction
Paediatric acute-onset neuropsychiatric syndrome (PANS) is a debilitating condition, characterised by abrupt-onset obsessive-compulsive disorder (OCD) or severely restrictive food intake with co-existing symptoms, including anxiety, depression, irritability, behavioural regression, cognitive deterioration, sensory or motor abnormalities, and somatic symptoms [ref. 1]. PANS often has a relapsing-remitting clinical course where periods of stability can be followed by relapses [ref. 2], triggered by infections or other stressors.
A recent American Academy of Paediatrics statement recognises PANS as a clinical syndrome, however it is unclear whether PANS is a distinct neuroimmune entity or part of the neurodevelopmental continuum [ref. 3,ref. 4]. OCD affects 2 % of children and commonly co-exists with other neurodevelopmental disorders (NDDs), such as autism spectrum disorder, attention-deficit hyperactivity disorder (ADHD), and Tourette syndrome [ref. 5]. Rare, pathogenic DNA variants are found in only a small minority of patients with NDDs [ref. 6]. Instead, a combination of genetic and environmental factors is considered to contribute to the development of NDDs, through epigenetic, gene regulatory, and immune processes [ref. 7,ref. 8]. We propose that PANS is a clinical phenotype driven by gene-environment interactions, involving epigenetic dysregulation that impacts both the immune system and brain [ref. 9,ref. 10]. There are currently no approved treatments for PANS, and patients often have refractory symptoms, despite the use of conventional psychotherapy or psychiatric medications.
Cannabis sativa L. has long been used in medicine, and increasingly proposed as a treatment of psychiatric disorders and NDDs [ref. 11] The plant secretes a resin which contains a mix of cannabinoids with two principal components, Δ9-tetrahydrocannabinol (THC) and cannabidiol (CBD). Extensive research centres around CBD, which has generated much interest due to a lack of psychoactive activity (unlike THC) and excellent tolerability in humans, as well as anti-inflammatory capabilities [ref. 12]. While THC can elicit undesired psychoactive effects, emerging evidence suggests cannabis extracts with all components of the plant are more potent immunomodulators than CBD alone (or other isolated compounds), termed the “entourage effect” [ref. 13]. Other studies support the observation that botanical drugs are more efficacious than their isolated components [ref. 14].
Currently, the strongest evidence for use of CBD and other cannabinoids is in refractory epilepsy [ref. 15]. Recent pre-clinical studies propose cannabinoids act on various brain mechanisms and molecular targets, offering neuroprotective, anti-inflammatory, anxiolytic, and anti-psychiatric effects [ref. [16], ref. [17], ref. [18]]. These findings suggest cannabinoids may benefit neuropsychiatric disorders by modulating multiple signalling pathways, and randomised controlled trials are now emerging to clarify therapeutic indications [ref. [19], ref. [20], ref. [21]].
In this open-label study (FENPANS1, ClinicalTrials.gov ID: NCT06621888), the safety and efficacy of NTI164, a full-spectrum medicinal cannabis plant derived from proprietary strains of Cannabis sativa and containing cannabidiolic acid (CBDA), CBD, cannabigerolic acid (CBGA), cannabidivarin (CBDV), and low THC (<0·3 %) were investigated in a cohort of children diagnosed with PANS. NTI164 was administered orally twice daily at a dose of 20 mg/kg/day for 12 weeks. This open-label study measured changes in PANS clinical symptoms and explored the biological effects of NTI164 on peripheral immune cells through cytokine assays, transcriptomic, proteomic, phosphoproteomic, and DNA methylation analyses, at baseline and after treatment with NTI164 in children with PANS.
Methods
Ethics approval
Ethics approval for this study was granted by the Sydney Children’s Hospitals Network (SCHN) Human Research Ethics Committee (HREC reference: 2022/ETH02308) and Monash Health (HREC reference: RES-23-0000-333X).
Participant selection
Children were recruited from specialist neurology clinics at the Children’s Hospital at Westmead (CHW, clinic run by RD) or Monash Children’s Hospital (MCH, clinic run by MF and MK). All children were diagnosed with PANS according to the criteria from Swedo et al. [ref. 1], and Chang et al. [ref. 22], and also met eligibility criteria for the current study (for full inclusion/exclusion criteria, see Supplementary material, Appendix 1). All participants and their parent/guardian provided written informed consent for participation in this study. Age- and sex-matched healthy control children (children of hospital staff members) were recruited for biological investigations. The inclusion criteria for controls were the absence of neurodevelopmental disorders, autoimmune conditions, moderate-severe allergies, and no recent infections (within 2 weeks of blood sample collection).
Structured clinical assessment on REDCap and clinician-directed interview (questionnaires)
A structured clinical interview was conducted between parents of participants and trial staff to gather information regarding family background and familial medical history and symptoms at the time of recruitment. This allowed family medical history and information relating to patient symptoms to be collected in a standardised manner across participants and sites.
Study design
This study was split into four stages: up-titration, treatment, down-titration, and extension. In the up-titration phase, patients received a starting dose of NTI164 of 5 mg/kg/day. This was increased by 5 mg/kg weekly for four weeks, until the target dose of 20 mg/kg/day (or maximum tolerated dose as determined by the treating physician) was reached by Week 4 of the study.
In the treatment phase, children received NTI164 at 20 mg/kg/day (or their maximum tolerated dose) for eight weeks. Blood samples were collected at Week 12 (i.e. end of the treatment phase, primary endpoint), as well as questionnaires completed. At Week 12, children wishing to stop NTI164 were down-titrated by 5 mg/kg/week from 20 mg/kg/day (i.e. reversal of up-titration), and Week 16 was the end of the study for these children. At Week 12, children wishing to remain on NTI164 and enter the extension phase were able to do so.
Questionnaires
Questionnaires used in this study are gold-standard tools, see Supplementary Table 3 for more detail on individual questionnaires. Questionnaires were administered by trained personnel at baseline (i.e. prior to NTI164) and at Week 12 (i.e. completion of treatment phase). Parent/caregiver-rated questionnaires were completed at the same time points in the presence of study staff.
Sample collection
After written consent, venous blood was collected prior to starting NTI164 (baseline) as well as after 12 weeks of receiving the drug. See Supplementary Table 1 for detailed information about parameters investigated. For the plasma neuroinflammation panel, venous blood was collected into lithium heparin tubes and centrifuged at 1,300xG for 10 min at room temperature. Separated plasma was divided into 200 μL aliquots and stored at −80 °C until analysis. For bulk-RNA Sequencing, venous blood was collected directly into PAXGene™ Blood RNA tubes and stored at −80 °C until extraction and sequencing by the Australian Genome Research Facility Ltd (AGRF Ltd). For single-cell RNA sequencing, venous blood was collected into acid-citrate-dextrose (ACD) tubes, and red blood cells (RBCs) were removed following the EasySep™ RBC Depletion protocol from STEMCELL Technologies (catalogue #18170) using the EasySep™ Magnet (catalogue #18000, Stemcell Technologies). The resulting cell suspension was then loaded into the HIVE™ collectors, following the manufacturer’s protocol (Honeycomb Biotechnologies, Inc., USA) and stored at −80 °C until library preparation and sequencing by AGRF Ltd. Samples were always loaded into a HIVE™ collector within 1 h of collection to reduce neutrophil activation. For methylation, venous blood was collected directly into EDTA K2 tubes and stored at −80 °C until analysis by AGRF Ltd. For proteomics and phosphoproteomics, a minimum volume of 4 mL venous blood was collected into ACD tubes and diluted 1:1 with Dulbecco’s phosphate buffer saline (DPBS) -Ca2 ± Mg2+. Samples were then layered over Ficoll in SepMate™-15 tubes (STEMCELL Technologies, catalogue #85415) and centrifuged at 1,200xG for 15 min at room temperature (RT). Supernatant was discarded and the peripheral blood mononuclear cell (PBMC) layer was transferred via pipette into a 50 mL tube. Tubes were filled to 50 mL with DPBS and centrifuged at 1,000xG for 6 min at RT, twice.
Following the second spin, cells were resuspended in 200 μL lysis buffer with detergent (50 mM Tris, 0.8 % v/v Triton X-100, Complete EDTA-free Protease Inhibitor Cocktail (Merck), PhosSTOP (Merck)) + 2uL of 200 mM phenylmethylsulfonyl fluoride (PMSF) in ethanol, transferred to a 1·5 mL tube, heated at 85 °C for 10 min, and then stored at −80 °C until analysis.
Plasma neuroinflammation panel
A multiplex assay was performed on thawed plasma from eight children with PANS (mean age 11·4 years, 37·5 % females) and eight age- and sex-matched controls (mean age 11·8 years, 37·5 % females). The LEGENDplex™ Human Neuroinflammation Panel 1 (13-plex) assay was performed according to the manufacturer’s protocol by the Westmead Cytometry Core Facility at the Westmead Research Hub (Westmead, Australia).
Bulk blood RNA sequencing (library preparation and sequencing)
Bulk RNA sequencing was performed on whole blood from eight children with PANS (mean age 11·4 years, 37·5 % females) and eight age- and sex-matched controls (mean age 11·8 years, 37·5 % females). This workflow included RNA extraction, depletion of ribosomal RNA via hybrid capture (Illumina Ribo-Zero), and Illumina TruSeq Stranded Total RNA Library Preparation (input 200-1,000 ng of Total RNA). The stranded RNA samples were sequenced on the Illumina NovaSeq X sequencing platform (2 × 150 base pairs) for a depth of 50 million paired end reads. The cleaned sequence reads were aligned against the Homo sapiens genome (Build version hg38), and the STAR aligner (v2·5·3a) was used to map unique reads to the genomic sequences [ref. 23].
Single-cell RNA sequencing (HIVE, library preparation, and sequencing)
Single-cell RNA sequencing was performed on samples from four patients at baseline and after 12 weeks of NTI164 treatment (mean age 15·2 years, 50 % females), as well as four age- and sex-matched controls (mean age 14·5 years, 50 % females). Following a standard protocol (Honeycomb Biotechnologies, Inc., USA), cell-loaded HIVE™ devices were sealed with a semi-permeable membrane, allowing for a strong lysis solution and a hybridisation solution. After collection, beads with captured transcripts were extracted from the HIVE™ by centrifugation and transferred to a 96-well plate (AGRF Ltd., Westmead). The size profiles of the final libraries were determined on a TapeStation platform using a D5000 ScreenTape System (Agilent Technologies, Santa Clara, CA, USA), and the concentration of final pooled libraries was determined by qPCR. HIVE™ scRNAseq libraries were sequenced using specific primers contained in the kit on an Illumina® NovaSeq X sequencer (AGRF Ltd., Melbourne).
