
Deciphering Mitochondria: Unveiling Their Roles in Mechanosensing and Mechanotransduction
Abstract
Mitochondria are highly dynamic organelles that are responsible for essential cellular functions such as calcium regulation, reactive oxygen species (ROS) production, metabolism, and apoptosis initiation. Mitochondrial dysfunctions are associated with a variety of pathologies, and the onset and progression of disease are accompanied by alterations in extracellular biochemical and mechanical signals. Recent studies have demonstrated that physicochemical cues, especially mechanical cues, exert pivotal roles in the organization of mitochondrial network and their metabolic functions. Therefore, understanding the mechanisms that orchestrate mitochondrial morphology and function is essential for elucidating their role in both health and disease. This review discusses novel insights into the recent advances regarding mitochondrial dysfunction across a spectrum of diseases and describes the effect of various factors. It then highlights the recently discovered mechanisms, particularly those involving matrix mechanical cues and cellular mechanical cues, summarizing the multiple pathways of mechanotransduction, such as integrin, Piezo1/TRPV4, and YAP/TAZ signaling pathways. Last, the review explores the potential future directions, stressing that understanding mitochondrial dysfunction is crucial for developing effective therapies to improve mitochondrial function and address related diseases.
Affiliations: Department of Medical Oncology, Sichuan Clinical Research Center for Cancer, Sichuan Cancer Hospital & Institute, Sichuan Cancer Center, and School of Life Science and Technology, University of Electronic Science and Technology of China, Chengdu 610041, Sichuan, P. R. China.; TCM Regulating Metabolic Diseases Key Laboratory of Sichuan Province, Hospital of Chengdu University of Traditional Chinese Medicine, Chengdu 610072, Sichuan, P. R. China.; Department of Otorhinolaryngology, Hospital of Chengdu University of Traditional Chinese Medicine, Chengdu 610072, P. R. China.; Department of Urology, Deyang People’s Hospital, Deyang 618099, Sichuan, P. R. China.
License: Copyright © 2025 Jiaxuan Yu et al. CC BY 4.0 Exclusive licensee Science and Technology Review Publishing House. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution License (CC BY 4.0).
Article links: DOI: 10.34133/research.0816 | PubMed: 40785969 | PMC: PMC12332263
Relevance: Moderate: mentioned 3+ times in text
Full text: PDF (7.5 MB)
Introduction
Mitochondria are highly dynamic and autonomous organelles within the cell. As the central hubs of energy metabolism, mitochondria are involved not only in adenosine triphosphate (ATP) synthesis but also in the biosynthesis of fatty acids and the generation of reactive oxygen species (ROS). Beyond these metabolic functions, mitochondria are integral to a spectrum of signal transduction pathways, including oxidative stress, cell differentiation, apoptosis, and mitochondrial autophagy [ref. 1]. These features and functions render mitochondria a multifunctional central hub, essential for maintaining the normal physiological functions of the cells. The highly variable shapes, numbers, sizes, and locations resulting from mitochondrial fusion, division, mitophagy, and transport are collectively referred to as mitochondrial dynamics [ref. 2]. These dynamic processes are essential for maintaining mitochondrial functions and cellular functions [ref. 3]. Dysregulation of mitochondrial dynamics has been linked to the development of a variety of diseases, including neurodegenerative disease, metabolic syndrome, cardiovascular disease (CVD), and cancer [ref. 4]. Therefore, regulation of mitochondrial dynamics is considered a potential strategy for treating these diseases.
Mitochondrial dynamics vary greatly among different cell types, stages of cell growth, and stressful stimuli [ref. 5]. Cells can modify their shape and function in response to different extracellular biochemical and mechanical cues. They adjust to environmental shifts by managing mitochondrial dynamics and function. Historically, research has concentrated on extracellular biochemical signals such as glucose levels, pH values, and calcium concentration [ref. 6–ref. 9]. However, ongoing studies reveal that the extracellular microenvironment is intricate and variable, encompassing diverse cell types, their secreted factors, and the extracellular matrix (ECM) [ref. 7]. It is established that the mechanical characteristics of the matrix, including stiffness and viscoelasticity, can trigger mechanosensitive receptors and sensors on cell membranes [ref. 10]. These mechanical characteristics influence cell behavior and fate via intracellular mechanotransduction pathways. Given that diseases such as cardiovascular conditions and cancers often involve ECM fibrosis, targeting matrix stiffness has emerged as a potent therapeutic strategy [ref. 11]. The cytoskeleton, responsible for cell shape and movement through its assembly and disassembly, is crucial in the malignant migration of tumor cells. It also plays a crucial role in controlling mitochondrial dynamics and function [ref. 12].
In this review, we initially elucidate the critical role of mitochondrial dynamics and function in disease progression, subsequently focusing on how cells perceive extracellular mechanical signals to modulate mitochondrial dynamics and function through mechanotransduction, as well as the involvement of the cytoskeleton in regulating these processes.
Mitochondrial Dysfunction and Diseases
To maintain the stability and balance of cellular energy metabolism, mitochondria can flexibly respond to functional demands and adjust their internal composition and behavioral patterns through complex mitochondrial dynamics processes [ref. 13,ref. 14]. This process not only governs the increases or decreases in mitochondrial quantity, shape transformation, and size adjustment but also determines their distribution within the cell [ref. 15,ref. 16]. Therefore, the dysfunction caused by mitochondrial dynamics will affect the balance of cellular energy metabolism, thereby accelerating the deterioration of CVDs, neurodegenerative disorders (NDs), cancer, etc., which are illustrated in Fig. 1.
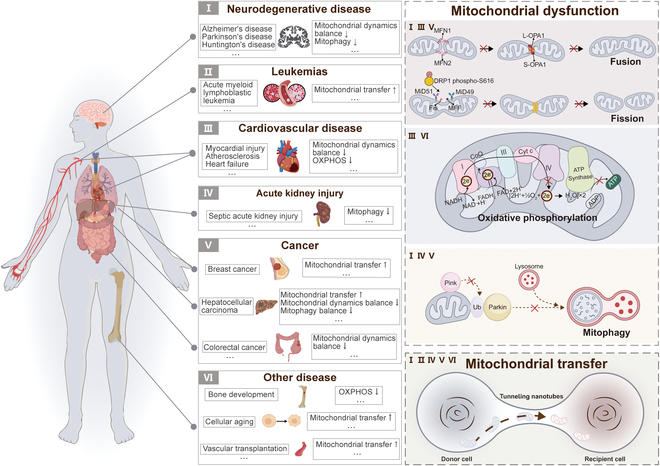
ND includes Alzheimer’s disease (AD), Parkinson’s disease (PD), and Huntington’s disease (HD), which is characterized by continuous neuronal degeneration and death, leading to a variety of neurological symptoms. Research has demonstrated that these disorders are accompanied by imbalances in mitochondrial fission and fusion [ref. 17–ref. 19], along with abnormal mitophagy [ref. 20]. In addition, studies of CVD patient samples have shown a significant inverse correlation between oxidative phosphorylation (OXPHOS) levels and the extent of atherosclerotic lesions [ref. 21]. Similarly, in heart failure (HF), by improving the mitochondrial OXPHOS process, it effectively alleviated cardiac insufficiency and slowed HF progression [ref. 22]. In cancer cells, the state of mitochondria shows a significant imbalance [ref. 23,ref. 24], which is closely related to the progression of cancer and the formation of drug resistance [ref. 25,ref. 26]. Although cancer cells prefer to undergo glycolysis as the primary pathway of energy metabolism in an aerobic environment, OXPHOS acts as a complementary pathway to provide the cells with an additional source of ATP in certain cancer cells [ref. 27]. In addition, if cancer cells regulate mitochondrial mass through mitophagy, they may develop drug resistance [ref. 28–ref. 30]. Indeed, OXPHOS is also involved in bone development [ref. 31]. Mitophagy is a process that contributes to tubular cell survival and protects and maintains renal function in a mouse model of septic acute kidney injury (AKI) [ref. 32]. Similarly, Jionoside A1-mediated increase in mitophagy attenuates ischemic/reperfusion injury in ischemic stroke by increasing mitochondrial content and ATP [ref. 33]. These studies suggest that targeting the regulation of mitochondrial dynamics and function could be an effective target for ameliorating and treating these diseases.
Mitochondrial Transfer
Delivery of mitochondria from one cell type to another has garnered widespread attention in recent years [ref. 34]. The transfer of mitochondria from donor cells to recipient cells is closely related to rescue respiration, tissue repair, and tumor progression [ref. 35]. The methods of mitochondrial transfer between cells mainly include tunneling nanotubes (TNTs), gap junctions, cell fusion, extracellular vesicles, and free mitochondria release [ref. 36,ref. 37]. Common modes of mitochondrial transfer are outlined in Fig. 2. Among them, TNTs are considered the most extensive method of mitochondrial transfer between cells and are also the current focus of research. TNTs are thought to be primarily composed of filamentous actin (F-actin), forming a membrane-like tubular structure with open ends suspended above the matrix. These structures exhibit diameters ranging from 50 to 1,000 nm and lengths ranging from 10 to 200 μm [ref. 38]. Previous research has revealed 2 types of formation patterns for TNTs [ref. 39]. In one pattern, a single cell or a pair of cells extends processes that then fuse with another cell, forming the nanotube. Another pattern, 2 cells that were initially in contact form a nanotube by moving away from each other in opposite directions. Nevertheless, the precise mechanism underlying the formation of TNTs remains ambiguous. Notably, TNTs can establish connections between distant cells to transport various materials apart from mitochondria, such as lysosomes, calcium ions, and other proteins [ref. 40].
