β-Carboline Alkaloids in Soy Sauce and Inhibition of Monoamine Oxidase (MAO)
Abstract
Monoamine oxidase (MAO) oxidizes neurotransmitters and xenobiotic amines, including vasopressor and neurotoxic amines such as the MPTP neurotoxin. Its inhibitors are useful as antidepressants and neuroprotectants. This work shows that diluted soy sauce (1/3) and soy sauce extracts inhibited human MAO-A and -B isozymes in vitro, which were measured with a chromatographic assay to avoid interferences, and it suggests the presence of MAO inhibitors. Chromatographic and spectrometric studies showed the occurrence of the β-carboline alkaloids harman and norharman in soy sauce extracts inhibiting MAO-A. Harman was isolated from soy sauce, and it was a potent and competitive inhibitor of MAO-A (0.4 µM, 44 % inhibition). The concentrations of harman and norharman were determined in commercial soy sauces, reaching 243 and 52 μg/L, respectively. Subsequently, the alkaloids 1,2,3,4-tetrahydro-β-carboline-3-carboxylic acid (THCA) and 1-methyl-1,2,3,4-tetrahydro-β-carboline-3-carboxylic acid (MTCA) were identified and analyzed in soy sauces reaching concentrations of 69 and 448 mg/L, respectively. The results show that MTCA was a precursor of harman under oxidative and heating conditions, and soy sauces increased the amount of harman under those conditions. This work shows that soy sauce contains bioactive β-carbolines and constitutes a dietary source of MAO-A and -B inhibitors.
Article type: Research Article
Keywords: monoamine oxidase, MAO inhibition, soy sauce, β-carbolines, norharman, harman, tetrahydro-β-carbolines, alkaloids, heterocyclic amines
License: © 2023 by the author. CC BY 4.0 Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Article links: DOI: 10.3390/molecules28062723 | PubMed: 36985694 | PMC: PMC10053526
Relevance: Relevant: mentioned in keywords or abstract
Full text: PDF (1.9 MB)
1. Introduction
Monoamine oxidase (MAO) is an enzyme located at the outer membranes of mitochondria in the brain, liver, intestinal mucosa, and other organs that catalyzes the oxidative deamination of biogenic amines, neurotransmitters, vasoactive dietary amines and xenobiotic amines, including dopamine, serotonin, norepinephrine, tyramine, tryptamine and the MPTP (1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine) neurotoxin. MAO appears as two isozymes, MAO-A and B, which are distinguished by substrate and inhibitor selectivity [ref. 1]. MAO-A preferentially catalyzes the oxidation of serotonin and norepinephrine and is inhibited by clorgyline, whereas MAO-B selectively catalyzes the oxidation of phenylethylamine and benzylamine and is inhibited by (R)-deprenyl. Tyramine, dopamine, and tryptamine are substrates for both enzymes. MAO isozymes play a significant role in the metabolism and regulation of neurotransmitters and biogenic amines and are involved in several diseases [ref. 1,ref. 2]. Thus, MAO-B has been implicated in neurological disorders and diseases and MAO-A in psychiatric conditions and depression [ref. 3]. An increased number of inhibitors of MAO, both of synthetic and natural product origin, are currently a matter of interest in drug discovery [ref. 1,ref. 4], both as antidepressants (MAO-A inhibitors) [ref. 3,ref. 5] or neuroprotectants (MAO-B inhibitors) [ref. 6,ref. 7,ref. 8,ref. 9,ref. 10]. The oxidation of biogenic amines and neurotransmitters by MAO produces hydrogen peroxide (H2O2), ammonia and aldehydes, which represent risk factors for oxidative cell injury and toxicity [ref. 11,ref. 12]. MAO also bioactivates proneurotoxins such as MPTP [ref. 13,ref. 14,ref. 15]. The inhibition of MAO could reduce reactive oxygen species or toxins [ref. 16,ref. 17]. On the other hand, the inhibition of peripheral MAO (e.g., intestinal and liver) has been linked to hypertensive crisis produced by a reduced metabolism of vasopressor dietary amines (e.g., tyramine) [ref. 1,ref. 18]. In this regard, the inhibition of MAO may potentiate the physiological effects of dietary vasoactive amines and exert possible food-drug interactions. The available inhibitors are sometimes irreversible inhibitors that may produce hypertensive crises, and in this regard, new inhibitors devoid of those undesirable effects are currently needed.
Dietary and environmental factors such as smoking, foods, herbal preparations or drugs might affect the action of MAO. MAO is reduced in smokers compared to nonsmokers [ref. 19,ref. 20], and cigarette smoke inhibits MAO-A and -B isozymes [ref. 21]. Naturally occurring β-carboline alkaloids have been identified in the inhibition of MAO by cigarette smoke [ref. 21,ref. 22], coffee [ref. 23] and raisins [ref. 24]. The inhibition of MAO in smokers and coffee drinkers might be linked with some biological actions, such as the addictive properties of cigarettes and depression [ref. 25,ref. 26] and the lowest incidence of Parkinson’s disease (PD) [ref. 27,ref. 28,ref. 29,ref. 30]. More research is currently needed to study the effects of diet and environmental factors on MAO enzymes. On the other hand, new inhibitors of MAO devoid of undesirable effects might arise from naturally occurring compounds and sources. β-Carbolines are naturally occurring bioactive alkaloids that exhibit an array of pharmacological effects such as the binding to benzodiazepine, imidazoline, serotonin and opiate receptors as well as inhibition of kinases and MAO [ref. 14,ref. 31,ref. 32,ref. 33,ref. 34,ref. 35,ref. 36,ref. 37,ref. 38,ref. 39]. Some of these substances are also involved in toxicological effects [ref. 14,ref. 15]. β-Carbolines can be absorbed and accumulated in tissues contributing to their presence and actions in the biological systems [ref. 40,ref. 41,ref. 42]. These alkaloids have been found in foods and seasonings [ref. 22,ref. 24,ref. 43,ref. 44,ref. 45,ref. 46,ref. 47,ref. 48]. In this regard, soy sauce is a special seasoning made from salt, water, soybeans and wheat that, after heating processes, is traditionally fermented in a process involving molds, lactic bacteria and yeast lasting months, and this process may result in an increase of β-carboline alkaloids [ref. 49]. This research aimed to study the presence of β-carboline alkaloids in soy sauce and to assess the activity and inhibition of MAO by soy sauce using a chromatographic assay. It is shown that MAO isozymes are inhibited in vitro by soy sauce and soy sauce extracts, suggesting the presence of inhibitors. The aromatic β-carboline alkaloids norharman and harman were identified and analyzed in soy sauce, and harman was subsequently isolated as a potent and competitive inhibitor of MAO-A. Finally, tetrahydro-β-carboline-3-carboxylic acid (THβC-3-COOH) alkaloids were analyzed in soy sauce, and it was found that 1-methyl-1,2,3,4-tetrahydro-β-carboline-3-carboxylic acid (MTCA) was the precursor of the MAO inhibitor harman.
