
Could cannabinoids provide a new hope for ovarian cancer patients?
Abstract
It is known that gynecological cancers remain a worldwide problem and as shown by the statistics, there is a need for new gynecological cancer treatments. Cannabinoids, the pharmacologically active compounds of the Cannabis sativa plant, have been used for many centuries by individuals as a symptomatic treatment to alleviate pain, nausea, vomiting, and to help stimulate appetite. Research has revealed that cannabinoids also exert anti‐cancer activity such as anti‐proliferative and pro‐apoptotic effects through a variety of mechanisms. There is significant value in the development of these compounds as anti‐cancer therapies in clinical practice as they do not produce the typical toxic side effects that exist with conventional therapies and recent clinical trials have shown their great tolerability by patients at high doses. Cannabinoids can induce psychoactive effects that could limit their progression. Therefore, non‐psychoactive cannabinoids are attracting pharmacological interest due to their inability to produce psychological effects. Recent studies have focussed on non‐psychoactive cannabinoids in ovarian cancer and have revealed promising pre‐clinical results that indicate that these compounds may have potential benefits in the treatment of these cancers. However, there are still unanswered questions and research gaps that need to be addressed. This review summarizes the current understanding of this topic and identifies the current gaps in knowledge that provide a useful direction for future work.
Article type: Review Article
Keywords: cannabinoids, non‐psychoactive cannabinoids, ovarian cancer
Affiliations: Department of Pharmacy, School of Applied Sciences University of Huddersfield Huddersfield UK
License: © 2023 The Authors. Pharmacology Research & Perspectives published by British Pharmacological Society and American Society for Pharmacology and Experimental Therapeutics and John Wiley & Sons Ltd. CC BY 4.0 This is an open access article under the terms of the http://creativecommons.org/licenses/by/4.0/ License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
Article links: DOI: 10.1002/prp2.1122 | PubMed: 37526235 | PMC: PMC10391716
Relevance: Moderate: mentioned 3+ times in text
Full text: PDF (830 KB)
INTRODUCTION
Gynecological cancers are cancers of the female reproductive system and consist of cancers of the cervix, ovaries, vagina, vulva, and uterus.ref. prp21122-bib-0001 Among gynecological cancers, ovarian cancer has the highest morbidity and mortality rates.ref. prp21122-bib-0002 Globally, the number of patients diagnosed in 2018 with gynecological cancers was >295 000 for ovarian. In 2018, approximately 185 000 women died from ovarian cancer.ref. prp21122-bib-0003 It is clear that there are unfavorable outcomes and that today, there are still unmet clinical needs.
Despite improvements in treatment strategies (Table 1), tumor recurrence, drug resistance, and toxic effects remain a major challenge, indicating the need for a new approach to the treatment of ovarian cancer. Recently, cannabinoids have gained attention as potential anti‐cancer therapies due to their ability to modulate the intracellular signaling pathways involved in cancer progression.ref. prp21122-bib-0004 However, the presence of psychoactive effects of cannabinoids could limit their progress in this field, therefore recent studies have highlighted the value of non‐psychoactive cannabinoids such as cannabidiol (CBD).ref. prp21122-bib-0005 CBD has been shown to demonstrate a broad array of anti‐carcinogenic properties, such as anti‐proliferative action toward breast cancer cells,ref. prp21122-bib-0006 anti‐invasive and anti‐metastatic effects on lung cancer cellsref. prp21122-bib-0007 as well as induction of apoptosis.ref. prp21122-bib-0008
TABLE 1: Current treatment strategies for ovarian cancer.prp21122-note-0001
| Surgery | First‐line chemotherapy | Maintenance treatment | |
|---|---|---|---|
| Stage I | Yes | Only for stage Ic or grade 3—six cycles of carboplatin | Not recommended |
| Stage II | Yes | Paclitaxel plus platinum‐based compound or platinum‐based therapy alone (cisplatin or carboplatin) | Not recommended |
| Stage III | Yes | Paclitaxel plus platinum‐based compound or platinum‐based therapy alone (cisplatin or carboplatin) | Olaparib plus bevacizumab or niraparib aloneOlaparib for BRCA mutation‐positive ovarian cancer |
| Stage IV | Yes | Paclitaxel plus platinum‐based compound or platinum‐based therapy alone (cisplatin or carboplatin) | Olaparib plus bevacizumab or niraparib aloneOlaparib for BRCA mutation‐positive ovarian cancer |
National Institute for Health and Care Excellence (NICE) Guidance Ovarian Cancer: Recognition and Initial Management (2011).9
In this review, the current knowledge on ovarian cancer treatment and the rationale for new therapies are discussed. In addition, this review provides a summary of the endocannabinoid system, the action of cannabinoids in cancer settings, and their anti‐tumor properties, as well as the effects of non‐psychoactive cannabinoids in ovarian cancer. Finally, recommendations for future work will be included in relation to their potential as a novel gynecological cancer treatment.