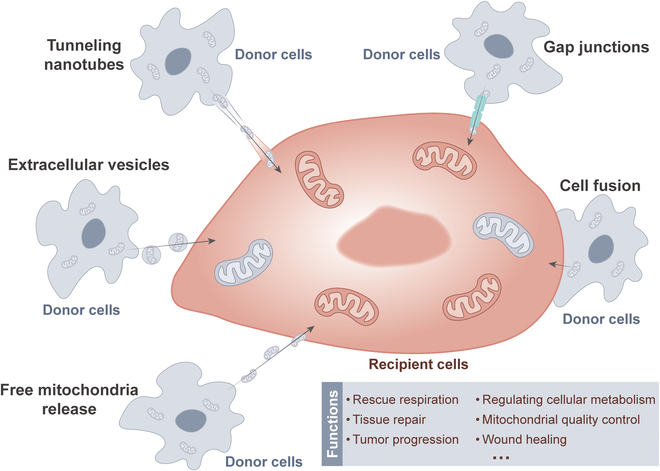
Cell-to-cell mitochondrial transfer via TNTs has been extensively reported to play a role in in vivo and in vitro settings, as well as in a variety of disease states, including cancer [ref. 41–ref. 46]. For instance, when stressed cells in the early stage of apoptosis, such as ultraviolet-treated rat pheochromocytoma cells (PC12), are cocultured with healthy cells, TNTs form between them [ref. 47]. These TNTs facilitate the transfer of functional mitochondria from healthy cells to stressed cells, effectively reversing the early apoptotic process in stressed cells. Similarly, in a coculture model of young and aged granulosa cells (GCs) in humans, the transfer of mitochondria from young to aged GCs via TNTs has been confirmed, improving the mitochondrial bioenergetic function of aged GCs [ref. 48]. Moreover, in the bone marrow microenvironment, acute myeloid leukemia (AML) cells receive mitochondrial transfer from mesenchymal stem cells (MSCs) via TNTs, thereby enhancing resistance to OXPHOS inhibitors [ref. 49].
Donor cells transfer their healthy mitochondria to stimulated or damaged recipient cells through TNTs. This process helps to restore the bioenergetic characteristics of the recipient cells, enhance cell viability, reduce inflammatory processes, and promote the normalization of cell function [ref. 50]. Consistent with this concept, cancer cells use TNTs to acquire immune cell mitochondria, enhance their own metabolism, weaken immunity, and accelerate proliferation and immune evasion [ref. 51]. A recent publication also reported that cancer-associated fibroblasts (CAFs) exchange goods with breast cancer cells through TNTs, wherein the mitochondria of CAFs serve as critical cargo, significantly enhancing the 3-dimensional (3D) migratory capacity of cancer cells [ref. 52]. Interestingly, a recent study reported that low invasive hepatocellular carcinoma (HCC) acquires mitochondria from highly invasive HCC through TNT mechanism, which significantly enhances the migration and invasion capabilities of low invasive HCC [ref. 53]. The acquisition of mitochondria by TNTs is beneficial for cancer cell invasion. This cellular process may also be a novel target for cancer therapy.
In addition to the fact that the transfer of mitochondria through TNTs has been intensively studied in the field of cancer, this phenomenon has received similar reports and attention in other fields of biology and medicine. For instance, a study has reported that in the context of vascular transplantation, MSCs can transport mitochondria to endothelial cells (ECs) via TNTs [ref. 54]. This process is crucial for the successful implantation and functioning of ECs in vivo, and thus offers significant clinical potential. Another study suggests that promoting the formation of TNTs between senescent bone marrow mesenchymal stromal cells (BMSCs) can accelerate the transfer of mitochondria between these cells, thereby restoring mitochondrial function and ultimately significantly enhancing the viability of senescent BMSCs [ref. 55]. According to these studies, we deeply realize that mitochondrial transfer through TNTs plays a pivotal role in biology and medicine, and its far-reaching impact and great potential indicate that this field is worthy of further exploration and research.
Cues from Biochemical and Mechanical Sources Reorganize and Regulate Mitochondrial Networks and Functions
The biochemical and mechanical characteristics of the cellular microenvironment are altered by various diseases, and cells sense these mechanical cues to regulate their morphology, function, and metabolism [ref. 7]. Therapeutic strategies targeting the extracellular microenvironment hold promise for addressing the progression of various diseases, particularly in cancer [ref. 56]. There is growing evidence that mitochondrial dysfunction results from abnormal changes in both the biochemical composition and physical properties of the microenvironment. We will then discuss how these cues, including biochemical cues such as glucose concentration [ref. 9], potential of hydrogen (pH) [ref. 57], and Ca2+ [ref. 58] and mechanical cues such as stiffness [ref. 59], influence cellular activity, especially their role in controlling mitochondrial dynamics and functions (Fig. 3).
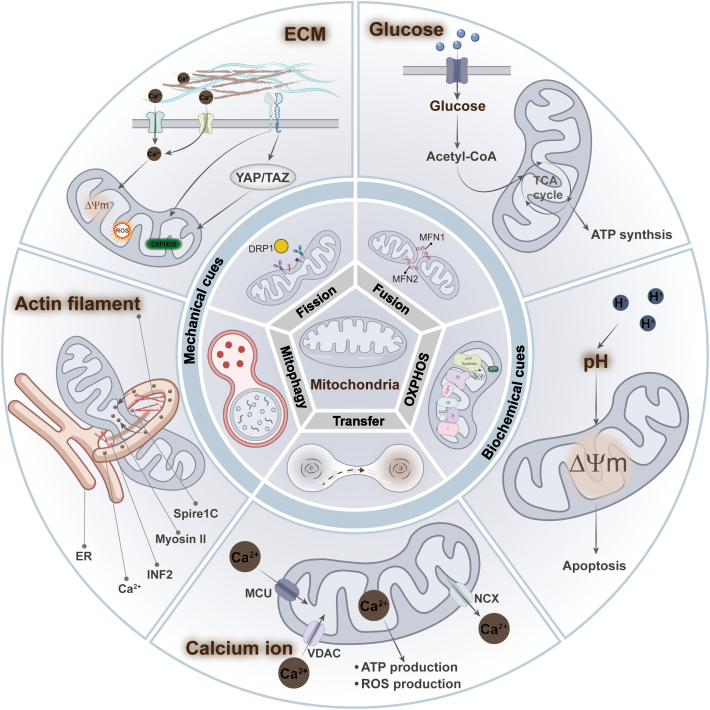
Biochemical cues
High glucose
High glucose, as a biochemical stressor, induces a rapid shift in mitochondrial morphology from filamentous networks to fragmented structures in human mammary epithelial cells. This process result in increased intracellular ROS levels and increased levels of Ca2+ in mitochondria, ultimately triggering oxidative stress [ref. 9]. Mechanistically, this may be due to the high glucose-induced increase in intracellular ROS levels, which inhibits adenosine monophosphate (AMP)-activated protein kinase (AMPK) activity and indirectly promotes Dynamin-related protein 1 (DRP1) activation, leading to mitochondrial division [ref. 60]. Similarly, high glucose-induced impairment of mitochondrial integrity and reduced mitochondrial respiration, as evidenced by short and small mitochondria, cytoplasmic cytochrome c levels, and reduced mitochondrial ATP levels, were observed in primary retinal cells [ref. 61].
pH
The changes in extracellular pH significantly impact the activity of various ion channels and transporter proteins on the cell membrane, which in turn affects cell function [ref. 62]. However, some studies have shown that increasing cellular pH can induce mitochondrial dysfunction-dependent cancer cell death by triggering a series of molecular events. For example, bicarbonate increased intracellular cytoplasmic and mitochondrial pH in liver cancer cells (SK-HEP-1) and induced mitochondrial persistent permeability (MPT) and mitochondrial damage, which resulted in a disruption of OXPHOS. It leads to a significant increase in AMP, which activates classical AMPK-mediated autophagy, leading to cell death [ref. 57]. Exposure to high pH buffers raises the pH of the mitochondrial matrix, inhibiting mitochondrial respiration and increasing ROS production. On the one hand, this may have led to spontaneous transient depolarization of mitochondria to reduce ROS production by suppressing continued elevated matrix pH and protecting cells from damage [ref. 6].
Calcium levels
Notably, a growing number of studies have demonstrated a reciprocal regulatory relationship between Ca2+ and mitochondria. On the one hand, mitochondria are involved in the regulation of intracellular Ca2+ and Ca2+ signaling through their capacity to uptake and store Ca2+ [ref. 58]. Conversely, Ca2+ has been identified as the signature stimulatory signal for the activation of several mitochondrial enzymes that enhance respiratory chain activity [ref. 63]. In the presence of Piezo1, the intracellular concentration of Ca2+ is increased, which leads to an increase in mitochondrial oxygen consumption and ATP levels [ref. 64].