Proteomics
Proteomics analysis was performed in six patients (PANS-pre and PANS-post, mean age 12 years, 33 % female), as well as six age- and sex-matched control children (mean age 12·3 years, 33 % female). The samples were lysed, digested, and tagged using the TMTpro 18-plex system (n = 6 for each of three groups). Hydrophilic ion liquid chromatography fractionation was performed on a Vanquish Neo HPLC system with a 250 mm long and 1 mm inside diameter TSKgel Amide-80 column (Tosoh Biosciences, Inc., OH, USA). Fractions were collected into a 96-well plate using an FC204 fraction collector (Gilson) at 1 min intervals, monitored by absorbance of UV at 214 nm. Fractions were analysed by LC-MS/MS, performed using a Dionex UltiMate 3000 RSLC nano system and Q Exactive Plus hybrid quadrupole-orbitrap mass spectrometer (Thermo Fisher Scientific). An in-house 300 mm long 0·075 mm inside diameter column packed with ReproSil Pur C18 AQ 1·9 μm resin (Dr Maisch, Germany) was used. The instrument settings were as described previously [ref. 24].
Phosphoproteomics
Phosphoproteomics analysis was performed on the same samples as underwent proteomics analysis, and so all PBMC preparation was identical. Phosphopeptides were enriched and fractionated prior to LC-MS/MS analysis as described previously [ref. 25].
Searching of LC-MS/MS data
The raw LC-MS/MS data was processed with MaxQuant v1·6·7·0 using the following settings: variable modifications were oxidation (M), acetyl (protein N-terminus), deamidation (NQ) and phospho (STY); carbamidomethyl (C) was a fixed modification; digestion was set to trypsin/P with a maximum of three missed cleavages; the TMTpro correction factors were entered for lots XC344112 and XK350589; minimum reporter peptide ion faction was 0·6; the Homo sapiens reference proteome with canonical and isoform sequences downloaded March 4, 2024 with 82,485 entries and 20,597 genes; the inbuilt contaminants fasta file was also used; minimum peptide length was six and maximum peptide mass was 6,000 Da; second peptides search and dependent peptides searches were enabled; peptide spectrum matching and protein false discovery rates were set at 1 %. All other settings were default.
DNA methylation
Methylation analysis was performed on whole blood from eight children with PANS (mean age 11·4 years, 37·5 % females) and eight age- and sex-matched controls (mean age 11·8 years, 37·5 % females). Genome-wide DNA methylation was assessed using the Illumina MethylationEPIC v2·0 BeadChip (Illumina, Inc., San Diego, CA, USA), which interrogates >936,000 CpG sites. The BeadChips were scanned using an Illumina iScan. All analysis was undertaken using the R statistical environment [ref. 26]. Quality control and probe summaries were assessed using the lumi Bioconductor package [ref. 27]. All samples were retained following quality control. Differential methylation was assessed using the limma package [ref. 28].
Bioinformatic analysis
Omics bioinformatic analysis
Omics data were analysed in the R statistical environment [ref. 26] with tidyverse [ref. 29]. For bulk RNA sequencing and proteomics, filtering and normalisation steps were first performed. Subsequently, normalisation with removal of unwanted variation, via the remove unwanted variation (RUV) R package was performed [ref. 30]. In the bulk RNA sequencing (k = 13), proteomics (k = 4), and phosphoproteomics (k = 4) (factors of unwanted variation) were used respectively to remove genes that had minimal differential expression, compared to negative control genes. For linear modelling, the limma R package was used and the p-values were calculated using the empirical Bayes method ‘eBayes’ function [ref. 31]. The false discovery rate (FDR) correction was applied to the p-values by calculating the adjusted p-values. Significant differentially expressed genes/proteins were defined as those with adjusted p-values/false discovery rate (FDR) < 0·05.
For single cell transcriptomics, the data was analysed using the Seurat package [ref. 32]. Cells with a high mitochondrial transcript ratio (>0·15) were excluded. Normalisation was performed using SCTransform in Seurat and immune cell types were assigned with scType and scPred [ref. 33]. Merged data were then split by cell type and separately normalised, scaled, integrated between patient using harmony [ref. 34], then UMAP (uniform manifold approximation and projection) projections were made using the first 30 dimensions. Differentially expressed genes with significant FDR values < 0·05 were identified using FindMarkers.
DNA methylation profiling was performed using the Illumina Infinium MethylationEPIC v2·0 BeadChip (Illumina, Inc., San Diego, CA, USA). Raw IDAT files were imported and preprocessed using the minfi package [ref. 35] in R, following the Bioconductor methylation array analysis workflow [ref. 36]. Low-quality samples and probes were excluded based on detection p-values (>0·01) and known single nucleotide polymorphisms (SNPs). Probes on sex chromosomes and non-CpG probes were also removed. Data were normalised using preprocessQuantile.
Methylation levels were summarised as beta and M-values, with M-values used for statistical testing. To account for unwanted technical variation, RUV-III normalisation was applied to the M-values using a control set of probes and setting unwanted factors K = 6 [ref. 37]. Differentially methylated positions (DMPs) were identified using the limma package [ref. 31], applying empirical Bayes moderation and adjusting for multiple testing with the Benjamini-Hochberg method (FDR <0·05). Differentially methylated regions (DMRs) were detected using the DMRcate package [ref. 38], which applies kernel smoothing to identify spatially correlated methylation changes across genomic regions.
Pathway enrichment analysis
Gene set enrichment analysis (GSEA) for bulk RNA seq, scRNAseq, and proteomics
The genes and proteins were ranked based on their sign(logFC) x log10Pvalue scores [ref. 39,ref. 40]. Enriched gene sets were identified based on a running sum statistic (adjusted p values) and statistical significance based on the false discovery rate (FDR). Significant GSEA Gene Ontology (GO) pathways (FDR <0·05) were further simplified use the simplify function in clusterProfiler, and ranked by p.adjust [ref. 41].
Over representation enrichment (ORA) analysis for phosphoproteomics and methylation
We took an approach that deliberately reduced the significant pathways identified by limiting the background to only proteins we could detect by mass spectrometry, and using the ranked list method within gProfiler, which weights enrichment towards highly ranked genes. We separated up- and down-regulated phosphorylation, assigned the maximum positive and negative quantitative values for phospho-regulation to each protein, and ranked the proteins using both the quantitative value and significance of maximal change.
Bar and dot plots of GSEA results were plotted using ggplot2 package, and heatmaps of GSEA results were made using the pheatmap package. To further evaluate themes within pathways, gene subclusters were created based on most significant Reactome or molecular function pathways per gene set, and connectivity network (CNET) plots were created using enrichplot.
For methylation, probe annotations were retrieved from the IlluminaHumanMethylation450kanno·ilmn12·hg19 annotation package. Gene ontology enrichment for DMP or DMR-associated genes was conducted using the clusterProfiler package, employing enrichGO and enrichKEGG functions for Gene Ontology and KEGG pathway enrichment, respectively [ref. 42]. Reactome pathway analysis was also conducted using enrichPathway from the ReactomePA package [ref. 43]. The same approach was used to evaluate themes within pathways using enrichplot.
Role of funders
This study was monetarily supported by Fenix Innovation Group Pty Ltd and Neurotech International Ltd (Sponsors). The Sponsors had no role in patient recruitment, data collection, analysis, or presentation.
Results
Clinical characteristics of children with PANS
Family history
A total of 14 patients fulfilling PANS criteria [ref. 1,ref. 22] (full inclusion/exclusion criteria available in Appendix 1) from New South Wales and Victoria, Australia were recruited to the study from two tertiary neurology clinics. The mean age was 12·1 (range 4–17) years ((4 females, 29 %; 10 males, 71 %), Table 1), 64·3 % (n = 9) of the probands had a first-degree relative with an autoimmune condition, and 50 % (n = 7) of probands had a first-degree relative with a neurodevelopmental or neuropsychiatric disorder (Table 1).
Table 1: Clinical characteristics of PANS patients enrolled in the FENPANS1 study.