2. Results
The enzymatic activity of human MAO-A and -B was assayed by chromatographic means following HPLC analysis of 4-hydroxyquinoline (4-HQ) formed by enzymatic deamination of kynuramine (kyn) substrate (Figure 1A). This chromatographic analysis allows the determination of the inhibition of MAO without interferences that may occur when using inhibitors present in complex mixtures [ref. 17,ref. 50]. The activity of MAO was studied in the presence of increasing concentrations of soy sauce. Both MAO-A and -B isozymes were inhibited in the presence of soy sauce (Figure 2A). This inhibition was reversible for MAO-A (90%) and partly reversible for MAO-B (60%), as suggested from the activity recovered following incubation of MAO with soy sauce when compared with controls. The inhibition of MAO-A and -B obtained with different samples of soy sauces (1/3 diluted) ranged from 23–50% for MAO-A and 30–73% for MAO-B (Figure 2B). In the assays, the standard inhibitors of MAO, clorgyline (1 µM) and R-deprenyl (1 µM) provided more than 90% inhibition of MAO-A and -B, respectively.
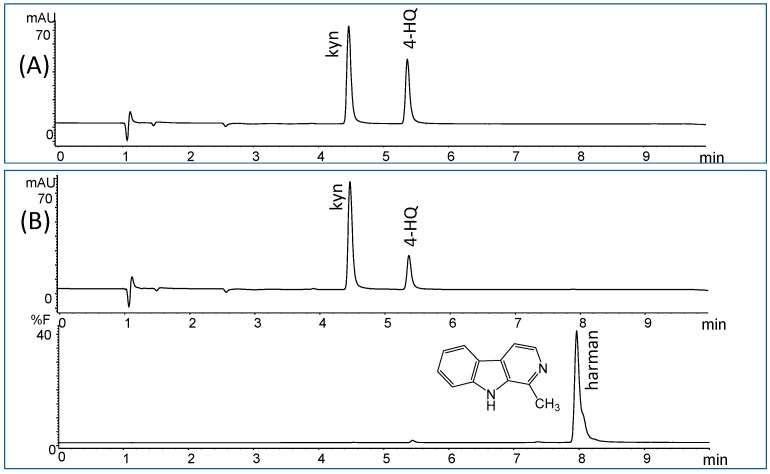
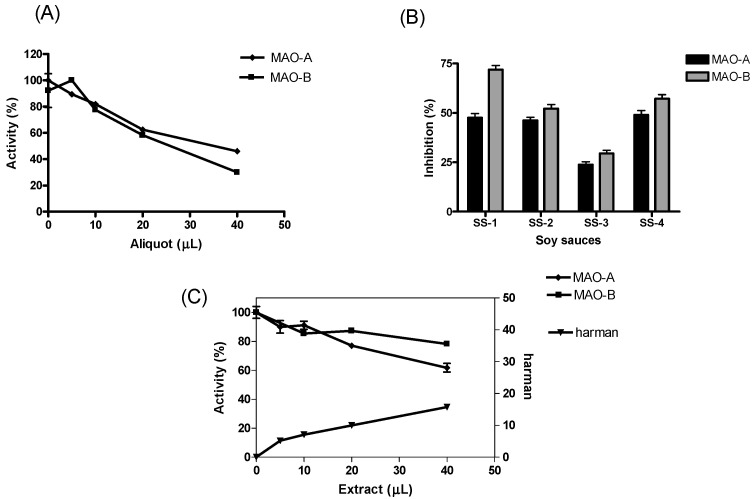
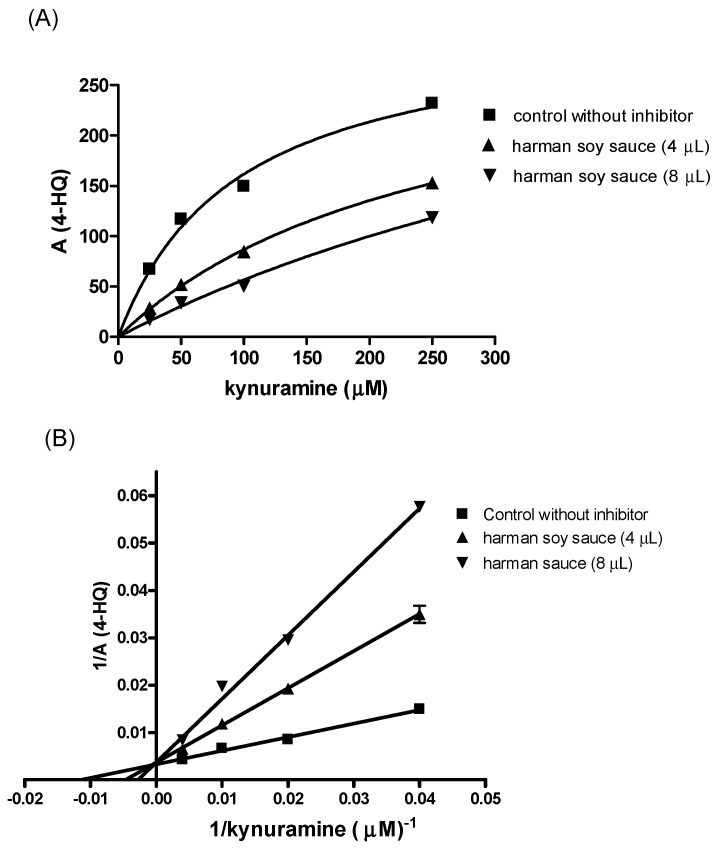
The inhibition of the deamination of kynuramine by MAO suggested the presence of inhibitors in soy sauces. Then, soy sauce with a relatively high inhibition of MAO was fractionated by solid phase extraction (SPE) using propylsulfonic (PRS) columns. The isolated extract inhibited MAO-A and, to a lesser extent, MAO-B (Figure 2C), suggesting that it contained compounds contributing to the inhibition of MAO-A. When considering dilutions, a similar degree of inhibition of MAO-A was observed with both soy sauce and their PRS extracts, suggesting that PRS extracts contained compounds that were contributors to MAO-A inhibition in soy sauce. The analysis of those PRS-extracts evidenced the presence of two β-carbolines that were identified by co-injection with standards, UV-VIS spectra and HPLC-MS as norharman (m/z at 169 (M + H)+) and harman (m/z at 183 (M + H)+; Figure 3A). Harman that was the major β-carboline in the PRS extracts, was isolated following successive RP-HPLC chromatographic injections by collecting the chromatographic peak at the end of the Diode Array Detector (DAD) and then used to study inhibition (Figure 1B and Figure 4A). This β-carboline from soy sauce was a potent inhibitor of MAO-A and showed a competitive type of inhibition when determined at different concentrations of substrate and inhibitor (Figure 4B). The inhibition of MAO-A by the harman isolated from soy sauce was in agreement with the concentration of harman included in the assays and its IC50 [ref. 21]. Thus, harman from soy sauce (0.4 μM) gave a 44 % inhibition of MAO-A using kynuramine (250 μM).