However, when there is an excess of intra-mitochondrial Ca2+, this can result in the opening of the mitochondrial permeability transition pore (mPTP), which leads to mitochondrial fragmentation and increased mitochondrial autophagy [ref. 65,ref. 66]. Additionally, the uptake of calcium ions by mitochondria can result in the production of ROS [ref. 67]. Interestingly, the high uptake of mitochondrial Ca2+ in the above study did not result in elevated ROS. Most studies have suggested that Ca2+-induced ROS elevation is associated with its impact on OXPHOS. These suggest a complex crosstalk between Ca2+ and the ROS pathway, the exact regulatory mechanism of which remains unclear.
Mechanical cues
Mechanical microenvironment of ECM
The mechanical forces provided by the ECM have been largely overlooked and have recently been shown in many studies to play a key role in disease progression [ref. 68]. ECM is not only a matrix for cell attachment and migration but also a dynamic signaling center that regulates various physiological functions of cells by providing mechanical, chemical, and other stimuli. Considerable articles have been published showing the association between extracellular mechanical cues, particularly ECM stiffness, and mitochondrial dynamics and function, especially mitochondrial metabolism (Fig. 4). For example, in human mammary epithelial cells, the morphology of mitochondria gradually changes from a filamentous network to a ring-like fragmented one as the matrix stiffness increases, which leads to increased intracellular ROS levels and decreased ATP levels [ref. 9]. During the spreading of MSCs on the stiff matrix, there is an increase in mitochondrial division and decrease in ATP levels, which increases YAP/TAZ-mediated osteogenic differentiation of MSCs [ref. 69]. Similarly, mitochondrial phosphoenolpyruvate carboxykinase (PCK2) expression increased with increasing matrix stiffness, which activated glycolysis to promote osteogenesis in MSCs [ref. 70]. These findings imply a multifaceted regulatory relationship between ECM stiffness and mitochondrial dynamics and function during the development of bone tissue.
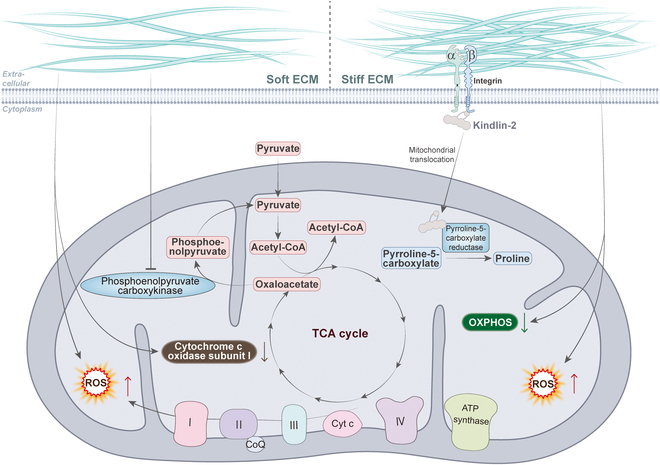
The development of cancer is usually accompanied by stiffening of the tissues. In lung adenocarcinoma, stiffening of the ECM promotes translocation of kindlin-2 to mitochondria and interaction with the enzyme pyrrolidine-5-carboxylate reductase 1 (PYCR1), which induces an increase in proline synthesis and cellular proliferation in tumor cells. This can be reversed by knockdown of kindlin-2, which serves as an effective strategy to inhibit tumor growth [ref. 71]. Similarly, in HCC cells, mitochondria were fragmented and granular in the softer 3D collagen environment, with reduced depolarization of the mitochondrial membrane potential and down-regulation of mitochondria-encoded cytochrome c oxidase I, which inhibited cell proliferation [ref. 72]. Softer matrix stiffness induced an increase in mitochondrial division and intracellular ROS levels in breast cancer cells [ref. 73]. These studies reveal new targets for inhibiting tumor growth and metastasis by regulating mitochondrial dynamics and function in response to ECM stiffness.
Cell mechanics
Mechanical forces acting on cells affect cell morphology, gene expression, and physiological function by regulating cytoskeletal dynamics [ref. 74,ref. 75]. In this process, the cytoskeleton plays a very important role, which forms a stable molecular network structure within the cell to generate mechanical forces to maintain cell morphology. In particular, actin filaments are the main generators of intracellular mechanical forces and are important for maintaining cell morphology and function [ref. 76]. The dynamics of actin filaments have been shown to be involved in a variety of life activities, especially tumor cell proliferation [ref. 77], motility [ref. 78], and mitochondrial division [ref. 79]. An increasing number of studies have demonstrated that the dynamics of the cytoskeleton play a crucial role in regulating mitochondrial morphology. It has been shown that the actin cytoskeleton can be involved in mitochondrial division in several ways. Firstly, by directly affecting mitochondrial division: An advanced technique called platinum replica electron microscopy (PREM) was used to visualize the structure of the cytoskeletal network and mitochondria. The researchers found that actin filaments cross the constrictions of mitochondria that are about to divide [ref. 80]. This particular network structure is associated with mitochondrial division, which may exert a tangential force on the mitochondria to promote their division. Secondly, by indirectly affecting mitochondrial division: Overexpression of the splice isoform of spire type actin nucleation factor 1 (Spire1C), which is specifically localized to mitochondria, promotes actin assembly around mitochondria by directly binding to and stimulating inverted formin 2 [INF2; an actin nucleation factor located on the endoplasmic reticulum (ER)] [ref. 81]. This leads to an increase in mitochondria–ER contact sites and increased mitochondrial division [ref. 82].
Actin works in conjunction with myosin to fulfill its function. Myosin binding and gliding can regulate the dynamics of actin network and influence the remodeling of the cytoskeleton [ref. 83]. Intriguingly, a number of studies have shown that myosin can be indirectly involved in mitochondrial division by interacting with actin [ref. 12]. For example, the mitochondria-localized myosin 19 tethers mitochondria to ER-associated actin to enhance ER contact with mitochondria and promote mitochondrial fission [ref. 84]. In addition, the contractile activity of the non-muscle myosin IIA (NMIIA) leads to stochastic inhomogeneous deformations of the cytoskeletal network. This highly dynamic mechanically active actin filament exerts a localized squeeze on mitochondria, which leads to localized invagination of the mitochondrial surface. This may be a critical point for initiating mitochondrial fission [ref. 80]. In general, actin and myosin play a key role in mitochondrial division and dynamic balance by remodeling the cytoskeletal network, recruiting division proteins, transporting and localizing them, and regulating mitochondrial morphology. This regulation of mitochondrial dynamics is important for cellular energy metabolism, signaling, and other processes.
Mechanotransduction in mitochondrial dynamics and function
Cells sense extracellular mechanical cues through different mechanisms, causing a series of biochemical reactions downstream within cells that ultimately affect various cellular behaviors; these processes are called mechanotransduction. Previously, we introduced that mechanical cues affect intracellular mitochondrial dynamics and function. In the subsequent sections, we will delineate the process of mechanotransduction that ensues when cells are subjected to mechanical stimuli, emphasizing the consequential effects on mitochondrial function and dynamics.
Mechanosensors
Cells respond to mechanical signals through transmembrane proteins localized to the cell membrane or proteins that sense changes in membrane tension caused by these signals. These proteins then activate downstream effector proteins, which ultimately cause spatial and temporal changes in gene expression, thereby regulating cellular function [ref. 85,ref. 86]. For example, cells can sense ECM stiffness through integrins, which are activated to transmit signals into the cell, causing changes in the dynamics of the cytoskeleton [ref. 87]. The cytoskeletal remodeling events in turn affect various cellular functions [ref. 76]. In addition, there are a number of mechanosensitive ion channels on the cell membrane in addition to adhesion receptors, such as Piezo1, a member of the Piezo family of mechanically activated cation channels. Piezo1 senses changes in membrane plasmodesmata caused by mechanical signals and transmits them to the cell, realized as changes in Ca2+ concentration [ref. 88]. This signaling process is involved in the regulation of various cellular physiological activities, such as cell migration, proliferation, and differentiation [ref. 89]. Collectively, these mechanosensitive molecules ultimately regulate cellular physiological activities such as gene expression, cell proliferation, differentiation, and apoptosis by participating in signal transduction pathways.
In addition to its role as an energy factory within the cell, mitochondria also play key roles in a variety of signaling pathways [ref. 4]. For example, mitochondria are capable of taking up and releasing Ca2+, which act as important second messengers in a variety of signaling cascades [ref. 58]. In addition, ROS produced by mitochondrial metabolism can act as signaling molecules to regulate cellular processes [ref. 90]. Mitochondrial metabolites such as nicotinamide adenine dinucleotide (NAD+) and adenosine diphosphate (ADP)/ATP can sense cellular energy status and transmit signals [ref. 91]. Eventually, signaling cascades such as the mitochondrial unfolded protein response are activated when mitochondrial dysfunction occurs [ref. 92]. Overall, mitochondria play an important role in the cellular signaling network and are involved in the regulation of a variety of key physiological processes.
Therefore, it is reasonable to question whether mitochondria are involved in mechanotransduction processes or extracellular mechanical forces modulate mitochondrial function to influence cellular behavior. Certainly, there have been many studies demonstrating that cellular mechanical forces can regulate mitochondrial function through different mechanisms (Fig. 5 and Table).