| Age at recruitment (yr), sex | 1st degree FH NDD | 1st degree FH autoimmunity | Age PANS onset (yr), trigger | PANS phenotype | NDDs at study commencement | Conventional Tx at study commencement | Previous rescue Tx | RCADS-P baseline score | CY-BOCS-II baseline score |
|---|---|---|---|---|---|---|---|---|---|
| 15, M | ADHD (sib, mat) | Grave’s disease (mat.) | 4.5, Strep throat | OCD, agitation | ASD, ADHD, OCD, Tourette | Methylphenidate, aripiprazole, risperidone, fluoxetine | Azithromycin | 84 | 36 |
| 11, M | – | Crohn’s disease (pat.); MS, Grave’s disease (mat.) | 4, unknown trigger | OCD, cognitive decline | OCD | Sertraline, aripiprazole | Azithromycin, amoxycillin-clavulanic acid | 75 | 33 |
| 7, M | ADHD (siblings) | T1D (pat.); post-partum thyroiditis (mat.) | 5, throat infection NOS | Behavioural regression, eating restriction, separation anxiety, rage | ASD, OCD, ADHD | Guanfacine, methylphenidate | Erythromycin | 87 | 28 |
| 7, F | – | – | 2, Staph-scalded skin syndrome | Separation anxiety, OCD, rage | ADHD, OCD, tics | Aripiprazole, methylphenidate, fluoxetine | Azithromycin, amoxycillin-clavulanic acid | 81 | 31 |
| 16, F | Depression (mat., pat.) | Thyroid NOS (mat.) | 3, URTI, immune response post-vaccine | OCD, anxiety, inattention | Tourette, OCD, | Aripiprazole | Azithromycin, amoxycillin-clavulanic acid | 91 | 40 |
| 17, M | – | Thyroid NOS (mat.) | 12, unknown trigger | OCD | OCD | Fluoxetine, clonidine | Azithromycin, amoxycillin-clavulanic acid | 95 | 37 |
| 14, F | ADHD, PTSD, eating disorder (mat.) | Raynaud’s disease (mat.) | 11, throat infection NOS | Insomnia, hallucinations, cognitive decline, OCD | OCD | Fluoxetine, risperidone, aripiprazole, quetiapine | Cephalexin | 79 | 29 |
| 9, M | Anxiety, depression (mat.) | Hashimoto’s thyroiditis (mat.) | 3.5, multiple infections NOS, pneumonia | Restricted eating, autistic regression, stutter, depression | ADHD, ASD, tics, OCD | Fluoxetine, quetiapine, dexamphetamine | Amoxycillin-clavulanic acid | 77 | 27 |
| 9, M | Depression, anxiety, PTSD (mat.) | Hashimoto’s thyroiditis, coeliac disease, psoriasis (mat.) | 1.5, croup | Autistic regression, OCD, irritability, sensory issues | ASD, OCD, ID | Methylphenidate, risperidone, guanfacine | Amoxycillin-clavulanic acid | anot performed | anot performed |
| 13, M | – | – | 2.5, bronchitis NOS | Restricted eating, separation anxiety | ASD, OCD | Fluoxetine | Amoxycillin-clavulanic acid | 91 | 40 |
| 4, M | – | – | 1, URTI, immune response post-vaccine | Autistic regression, catatonic state, OCD, cognitive decline | ASD, OCD | – | Azithromycin, amoxycillin-clavulanic acid | anot performed | anot performed |
| 16, M | Depression, PTSD, ADHD, eating disorder (mat.) | – | 8, throat infection NOS | OCD, tics, anxiety, language regression, agitation | ADHD, OCD, ASD | Quetiapine, olanzapine, risperidone, methylphenidate, sertraline | Azithromycin, IVIG | 86 | 10 |
| 13, M | – | Primary cholangitis, SLE, psoriasis, coeliac disease (mat.); ulcerative colitis (pat.) | 3, URTI, multiple infections NOS | Language regression, OCD, cognitive decline, agitation | ASD, OCD, tics, ADHD | Olanzapine, methylphenidate | IVIG, steroids, azithromycin | 129 | 41 |
| 13, F | – | – | 11, COVID-19 | OCD, eating restriction, oppositional behaviour, anxiety, inattention | OCD | Fluoxetine | Amoxycillin-clavulanic acid, azithromycin, steroids | 90 | 45 |
ADHD = attention-deficit hyperactivity disorder, ASD = autism spectrum disorder, FH = family history, GDD = global developmental delay, ID = intellectual disability, MS = multiple sclerosis, NDD = neurodevelopmental disorder, NOS = not otherwise specified, OCD = obsessive-compulsive disorder, PTSD = post-traumatic stress disorder, SIB = self-injurious behaviours, T1D = type 1 diabetes, URTI = upper respiratory tract infection.
PANS symptoms
The mean age of PANS onset was 5·2 years (range 1·5–12 years) and was typically triggered by infections (n = 12, 85·7 %). In our cohort, all patients demonstrated a relapsing-remitting phenotype and had an average of five PANS-related flares each year (range 2–8 flares a year), lasting 3–4 days to 6 months in duration, which were mainly infection-provoked. The PANS symptoms in our cohort were OCD (n = 14, 100 %), anxiety (n = 8, 57·1 %), regression/cognitive decline (n = 8, 57·1 %), inattention or agitation (n = 6, 42·9 %), tics/Tourette syndrome (n = 3, 21·4 %), rage or oppositional behaviours (n = 3, 21·4 %), and severe food restriction (n = 3, 21·4 %). In our cohort, other NDD and neuropsychiatric diagnoses diagnosed, in addition to their PANS diagnosis, included autism spectrum disorder (n = 8, 53·3 %) and ADHD (n = 6, 42·9 %). The average duration between PANS onset and enrolment into the current study was 6·7 (range 2–13) years.
Severity of PANS symptoms
At baseline, the mean Clinical Global Impression – Severity (CGI-S) score was 4·8 out of 7 (markedly ill; score range 4–6, higher score indicates severe disease), and baseline Revised Children’s Anxiety and Depression Scale – Parent-rated (RCADS-P) and Children’s Yale-Brown Obsessive-compulsive Scale (CY-BOCS) scores were 101·9 (score range 92–129) and 33·1 (score range 24–48) respectively, indicating severe emotional disorders and OCD.
Neuropsychiatric and PANS treatments
All patients had previously received or were receiving ongoing psychological support. Children were taking an average of 2·4 medications (range 1–5) at the time of trial enrolment, including selective serotonin reuptake inhibitors (SSRIs; n = 9, 64·3 %), antipsychotics (n = 9, 64·3 %), stimulants (n = 7, 50 %), and α-agonists (n = 3, 21·4 %). All patients had previously received antibiotics as a rescue treatment for acute episodes. Additionally, 14·3 % (n = 2) had been treated with steroids or intravenous immunoglobulin (IVIg) during prior flares. As part of inclusion criteria to the current study, IVIg, steroids, and antibiotic use was not permitted for at least 12 weeks before entering the study or throughout the study period.
Safety and tolerability of NTI164 in PANS patients
Oral NTI164 was well-tolerated in patients at 20 mg/kg/day, divided into 2 doses, with no significant abnormalities observed in laboratory results (full blood examination, including kidney and liver function tests, Supplementary Table 1) after 12 weeks of treatment. Adverse events (AEs) determined to be related to the Investigational Product (Supplementary Table 2) were experienced by 33 % of patients, with these being symptoms of gastrointestinal symptoms (i.e. nausea, diarrhoea), lethargy, or changes to sleep behaviour (i.e. taking longer to fall asleep). One patient required dose-adjustment due to diarrhoea (reduced to 15 mg/kg/day) and one patient due to lethargy (reduced to 17·5 mg/kg/day). All other AEs were self-resolving and did not impact the patients’ functioning. No patients discontinued NTI164 due to AEs during the study period.
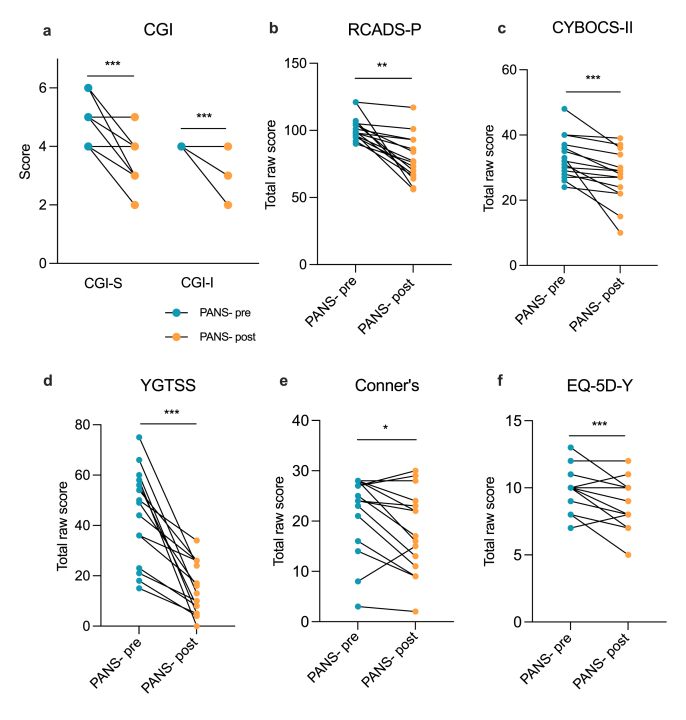
NTI64 improved clinical symptoms associated with PANS
Several gold-standard assessments of behaviour and tics (Supplementary Table 3) were implemented in the FENPANS1 study and were assessed by trained personnel at baseline and following 12 weeks of NTI164 oral administration (Fig. 1, Supplementary Table 4). At baseline, the mean CGI-S score was 4·8 (range 4–6) which was significantly reduced (i.e. improved) to 3·3 (range 2–5) after 12 weeks of NTI164 administration (Fig. 1A, p = 0·002). Similarly, NTI164 administration significantly improved emotional regulation (RCADS-P, Fig. 1B, p < 0·0001), OCD (CYBOCS-II, Fig. 1C, p = 0·0001), tics (YGTSS, Fig. 1D, p < 0·0001), ADHD (Conner’s, Fig. 1E, p = 0·028), and overall quality of life (EQ-5D-Y, Fig. 1F, p = 0·011). Notably, the RCADS-P showed improvements in all subdomains, including social phobia (p < 0·0001), panic disorder (p = 0·018), major depression (p = 0·0098), separation anxiety (p < 0·0001), generalised anxiety (p < 0·0001), and obsessive-compulsive symptoms (p = 0·001) (subdomains for the relevant surveys can be found in Supplementary Fig. 1 and Supplementary Table 4). Following 12 weeks of NTI164 treatment, 13 patients/caregivers (85·7 %) elected to continue receiving NTI164 into an Extension phase.
PANS children have dysregulated immune cell transcriptomic signatures, which were modified by NTI164
Cytokine assay
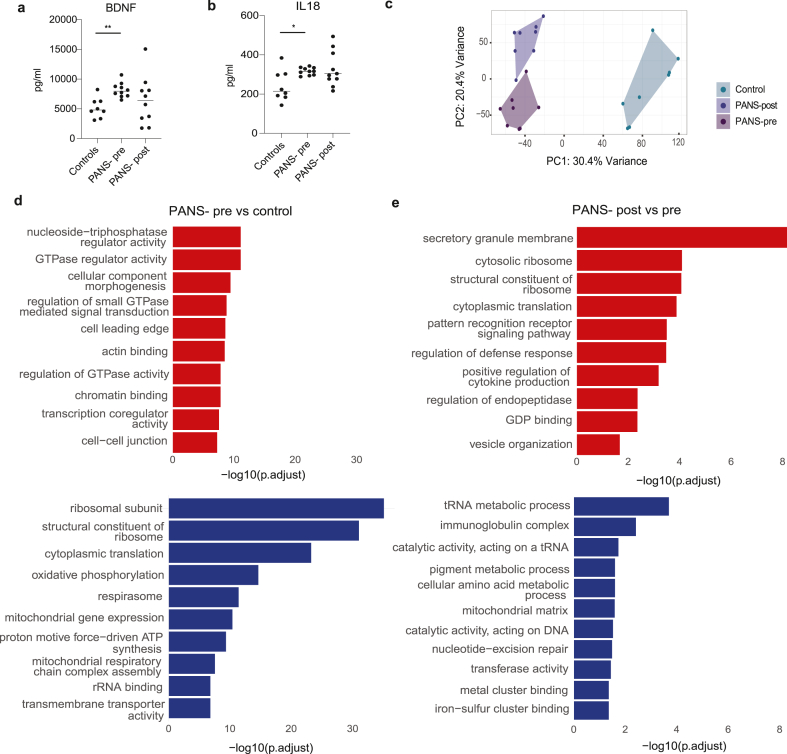
A multiplex 13-cytokine assay performed on plasma revealed elevated brain-derived neurotrophic factor (BDNF) (Fig. 2A, p = 0·0014) and interleukin-(IL)-18 (Fig. 2B, p = 0·0266) at baseline (PANS-pre) compared to controls. Other cytokines in the assay were generally increased in PANS-pre and reduced in patients with PANS post-NTI164, but no additional significant differences were found (Supplementary Fig. 2).