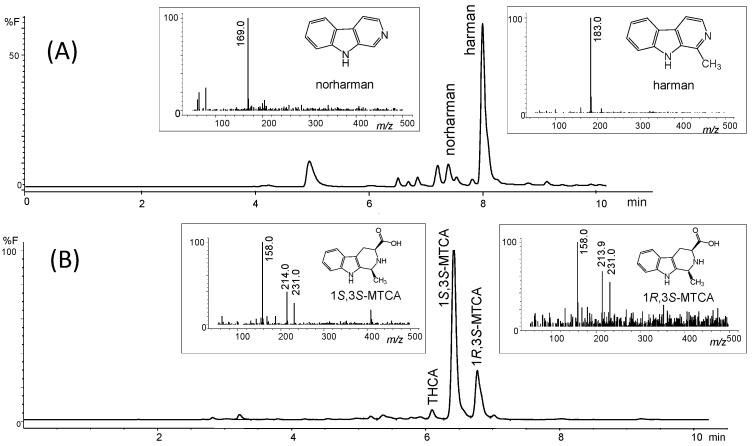
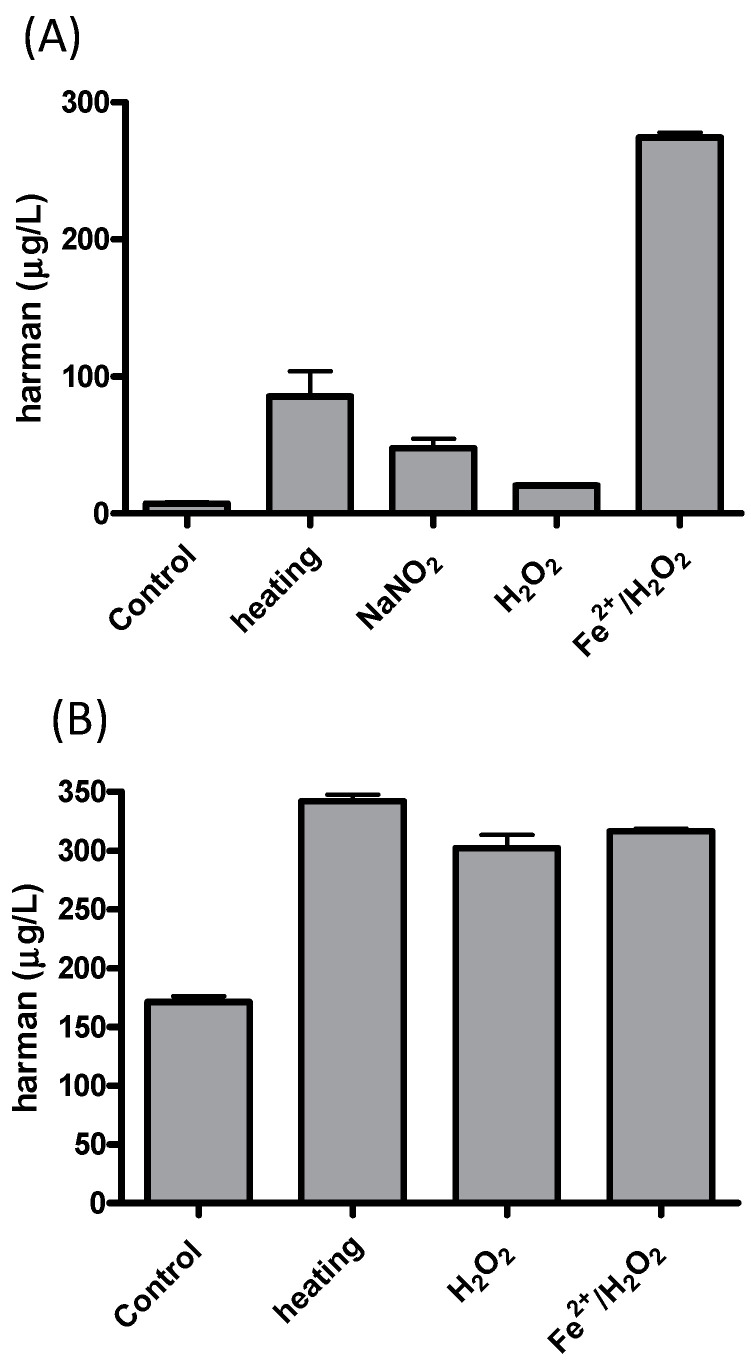
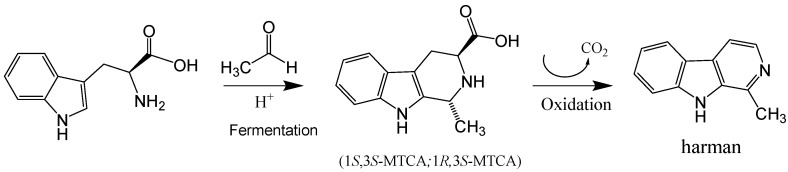
The aromatic β-carboline alkaloids norharman and harman were measured in commercial samples of soy sauces by SPE and HPLC-fluorescence detection (Table 1). Harman was a major aromatic β-carboline, and its concentration varied among different samples of soy sauces ranging from 46.7 to 243 μg/L, whereas norharman ranged from 11.6 to 52 μg/L. Subsequently, soy sauces were analyzed for the presence of tetrahydro-β-carboline-3-carboxylic acids (THβC-3-COOH) that are precursors of the aromatic β-carboline alkaloids [ref. 44,ref. 51,ref. 52,ref. 53]. Soy sauces contained 1,2,3,4-tetrahydro-β-carboline-3-carboxylic acid (THCA) and 1-methyl-1,2,3,4-tetrahydro-β-carboline-3-carboxylic acid (MTCA; Figure 3B) that were identified by chromatographic and mass spectrometric analysis (i.e., m/z at 217 (M + H)+ for THCA, and m/z at 231 (M + H)+ for MTCA). THβC-3-COOHs were determined following SPE, and their concentration was up to 1000 times higher than that of aromatic βCs (Table 1). The main THβC-3-COOH was MTCA which appeared as two diastereoisomers (1S,3S and 1R,3S) with the same spectra (Figure 3B) and concentration reached 450 mg/L. The compound MTCA was isolated from soy sauce by successive HPLC injections, and it afforded harman when heated (90 °C, 6 h; results not shown), showing that it is a direct precursor of this aromatic β-carboline. The formation of harman from MTCA was studied, and it afforded harman when treated under conditions of heating and oxidation (Figure 5A). Moreover, a sample of soy sauce increased the level of harman under conditions of heating and oxidation (Figure 5B). Therefore, these results indicate that MTCA was the precursor of harman, a potent inhibitor of MAO-A, in soy sauce (Figure 6).
Table 1: Concentration of aromatic β-carbolines and THβC-3-COOH alkaloids in different commercial soy sauces (n = 7).