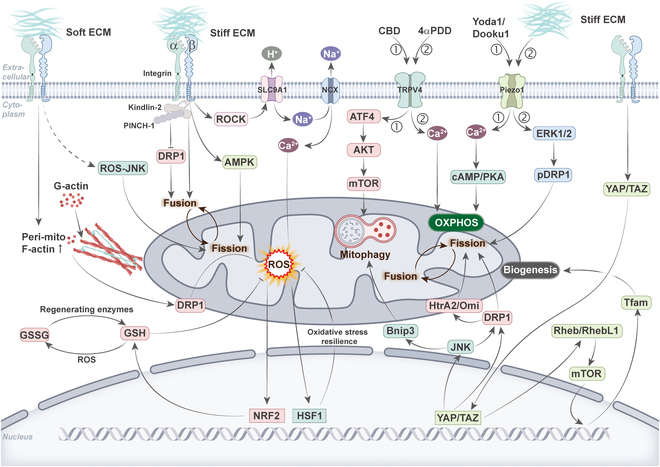
Table.: Summary of the mechanotransduction pathways related to mitochondrial morphology, function, and their impact on cell fate
| Influencing factors | The effect on mitochondria | Mechanisms involved | Results | References |
|---|---|---|---|---|
| Stiff ECM | Fusion | Integrin–kindlin-2/PINCH-1 | Cell spreading area | [ref. 94] |
| Stiff ECM | Fission, ROS↑ | Integrin–Ca2+–HSF1 | Oxidative stress resilience | [ref. 9] |
| Stiff ECM | Fission, OXPHOS | Integrin–AMPK | Osteogenic differentiation | [ref. 69] |
| Soft ECM | Fission | ROS–JNK | Apoptosis | [ref. 101] |
| Soft ECM | Fission, mitophagy | IP3R–Ca2+–pDRP1 | Cell survival | [ref. 66] |
| Soft ECM | Fission, ROS↑ | Integrin–ROS–NRF2– glutathione | Oxidative stress resilience | [ref. 73] |
| Yoda1/Dooku1 | OXPHOS | Piezo1–Ca2+–cAMP/PKA | ATP↑ | [ref. 109] |
| Stiff ECM | Fission | Piezo1–ERK1/2–pDRP1 | Apoptosis | [ref. 110] |
| 4αPDD | OXPHOS | TRPV4–Ca2+ | Osteoblast differentiation | [ref. 112] |
| CBD | Mitophagy | TRPV4–ATF4–AKT–mTOR | Cell death | [ref. 113] |
| Yoda1 | Depolarization | Piezo1–TRPV4–Ca2+ | Cell death | [ref. 115] |
| YAP deficiency | Mitophagy | YAP–JNK–Bnip3 | Cell migration | [ref. 119] |
| TAZ knockout | Biogenesis | TAZ–Rheb/RhebL1–mTOR–Tfam | Impaired exercise ability in mice | [ref. 120] |
| YAP deficiency | Fission | YAP–JNK/DRP1–HtrA2/Omi | Apoptosis | [ref. 121] |
| Stiff ECM | Fission, biogenesis | YAP–DRP1 | Osteogenic differentiation | [ref. 126] |
Integrins
Cell–ECM interactions are primarily facilitated by integrins and transmembrane receptors for ECM proteins. The binding of integrins to ECM proteins recruits a multitude of adhesion proteins, such as talin, kindlin-2, paxillin, integrin-linked kinase (ILK), and PINCH-1 [ref. 93], to cell–ECM adhesion sites where they transmit signals to downstream effectors and influence cellular behavior. A study has demonstrated that alterations in mitochondrial dynamics induced by ECM rigidity in human MSCs are mediated through 2 distinct integrin-dependent signaling pathways: specifically, the up-regulation of mitochondrial fusion via kindlin-2 and the inhibition of DRP1 expression and subsequent mitochondrial fission through PINCH-1 [ref. 94]. High matrix stiffness is thought to activate integrins, which will activate a series of intracellular signaling pathways to regulate physiological processes such as cell proliferation, migration, and survival [ref. 87]. Interestingly, in human mammary epithelial cells, high matrix stiffness activates the integrin–rho-associated protein kinase (ROCK) signaling pathway, which regulates the activity of the H+ transporter protein–solute carrier family 9A1 (SLC9A1) and Na+/Ca2+ exchangers (NCX) at the cell membrane, resulting in mitochondrial Ca2+ loading and ROS production [ref. 95,ref. 96]. Meanwhile, activation of heat shock factor 1 (HSF1) and its target gene expression in a heat stress-independent manner conferred cellular resistance to oxidative stress by limiting mitochondrial respiration and causing mitochondrial reprogramming [ref. 97]. The dynamics of cell spreading is the first step in the interaction between the cell and the ECM, which is based on the interaction of integrins with the ECM [ref. 98]. Research has shown that MSCs have different mitochondrial morphology and different ATP levels during cell spreading on different matrix stiffness. On stiffer substrates, the cells spread over a larger area, the intracellular mitochondria were in a divided state, and the total ATP level was higher due to the activation of AMPK by the stiff substrate, which in turn regulates mitochondrial function [ref. 69].
Tumor progression is accompanied by a continuous hardening of the ECM, while cancer cells usually have more mitochondria to meet the demands of their rapid proliferation and energy metabolism [ref. 11,ref. 99,ref. 100]. However, when tumors metastasize to other tissues, they encounter a softer ECM microenvironment again. A study showed that soft substrates activated DRP1-mediated mitochondrial fission with polymerization of actin around mitochondria in breast cancer cells, resulting in elevated levels of mitochondrial ROS and inducing intracellular oxidative stress. In response to the stress, the cells activated nuclear factor erythroid 2-related factor 2 (NRF2)-mediated expression of genes related to the antioxidant stress response, increasing cystine uptake and glutathione metabolism levels, which, in turn, conferred resistance to ROS-dependent drugs, thereby promoting cell survival [ref. 73]. Similarly, soft matrix activated the ROS–JNK (c-Jun N-terminal kinase) signaling pathway in breast cancer cells, which induced apoptosis through the mitochondrial pathway. Interestingly, soft matrix simultaneously induced intracellular protective autophagy, which inhibited apoptosis to promote cell survival [ref. 101]. However, this article does not mention integrins, and their upstream mechanisms remain as in need of further study. Significantly, the same conditions, such as a softer ECM, which promoted mitochondrial division in breast cancer cells, had the opposite result in normal cells. This may be influenced by the distinctive metabolic characteristics of cancer cells and the tumor microenvironment. Further investigation is required to validate this finding in other cancer cells.
Transverse aortic constriction (TAC) is one of the most commonly used disease models of chronic ventricular hypertrophy and can be used to mimic hypertrophic cardiomyopathy caused by hypertension or increased intraventricular pressure [ref. 102]. Melusin is a chaperone protein that is selectively expressed in the heart and binds to the cytoplasmic region of β1 integrins to transduce mechanical signals [ref. 103]. It was found that in a mouse model of TAC, melusin enters mitochondria to bind and inhibit mitochondrial trifunctional protein (MTP), inhibit fatty acid oxidation (FAO), and inhibit ROS production, which protects mice from TAC-induced pressure overload [ref. 104]. Mechanistically, this may be related to the mechanotransduction induced by changes in blood flow shear caused by TAC.
Ion channels: Piezo1 and TRPV4
Piezo1 and transient receptor potential cation channel subfamily V member 4 (TRPV4) have been demonstrated to promote extracellular Ca2+ influx and activate intracellular Ca2+-mediated signaling pathways, which in turn affect cellular metabolism and function [ref. 105,ref. 106]. Under pathological conditions such as hypertension and vascular occlusion, ECs respond to changes in blood flow shear via Piezo1/TRPV4, which triggers an increase in intracellular Ca2+ concentration, which in turn regulates processes such as angiogenesis and vasodilation. In addition, Piezo1/TRPV4 has been associated with malignant angiogenesis in tumors [ref. 107,ref. 108]. Due to the significant interconnection between Ca2+ and mitochondria, we aimed to elucidate the relationship between these ion channels and mitochondrial dynamics and function.
The specific Piezo1 channel activator Yoda1 or its analog Dooku1 has been observed to increase the concentration of Ca2+ in human umbilical vein ECs (HUVECs) and in mitochondria [ref. 64]. This ultimately enhances mitochondrial respiration and glycolysis, stimulating ATP production. Similarly, another study has demonstrated that the activation of Piezo1 results in the uptake of mitochondrial calcium and the subsequent induction of OXPHOS. A more comprehensive investigation revealed that Piezo1 enhances mitochondrial functionality by elevating cAMP signaling [ref. 109]. A study has demonstrated that the presence of a stiff matrix stimulates the activation of the Piezo1 channel, which in turn facilitates the influx of calcium ions. This initiates a cascade of events involving the activation of extracellular signal-regulated kinases 1 and 2 (ERK1/2) and the phosphorylation of DRP1, ultimately leading to the fragmentation of mitochondria and the subsequent induction of apoptosis [ref. 110].