Bulk-RNA sequencing
Bulk-RNA sequencing was performed in eight patients at baseline (PANS-pre) and after 12 weeks of NTI164 treatment (PANS-post; mean age 11·4 (range 4–17) years), 37·5 % females), and eight age- and sex-matched controls (mean age 11·8 (range 7–17) years, 37·5 % females).
Principal component analysis and differentially expressed genes
Post RUV normalisation (Supplementary Fig. 3A–C), principal component analysis (PCA) of bulk RNA sequencing showed clear discrimination between healthy controls, PANS-pre, and PANS-post (Fig. 2C). In the PANS-pre vs control, there were 7,710 differentially expressed genes (DEGs) FDR <0·05, with 4,028 DEGs up-regulated and 3,682 down-regulated. In PANS-post vs PANS-pre, there were 4,974 DEGS FDR <0·05, with 2,794 up-regulated and 2,180 down-regulated.
Pathway analysis
In PANS-pre vs control, top 10 up-regulated GSEA GO pathways included cellular signalling (GTPase-related activities, ‘actin binding’), chromatin binding, and transcription-related pathways (Fig. 2D, red). Top 10 down-regulated pathways related to ribosome formation, cytoplasmic translation, mitochondrial function, and oxidative phosphorylation (Fig. 2D, blue).
After NTI164 treatment, the ribosomal and translational pathways (e.g. ‘structural constituent of ribosome’, ‘cytoplasmic translation’) that were down-regulated at baseline (PANS-pre vs controls), were up-regulated (PANS-post vs pre) (Fig. 2E). Additionally, several immune pathways (‘pattern recognition receptor signalling pathway’, ‘regulation of defense response’ and ‘positive regulation of cytokine production’) were up-regulated after NTI164 treatment (PANS-post vs pre). However, other immune pathways (‘immunoglobulin complex’) were down-regulated after NTI64 treatment (PANS-post vs pre), supporting broad immune modulatory effects of NTI164.
Single-cell RNA sequencing
Based on the findings from bulk RNA sequencing, a deeper investigation into cell-specific gene expression patterns was conducted via single-cell RNA sequencing (scRNA-seq). We performed scRNA-seq in four children with PANS at baseline (PANS-pre) and after 12 weeks of NTI164 treatment (PANS-post) (mean age 15·2 (range 14–17) years, 50 % females), and four age- and sex-matched healthy controls (mean age 14·5 years (range 12–17) years, 50 % females).
Uniform manifold approximation and projection (UMAP) and differentially expressed genes
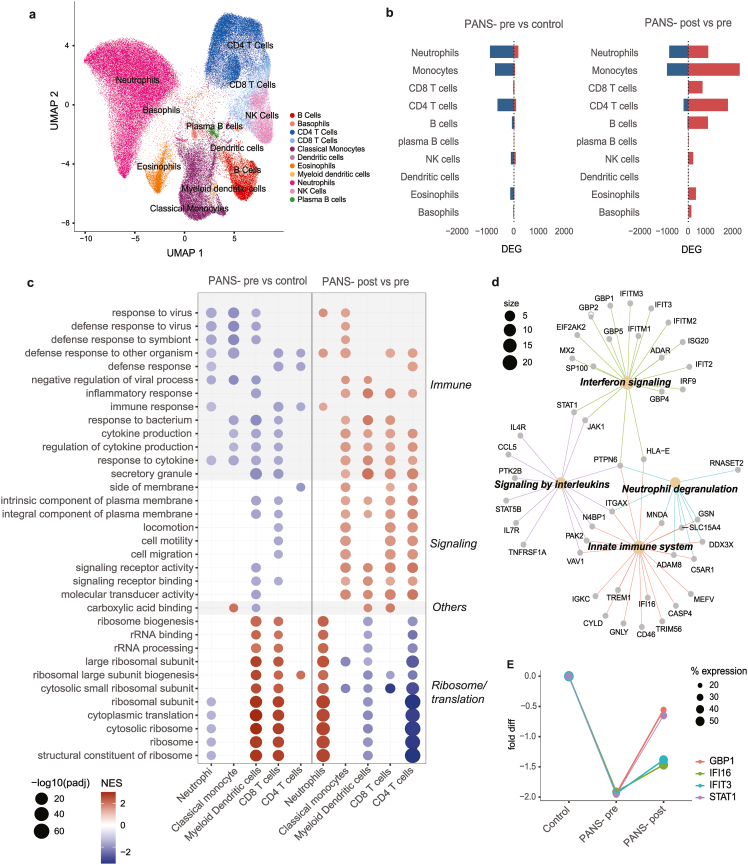
Based on cell markers, 11 distinct cell types were identified (Fig. 3A). Neutrophils constituted the largest proportion of cell type across samples. There were no significant differences observed in cell distribution between healthy controls, patients at baseline (PANS-pre), and after treatment (PANS-post) (Supplementary Fig. 4A–C). In PANS-pre vs control, DEGs (FDR <0·05) were predominantly down-regulated, most notably in neutrophils, monocytes, and CD4+ T cells (Fig. 3B, left). In PANS-post vs pre, DEGs (FDR <0·05) were predominantly up-regulated, most notably in neutrophils, monocytes, CD8+ T cells, CD4+ T cells, and B cells (Fig. 3B, right).
Pathway analysis
The most significantly enriched GSEA GO pathways were identified in neutrophils, classical monocytes, myeloid dendritic cells, CD8+ T cells, and CD4+ T cells (Fig. 3C). In PANS-pre vs controls, top down-regulated GSEA GO pathways were in immune function (‘defense response to virus/other symbiont/bacterium’, ‘cytokine production’, ‘inflammatory response’) and cellular signalling (Fig. 3C, left column). Additionally, ribosomal and RNA processing pathways showed cell-specific dysregulation in PANS-pre vs control, with down-regulation in neutrophils but up-regulation in myeloid dendritic cells and CD8+ T cells.
After NTI164 treatment, the immune function and cellular signalling pathways that were down-regulated at baseline (PANS-pre vs control), were generally up-regulated (PANS-post vs pre) (Fig. 3C, right column). The translation and ribosomal pathways that were dysregulated at baseline (PANS-pre vs controls) also demonstrated reversal after NTI164 treatment (PANS-post vs pre), most evident in neutrophils and myeloid dendritic cells.
We focussed on the 69 genes from the classical monocyte ‘defense response to other organism’ pathway that were significantly down-regulated (total 119 genes) in PANS-pre vs control, and up-regulated in PANS-post vs pre (total 143 genes) (Fig. 3D). Connectivity enrichment plot (CNET) of the 69 overlapping genes revealed GO molecular function subclusters, including interferon signalling (enriched by interferon-related genes: IFIT2, IFIT3, IFITM1, IFITM2, IFITM3), signalling by interleukins (enriched by signal transducer and activator of transcription genes: STAT1, STAT5B, Janus kinase genes: JAK1)), neutrophil degranulation and innate immune system (enriched by human leucocyte antigen genes: HLA-E, complement genes: C5AR1, integrin genes: ITGAX).
Gene expression of key genes
We further investigated trends of gene expression in key genes within the neutrophil GSEA GO pathway ‘defense response to other organisms’, including GBP1, IFI6, IFIT3, and STAT3 (Fig. 3E). These genes were significantly down-regulated in PANS-pre (compared to control), and increased in PANS-post, although not completely normalised to the level of healthy controls.
Immune and epigenetic pathways in the proteome and phosphoproteome in PANS are modified by NTI164
Proteomics
Bulk proteomics was performed on peripheral blood mononuclear cells (PBMCs) from six children with PANS at baseline (PANS-pre) and after 12 weeks of NTI164 treatment (PANS-post) (mean age 12 [ref. [4], ref. [5], ref. [6], ref. [7], ref. [8], ref. [9], ref. [10], ref. [11], ref. [12], ref. [13], ref. [14], ref. [15], ref. [16], ref. [17]] years, 33·3 % females), and six age- and sex-matched controls (mean age 12·3 [ref. [7], ref. [8], ref. [9], ref. [10], ref. [11], ref. [12], ref. [13], ref. [14], ref. [15], ref. [16], ref. [17]] years, 33·3 % females).