| β-Carbolines | X (μg/L) | Range |
|---|---|---|
| Norharman | 34.9 | 11.6–52 |
| Harman | 165.4 | 46.7–243 |
| THβC-3-COOH | X (mg/L) | Range |
| THCA | 18.8 | 2.2–69.6 |
| 1S,3S-MTCA | 161.6 | 43.6–360.5 |
| 1R,3S-MTCA | 42.8 | 8.9–88.2 |
3. Discussion
The results of this work have shown that MAO isozymes (MAO-A and -B) are inhibited in vitro by soy sauce aliquots and soy sauce extracts suggesting the presence of inhibitors in soy sauce. They also have shown the occurrence of aromatic β-carboline alkaloids in soy sauce extracts inhibiting MAO. The β-carbolines, norharman and harman, were identified and analyzed, and among them, harman was a major compound. Subsequently, harman was isolated from soy sauce, and it was a potent inhibitor of MAO-A. The level of β-carboline alkaloids highly varied among the samples of soy sauces analyzed (46 to 243 μg/L for harman). The amount of β-carbolines in soy sauce is relatively high when compared with other foodstuffs [ref. 22]. Besides aromatic β-carboline alkaloids, the results in this work also show the presence of the tetrahydro-β-carboline-3-carboxylic acids (THβC-3-COOHs) THCA and MTCA in soy sauce. THβC-3-COOHs in foods form through the reaction of tryptophan with formaldehyde (THCA) or acetaldehyde (MTCA) [ref. 45]. Thus, MTCA forms during fermentation and storage as tryptophan reacts with acetaldehyde released (Figure 6) [ref. 45,ref. 49,ref. 54,ref. 55]. Soy sauce is made from a mixture of salt, water, soybeans and wheat that, after heating processes, is fermented in a process successively involving molds (Aspergillus oryzae), lactic bacteria and yeast that may last months, and that is finally filtered and heat-treated for sterilization. Alternatively, elaboration methods may involve the chemical hydrolysis of soybeans or chemical hydrolysis and fermentation. Then, factors such as the fermentation process, time of storage and heating processes could determine the content of THβC-3-COOH. For instance, soy sauces contain a high level of THβC-3-COOHs [ref. 45], and MTCA appeared up to 1000 times higher in concentration than harman (Table 1). Results here have shown that MTCA is the precursor of harman in soy sauce (Figure 5 and Figure 6). Harman increased in soy sauce under heating and oxidative conditions. Therefore, technological processes such as high temperatures and long storage time or oxidation could increase the levels of the MAO inhibitor harman in soy sauce. These results agree with previous results on the formation of β-carbolines from THβC-3-COOHs [ref. 44,ref. 51,ref. 53,ref. 56]. Previously, MTCA received attention as a possible precursor of mutagens when reacted with nitrite [ref. 57]. The reaction of MTCA with nitrite affords harman among other compounds, as proved during nitrosation of THβCs [ref. 56].
β-Carbolines are pyrido-indole alkaloids that occur and accumulate in biological tissues [ref. 31,ref. 34,ref. 41,ref. 42,ref. 58,ref. 59,ref. 60,ref. 61,ref. 62]. β-Carbolines also occur in foods and tobacco smoke, suggesting daily exposure to these compounds [ref. 22,ref. 44,ref. 46]. It is assumed that their occurrence in foods such as soy sauce, along with their absorption, may contribute to their physiological occurrence and accumulation in the body. Remarkably, β-carbolines exert various pharmacological and psychopharmacological effects, including antidepressant-like effects [ref. 32,ref. 37,ref. 38,ref. 61,ref. 63]. They alter the concentrations of brain neurotransmitters by interaction with serotonin, benzodiazepine, opioid and imidazoline receptors and also interact with MAO, kinases and cytochrome P450 enzymes [ref. 14,ref. 31,ref. 32,ref. 33,ref. 34,ref. 35,ref. 39,ref. 64,ref. 65]. The presence of these compounds in the human brain has been involved in alcoholism and addiction and they have been investigated as potential endogenous and/or environmental proneurotoxins involved in Parkinson’s disease [ref. 13,ref. 14,ref. 15,ref. 59,ref. 66,ref. 67]. The latter is based on the fact that under bioactivation by N-methyltransferases, β-carbolines afford neurotoxic N-methyl-β-carbolinium cations structurally resembling the neurotoxin MPP+ that is produced from MPTP with the participation of MAO [ref. 62,ref. 67]. Moreover, endogenous β-carbolines might somehow affect the metabolism of exogenous amines and neuroamines, exerting potential biological actions [ref. 23,ref. 24,ref. 61].
Further studies are needed to fully delineate the implications of dietary, environmental, and endogenous β-carboline alkaloids in human health. Some of the effects attributed to β-carboline alkaloids could be produced by their interaction with MAO enzymes [ref. 21,ref. 23,ref. 68]. The primary role of MAO isozymes in the CNS lies in the metabolism of amines and the regulation of neurotransmitter levels and intracellular amine stores. In the gastrointestinal tract, the circulatory system and the liver, MAO regulates the levels of exogenous dietary amines that exert vasopressor effects serving a protective function. The use of MAO inhibitors, particularly irreversibly inhibitors, may cause a hypertensive crisis when the patients consume tyramine-containing foods (the so-called “cheese effect”) [ref. 1,ref. 18,ref. 69,ref. 70]. MAO also metabolizes toxic xenobiotic amines such as the neurotoxin MPTP and, in this regard, the inhibitors of MAO can be protective agents [ref. 13,ref. 14,ref. 71,ref. 72]. Moreover, the oxidation of amine substrates by MAO results in the production of hydrogen peroxide, ammonia, and aldehydes, which are risk factors for cell oxidative injury [ref. 12,ref. 17,ref. 73]. Therefore, the use of MAO-inhibiting substances can protect against toxicants and oxidative stress [ref. 11,ref. 17,ref. 71,ref. 74]. Currently, MAO inhibitors are being developed against Parkinsonism and neurodegeneration and as antidepressant drugs [ref. 1,ref. 3,ref. 4,ref. 5,ref. 6,ref. 7,ref. 8,ref. 9]. Inhibition of MAO could help to spare neurotransmitters such as dopamine, serotonin and others while reducing reactive oxygen species. Alternatively, the inhibition of peripheral MAO could eventually affect the metabolism of exogenous and dietary amines like tyramine or tryptamine, potentiating vasopressor effects [ref. 1,ref. 3]. This work has shown the presence of β-carboline alkaloids in soy sauce, which are inhibitors of MAO. Based on concentration, other substances can contribute to MAO inhibition in soy sauce in addition to β-carbolines. However, β-carbolines could be absorbed and accumulated in tissues exhibiting MAO inhibition [ref. 62]. Soy sauce products might be beneficial against neurodegenerative diseases [ref. 75,ref. 76]. This study shows that soy sauce contains MAO inhibitors, and the inhibition of MAO is a target in neuroprotection.
4. Materials and Methods
Kynuramine, 4-hydroxyquinoline, norharman (9H-pyrido-(3,4-b)-indole), harman (1-methyl-9H-pyrido-(3,4-b)-indole) and 1,2,3,4-tetrahydro-β-carboline-3-carboxilic acid (THCA) and 1-methyl-1,2,3,4-tetrahydro-β-carboline-3-carboxilic acid (1S,3S-MTCA) were purchased from Sigma. Recombinant human monoamine oxidase A and B were obtained from Gentest. HPLC grade acetonitrile, methanol and dimethyl sulfoxide (DMSO) were from Scharlau, Barcelona (Spain), and dichloromethane from Merck, Darmstad (Germany). Commercial samples of soy sauces from different producers and origins, also labeled as produced from natural fermentation, were purchased in local supermarkets. Sample preparation of soy sauces for analysis and enzyme inhibition was carried out in several ways: (a) diluted soy sauces (1/3) with phosphate buffer pH 7.4–10% DMSO were used for MAO inhibition; (b) soy sauces were fractionated by SPE, and the eluting fraction of TP-MeOH used for analysis and MAO inhibition; and (c) the β-carboline harman was isolated by HPLC and subsequently used for MAO inhibition and kinetic studies.