Similar to Piezo1, TRPV4 has been the subject of extensive study as a mechanosensitive ion channel, particularly in the context of osteoblast research [ref. 111]. In osteoblasts, the activation of TRPV4 has been observed to increase intracellular as well as mitochondrial Ca2+ levels, and promote basal and maximal mitochondrial respiration and ATP production [ref. 112]. This has been demonstrated to facilitate osteoblast differentiation. Moreover, TRPV4 activation has been demonstrated to regulate mitochondrial morphology, although the precise mechanisms involved remain uncertain. Another study indicated the non-psychoactive phytocannabinoid cannabidiol (CBD), a drug used to treat gliomas, possesses a mechanism of action that induces mitochondrial autophagy through the activation of TRPV4, which ultimately results in cell death [ref. 113]. Mechanistically, the ATF4 (activating transcription factor 4)–DDIT3 (DNA damage inducible transcript 3)–TRIB3 (tribbles pseudokinase 3)–AKT (serine-threonine protein kinase )–mTOR (mammalian target of rapamycin) pathway [ref. 114], which is activated by TRPV4, plays a crucial role in CBD-induced mitochondrial autophagy. It is noteworthy that in pancreatic acinar cells, Piezo1 activation resulted in a transient elevation of intracellular calcium levels, whereas Piezo1 activation-induced opening of TRPV4 channels led to a prolonged elevation of intracellular calcium levels, mitochondrial depolarization, intracellular trypsin activation, and cell death [ref. 115]. These studies have identified Ca2+ signaling pathway in which Piezo1 and TRPV4 act in concert.
YAP and TAZ
YAP and TAZ have attracted intense interest due to their remarkable biological properties in cell differentiation, tissue development, and cancer progression [ref. 116,ref. 117]. Moreover, the activities of YAP and TAZ as mechanosensitive transcription factors are directly regulated by ECM stiffness, cell shape, and cytoskeletal tension. To illustrate, when cells are cultured on a stiffer ECM, YAP and TAZ exhibit increased nuclear localization and transcriptional activity, whereas on a softer ECM they are repressed and relocated to the cytoplasm [ref. 118]. In recent years, a number of studies have demonstrated that YAP/TAZ activity is related to mitochondrial function. For example, deletion of YAP results in inhibition of HCC migration. The mechanism is that knockdown of YAP induces phosphorylation of JNK, which binds to the Bnip3 promoter and promotes Bnip3 expression. Higher Bnip3 activity triggers excessive mitochondrial autophagy, leading to mitochondrial dysfunction and ATP shortage [ref. 119]. Similarly, specific knockout of TAZ reduced mitochondrial biogenesis, respiratory metabolism, and locomotor activity in mice, which is related to the Ras homolog enriched in brain (Rheb)/Rheb like 1 (RhebL1)–mTOR–mitochondrial transcription factor A (Tfam) pathway [ref. 120]. Furthermore, YAP/TAZ plays a role in the regulation of mitochondrial dynamics. To illustrate, the silencing of YAP has been observed to result in JNK phosphorylation, which in turn induces DRP1 activation and translocation to the mitochondrial surface, thereby initiating mitochondrial fission. Excessive mitochondrial fission has been demonstrated to mediate the leakage of high-human temperature requirement A member (HtrA2/Omi) from mitochondria into the cytoplasm [ref. 121]. This results in the triggering of apoptosis in human rectal cancer cells via the mitochondrial apoptotic pathway. On the contrary, another study indicated the down-regulation of YAP was observed to inhibit myofibroblast differentiation and result in a reduction of DRP1 levels. This, in turn, led to the elongation of mitochondria, fusion of mitochondrial network, and collapse of the mitochondrial membrane potential [ref. 122].
Moreover, studies have demonstrated that YAP/TAZ directly regulates mitochondrial function in response to mechanical stimuli [ref. 123]. The transfer of phosphate to ADP, catalyzed by cytoplasmic creatine kinase type B (CKB), enables the regeneration of ATP, thereby maintaining the local ATP gradient [ref. 124]. In a stiff environment, pancreatic ductal adenocarcinoma (PDAC) cells not only support ATP production by increasing mitochondrial activity and OXPHOS but also participate in the phosphocreatine-dependent ATP cycling mechanism by expressing CKB via the integrin–YAP pathway [ref. 125]. Furthermore, it has been demonstrated that in the absence of a mineralization-inducing medium, MSCs can respond to alterations in ECM stiffness by modulating mitochondrial dynamics and biogenesis for osteogenic differentiation through the regulation of YAP activity. In the stiff substrate, YAP activation inhibits DRP1 activity and enhances Tfam-dependent mitochondrial biosynthesis, resulting in an elongated and fused mitochondrial network [ref. 126].
Intracellular mechanotransduction
In the previous section, we have learned that cytoskeleton remodeling plays a key role in the regulation of mitochondrial dynamics. This suggests that the cytoskeleton may play a major role in the influence of extracellular mechanical signals on mitochondrial dynamics and function. In the following sections, we will describe how the cytoskeleton responds to mechanical signaling to affect mitochondrial dynamics and function (Fig. 6).
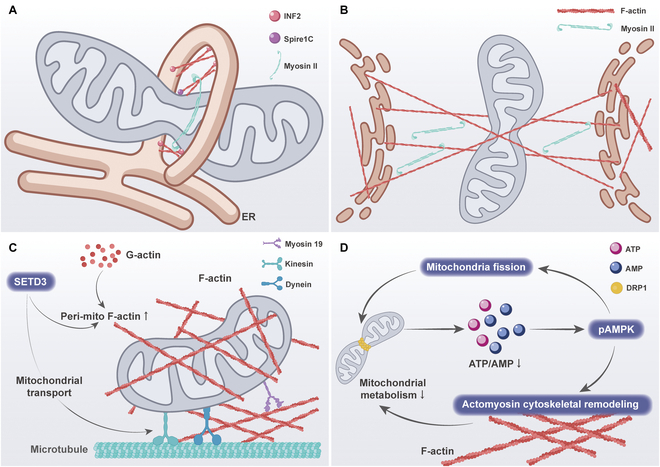
SET domain protein 3 (SETD3) is a mechanosensitive enzyme that is localized on the outer mitochondrial membrane and promotes actin polymerization around mitochondria [ref. 127]. Loss of SETD3 function resulted in a reduction in peri-mitochondrial F-actin, as well as a reduction in mitochondrial branching length, the number of branches, and mitochondrial movement [ref. 128]. These findings provide new insights into the mechanism of peri-mitochondrial F-actin polymerization. Similarly, another study demonstrated that when breast cancer cells are cultured on soft substrates, it is observed that F-actin accumulates around mitochondria in the presence of the actin-related protein 2/3 (Arp2/3) complex and the Spire1C protein, which favors the DRP1-regulated mitochondrial division [ref. 83]. Interestingly, further inhibition of ROCK and myosin light chain kinase (MLCK) activity increased the contact of F-actin with mitochondria [ref. 73]. This suggests that there may be crosstalk based on different F-actin pools.
Furthermore, as previously stated, AMPK is capable of detecting alterations in the stiffness of the ECM, thereby regulating mitochondrial morphology. There are 2 modes of cell migration across the matrix: elongated mesenchymal and rounded-amoeboid modes of movement. The different migration patterns are related to the levels of intracellular metabolism and actomyosin skeleton activity. One study showed lower levels of adhesion to the substrate and lower levels of ATP within rounded-amoeboid cells, which activated pAMPK [ref. 129]. Activation of AMPK results in the direct phosphorylation and inactivation of myosin phosphatase, which in turn leads to increased phosphorylation of myosin light chains and increased overall myosin II activity. These determine the pattern of cell movement and the level of adhesion to the substrate. Concurrently, AMPK preserves the balance in energy levels by inducing mitochondrial fission, which further promotes AMPK signaling and myosin II activation. These studies demonstrate that mitochondrial dynamics can indirectly regulate the cytoskeleton by altering energy levels.
The actin skeleton plays a role in the regulation of mitochondrial dynamics, as well as in the processes of mitochondrial transport and localization. Integrin-based mechanosensing of ECM stiffness has been observed to result in the formation of diffuse actin structures in cells exposed to soft ECM, while cells on hard ECM exhibit prominent actin stress fibers [ref. 69]. This phenomenon has been linked to increased mitochondrial motility and reduced perinuclear localization of mitochondria in comparison to those observed in cells on soft ECM [ref. 130].
Motor-driven mitochondrial transfer along the cytoskeleton
To explore the mechanical signals involved in the transfer of mitochondria between cells via TNTs, the research has concentrated on the biomechanical changes occurring within cells. Therefore, a deeper understanding of the composition, structure, and formation mechanism of TNTs is essential for elucidating the mechanical signal transmission during this process.
Mitochondrial transport in TNTs depends on the action of dynein and kinesin motor complexes that move along the cytoskeleton, such as F-actin and microtubules (Fig. 7). In particular, these motor complexes contain adaptor proteins such as Mitochondrial Rho guanosine triphosphatases (GTPases) 1 and 2 (Miro1 and Miro2), which are tightly anchored to the mitochondrial outer membrane and effectively facilitate unidirectional or bidirectional mitochondrial movement on the cytoskeleton within the TNTs [ref. 36,ref. 131]. Intriguingly, a recent study showed that mitochondrial precursor proteins accumulated in the cytoplasm did not migrate spontaneously in TNTs, indicating that mitochondrial transport through TNTs depends on an active mechanism rather than a simple passive diffusion [ref. 132].