Differentially expressed proteins
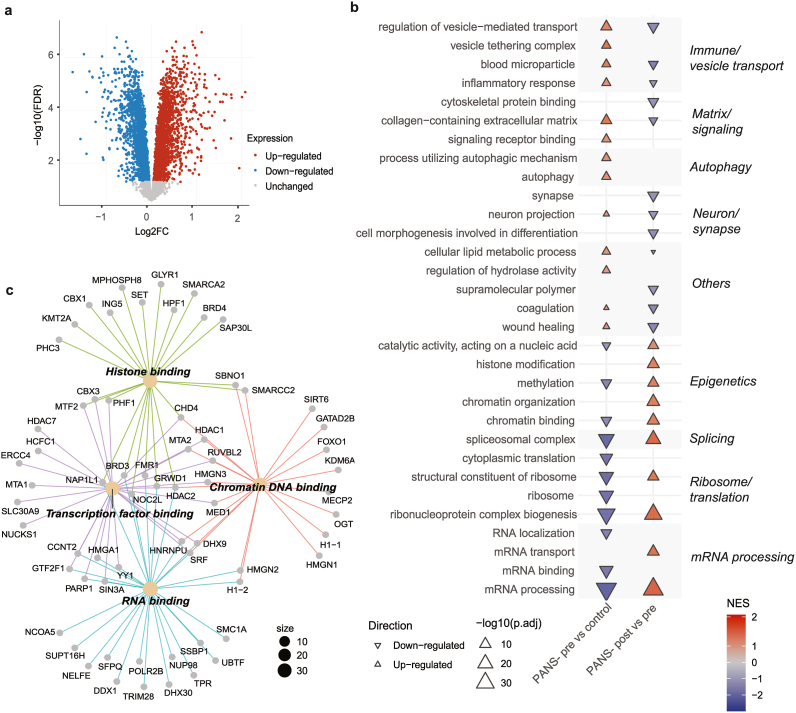
Post RUV normalisation, principal component analysis (PCA) of bulk proteomics showed clear discrimination between healthy controls, PANS-pre, and PANS-post (Supplementary Fig. 5A). In PANS-pre vs control, there were 4,518 differentially expressed proteins (DEPs) FDR <0·05, with 2,349 significantly up-regulated DEPs (Fig. 4A, red) and 2,169 down-regulated DEPs (Fig. 4A, blue). In PANS-post vs pre there were 3,502 DEPs FDR <0·05, with 1,654 up-regulated DEPs and 1,848 down-regulated DEPs (Supplementary Fig. 5B–D).
Pathway analysis
In PANS-pre vs control, top 10 up-regulated GSEA GO pathways were related to immune function, signalling, and autophagy (Fig. 4B, left column, red). Top 10 down-regulated pathways were related to mRNA processing, ribosome/translation, epigenetics (‘chromatin binding’, ‘methylation’) (Fig. 4B, left column, blue). NTI164 treatment considerably modified protein expression in many pathways (Fig. 4B, right column). Immune pathways that were up-regulated at baseline (PANS-pre vs control) showed significant down-regulation post treatment (PANS-post vs pre). Additionally, epigenetic and ribosome/translational pathways which were down-regulated at baseline (PANS-pre vs control) were significantly up-regulated after treatment (PANS-post vs pre).
We further investigated the 87 overlapping proteins from the ‘chromatin binding’ pathway that were significantly down-regulated in PANS-pre vs control (total 129 proteins), and up-regulated in the PANS-post vs pre (total 121 proteins) (Fig. 4C). CNET of the 87 overlapping proteins revealed GO molecular function subclusters, including histone binding (enriched by histone deacetylases: HDAC2, lysine methyltransferase: KMT2A, epigenetic regulators: BRD3, BRD4), transcription factor binding (enriched by histone deacetylases: HDAC1, HDAC7), chromatin DNA binding (enriched by lysine demethylase: KDM5A, histone genes: H1-1, epigenetic regulator: MeCP2, transcription factor: FOXO1), and RNA binding (enriched by histone genes: H1-2).
Phosphoproteomics
Bulk phosphoproteomics was performed on the same samples which underwent proteomics analysis (n = 6 patients pre and post and n = 6 controls).
Differentially abundant phosphopeptides
Post RUV normalisation, principal component analysis (PCA) of bulk phosphoproteomics showed clear discrimination between healthy controls, PANS-pre, and PANS-post (Supplementary Fig. 6A). In the PANS-pre vs control, there were 4,928 differentially abundant phosphopeptides (FDR <0·05), with 2,517 significantly up-regulated and 2,411 down-regulated differentially abundant phosphopeptides. In the PANS-post vs pre comparison there were 4,271 differentially abundant phosphopeptides (FDR <0·05), with 2,103 up-regulated and 2,168 down-regulated differentially abundant phosphopeptides (Supplementary Fig. 6B–D).
Pathway analysis
In PANS-pre vs control, top 10 ORA GO up-regulated pathways were cadherin binding, actin and protein kinase pathways (Fig. 5A, left column, red). The top 10 ORA GO down-regulated pathways were GTPase-related activity, chromatin (‘chromatin binding’, ‘mRNA processing’), and transcription pathways (Fig. 5A, left column, blue).
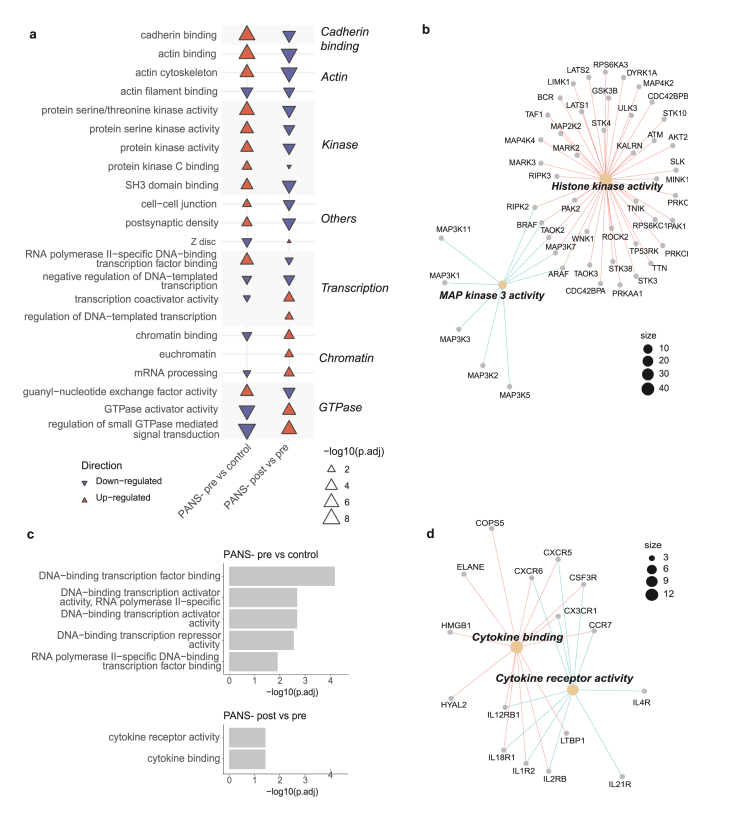
Modification of these pathways was seen following 12 weeks of NTI164 administration (PANS-post vs pre, Fig. 5A, right column). The cadherin binding, actin, and protein kinase activity pathways that were up-regulated in patients at baseline (PANS-pre vs control) were down-regulated after NTI164 treatment (PANS-post vs pre). Additionally, many GTPase signalling, chromatin, and transcription pathways that were down-regulated at baseline (PANS-pre vs control) were up-regulated following NTI164 treatment (PANS-post vs pre).
We focussed on the 53 proteins enriched by phosphopeptides from the ‘protein serine/threonine kinase activity’ pathway that were significantly up-regulated in the PANS-pre vs control comparison (74 proteins), and down-regulated in the PANS-post vs pre comparison (62 proteins) (Fig. 5B). CNET of these 53 overlapping proteins enriched by phosphopeptides revealed GO molecular function subclusters, including histone kinase activity, and MAP kinase 3 activity.
DNA methylation in PANS is modified by NTI164
DNA methylation was performed on whole blood from eight children with PANS at baseline (PANS-pre) and after 12 weeks of NTI164 treatment (PANS-post; mean age 11·4 (range 4–17) years, 37·5 % females), and eight age- and sex-matched controls (mean age 11·8 (range 7–17) years, 37·5 % females).
Differentially methylated positions
Post RUV normalisation (Supplementary Fig. 7A–C), principal component analysis (PCA) of DNA methylation showed discrimination between healthy controls, PANS-pre, and PANS-post (Supplementary Fig. 7D). In the PANS-pre vs control, there were 63,749 differentially methylated positions (FDR <0·05). In the PANS-post vs pre comparison there were 17,930 differentially methylated positions (FDR <0·05).
Differentially methylated region pathway analysis
Differentially methylated positions were assigned to differentially methylated regions. In PANS-pre vs control, top 5 significant ORA GO MF dysregulated pathways were all related to DNA transcription and included HDAC genes and TBX genes (Fig. 5C, top). Following 12 weeks of NTI164 administration (PANS-post vs pre, Fig. 5C, bottom), the 2 significant ORA GO Molecular Function dysregulated pathways involved cytokine activity. A CNET of these cytokine activity pathways revealed chemokine and interleukin genes corresponding to differentially methylated regions (Fig. 5D).
Discussion
In this open-label clinical trial of full-spectrum medicinal cannabis in children with PANS, we integrated clinical outcomes with multi-omics analyses to evaluate NTI164. Based on our hypothesis that PANS is an epigenetic disorder of immune and brain function, we selected a cannabis-based treatment due to its recognised epigenetic and immune modulating effects. NTI164 was well-tolerated at doses up to 20 mg/kg/day, with only one serious adverse event (diarrhoea) reported during the trial, and the child and family chose to remain on the drug. We observed significant clinical and functional improvements in children with PANS following treatment with NTI164. Using a multi-omics approach, we also identified dysregulated epigenetic (chromatin, methylation, histone modifications, transcription factor), ribosomal, mRNA processing/transcriptional, immune, and signalling (GTPase, actin) pathways in children with PANS, and showed that this dysregulation was modifiable with NTI164. We previously identified an immune, ribosomal, and epigenetic signature in peripheral immune cells from children with PANS [ref. 44]. This abnormal signature was reversed following treatment with the immunomodulators, such as intravenous immunoglobulin (IVIg) [ref. 44,ref. 45] and butyrate [ref. 46]. Here, we replicated our previous baseline findings in children with PANS and demonstrated similar therapeutic benefits using a proprietary strain of medicinal cannabis (NTI164).
Prior to treatment with NTI164, the PANS patients in our cohort were very debilitated with substantial functional impairment. All patients had previously trialled multiple medications and conventional therapy, and remained seriously functionally impaired. Additionally, the children suffered ongoing infection-triggered relapses which exacerbated impairments. Improvement in emotional assessments was the primary outcome of this trial. Following treatment with NTI164, clinical assessments demonstrated improvement in emotional regulation, anxiety, OCD, ADHD, tics, and overall Quality of Life (Fig. 1, Supplementary Fig. 1, Supplementary Table 4). There were significant improvements in social phobia, panic disorder, major depression, separation anxiety, generalised anxiety, and obsessive-compulsive symptoms, as assessed by the RCADS-P. A previous study of NTI164 in autistic spectrum disorder also showed similar clinical improvements in areas of anxiety, emotional regulation, and attention [ref. 47].