4.1. Isolation of Tetrahydro-β-carboline-3-carboxylic Acid and β-Carboline Alkaloids from Soy Sauces
(a) Tetrahydro-β-carboline-3-carboxylic acids (THβC-3-COOHs) were analyzed from soy sauces by using benzenesulfonic acid (SCX)-columns following a previously described procedure that affords good performance and reliability with recoveries higher than 90% [ref. 45,ref. 54]. The elution fraction of 0.4 M phosphate buffer-methanol (1:1), pH 9, containing the THβC-3-COOH was injected into RP-HPLC-fluorescence (excitation, 270 nm; emission, 343 nm) for quantitative analysis and into HPLC-MS for identification, as mentioned below. Quantitation was carried out from calibration curves constructed with standards of THβC-3-COOH isolated under the same procedure [ref. 45,ref. 54].
(b) The aromatic β-carbolines were isolated for subsequent chromatographic analysis, identification and MAO inhibition studies by using a solid phase extraction procedure reported before that affords good performance and reliability with recoveries higher than 90% [ref. 22,ref. 46]. Briefly, soy sauce diluted with 0.6 M HClO4 and added with 125 μL 1-ethyl-β-carboline (0.2 mg/L) as internal standard was passed through a propylsulfonic acid (PRS)-derivatized silica column that was eluted with water (6 mL), 0.4 M phosphate buffer, pH 9 (3 mL) and 0.2 M buffer phosphate-methanol (1:1), pH 9 (3 mL). The eluates of 0.2 M buffer phosphate-methanol (1:1), pH 9 (3 mL) containing the β-carbolines, were analyzed by HPLC and norharman and harman detected by fluorescence (300 nm, excitation and 433 nm, emission). Quantitation was obtained from calibration curves of standards isolated under the same procedure. The same PRS-SPE procedure but without an internal standard was used to isolate the fractions containing β-carboline for subsequent MAO inhibition studies. In this case, the eluting fractions of 0.4 M phosphate buffer, pH 9 (3 mL) and 0.2 M buffer phosphate-methanol (1:1), pH 9 (3 mL) were mixed and used for MAO inhibition. In order to isolate the β-carboline harman, PRS fractions of 0.2 M buffer phosphate-methanol (1:1) pH 9 were evaporated under vacuum and extracted with dichloromethane (Merck). The organic phase was evaporated, redissolved and injected into HPLC as mentioned below, with the peak corresponding to the β-carboline harman collected from successive HPLC injections. After removing acetonitrile, harman was extracted with dichloromethane in pH 9, concentrated to dryness, and redissolved in phosphate buffer containing 30% DMSO, and used for inhibition of MAO. Corresponding blanks following the same procedures but without β-carbolines were used in the assays.
4.2. Formation of Harman from 1-Methyl-1,2,3,4-Tetrahydro-β-Carboline-3-Carboxylic Acid (MTCA)
To study the formation of harman, a standard solution of 1S,3S-MTCA (Sigma) 50 μM in 100 mM phosphate buffer pH 4 was heated (80 °C, 3 h) or added to H2O2 (2 mM) (1 h), NaNO2 (100 μM) (1 h), or H2O2 (50 μM) plus FeSO4 (50 μM) (1 h), and subsequently analyzed by RP-HPLC for harman as indicated below. Also, the PRS extracts isolated from soy sauce were successively injected into the RP-HPLC as mentioned below, and the peaks corresponding to 1S,3S-MTCA and 1R,3S-MTCA collected and heated at 90 °C for 3–6 h and subsequently analyzed for harman as indicated below. On the other hand, a sample of soy sauce was treated under several conditions: control (37 °C, 2 h), heating (90 °C, 2 h), H2O2 (5 mM) or FeSO4 (5 mM) plus H2O2 (5 mM) (37 °C, 2 h), and subsequently analyzed for harman.
4.3. Monoamine Oxidase (MAO-A and B) Assay and Inhibition by Soy Sauce and β-Carbolines Isolated from Soy Sauce
A chromatographic assay was carried out to determine MAO activity [ref. 17,ref. 21,ref. 50]. It was performed with membrane protein fractions containing MAO-A or MAO-B that were diluted to the desired concentrations in 100 mM potassium phosphate buffer (pH 7.4) [ref. 21,ref. 50]. A 0.2 mL reaction mixture containing 0.01 mg/mL protein and 0.25 mM kynuramine in 100 mM potassium phosphate (pH 7.4) was incubated at 37 °C for 40 min. After incubation, the reaction was stopped by the addition of 2N NaOH (75 μL), followed by the addition of 70% HClO4 (25 μL), and the sample was centrifuged (10,000× g) for 5 min. Under these conditions, kynuramine deaminated by MAO spontaneously cyclizes to give 4-hydroxyquinoline. An aliquot of the supernatant (20 μL) was injected into the HPLC, and the deamination product of kynuramine (i.e., 4-hydroxyquinoline) formed during the enzymatic reaction determined by RP-HPLC-DAD at 320 nm. A response curve of area versus concentration was constructed to calculate the concentration of 4-hydroxyquinoline. The standard inhibitors of MAO, clorgyline (MAO-A inhibitor) and deprenyl (MAO-B inhibitor) were used as reference inhibitors in the assays (1 µM).
The inhibition assays were performed with: (a) aliquots of diluted soy sauce (diluted 1/3 in 100 mM phosphate buffer pH 7.4 with 10% of DMSO), (b) soy sauce extracts and fractions prepared from SPE containing the β-carbolines (i.e., buffer-methanol PRS fractions), and (c) samples of isolated harman from soy sauces. For that, aliquots of those samples were added to reaction mixtures containing kynuramine (0.25 mM) and MAO enzyme (A or -B; 0.01 mg/mL membrane protein) in 100 mM potassium phosphate buffer (pH 7.4), as above. Corresponding blanks and controls were used. The MAO kinetic and the mechanism of inhibition were assessed by analyzing the corresponding Michaelis-Menten curves and double reciprocal Lineweaver-Burk plots obtained at different concentrations of the substrate kynuramine. To determine MAO-binding reversibility, membrane proteins of MAO-A and B (0.12 mg/mL) in 100 mM phosphate buffer (pH 7.4) were preincubated (37 °C, 40 min) with diluted soy sauce aliquots. The mixtures were centrifuged (15,000× g) for 15 min to pellet membrane proteins, washed twice with 100 mM phosphate buffer, and finally, the pellet was resuspended in 100 mM phosphate buffer + 10% DMSO (0.1 mL). An aliquot (40 μL) was used to measure MAO activity and compared with corresponding controls.
4.4. RP-HPLC chromatographic Analysis and Chemical Identification by MS
The analysis of 4-hydroxyquinoline (kynuramine deamination product), β-carbolines and tetrahydro-β-carbolines was performed by RP-HPLC with uv-DAD and fluorescence detection using an HPLC 1050 (Agilent Technologies, Santa Clara, CA, USA) with a Diode Array Detector (DAD) and a 1046A-fluorescence detector. A 150 mm × 3.9 mm, 4 μm, Nova-pak C18 column (Waters, Milford, MA, USA) was used for chromatographic separation. Chromatographic conditions were 50 mM ammonium phosphate buffer (pH 3) (buffer A) and 20% of A in acetonitrile (buffer B). Gradient programmed from 0% (100% A) to 32% B in 8 min, and 90% B in 15 min. The flow rate was 1 mL/min, the column temperature was 40 °C, and the injection volume was 20 μL. Absorbance detection was set at 320 nm (analysis of 4-hydroxyquinoline), whereas fluorescence detection was used for tetrahydro-β-carbolines (270 nm, excitation and 343 nm, emission) and norharman and harman (300 nm, excitation and 433 nm, emission). Identification of compounds was done by UV, fluorescence and mass spectrometry. The identification by HPLC-ESI-mass spectrometry of 4-hydroxyquinoline in MAO assays was carried out as previously [ref. 50]. Identification of the β-carbolines norharman and harman in soy sauce was carried out in the PRS extracts, and the THβC-3-COOH in SCX extracts was obtained as mentioned above. They were analyzed with a 2.1 × 150 mm Zorbax SB-C18, 5 μm, column (Agilent Technologies, Santa Clara, CA, USA) by using an HPLC-MSD series 1100 (Agilent) (electrospray-positive ion mode). Eluent A: formic acid (0.5%); B: formic acid (0.5%) in acetonitrile; 80% B in 30 min, flow rate 0.25 mL/min.; T: 40 °C; mass range 50–700 u and cone voltage 100 V.