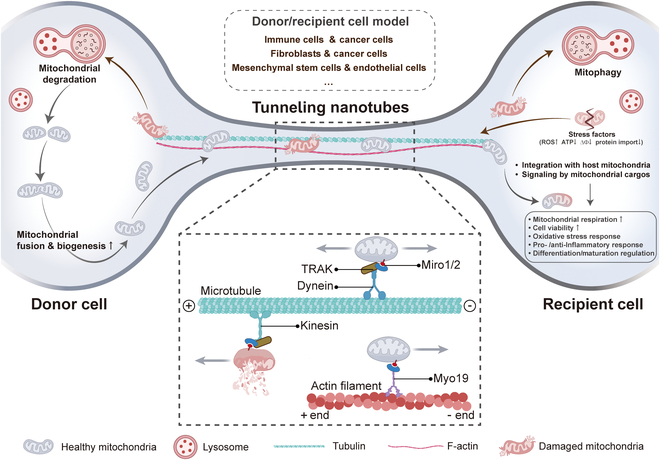
Mitochondria exhibit 2 modes of intracellular movement: one is long-distance transport driven by microtubule mechanisms, and the other is short-distance actin-based transport [ref. 133]. Specifically, mitochondrial transport through TNTs is mediated by mitochondrial Miro1, allowing it to move along the cytoskeleton within TNTs. Mechanically, Miro1 acts synergistically with the auxiliary proteins Miro2, transport-associated protein 1 and 2 (TRAK1 and TRAK2), and myosin 19 (Myo19) to drive mitochondrial binding to kinesin family motor protein 5 (KIF5)-driven proteins [ref. 134]. When combined, they form a motor adaptor complex that facilitates the transport of mitochondria within the TNTs along the cytoskeleton and regulates their motility.
The mechanism of mitochondrial transport within TNTs is not exclusively reliant on a singular cytoskeletal element [ref. 135]. Consequently, in F-actin-only TNTs, mitochondria are predominantly transported along actin filament. In contrast, in TNTs that incorporates both F-actin and microtubules, mitochondrial transport may exhibit greater flexibility, potentially utilizing either F-actin or microtubules—or even both simultaneously. This observation further highlights the intricate complexity and adaptability inherent to mitochondrial transport within TNTs. If mitochondria are transported along F-actin within TNTs, it is critical that Miro proteins recruit and stabilize Myo19 on mitochondria, as Myo19 is a key molecule linking mitochondria to the actin cytoskeleton. Myo19 directly interacts with Miro to participate in mitochondrial transport without the involvement of other complexes. By contrast, Miro cooperates with KIF5 and TRAK to form a complex during mitochondrial transport along TNT microtubules [ref. 136]. The complex effectively drives mitochondrial movement along the microtubules of TNTs under the regulation of KIF5. For instance, between cardiomyocytes and cardiac fibroblasts, mitochondria are efficiently transported along microtubules by TNTs using KIF5 [ref. 46].
Mitochondrial movement mediated by kinesin and dynein motors on microtubules enables efficient bidirectional movement along microtubules [ref. 137]. The KIF family is responsible for the directed transport of mitochondria along the plus end of microtubules, while dynein drives mitochondria along the minus end of microtubules [ref. 138]. The adaptor proteins Milton and Miro interact to become a key bridge connecting mitochondria to motor proteins, regulating the bidirectional movement of mitochondria on microtubules [ref. 139]. Through the tight association of Miro with Milton, motor proteins, particularly kinesin-1, are efficiently recruited to mitochondria, leading to efficient mitochondrial transport along microtubules. Furthermore, Miro1 is required by TRAK2 for minus-end directed mitochondrial transport in fibroblasts. However, in the presence of overexpression of KIF5, TRAK2 regulates plus-end directed transport of mitochondria even in the absence of Miro1. Remarkably, TRAK1 and TRAK2 were still able to localize to mitochondria even in the absence of Miro1 and Miro2 [ref. 136]. This suggests that their binding to mitochondria may not be entirely dependent on Miro proteins or that other unknown interaction mechanisms exist.
It is important to note that we have identified an unusual member of the myosin family, human Myo19. This protein is unique in that it is specifically anchored to mitochondria through its tail domain, but is not directly inserted into the mitochondrial outer membrane [ref. 140]. In contrast, human Myo19 was shown to bind tightly to the Miro GTPase, and this association constitutes an important bridge between the myosin motor and mitochondria, thereby enabling their coordination and connection [ref. 136]. Consistent with these findings, a study reported that Myo19 dimers exhibit efficient transport ability on actin tracks in the cell. These findings not only reveal the intracellular motility properties of Myo19 molecules but also provide strong evidence that Myo19 molecules are able to transport mitochondria directly on actin tracks and are essential for maintaining cellular function and homeostasis. In addition, the importance of actin dynamics in mitochondrial transport has been illustrated in numerous review articles [ref. 135,ref. 139]. Although the process of mitochondrial movement along actin filaments has been widely described, there are still many unanswered questions in this complex process that need to be further explored.
Conclusion and Prospects
Disruption of mitochondrial function leads to abnormal cell fate and a range of diseases. In this review, we summarize the roles of mitochondrial dynamics and function in diseases such as metabolic diseases, CVDs, neurodegenerative diseases, and cancer, including mitochondrial fusion and fission, mitochondrial autophagy and transport, and mitochondrial function, and describe the roles played by a series of key proteins that provide new insights into targeting mitochondria to regulate disease.
In addition, mitochondria are highly plastic and dynamic organelles that regulate their morphology and function in response to a variety of stimuli both inside and outside the cells [ref. 5]. In this review, we summarize the effects of intra- and extracellular biochemical and mechanical cues on mitochondrial dynamics, function, and transfer, and highlight the specific molecular mechanisms by which intra- and extracellular mechanical cues regulate mitochondrial morphology and function and transfer. Alterations in these mechanical signals arise with diseases such as aging, fibrosis, and cancer. Altered mechanical signals can in turn be transmitted to mitochondria via intra- and extracellular mechanotransduction molecules, thereby altering their morphology and function, ultimately leading to malignant cellular behaviors. Therefore, studying the results of tissue and cell mechanics associated with common diseases may be a new way to treat them.
Although we have identified many mechano-regulated phenomena of mitochondrial dynamics and function, the underlying mechanotransduction mechanisms remain to be refined. Ca2+ are usually thought to enter mitochondria through the mitochondrial calcium uniporter (MCU) on the mitochondrial membrane and regulate its function [ref. 141]. Recent studies have shown that connexins are also localized to mitochondria, particularly in the heart, where they regulate mitochondrial ion homeostasis (K+, Ca2+) by forming mitochondrial hemichannels [ref. 142]. Whether mitochondrial connexin 43 responds to Piezo1/TRPV4 remains to be investigated. In addition, most of the experiments did not investigate the key mechanosensitive proteins and specific signaling pathways, while others were limited to cell lines and experimental conditions, and the conclusions obtained could not be applied to other cell lines. For example, normal cells and tumor cells show different mitochondrial morphology and function under similar matrix stiffness conditions, and the specific regulatory substrates behind this need to be investigated. However, what we can affirm is the indispensable and important role of mitochondrial dynamics in mechanotransduction [ref. 123].
In addition, most in vitro studies are established in 2D conditions, whereas in vivo cells are in more complex environments and are subjected to multifaceted mechanical stimuli; thus, further optimization of in vitro models to reflect the true state of the cells under physiological or pathological conditions is needed in the future. For example, in vivo cells often move in irregular channels, and we would like to understand how the mechanical stimuli to which the cells are subjected change during such complex movements, and how the cells regulate mitochondrial dynamics and function to adapt to the environmental stimuli. Therefore, we need to establish a reasonable and perfect in vitro model for the study. Moreover, when performing in vivo experiments, we hope to develop an in vivo real-time mitochondrial tracer model in the future to replace the traditional immunohistochemistry to better observe the changes of mitochondrial dynamics. Last but most importantly, as our research continues, we need to consider how to turn these mechanosignaling molecules into powerful targets for the treatment of various diseases.