We incorporated a multi-omics approach to explore the ex vivo biological effects of NTI164. In this study of children with PANS, we observed abnormal gene and protein expression at baseline, along with altered DNA methylation, enriched in pathways related to immune function, which were modulated by NTI164 treatment. Based on single-cell RNA sequencing, we observed down-regulation of immune pathways in multiple cell types in PANS, affecting both adaptive and innate immune cell types, which were reversed with NTI164 treatment. Immune system dysfunction in PANS children pre-NTI164 was also suggested through cytokine testing, although only IL-18 and BDNF were also found to be significantly elevated in PANS children at baseline compared to healthy controls. The role of BDNF in CNS development and synaptic plasticity is well known [ref. 48,ref. 49], and a previous study in ASD patients suggests a link between increased BDNF levels and intellectual disability [ref. 50]. Although the cytokine and BDNF findings added some insights, these approaches were statistically less powerful compared to the RNA/proteomic approaches.
Cannabis, and in particular the isolated cannabinoids CBD and THC, has demonstrated clinical benefit in various immune and autoimmune related neurological conditions, including multiple sclerosis, neuropathic pain, and neurodegenerative conditions, in part due to its potent anti-inflammatory effects and reduction of oxidative stress [ref. 51]. Cannabinoid receptor CB2 is predominantly expressed on immune cells and interaction between the receptor and various cannabinoids can suppress inflammatory cytokine production, and alter T cell differentiation, macrophage activity, and B cell function [ref. [52], ref. [53], ref. [54]]. NTI164 has similarly been shown to suppress production of inflammatory cytokines and reduce expression of microglia activation state markers [ref. 14].
Although we focus on peripheral immune cells in this study, microglia, which are resident immune cells within the central nervous system, are hypothesised to be chronically activated in PANS [ref. 55]. Disrupted microglia-dependent synaptic pruning is a key process in the pathophysiology of NDDs [ref. 56], and epigenetic ‘priming’ of microglia and peripheral immune cells can lead to widespread immune system dysregulation [ref. 57,ref. 58]. A ‘two-hit’ model has been proposed for the development of NDDs, in which prenatal disruptions, such as maternal immune activation and chronic inflammation in utero, prime the foetal brain [ref. 8,ref. 59]. A subsequent postnatal ‘second hit’ postnatally such as infection or psychosocial stress, is thought to trigger immune dysregulation, leading to aberrant microglial activation and the clinical manifestation of NDDs [ref. 55]. We found high rates of maternal autoimmunity in our cohort, which has been reported in other PANS cohorts [ref. 59], and represents a possible example of the genetic and environmental (immune activation in pregnancy) factors relevant to PANS expression [ref. 8]. Maternal autoimmunity is known to be associated with increased expression of neurodevelopmental disorders (ASD, ADHD, Tourette) in offspring [ref. 8,ref. 60]. CBD is known to modulate an extensive array of microglial genes involved in regulating stress and inflammation [ref. 61], ameliorating microglia-dependent inflammation [ref. [62], ref. [63], ref. [64]], and limiting neuronal damage in excitotoxic environments [ref. 65]. In vitro studies of NTI164 have shown attenuation of microglial activation and associated pro-inflammatory cytokines, and reduced neuronal cell death following a stressor in cells treated with NTI164 [ref. 14], and these may be mechanisms through which our observed clinical dysregulation is normalised.
Secondly, we observed ribosomal and translational dysregulation in children with PANS compared to healthy controls through RNA and proteomic sequencing. Interestingly, the direction of dysregulation of translation-related pathways varied across cell types. For instance, ‘structural constituent of ribosome’ was down-regulated at baseline in neutrophils, and up-regulated in myeloid dendritic cells and CD8+ T cells. There are well-documented links between translation and ribosomal protein abnormalities in NDDs [ref. [66], ref. [67], ref. [68]]. Ribosomal function is fundamental for maintaining cellular homeostasis, through translation and regulation of protein synthesis. Disruption to ribosome biogenesis can reduce synthesis of proteins critical for dendritic and synaptic function, contributing to changes in neuronal function and connectivity in NDDs [ref. 69]. Additionally, ribosomal proteins can be involved in immune function [ref. 70]. Ribosome stress can trigger production of ribosomal proteins involved in signalling pathways, potentially contributing to downstream immune system dysregulation observed in PANS [ref. 44].
Thirdly, we identified broad epigenetic dysregulation relating to chromatin, methylation, histone modification and transcription factors at baseline across all omics methodologies, which was significantly modified with NTI164 treatment. We also identified dysregulation of GTPase and actin cytoskeletal pathways in children with PANS, which were reversed with NTI164 treatment. Increasing evidence supports the role of GTPase signalling and actin dynamics in chromatin remodelling, suggesting that they are mechanisms under epigenetic control [ref. 71,ref. 72]. Epigenetic dysregulation plays a crucial role in onset and progression of various NDDs [ref. 44,ref. 73]. Epigenetic mechanisms such as chromatin modification, DNA methylation, and transcription factors are essential for regulating gene expression and disruptions in these processes can lead to abnormal neural differentiation and connectivity. Chromatin abnormalities have been implicated in NDDs, and chromatin structure tightly regulated by histone modifications [ref. 74,ref. 75]. Proteomics analysis in our current study revealed proteins contributing to chromatin binding, specifically histone deacetylases, lysine demethylases, lysine methyltransferases, which were down-regulated at baseline and up-regulated with NTI164. Our study showed that protein kinases with histone kinase activity were abnormal in PANS children and modified by NTI164; histone phosphorylation by kinases is also involved in chromatin structure and function [ref. 76]. Further research is needed to explore the effects of NTI164 on histone kinases and phosphorylation of histones, as well as downstream effects on chromatin structure and gene expression, plus other histone modification (methylation, acetylation etc).
Analysis of DNA methylation, another epigenetic modification, in our current study revealed PANS patients have differences in methylation in DNA transcription-related regions, supporting the concept that PANS is an epigenetic disorder with associated abnormal gene expression and downstream effects on the immune system. Some studies suggest methylation changes in genes relating to synaptic or neuronal development leads to altered brain function contributing to OCD, anxiety, and tics [ref. 77,ref. 78].
The epigenetic effects of cannabis are only beginning to be explored [ref. 79,ref. 80]. Isolated CBD can affect histone acetyltransferases and histone deacetylases, and a study in adult mice demonstrated CBD can also work synergistically with other cannabinoids to induce histone modifications in the mesolimbic system and improve motor outcomes and anxiety [ref. 81]. Cannabis’ effect on epigenetic ‘reprogramming’ has also been suggested to involve repression of genes relating to inflammation and thus dampening immune responses, and promoting immunosuppressive effects [ref. 82]. Cannabinoids bound to CB1 and CB2 receptors can inhibit transcription factors associated with modification of inflammatory gene expression [ref. 53,ref. 83], and certain histone modifications induced by THC and CBD have been shown to promote heterochromatin formation and gene ‘silencing’, leading to long-term suppression of specific transcripts [ref. 84,ref. 85]. More research into the epigenetic effects of cannabis and NTI164 is required, utilising standardised drug compositions and assessments. This should include analysis of the effects of NTI164 on histone modifications, chromatin function, and methylation, and its downstream effects on expression of NDD and immune genes. Chromatin analyses (e.g. ATAC-seq or ChIP-seq) and microRNA analysis would be useful in further exploring epigenetic dysregulation in PANS [ref. 86].
Limitations of the current study include the small sample size and open-label nature of our clinical trial. As this was a Phase I/II, proof-of-concept trial, an open-label design was pursued. A Phase II/III, randomised, placebo-controlled, double-blind clinical trial of NTI164 in PANS is currently under development, and we acknowledge future research involving larger randomised controlled cohorts, including comparison of responders with non-responders, will be necessary to validate these findings and better evaluate mechanistic pathways. Our patients were also on various other medications, including anti-psychotics and anti-depressants, which could influence baseline RNA or protein expression, but we controlled for this by prohibiting changes to medication regimes in the three months prior to enrolment and during the study period. Also, our multi-omics investigations were based on peripheral immune cells and not central nervous system cells, however emerging preclinical research does show comparable dysregulation between the periphery and brain tissue in an animal model of NDDs [ref. 87]. Further investigation using animal models or patient-derived induced pluripotent stem cells (iPSCs) would be useful to determine the effects of NTI164 on brain cells in the context of PANS. Additional studies are also needed to further validate our findings, including functional immune testing, ribosomal profiling, epigenetic and phosphorylation studies, and to investigate the long-term effects and safety of cannabis. Future studies could also consider additional validation of key genes and proteins modulated by medicinal cannabis using real-time PCR and protein quantification using Western blotting.
This study demonstrates the clinical benefits of NTI164, a novel full-spectrum medicinal cannabis plant extract, in children with PANS. Our results show NTI164 has epigenetic, ribosomal and immune modulatory effects, and can reverse baseline abnormalities observed in PANS. While further studies are needed, these results show significant potential for NTI164 in PANS and potentially other neurological conditions involving epigenetic and immune dysregulation.
Resource availability and data sharing
Additional data and code available upon request by qualified Investigators. Deidentified patient standardised bulk RNA sequencing data and single-cell RNA sequencing data have been deposited at Gene Expression Omnibus as GSE299764 and GSE301611, respectively. Deidentified patient bulk proteomics and phosphoproteomics data has been deposited at ProteomeXchange via the PRIDE database with identifier PXD064184. Requests for further information should be directed to the corresponding author, Prof Russell Dale.
Author contributions
BAK, MK, MCF, SP, and RCD designed the trial. BAK, VXH, HN, SSM, MK, MCF, SP, and RCD recruited the patients and controls. BAK, MK, MCF, and RCD performed clinical assessments. BAK, VXH, HN, XL, RD, SD, MG, SP, and RCD conducted experiments and acquired and analysed the data. BAK, SSM, MK, MCF, SP, and RCD oversaw the trial and data analysis. VXH, HN, NA, LLM, BSG, and MG performed computational analyses. BAK and VXH drafted this manuscript and shared responsibilities as co-first authors. SP and RCD critically revised this manuscript and shared responsibilities as co-senior authors.