5. Conclusions
This work has shown that soy sauce inhibits MAO-A and -B isozymes in vitro, as evidenced by an assay of kynuramine deamination performed by chromatographic means, suggesting the occurrence of MAO inhibitors in soy sauces. The inhibition of MAO was reversible for MAO-A and partly reversible for MAO-B. Soy sauces were analyzed, and the β-carboline alkaloids, norharman and harman, were identified. Subsequently, harman was isolated from soy sauce, and it potently inhibited MAO-A in a competitive mode. This β-carboline was analyzed in several soy sauces, and it reached a concentration of 243 μg/L. In addition, the THβC-3-COOH compounds, THCA and MTCA, were identified and analyzed in soy sauces. MTCA isolated from soy sauce afforded harman under conditions of heating and oxidation. Also, under heating and oxidative conditions, soy sauces increased the levels of harman, showing that MTCA was the precursor of harman in soy sauces. Then, soy sauce contained β-carboline alkaloids that are considered bioactive compounds, and among them, harman is a potent inhibitor of MAO-A, as shown here. Results suggest that soy sauce constitutes a dietary source of MAO inhibitors.
References
- M.B.H. Youdim, D. Edmondson, K.F. Tipton. The therapeutic potential of monoamine oxidase inhibitors. Nat. Rev. Neurosci., 2006. [DOI | PubMed]
- J.C. Shih, K. Chen, M.J. Ridd. Monoamine oxidase: From genes to behavior. Annu. Rev. Neurosci., 1999. [DOI | PubMed]
- M. Yamada, H. Yasuhara. Clinical pharmacology of MAO inhibitors: Safety and future. Neurotoxicology, 2004. [DOI | PubMed]
- R.R. Ramsay, K.F. Tipton. Assessment of enzyme inhibition: A review with examples from the development of monoamine oxidase and cholinesterase inhibitory drugs. Molecules, 2017. [DOI | PubMed]
- W.A. Nolen, E. Hoencamp, P.F. Bouvy, P.M.J. Haffmans. Reversible monoamine oxidase-A inhibitors in resistant major depression. Clin. Neuropharmacol., 1993. [DOI | PubMed]
- S. Carradori, M. Fantacuzzi, A. Ammazzalorso, A. Angeli, B. De Filippis, S. Galati, A. Petzer, J.P. Petzer, G. Poli, T. Tuccinardi. Resveratrol analogues as dual inhibitors of monoamine oxidase B and carbonic anhydrase VII: A new multi-target combination for neurodegenerative diseases?. Molecules, 2022. [DOI | PubMed]
- T. Behl, D. Kaur, A. Sehgal, S. Singh, N. Sharma, G. Zengin, F.L. Andronie-Cioara, M.M. Toma, S. Bungau, A.G. Bumbu. Role of monoamine oxidase activity in Alzheimer’s Disease: An insight into the therapeutic potential of inhibitors. Molecules, 2021. [DOI | PubMed]
- N.D. Chaurasiya, F. Leon, I. Muhammad, B.L. Tekwani. Natural products inhibitors of monoamine oxidases: Potential new drug leads for neuroprotection, neurological disorders, and neuroblastoma. Molecules, 2022. [DOI | PubMed]
- L. Pisani, M. Catto, F. Leonetti, O. Nicolotti, A. Stefanachi, F. Campagna, A. Carotti. Targeting monoamine oxidases with multipotent ligands: An emerging strategy in the search of new drugs against neurodegenerative diseases. Curr. Med. Chem., 2011. [DOI | PubMed]
- N.D. Chaurasiya, M.A. Ibrahim, I. Muhammad, L.A. Walker, B.L. Tekwani. Monoamine oxidase inhibitory constituents of propolis: Kinetics and mechanism of inhibition of recombinant human MAO-A and MAO-B. Molecules, 2014. [DOI | PubMed]
- G. Cohen, R. Farooqui, N. Kesler. Parkinson disease: A new link between monoamine oxidase and mitochondrial electron flow. Proc. Natl. Acad. Sci. USA, 1997. [DOI | PubMed]
- N. Hauptmann, J. Grimsby, J.C. Shih, E. Cadenas. The metabolism of tyramine by monoamine oxidase A/B causes oxidative damage to mitochondrial DNA. Arch. Biochem. Biophys., 1996. [DOI | PubMed]
- J.W. Langston, I. Irwin, E.B. Langston, L.S. Forno. 1-Methyl-4-phenylpyridinium ion (MPP+): Identification of a metabolite of MPTP, a toxin selective to the substantia nigra. Neurosci. Lett., 1984. [DOI | PubMed]
- T. Herraiz, H. Guillén, V.J. Arán, J.R. Idle, F.J. Gonzalez. Comparative aromatic hydroxylation and N-demethylation of MPTP neurotoxin and its analogs, N-methylated β-carboline and isoquinoline alkaloids, by human cytochrome P450 2D6. Toxicol. Appl. Pharm., 2006. [DOI]
- T. Herraiz, H. Guillen, J. Galisteo. N-methyltetrahydro-β-carboline analogs of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) neurotoxin are oxidized to neurotoxic β-carbolinium cations by heme peroxidases. Biochem. Biophys. Res Commun., 2007. [DOI | PubMed]
- T. Herraiz, H. Guillén. Monoamine oxidase-A inhibition and associated antioxidant activity in plant extracts with potential antidepressant actions. BioMed Res. Int., 2018. [DOI | PubMed]
- T. Herraiz, A. Flores, L. Fernández. Analysis of monoamine oxidase (MAO) enzymatic activity by high-performance liquid chromatography-diode array detection combined with an assay of oxidation with a peroxidase and its application to MAO inhibitors from foods and plants. J. Chromatogr. B, 2018. [DOI]
- E.E. Billett. Monoamine oxidase (MAO) in human peripheral tissues. Neurotoxicology, 2004. [DOI | PubMed]
- J.S. Fowler, N.D. Volkow, G.J. Wang, N. Pappas, J. Logan, R. MacGregor, D. Alexoff, C. Shea, D. Schlyer, A.P. Wolf. Inhibition of monoamine oxidase B in the brains of smokers. Nature, 1996. [DOI | PubMed]
- J.S. Fowler, N.D. Volkow, G.J. Wang, N. Pappas, J. Logan, C. Shea, D. Alexoff, R.R. MacGregor, D.J. Schlyer, I. Zezulkova. Brain monoamine oxidase A inhibition in cigarette smokers. Proc. Natl. Acad. Sci. USA, 1996. [DOI | PubMed]
- T. Herraiz, C. Chaparro. Human monoamine oxidase is inhibited by tobacco smoke: β-carboline alkaloids act as potent and reversible inhibitors. Biochem. Biophys. Res. Commun., 2005. [DOI | PubMed]