References
- Mitochondria as cellular and organismal signaling hubs.. Annu Rev Cell Dev Biol., 2022. [PubMed]
- Mitochondrial dynamics and mitochondrial quality control.. Redox Biol., 2015. [PubMed]
- Active control of mitochondrial network morphology by metabolism-driven redox state.. Proc Natl Acad Sci USA., 2025. [PubMed]
- Mitochondrial dynamics and its involvement in disease.. Annu Rev Pathol., 2020. [PubMed]
- Mitochondrial dynamics in health and disease: Mechanisms and potential targets.. Signal Transduct Target Ther., 2023. [PubMed]
- Effects of matrix pH on spontaneous transient depolarization and reactive oxygen species production in mitochondria.. Front Cell Dev Biol., 2021. [PubMed]
- How the ageing microenvironment influences tumour progression.. Nat Rev Cancer., 2020. [PubMed]
- High glucose-induced endothelial progenitor cell dysfunction.. Diab Vasc Dis Res., 2017. [PubMed]
- Adhesion-mediated mechanosignaling forces mitohormesis.. Cell Metab., 2021. [PubMed]
- Tuning immunity through tissue mechanotransduction.. Nat Rev Immunol., 2023. [PubMed]
- Matrix stiffness contributes to cancer progression by regulating transcription factors.. Cancers (Basel)., 2022. [PubMed]
- The multiple links between actin and mitochondria.. Nat Rev Mol Cell Biol., 2023. [PubMed]
- Mitochondrial dynamics, diabetes, and cardiovascular disease.. Diabetes., 2024. [PubMed]
- Caveolin-1 controls mitochondrial damage and ROS production by regulating fission-fusion dynamics and mitophagy.. Redox Biol., 2022. [PubMed]
- Mitochondria in disease: Changes in shapes and dynamics.. Trends Biochem Sci., 2024. [PubMed]
- Mitochondrial dynamics and psychiatric disorders: The missing link.. Neurosci Biobehav Rev., 2024. [PubMed]
- Impaired mitochondrial dynamics in the blood of patients with Alzheimer’s disease and Lewy body dementia.. J Alzheimer’s Assoc., 2023
- Huntington’s disease affects mitochondrial network dynamics predisposing to pathogenic mitochondrial DNA mutations.. Brain., 2024. [PubMed]
- Changes in Drp1 function and mitochondrial morphology are associated with the alpha-synuclein pathology in a transgenic mouse model of Parkinson’s disease.. Cells., 2021. [PubMed]
- Mitophagy and cGAS-STING crosstalk in neuroinflammation.. Acta Pharm Sin B., 2024. [PubMed]
- Impaired mitochondrial oxidative phosphorylation capacity in epicardial adipose tissue is associated with decreased concentration of adiponectin and severity of coronary atherosclerosis.. Sci Rep., 2019. [PubMed]
- Direct cardiac actions of the sodium glucose co-transporter 2 inhibitor empagliflozin improve myocardial oxidative phosphorylation and attenuate pressure-overload heart failure.. J Am Heart Assoc., 2021. [PubMed]
- Mitochondrial dynamics and colorectal cancer biology: Mechanisms and potential targets.. Cell Commun Signal., 2024. [PubMed]
- MTFR2-dependent mitochondrial fission promotes HCC progression.. J Transl Med., 2024. [PubMed]
- Drp1 splice variants regulate ovarian cancer mitochondrial dynamics and tumor progression.. EMBO Rep., 2024. [PubMed]
- Caveolin-1 signaling-driven mitochondrial fission and cytoskeleton remodeling promotes breast cancer migration.. Int J Biochem Cell Biol., 2022. [PubMed]
- Oxidative phosphorylation as an emerging target in cancer therapy.. Clin Cancer Res., 2018. [PubMed]
- p52-ZER6/IGF1R axis maintains cancer stem cell population to promote cancer progression by enhancing pro-survival mitophagy.. Oncogene., 2024. [PubMed]
- USP26 promotes colorectal cancer tumorigenesis by restraining PRKN-mediated mitophagy.. Oncogene., 2024. [PubMed]
- Fighting against drug-resistant tumor by the induction of excessive mitophagy with transferrin nanomedicine.. Macromol Biosci., 2024. [PubMed]
- Impact of the SIK3 pathway inhibition on osteoclast differentiation via oxidative phosphorylation.. J Bone Miner Res., 2024. [PubMed]
- The PINK1/PARK2/optineurin pathway of mitophagy is activated for protection in septic acute kidney injury.. Redox Biol., 2021. [PubMed]
- Jionoside A1 alleviates ischemic stroke ischemia/reperfusion injury by promoting Nix-mediated mitophagy.. Cell Mol Biol., 2023
- The power and potential of mitochondria transfer.. Nature., 2023. [PubMed]
- Intercellular mitochondrial transfer as a means of tissue revitalization.. Signal Transduct Target Ther., 2021. [PubMed]
- Mitochondria on the move: Horizontal mitochondrial transfer in disease and health.. J Cell Biol., 2023. [PubMed]
- Targeting mitochondrial transfer as a promising therapeutic strategy.. Trends Mol Med., 2025
- Tunneling nanotubes: The fuel of tumor progression?. Trends Cancer., 2020. [PubMed]
- Tunneling nanotubes and related structures: Molecular mechanisms of formation and function.. Biochem J., 2021. [PubMed]
- Immune cell connection by tunneling nanotubes: The impact of intercellular cross-talk on the immune response and its therapeutic applications.. Mol Pharm., 2021. [PubMed]
- Miro1 enhances mitochondria transfer from multipotent mesenchymal stem cells (MMSC) to neural cells and improves the efficacy of cell recovery.. Molecules., 2018. [PubMed]
- Cancer-associated fibroblasts promote prostate cancer malignancy via metabolic rewiring and mitochondrial transfer.. Oncogene., 2019. [PubMed]
- Tunneling nanotubes promote intercellular mitochondria transfer followed by increased invasiveness in bladder cancer cells.. Oncotarget., 2017. [PubMed]
- NADPH oxidase-2 derived superoxide drives mitochondrial transfer from bone marrow stromal cells to leukemic blasts.. Blood., 2017. [PubMed]
- Control of long-distance cell-to-cell communication and autophagosome transfer in squamous cell carcinoma via tunneling nanotubes.. Oncotarget., 2017. [PubMed]
- Mitochondria are transported along microtubules in membrane nanotubes to rescue distressed cardiomyocytes from apoptosis.. Cell Death Dis., 2018. [PubMed]
- Transfer of mitochondria via tunneling nanotubes rescues apoptotic PC12 cells.. Cell Death Differ., 2015. [PubMed]
- Enhancement of human granulosa cells self-rescue by mitochondrial transfer via tunneling nanotubes.. Fertil Steril., 2020
- Oxphos inhibition induces formation of tunneling nanotubes in AML cells and facilitates mitochondrial transfer from BM stroma to AML that contributes to microenvironment-mediated drug-resistance of AML.. Blood., 2019. [PubMed]
- Horizontal mitochondrial transfer as a novel bioenergetic tool for mesenchymal stromal/stem cells: Molecular mechanisms and therapeutic potential in a variety of diseases.. J Transl Med., 2024. [PubMed]
- Intercellular nanotubes mediate mitochondrial trafficking between cancer and immune cells.. Nat Nanotechnol., 2022. [PubMed]
- Mitochondrial transfer from cancer-associated fibroblasts increases migration in aggressive breast cancer.. J Cell Sci., 2023. [PubMed]
- HMGB1 promotes mitochondrial transfer between hepatocellular carcinoma cells through RHOT1 and RAC1 under hypoxia.. Cell Death Dis., 2024. [PubMed]
- Mitochondrial transfer mediates endothelial cell engraftment through mitophagy.. Nature., 2024. [PubMed]
- Melatonin-loaded bioactive microspheres accelerate aged bone regeneration by formation of tunneling nanotubes to enhance mitochondrial transfer.. Mater Today Bio., 2024
- Tumor microenvironment-specific chemical internalization for enhanced gene therapy of metastatic breast cancer.. Research., 2021. [PubMed]
- Alkalization of cellular pH leads to cancer cell death by disrupting autophagy and mitochondrial function.. Oncogene., 2022. [PubMed]
- Mitochondrial Ca2+ signaling.. Pharmacol Ther., 2018. [PubMed]
- Matrix stiffness regulates osteoclast fate through integrin-dependent mechanotransduction.. Bioact Mater., 2023. [PubMed]
- Empagliflozin improves diabetic renal tubular injury by alleviating mitochondrial fission via AMPK/SP1/PGAM5 pathway.. Metabolism., 2020. [PubMed]
- Resolvin D1 reduces mitochondrial damage to photoreceptors of primary retinal cells exposed to high glucose.. J Cell Physiol., 2020. [PubMed]
- The volume-regulated anion channel (LRRC8) in nodose neurons is sensitive to acidic pH.. JCI Insight., 2017. [PubMed]
- Beyond the TCA cycle: New insights into mitochondrial calcium regulation of oxidative phosphorylation.. Biochem Soc Trans., 2023. [PubMed]
- Piezo1 channel activation stimulates ATP production through enhancing mitochondrial respiration and glycolysis in vascular endothelial cells.. Br J Pharmacol., 2023. [PubMed]
- Cancer cells with defective oxidative phosphorylation require endoplasmic reticulum-to-mitochondria Ca(2+) transfer for survival.. Sci Signal., 2020. [PubMed]
- Endoplasmic reticulum-mitochondrial calcium transport contributes to soft extracellular matrix-triggered mitochondrial dynamics and mitophagy in breast carcinoma cells.. Acta Biomater., 2023. [PubMed]
- Crosstalk signaling between mitochondrial Ca2+ and ROS.. Front Biosci., 2009
- Extracellular matrix and its therapeutic potential for cancer treatment.. Signal Transduct Target Ther., 2021. [PubMed]