Declaration of competing interest
The authors declare the following financial interests/personal relationships which may be considered as potential competing interests: Russell C Dale reports financial support was provided by Fenix Innovation Group Pty Ltd. Russell C Dale reports financial support was provided by Neurotech International Ltd. Brooke A Keating reports a relationship with Fenix Innovation Group Pty Ltd that includes: employment. BAK became affiliated with Fenix Innovation Group Pty Ltd (Sponsor) following completion of this trial. BAK was not affiliated with Fenix Innovation Group Pty Ltd during collection or analysis of the data presented in this paper. All other authors declare no conflicting interests.
If there are other authors, they declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- S.E. Swedo, J. Seidlitz, M. Kovacevic, M.E. Latimer, R. Hommer, L. Lougee. Clinical presentation of pediatric autoimmune neuropsychiatric disorders associated with streptococcal infections in research and community settings. J Child Adolesc Psychopharmacol, 2015. [PubMed]
- K.D. Brown, C. Farmer, G.M. Freeman, E.J. Spartz, B. Farhadian, M. Thienemann. Effect of early and prophylactic nonsteroidal anti-inflammatory drugs on flare duration in pediatric acute-onset neuropsychiatric syndrome: an observational study of patients followed by an academic community-based pediatric acute-onset neuropsychiatric syndrome clinic. J Child Adolesc Psychopharmacol, 2017. [PubMed]
- M.E. Franklin, S. Eken, E. Osterlund. Current research updates on PANDAS and PANS. Curr Develop Disord Rep, 2023
- Board of Directors. Pediatric Acute-Onset Neuropsychiatric Syndrome (PANS): clinical report. Pediatrics, 2025 Mar 1
- B. Zablotsky, L.I. Black, M.J. Maenner, L.A. Schieve, M.L. Danielson, R.H. Bitsko. Prevalence and trends of developmental disabilities among children in the United States: 2009-2017. Pediatrics, 2019
- R. Trifiletti, H.M. Lachman, O. Manusama, D. Zheng, A. Spalice, P. Chiurazzi. Identification of ultra-rare genetic variants in pediatric acute onset neuropsychiatric syndrome (PANS) by exome and whole genome sequencing. Sci Rep, 2022
- S. Pallanti, M. Di Ponzio. PANDAS/PANS in the COVID-19 age: autoimmunity and Epstein–Barr Virus reactivation as trigger agents?. Children, 2023. [PubMed]
- V.X. Han, S. Patel, H.F. Jones, T.C. Nielsen, S.S. Mohammad, M.J. Hofer. Maternal acute and chronic inflammation in pregnancy is associated with common neurodevelopmental disorders: a systematic review. Transl Psychiatry, 2021. [PubMed]
- R.C. Dale, S. Mohammad, V.X. Han, H. Nishida, H. Goel, S.G. Tangye. Pathogenic variants in chromatin-related genes: linking immune dysregulation to neuroregression and acute neuropsychiatric disorders. Dev Med Child Neurol, 2025 Aug. [PubMed]
- H. Nishida, V.X. Han, B.A. Keating, K.G. Zyner, B. Gloss, N. Aryamanesh. Chromatin, transcriptional and immune dysregulation in children with neurodevelopmental regression. medRxiv, 2025. [DOI]
- M.B. Bridgeman, D.T. Abazia. Medicinal cannabis: history, pharmacology, and implications for the acute care setting. P t, 2017. [PubMed]
- J.C. Arnold, D. McCartney, A. Suraev, I.S. McGregor. The safety and efficacy of low oral doses of cannabidiol: an evaluation of the evidence. Clin Transl Sci, 2023. [PubMed]
- E.B. Russo. The case for the entourage effect and conventional breeding of clinical cannabis: no “Strain,” no gain. Front Plant Sci, 2019
- E. Ross-Munro, E. Isikgel, B. Fleiss. Evaluation of the efficacy of a full-spectrum Low-THC cannabis plant extract using In vitro models of inflammation and excitotoxicity. Biomolecules, 2024
- O. Devinsky, N.A. Jones, M.O. Cunningham, B.A.P. Jayasekera, S. Devore, B.J. Whalley. Cannabinoid treatments in epilepsy and seizure disorders. Physiol Rev, 2024. [PubMed]
- O.J. Manzoni, A. Manduca, V. Trezza. Therapeutic potential of cannabidiol polypharmacology in neuropsychiatric disorders. Trends Pharmacol Sci, 2025. [PubMed]
- Naya N. Martinez, J. Kelly, G. Corna, M. Golino, A. Abbate, S. Toldo. Molecular and cellular mechanisms of action of cannabidiol. Molecules, 2023
- A.C. Campos, F.A. Moreira, F.V. Gomes, E.A. Del Bel, F.S. Guimarães. Multiple mechanisms involved in the large-spectrum therapeutic potential of cannabidiol in psychiatric disorders. Philos Trans R Soc Lond B Biol Sci, 2012. [PubMed]
- J.A. Crippa, F.S. Guimarães, A.C. Campos, A.W. Zuardi. Translational investigation of the therapeutic potential of cannabidiol (CBD): toward a new age. Front Immunol, 2018. [PubMed]
- P. McGuire, P. Robson, W.J. Cubala, D. Vasile, P.D. Morrison, R. Barron. Cannabidiol (CBD) as an adjunctive therapy in schizophrenia: a multicenter randomized controlled trial. Am J Psychiatr, 2018. [PubMed]
- S. Bhattacharyya, R. Wilson, E. Appiah-Kusi, A. O’Neill, M. Brammer, J. Perez. Effect of cannabidiol on medial temporal, midbrain, and striatal dysfunction in people at clinical high risk of psychosis: a randomized clinical trial. JAMA Psychiatry, 2018. [PubMed]
- K. Chang, J. Frankovich, M. Cooperstock, M.W. Cunningham, M.E. Latimer, T.K. Murphy. Clinical evaluation of youth with pediatric acute-onset neuropsychiatric syndrome (PANS): recommendations from the 2013 PANS consensus conference. J Child Adolesc Psychopharmacol, 2015. [PubMed]
- A. Dobin, C.A. Davis, F. Schlesinger, J. Drenkow, C. Zaleski, S. Jha. STAR: ultrafast universal RNA-seq aligner. Bioinformatics, 2013. [PubMed]
- E. Tsang, V.X. Han, C. Flutter, S. Alshammery, B.A. Keating, T. Williams. Ketogenic diet modifies ribosomal protein dysregulation in KMT2D Kabuki syndrome. EBioMedicine, 2024
- E. Cruz, R.M. Nisbet, P. Padmanabhan, A.J. van Waardenberg, M.E. Graham, G. Nkajja. Proteostasis as a fundamental principle of Tau immunotherapy. Brain, 2025. [PubMed]
- R.D.C.R. Team. 2010
- W. Huber, V.J. Carey, R. Gentleman, S. Anders, M. Carlson, B.S. Carvalho. Orchestrating high-throughput genomic analysis with bioconductor. Nat Methods, 2015. [PubMed]
- G.K. Smyth. Bioinformatics and computational biology solutions using R and Bioconductor, 2005
- H. Wickham, M. Averick, J. Bryan, W. Chang, L.D.A. McGowan, R. François. Welcome to the tidyverse. J Open Source Softw, 2019
- D. Risso, J. Ngai, T.P. Speed, S. Dudoit. Normalization of RNA-seq data using factor analysis of control genes or samples. Nat Biotechnol, 2014. [PubMed]
- M.E. Ritchie, B. Phipson, D. Wu, Y. Hu, C.W. Law, W. Shi. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res, 2015. [PubMed]
- Y. Hao, S. Hao, E. Andersen-Nissen, W.M. Mauck, S. Zheng, A. Butler. Integrated analysis of multimodal single-cell data. Cell, 2021. [PubMed]
- J. Alquicira-Hernandez, A. Sathe, H.P. Ji, Q. Nguyen, J.E. Powell. scPred: accurate supervised method for cell-type classification from single-cell RNA-seq data. Genome Biol, 2019. [PubMed]
- I. Korsunsky, N. Millard, J. Fan, K. Slowikowski, F. Zhang, K. Wei. Fast, sensitive and accurate integration of single-cell data with Harmony. Nat Methods, 2019. [PubMed]
- M.J. Aryee, A.E. Jaffe, H. Corrada-Bravo, C. Ladd-Acosta, A.P. Feinberg, K.D. Hansen. Minfi: a flexible and comprehensive Bioconductor package for the analysis of Infinium DNA methylation microarrays. Bioinformatics, 2014. [PubMed]
- J. Maksimovic, B. Phipson, A. Oshlack. A cross-package bioconductor workflow for analysing methylation array data. F1000Res, 2016. [PubMed]
- R. Molania, J.A. Gagnon-Bartsch, A. Dobrovic, T.P. Speed. A new normalization for nanostring nCounter gene expression data. Nucleic Acids Res, 2019. [PubMed]
- T.J. Peters, M.J. Buckley, A.L. Statham, R. Pidsley, K. Samaras, R. V Lord. De novo identification of differentially methylated regions in the human genome. Epigenetics Chromatin, 2015. [PubMed]
- A. Subramanian, P. Tamayo, V.K. Mootha, S. Mukherjee, B.L. Ebert, M.A. Gillette. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc Natl Acad Sci, 2005. [PubMed]
- J. Reimand, R. Isserlin, V. Voisin, M. Kucera, C. Tannus-Lopes, A. Rostamianfar. Pathway enrichment analysis and visualization of omics data using g: profiler, GSEA, Cytoscape and EnrichmentMap. Nat Protoc, 2019. [PubMed]
- T. Wu, E. Hu, S. Xu, M. Chen, P. Guo, Z. Dai. clusterProfiler 4.0: a universal enrichment tool for interpreting omics data. Innovation, 2021
- G. Yu, L.G. Wang, Y. Han, Q.Y. He. clusterProfiler: an R package for comparing biological themes among gene clusters. OMICS, 2012. [PubMed]
- G. Yu, Q.Y. He. ReactomePA: an R/Bioconductor package for reactome pathway analysis and visualization. Mol Biosyst, 2016. [PubMed]
- V.X. Han, S. Alshammery, B.A. Keating, B.S. Gloss, M.J. Hofer, M.E. Graham. Epigenetic, ribosomal, and immune dysregulation in paediatric acute-onset neuropsychiatric syndrome. Mol Psychiatr, 2025
- V.X. Han, H. Nishida, B.A. Keating, B.S. Gloss, X. Lau, R. Dissanayake. IV immunoglobulin is associated with epigenetic, ribosomal, and immune changes in pediatric acute-onset neuropsychiatric syndrome. Neurol Neuroimmunol Neuroinflamm, 2025
- J.P. Hayes, V.X. Han, B.A. Keating, H. Nishida, E. Tsang, X. Lau. Butyrate modifies epigenetic and immune pathways in peripheral mononuclear cells from children with neurodevelopmental disorders associated with chromatin dysregulation. Neurotherapeutics, 2025
- D. El Sukkari, K. Sharma, B. Fleiss, D.L. Braganza, B.A. Keating, Y. Ogru. Safety and efficacy of orally administered full-spectrum Medicinal Cannabis Plant Extract 0.08% THC (NTI164) in children with autism spectrum disorder. Adv Comp Alter Med, 2025. [DOI]
- S. Camuso, P. La Rosa, M.T. Fiorenza, S. Canterini. Pleiotropic effects of BDNF on the cerebellum and hippocampus: implications for neurodevelopmental disorders. Neurobiol Dis, 2022
- Y. Li, F. Li, D. Qin, H. Chen, J. Wang, J. Wang. The role of brain derived neurotrophic factor in central nervous system. Front Aging Neurosci, 2022
- V. Bryn, B. Halvorsen, T. Ueland, J. Isaksen, K. Kolkova, K. Ravn. Brain derived neurotrophic factor (BDNF) and autism spectrum disorders (ASD) in childhood. Eur J Paediatr Neurol, 2015. [PubMed]
- 51Nichols JM, Kaplan BLF. Immune responses regulated by Cannabidiol. (2578-5125 (Print)).