- T. Herraiz. Relative exposure to β-carbolines norharman and harman from foods and tobacco smoke. Food Addit. Contam., 2004. [DOI | PubMed]
- T. Herraiz, C. Chaparro. Human monoamine oxidase enzyme inhibition by coffee and β-carbolines norharman and harman isolated from coffee. Life Sci., 2006. [DOI | PubMed]
- T. Herraiz. Identification and occurrence of β-carboline alkaloids in raisins and inhibition of monoamine oxidase (MAO). J. Agric. Food Chem., 2007. [DOI | PubMed]
- I. Berlin, O. SpreuxVaroquaux, S. Said, J.M. Launay. Effects of past history of major depression on smoking characteristics, monoamine oxidase-A and -B activities and withdrawal symptoms in dependent smokers. Drug Alcohol Depend., 1997. [DOI | PubMed]
- A.C. Harris, P. Muelken, A. Alcheva, I. Stepanov, M.G. LeSage. Cigarette smoke extract, but not electronic cigarette aerosol extract, inhibits monoamine oxidase in vitro and produces greater acute aversive/anhedonic effects than nicotine alone on intracranial self-stimulation in rats. Front. Neurosci., 2022. [DOI | PubMed]
- M.A. Hernan, B. Takkouche, F. Caamano-Isorna, J.J. Gestal-Otero. A meta-analysis of coffee drinking, cigarette smoking, and the risk of Parkinson’s disease. Ann. Neurol., 2002. [DOI | PubMed]
- P. Ragonese, G. Salemi, L. Morgante, P. Aridon, A. Epifanio, D. Buffa, F. Scoppa, G. Savettieri. A case-control study on cigarette, alcohol, and coffee consumption preceding Parkinson’s disease. Neuroepidemiology, 2003. [DOI | PubMed]
- S.M. Carneiro, M.B.P. Oliveira, R.C. Alves. Neuroprotective properties of coffee: An update. Trends Food Sci. Technol., 2021. [DOI]
- J. Grzelczyk, G. Budryn, J. Peña-García, D. Szwajgier, I. Gałązka-Czarnecka, J. Oracz, H. Pérez-Sánchez. Evaluation of the inhibition of monoamine oxidase A by bioactive coffee compounds protecting serotonin degradation. Food Chem., 2021. [DOI | PubMed]
- M.M. Airaksinen, I. Kari. Beta-Carbolines, psychoactive compounds in the mammalian body. Part II: Effects. Med. Biol., 1981. [PubMed]
- S.S. Baum, R. Hill, H. Rommelspacher. Harman-induced changes of extracellular concentrations of neurotransmitters in the nucleus accumbens of rats. Eur. J. Pharmacol., 1996. [DOI | PubMed]
- S.M. Husbands, R.A. Glennon, S. Gorgerat, R. Gough, R. Tyacke, J. Crosby, D.J. Nutt, J.W. Lewis, A.L. Hudson. β-Carboline binding to imidazoline receptors. Drug Alcohol Depend., 2001. [DOI | PubMed]
- A.M. Yu, J.R. Idle, T. Herraiz, A. Kupfer, F.J. Gonzalez. Screening for endogenous substrates reveals that CYP2D6 is a 5-methoxyindolethylamine O-demethylase. Pharmacogenetics, 2003. [DOI | PubMed]
- E. Ergene, E.P. Schoener. Effects of harmane (1-methyl-β-carboline) on neurons in the nucleus accumbens of the rat. Pharm. Biochem. Behav., 1993. [DOI]
- A. Miralles, S. Esteban, A. Sastre-Coll, D. Moranta, V.J. Asensio, J.A. Garcia-Sevilla. High-affinity binding of β-carbolines to imidazoline I-2B receptors and MAO-A in rat tissues: Norharman blocks the effect of morphine withdrawal on DOPA/noradrenaline synthesis in the brain. Eur. J. Pharmacol., 2005. [DOI | PubMed]
- A. Beato, A. Gori, B. Boucherle, M. Peuchmaur, R. Haudecoeur. β-Carboline as a privileged scaffold for multitarget strategies in Alzheimer’s disease therapy. J. Med. Chem., 2021. [DOI | PubMed]
- M. Tarpley, H.O. Oladapo, D. Strepay, T.B. Caligan, L. Chdid, H. Shehata, J.R. Roques, R. Thomas, C.P. Laudeman, R.U. Onyenwoke. Identification of harmine and β-carboline analogs from a high-throughput screen of an approved drug collection; profiling as differential inhibitors of DYRK1A and monoamine oxidase A and for in vitro and in vivo anti-cancer studies. Eur. J. Pharm. Sci., 2021. [DOI | PubMed]
- A. Prah, T. Gavranić, A. Perdih, M. Sollner Dolenc, J. Mavri. Computational insights into β-carboline inhibition of monoamine oxidase A. Molecules, 2022. [DOI | PubMed]
- A. Ostergren, A. Annas, K. Skog, N.G. Lindquist, E.B. Brittebo. Long-term retention of neurotoxic β-carbolines in brain neuromelanin. J. Neural Transm., 2004. [DOI | PubMed]
- C.A. Parker, N.J. Anderson, E.S.J. Robinson, R. Price, R.J. Tyacke, S.M. Husbands, M.P. Dillon, R.M. Eglen, A.L. Hudson, D.J. Nutt. Harmane and harmalan are bioactive components of classical clonidine-displacing substance. Biochemistry, 2004. [DOI | PubMed]
- N.J. Anderson, R.J. Tyacke, S.M. Husbands, D.J. Nutt, A.L. Hudson, E.S.J. Robinson. In vitro and ex vivo distribution of H-3 harmane, an endogenous β-carboline, in rat brain. Neuropharmacology, 2006. [DOI | PubMed]
- T. Herraiz. Analysis of the bioactive alkaloids tetrahydro-β-carboline and β-carboline in food. J. Chromatogr. A, 2000. [DOI | PubMed]
- T. Herraiz. Tetrahydro-β-carboline bioactive alkaloids in beverages and foods. Nutraceutical Beverages: Chemistry, Nutrition, and Health Effects, 2004
- T. Herraiz. Occurrence of tetrahydro-β-carboline-3-carboxylic acids in commercial foodstuffs. J. Agric. Food Chem., 1996. [DOI]
- T. Herraiz. Identification and occurrence of the bioactive β-carbolines norharman and harman in coffee brews. Food Addit. Contam., 2002. [DOI | PubMed]
- T. Herraiz, A. Peña, H. Mateo, M. Herraiz, A. Salgado. Formation, characterization, and occurrence of β-carboline alkaloids derived from α-dicarbonyl compounds and L-tryptophan. J. Agric. Food Chem., 2022. [DOI | PubMed]
- T. Herraiz, F. Vera. Occurrence, formation from D-fructose and 3-deoxyglucosone, and activity of the carbohydrate-derived β-carbolines in foods. J. Agric. Food Chem., 2021. [DOI | PubMed]
- T. Herraiz, C.S. Ough. Chemical and technological factors determining tetrahydro-β-carboline-3-carboxylic acid content in fermented alcoholic beverages. J. Agric. Food Chem., 1993. [DOI]