- Energy expenditure during cell spreading influences the cellular response to matrix stiffness.. Biomaterials., 2021. [PubMed]
- The PCK2-glycolysis axis assists three-dimensional-stiffness maintaining stem cell osteogenesis.. Bioact Mater., 2022. [PubMed]
- Kindlin-2 links mechano-environment to proline synthesis and tumor growth.. Nat Commun., 2019. [PubMed]
- Mechanical regulation of mitochondrial dynamics and function in a 3D-engineered liver tumor microenvironment.. ACS Biomater Sci Eng., 2023. [PubMed]
- Mitochondrial fission links ECM mechanotransduction to metabolic redox homeostasis and metastatic chemotherapy resistance.. Nat Cell Biol., 2022. [PubMed]
- The matrix in cancer.. Nat Rev Cancer., 2021. [PubMed]
- Microtubule assists actomyosin to regulate cell nuclear mechanics and chromatin accessibility.. Research., 2023. [PubMed]
- Biochemical and mechanical regulation of actin dynamics.. Nat Rev Mol Cell Biol., 2022. [PubMed]
- The mechanics of tumor cells dictate malignancy via cytoskeleton-mediated APC/Wnt/beta-catenin signaling.. Research., 2023. [PubMed]
- Actin dynamics in cell migration.. Essays Biochem., 2019. [PubMed]
- The interplay between cell-extracellular matrix interaction and mitochondria dynamics in cancer.. Cancers (Basel)., 2022. [PubMed]
- Ultrastructure and dynamics of the actin-myosin II cytoskeleton during mitochondrial fission.. Nat Cell Biol., 2019. [PubMed]
- An actin-dependent step in mitochondrial fission mediated by the ER-associated formin INF2.. Science., 2013. [PubMed]
- A mitochondria-anchored isoform of the actin-nucleating spire protein regulates mitochondrial division.. eLife., 2015. [PubMed]
- Actin structure and function.. Annu Rev Biophys., 2011. [PubMed]
- Myo19 tethers mitochondria to endoplasmic reticulum-associated actin to promote mitochondrial fission.. J Cell Sci., 2023. [PubMed]
- Interplay between mechanics and signalling in regulating cell fate.. Nat Rev Mol Cell Bio., 2022. [PubMed]
- Mechanical regulation of chromatin and transcription.. Nat Rev Genet., 2022. [PubMed]
- Targeting integrin pathways: Mechanisms and advances in therapy.. Signal Transduct Target Ther., 2023. [PubMed]
- Mechanosensing by Piezo1 and its implications for physiology and various pathologies.. Biol Rev Camb Philos Soc., 2022. [PubMed]
- Calcium signaling.. Cell., 2007. [PubMed]
- Mitochondrial ROS signaling in organismal homeostasis.. Cell., 2015. [PubMed]
- NAD+ and sirtuins in aging and disease.. Trends Cell Biol., 2014. [PubMed]
- The mitochondrial unfolded protein response (UPR(mt)): Shielding against toxicity to mitochondria in cancer.. J Hematol Oncol., 2022. [PubMed]
- Chapter 22: Structural and signaling functions of integrins.. Biochim Biophys Acta., 2020
- Extracellular matrix stiffness regulates mitochondrial dynamics through PINCH-1- and kindlin-2-mediated signalling.. Curr Res Cell Biol., 2021
- Elevated Na/H exchanger 1 (SLC9A1) emerges as a marker for tumorigenesis and prognosis in gliomas.. J Exp Clin Cancer Res., 2018. [PubMed]
- Structure-based function and regulation of NCX variants: Updates and challenges.. Int J Mol Sci., 2022. [PubMed]
- Targeting HSF1 for cancer treatment: Mechanisms and inhibitor development.. Theranostics., 2023. [PubMed]
- The universal dynamics of cell spreading.. Curr Biol., 2007. [PubMed]
- Extracellular cell matrix stiffness-driven drug resistance of breast cancer cells via EGFR activation.. Mechanobiol Med., 2023. [PubMed]
- Therapeutic potential of targeting mitochondrial dynamics in cancer.. Biochem Pharmacol., 2020. [PubMed]
- Protective autophagy attenuates soft substrate-induced apoptosis through ROS/JNK signaling pathway in breast cancer cells.. Free Radic Biol Med., 2021. [PubMed]
- The extracellular matrix and cardiac pressure overload: Focus on novel treatment targets.. Cells., 2024. [PubMed]
- Integrin signalling: The tug-of-war in heart hypertrophy.. Cardiovasc Res., 2006. [PubMed]
- An intrinsic mechanism of metabolic tuning promotes cardiac resilience to stress.. EMBO Mol Med., 2024. [PubMed]
- Ca2+ signaling, mitochondria and cell death.. Curr Mol Med., 2008. [PubMed]
- The Janus-faced role of Piezo1 in cardiovascular health under mechanical stimulation.. Genes Dis., 2023. [PubMed]
- Endothelial transient receptor potential channels and vascular remodeling: Extracellular Ca2+ entry for angiogenesis, arteriogenesis and vasculogenesis.. Front Physiol., 2020. [PubMed]
- Stay connected: The myoendothelial junction proteins in vascular function and dysfunction.. Vasc Pharmacol., 2025
- Piezo1 stimulates mitochondrial function via cAMP signaling.. FASEB J., 2022. [PubMed]
- Matrix stiffness induces Drp1-mediated mitochondrial fission through Piezo1 mechanotransduction in human intervertebral disc degeneration.. J Transl Med., 2023. [PubMed]
- Transient receptor potential vanilloid 4: The sixth sense of the musculoskeletal system?. Ann N Y Acad Sci., 2010. [PubMed]
- TRPV4 regulates osteoblast differentiation and mitochondrial function that are relevant for channelopathy.. Front Cell Dev Biol., 2023. [PubMed]
- Cannabidiol inhibits human glioma by induction of lethal mitophagy through activating TRPV4.. Autophagy., 2021. [PubMed]
- Surviving stress: Modulation of ATF4-mediated stress responses in normal and malignant cells.. Trends Endocrinol Metab., 2017. [PubMed]
- TRPV4 channel opening mediates pressure-induced pancreatitis initiated by Piezo1 activation.. J Clin Invest., 2020. [PubMed]
- Mechano-sensitive TET1s inhibits endothelial athero-susceptible phenotype through regulating YAP phosphorylation.. Genes Dis., 2023. [PubMed]
- Hippo-YAP/TAZ signalling in organ regeneration and regenerative medicine.. Nat Rev Mol Cell Biol., 2019. [PubMed]
- YAP/TAZ at the roots of cancer.. Cancer Cell., 2016. [PubMed]
- Yap promotes hepatocellular carcinoma metastasis and mobilization via governing cofilin/F-actin/lamellipodium axis by regulation of JNK/Bnip3/SERCA/CaMKII pathways.. Redox Biol., 2018. [PubMed]
- TAZ links exercise to mitochondrial biogenesis via mitochondrial transcription factor A.. Nat Commun., 2022. [PubMed]
- YAP inhibits the apoptosis and migration of human rectal cancer cells via suppression of JNK-Drp1-mitochondrial fission-HtrA2/Omi pathways.. Cell Physiol Biochem., 2017. [PubMed]
- Yap regulates mitochondrial structural remodeling during myoblast differentiation.. Am J Physiol Cell Physiol., 2018. [PubMed]
- Mitochondrial mechanotransduction through MIEF1 coordinates the nuclear response to forces.. Nat Cell Biol., 2024. [PubMed]
- Metabolic basis of creatine in health and disease: A bioinformatics-assisted review.. Nutrients., 2021. [PubMed]
- The creatine–phosphagen system is mechanoresponsive in pancreatic adenocarcinoma and fuels invasion and metastasis.. Nat Metab., 2020. [PubMed]
- Extracellular matrix stiffness as an energy metabolism regulator drives osteogenic differentiation in mesenchymal stem cells.. Bioact Mater., 2024. [PubMed]
- The methyltransferase SETD3-mediated histidine methylation: Biological functions and potential implications in cancers.. Biochim Biophys Acta Rev Cancer., 2021. [PubMed]
- SETD3 is a mechanosensitive enzyme that methylates actin on His73 to regulate mitochondrial dynamics and function.. J Cell Sci., 2024. [PubMed]
- AMPK is a mechano-metabolic sensor linking cell adhesion and mitochondrial dynamics to myosin-dependent cell migration.. Nat Commun., 2023. [PubMed]
- Matrix stiffening promotes perinuclear clustering of mitochondria.. Mol Biol Cell., 2024. [PubMed]
- Mitochondrial transfer in cancer: A comprehensive review.. Int J Mol Sci., 2021. [PubMed]
- Rescue of mitochondrial import failure by intercellular organellar transfer.. Nat Commun., 2024. [PubMed]
- Microtubule-based mitochondrial dynamics as a valuable therapeutic target in cancer.. Cancers (Basel)., 2021. [PubMed]
- Miro1 regulates intercellular mitochondrial transport & enhances mesenchymal stem cell rescue efficacy.. EMBO J., 2014. [PubMed]
- Miro GTPases at the crossroads of cytoskeletal dynamics and mitochondrial trafficking.. Cells., 2024. [PubMed]
- Miro proteins coordinate microtubule- and actin-dependent mitochondrial transport and distribution.. EMBO J., 2018. [PubMed]
- Miro (mitochondrial Rho GTPase), a key player of mitochondrial axonal transport and mitochondrial dynamics in neurodegenerative diseases.. Mitochondrion., 2021. [PubMed]
- Regulation of long-distance transport of mitochondria along microtubules.. Cell Mol Life Sci., 2018. [PubMed]
- Mitochondrial dynamics, positioning and function mediated by cytoskeletal interactions.. Cell Mol Life Sci., 2021. [PubMed]
- Human Myo19 is a novel myosin that associates with mitochondria.. Curr Biol., 2009. [PubMed]
- MCU complex: Exploring emerging targets and mechanisms of mitochondrial physiology and pathology.. J Adv Res., 2025. [PubMed]
- Connexin 43 in mitochondria: What do we really know about its function?. Front Physiol., 2022. [PubMed]