- S. Devi, A.M. Zimmermann-Klemd, B.L. Fiebich, M. Heinrich, C. Gründemann, P. Steinberger. Immunosuppressive activity of non-psychoactive Cannabis sativa L. extract on the function of human T lymphocytes. Int Immunopharmacol, 2022
- E.G. Gojani, B. Wang, D.-P. Li, O. Kovalchuk, I. Kovalchuk. Anti-inflammatory effects of minor cannabinoids CBC, THCV, and CBN in human macrophages. Molecules, 2023. [PubMed]
- M.-C. Lampron, I. Paré, M. Al-Zharani, A. Semlali, L. Loubaki. Cannabinoid mixture affects the fate and functions of B cells through the modulation of the caspase and MAP kinase pathways. Cells, 2023. [PubMed]
- L. Frick, C. Pittenger. Microglial dysregulation in OCD, Tourette syndrome, and PANDAS. J Immunol Res, 2016
- F. Ginhoux, M. Prinz. Origin of microglia: current concepts and past controversies. Cold Spring Harbor Perspect Biol, 2015
- M. Bordeleau, C. Lacabanne, L. Fernández de Cossío, N. Vernoux, J.C. Savage, F. González-Ibáñez. Microglial and peripheral immune priming is partially sexually dimorphic in adolescent mouse offspring exposed to maternal high-fat diet. J Neuroinflammation, 2020. [PubMed]
- L.N. Hayes, K. An, E. Carloni, F. Li, E. Vincent, C. Trippaers. Prenatal immune stress blunts microglia reactivity, impairing neurocircuitry. Nature, 2022. [PubMed]
- H.F. Jones, V.X. Han, S. Patel, B.S. Gloss, N. Soler, A. Ho. Maternal autoimmunity and inflammation are associated with childhood tics and obsessive-compulsive disorder: transcriptomic data show common enriched innate immune pathways. Brain Behav Immun, 2021. [PubMed]
- T.C. Nielsen, N. Nassar, A.W. Shand, H.F. Jones, V.X. Han, S. Patel. Association of maternal autoimmune disease and early childhood infections with offspring autism spectrum disorder: a population-based cohort study. Autism Res, 2022. [PubMed]
- A. Juknat, N. Rimmerman, R. Levy, Z. Vogel, E. Kozela. Cannabidiol affects the expression of genes involved in zinc homeostasis in BV-2 microglial cells. Neurochem Int, 2012. [PubMed]
- L. Barata, L. Arruza, M.-J. Rodríguez, E. Aleo, E. Vierge, E. Criado. Neuroprotection by cannabidiol and hypothermia in a piglet model of newborn hypoxic-ischemic brain damage. Neuropharmacology, 2019. [PubMed]
- 63Dos-Santos-Pereira M, Guimarães FS, Del-Bel EA-O, Raisman-Vozari R, Michel PA-O. Cannabidiol prevents LPS-induced microglial inflammation by inhibiting ROS/NF-κB-dependent signaling and glucose consumption. (1098-1136 (Electronic)).
- A.M. Martín-Moreno, D. Reigada, B.G. Ramírez, R. Mechoulam, N. Innamorato, A. Cuadrado. Cannabidiol and other cannabinoids reduce microglial activation in vitro and in vivo: relevance to Alzheimer’s disease. Mol Pharmacol, 2011. [PubMed]
- E. Landucci, C. Mazzantini, D. Lana, M. Calvani, G. Magni, M.G. Giovannini. Cannabidiol inhibits microglia activation and mitigates neuronal damage induced by kainate in an in-vitro seizure model. Neurobiol Dis, 2022
- Y.C. Chen, Y.W. Chang, Y.S. Huang. Dysregulated translation in neurodevelopmental disorders: an overview of autism-risk genes involved in translation. Dev Neurobiol, 2019. [PubMed]
- M. Hetman, L.P. Slomnicki. Ribosomal biogenesis as an emerging target of neurodevelopmental pathologies. J Neurochem, 2019. [PubMed]
- Y. Park, N. Page, I. Salamon, D. Li, M.R. Rasin. Making sense of mRNA landscapes: translation control in neurodevelopment. Wiley Interdiscip Rev RNA, 2022
- W. Szaflarski, M. Leśniczak-Staszak, M. Sowiński, S. Ojha, A. Aulas, D. Dave. Early rRNA processing is a stress-dependent regulatory event whose inhibition maintains nucleolar integrity. Nucleic Acids Res, 2022. [PubMed]
- X. Zhou, W.-J. Liao, J.-M. Liao, P. Liao, H. Lu. Ribosomal proteins: functions beyond the ribosome. J Mol Cell Biol, 2015. [PubMed]
- M. Lasser, J. Tiber, L.A. Lowery. The role of the microtubule cytoskeleton in neurodevelopmental disorders. Front Cell Neurosci, 2018. [PubMed]
- A. Reichova, M. Zatkova, Z. Bacova, J. Bakos. Abnormalities in interactions of Rho GTPases with scaffolding proteins contribute to neurodevelopmental disorders. J Neurosci Res, 2018. [PubMed]
- A. Gagliano, A. Carta, M.G. Tanca, S. Sotgiu. Pediatric acute-onset neuropsychiatric syndrome: current perspectives. Neuropsychiatric Dis Treat, 2023
- B. Mossink, M. Negwer, D. Schubert, N. Nadif Kasri. The emerging role of chromatin remodelers in neurodevelopmental disorders: a developmental perspective. Cell Mol Life Sci, 2021. [PubMed]
- J. Reichard, G. Zimmer-Bensch. The epigenome in neurodevelopmental disorders. Front Neurosci, 2021
- D. Rossetto, N. Avvakumov, J. Côté. Histone phosphorylation: a chromatin modification involved in diverse nuclear events. Epigenetics, 2012. [PubMed]
- S.J. Goodman, C.L. Burton, D.T. Butcher, M.T. Siu, M. Lemire, E. Chater-Diehl. Obsessive-compulsive disorder and attention-deficit/hyperactivity disorder: distinct associations with DNA methylation and genetic variation. J Neurodev Disord, 2020. [PubMed]
- M.A. Schiele, J. Lipovsek, P. Schlosser, M. Soutschek, G. Schratt, M. Zaudig. Epigenome-wide DNA methylation in obsessive-compulsive disorder. Transl Psychiatry, 2022. [PubMed]
- H. Szutorisz, Y.L. Hurd. Epigenetic effects of cannabis exposure. Biol Psychiatry, 2016. [PubMed]
- A. Smith, F. Kaufman, M.S. Sandy, A. Cardenas. Cannabis exposure during critical windows of development: epigenetic and molecular pathways implicated in neuropsychiatric disease. Curr Environ Health Rep, 2020. [PubMed]
- S.M. Todd, C. Zhou, D.J. Clarke, T.W. Chohan, D. Bahceci, J.C. Arnold. Interactions between cannabidiol and Δ9-THC following acute and repeated dosing: rebound hyperactivity, sensorimotor gating and epigenetic and neuroadaptive changes in the mesolimbic pathway. Eur Neuropsychopharmacol, 2017. [PubMed]
- B. Wang, D. Li, A. Fiselier, I. Kovalchuk, O. Kovalchuk. High-CBD cannabis extracts inhibit the expression of proinflammatory factors via miRNA-mediated silencing in human small intestinal epithelial cells. Heliyon, 2023
- R.L. Silva, G.T. Silveira, C.W. Wanderlei, N.T. Cecilio, A.G. Maganin, M. Franchin. DMH-CBD, a cannabidiol analog with reduced cytotoxicity, inhibits TNF production by targeting NF-kB activity dependent on A2A receptor. Toxicol Appl Pharmacol, 2019. [PubMed]
- X. Yang, V.L. Hegde, R. Rao, J. Zhang, P.S. Nagarkatti, M. Nagarkatti. Histone modifications are associated with Δ9-tetrahydrocannabinol-mediated alterations in antigen-specific T cell responses. J Biol Chem, 2014. [PubMed]
- B.L. Holloman, M. Nagarkatti, P. Nagarkatti. Epigenetic regulation of cannabinoid-mediated attenuation of inflammation and its impact on the use of cannabinoids to treat autoimmune diseases. Int J Mol Sci, 2021. [PubMed]
- T.-T. Yao, L. Chen, Y. Du, Z.-Y. Jiang, Y. Cheng. MicroRNAs as regulators, biomarkers, and therapeutic targets in autism spectrum disorder. Mol Neurobiol, 2025. [PubMed]
- M.C. Bucknor, B.A. Keating, V.X. Han, B.S. Gloss, P. Dey, N. Aryamanesh. Cumulative pregnancy and postnatal environmental exposures impact social behaviour in male mice associated with epigenetic, ribosomal, and immune dysregulation. bioRxiv, 2025