- T. Herraiz, C. Chaparro. Analysis of monoamine oxidase enzymatic activity by reversed-phase high performance liquid chromatography and inhibition by β-carboline alkaloids occurring in foods and plants. J. Chromatogr. A, 2006. [DOI | PubMed]
- T. Herraiz. Tetrahydro-β-carboline-3-carboxylic acid compounds in fish and meat: Possible precursors of co-mutagenic β-carbolines norharman and harman in cooked foods. Food Addit. Contam., 2000. [DOI | PubMed]
- T. Herraiz, J. Galisteo. Tetrahydro-β-carboline alkaloids occur in fruits and fruit juices. Activity as antioxidants and radical scavengers. J. Agric. Food Chem., 2003. [DOI | PubMed]
- S. Diem, B. Gutsche, M. Herderich. Degradation of tetrahydro-β-carbolines in the presence of nitrite: HPLC-MS analysis of the reaction products. J. Agric. Food Chem., 2001. [DOI | PubMed]
- T. Herraiz. Occurrence of 1,2,3,4-tetrahydro-β-carboline-3-carboxylic acid and 1-methyl-1,2,3,4-tetrahydro-β-carboline-3-carboxylic acid in fruit juices, purees, and jams. J. Agric. Food Chem., 1998. [DOI]
- T. Herraiz. 1-methyl-1,2,3,4-tetrahydro-β-carboline-3-carboxylic acid and 1,2,3,4-tetrahydro-β-carboline-3-carboxylic acid in fruits. J. Agric. Food Chem., 1999. [DOI | PubMed]
- T. Herraiz, J. Galisteo. Nitrosative deamination of 2’-deoxyguanosine and DNA by nitrite, and antinitrosating activity of β-carboline alkaloids and antioxidants. Food Chem. Toxicol., 2018. [DOI | PubMed]
- K. Wakabayashi, M. Ochiai, H. Saito, M. Tsuda, Y. Suwa, M. Nagao, T. Sugimura. Presence of 1-methyl-1,2,3,4-tetrahydro-β-carboline-3-carboxylic acid, a precursor of a mutagenic nitroso compound, in soy sauce. Proc. Natl. Acad. Sci. USA, 1983. [DOI | PubMed]
- D. Fekkes, W.T. Bode. Occurrence and partition of the β-carboline norharman in rat organs. Life Sci., 1993. [DOI | PubMed]
- K. Matsubara, T. Gonda, H. Sawada, T. Uezono, Y. Kobayashi, T. Kawamura, K. Ohtaki, K. Kimura, A. Akaike. Endogenously occurring β-carboline induces parkinsonism in nonprimate animals: A possible causative protoxin in idiopathic Parkinson’s disease. J. Neurochem., 1998. [DOI | PubMed]
- T. May, A. Greube, S. Strauss, D. Heineke, J. Lehmann, H. Rommelspacher. Comparison of the in-vitro binding characteristics of the β-carbolines harman and norharman in rat-brain and liver and in bovine adrenal-medulla. Naunyn-Schmiedeberg’s Arch. Pharm., 1994. [DOI | PubMed]
- E.S.J. Robinson, N.J. Anderson, J. Crosby, D.J. Nutt, A.L. Hudson. Endogenous β-carbolines as clonidine-displacing substances. Ann. N. Y. Acad. Sci., 2003. [DOI | PubMed]
- T. Herraiz. N-methyltetrahydropyridines and pyridinium cations as toxins and comparison with naturally-occurring alkaloids. Food Chem. Toxicol., 2016. [DOI | PubMed]
- F. Aricioglu, H. Altunbas. Harmane induces anxiolysis and antidepressant-like effects in rats. Ann. N. Y. Acad. Sci., 2003. [DOI | PubMed]
- K. Kawanishi, N. Eguchi, T. Hayashi, Y. Hashimoto. Relationship between occurrence of tremor/convulsion and level of β-carbolines in the brain after administration of β-carbolines into mice. Pharm. Biochem. Behav., 1994. [DOI | PubMed]
- T. Herraiz, H. Guillén, V.J. Arán. Oxidative metabolism of the bioactive and naturally occurring β-carboline alkaloids, norharman and harman, by human cytochrome P450 enzymes. Chem. Res. Toxicol., 2008. [DOI | PubMed]
- D.A. Gearhart, P.F. Toole, J.W. Beach. Identification of brain proteins that interact with 2-methylnorharman: An analog of the parkinsonian-inducing toxin, MPP+. Neurosci. Res., 2002. [DOI | PubMed]
- M.A. Collins, E.J. Neafsey. β-Carboline analogs of N-methyl-4-phenyl-1,2,5,6-tetrahydropyridine (MPTP): Endogenous factors underlying idiopathic Parkinsonism. Neurosci. Lett., 1985. [DOI | PubMed]
- H. Kim, S.O. Sablin, R.R. Ramsay. Inhibition of monoamine oxidase A by β-carboline derivatives. Arch. Biochem. Biophys., 1997. [DOI | PubMed]
- M.B.H. Youdim, M. Weinstock. Therapeutic applications of selective and non-selective inhibitors of monoamine oxidase A and B that do not cause significant tyramine potentiation. Neurotoxicology, 2004. [DOI | PubMed]
- B. Davies, R. Bannister, P. Sever. Pressor amines and monoamine oxidase inhibitors for treatment of postural hypotension in autonomic failure: Llimitations and hazards. Lancet, 1978. [DOI | PubMed]
- T. Herraiz, H. Guillen. Inhibition of the bioactivation of the neurotoxin MPTP by antioxidants, redox agents and monoamine oxidase inhibitors. Food Chem. Toxicol., 2011. [DOI | PubMed]
- T. Herraiz, V.J. Aran, H. Guillen. Nitroindazole compounds inhibit the oxidative activation of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) neurotoxin to neurotoxic pyridinium cations by human monoamine oxidase (MAO). Free Radic. Res., 2009. [DOI | PubMed]
- C. Vindis, M.H. Seguelas, P. Bianchi, A. Parini, C. Cambon. Monoamine oxidase B induces ERK-dependent cell mitogenesis by hydrogen peroxide generation. Biochem. Biophys. Res. Commun., 2000. [DOI | PubMed]
- R.E. Heikkila, L. Manzino, F.S. Cabbat, R.C. Duvoisin. Protection against the dopaminergic neurotoxicity of 1-methyl-4-phenyl-1,2,5,6-tetrahydropyridine by monoamine-oxidase inhibitors. Nature, 1984. [DOI | PubMed]
- C.H. Jang, J. Oh, J.S. Lim, H.J. Kim, J.-S. Kim. Fermented soy products: Beneficial potential in neurodegenerative diseases. Foods, 2021. [DOI | PubMed]
- D.-C. Kim, T.H. Quang, C.-S. Yoon, N.T. Thanh Ngan, S.I. Lim, S.Y. Lee, Y.C. Kim, H. Oh. Anti-neuroinflammatory activities of indole alkaloids from kanjang (Korean fermented soy source) in lipopolysaccharide-induced BV2 microglial cells. Food Chem., 2016. [DOI | PubMed]