CURRENT CLINICAL STATUS OF OVARIAN CANCER AND RATIONALE FOR NOVEL THERAPIES
The current standard of care for patients with ovarian cancer is a combination of optimal cytoreductive surgery and platinum‐based chemotherapy.ref. prp21122-bib-0010 Ovarian cancer usually metastasizes within the peritoneal cavity within the abdomen first and in these cases, surgical debulking is used to inform staging and adjuvant therapy. Key advances in radical surgery and chemotherapy strategies have led to improved, yet modest, clinical outcomes. Despite this, there remains a significant risk of resistance to drug therapy and tumor recurrence. Due to the site of presentation and the shortage of promising screening tools for early‐stage detection, ovarian cancer often presents at a late stage, resulting in a poor 5‐year survival rate for these patients even with optimal care.ref. prp21122-bib-0010 Patients with stage III or IV disease have a 70%–75% chance of recurrence within 2 years of diagnosis. Treatment options are less effective at each recurrence, highlighting the need for novel maintenance therapies (Table 1).ref. prp21122-bib-0010 Further treatments after relapse are more intense, resulting in increased drug toxicity, drug resistance, and financial burden to patients with poor quality of life.ref. prp21122-bib-0011 Thus, there is a clear need for new and improved therapies to address the issues that remain with current conventional treatments.
THE HISTORY OF CANNABINOIDS
The first discovered and most important source of cannabinoids was the Cannabis sativa plant, which has been used as a herbal remedy for centuries. The earliest archeological evidence of the use of medicinal cannabis use was in ancient China where it was recommended for rheumatic pain, constipation, disorders of the female reproductive tract, and malaria, among other conditions.ref. prp21122-bib-0012 In Western medicine, the use of cannabis was introduced by William B. O’Shaughnessy (an Irish physician) and Jacques‐Joseph Moreau (a French psychiatrist) in the mid‐19th century. They described positive effects of cannabis preparations on pain, vomiting, convulsions, rheumatism, tetanus, and mental ability. From 1851, cannabis was recognized as a medicine in the United States (US) Pharmacopeia, in the form of tinctures, extracts, and resins. However, at the beginning of the 20th century, cannabis use decreased due to its increased use as a recreational drug, abuse potential, variability in the quality of herbal material, unidentified active compounds, and alternative medications with known efficacy being introduced to treat the same symptoms.ref. prp21122-bib-0013, ref. prp21122-bib-0014 In 1941, as the result of many legal restrictions, cannabis was considered to be in the same group as other illicit drugs and was removed from the American Pharmacopeia.ref. prp21122-bib-0014 Consequently, the exploration of medicinal uses of cannabis considerably slowed down for more than half a century.
However, in the late 20th century, studies conducted on cannabis led to the identification and extraction of pharmacologically active components named cannabinoids.ref. prp21122-bib-0015 The principal psychoactive constituent (or cannabinoid) of the cannabis plant is Δ9‐tetrahydrocannabinol (THC). This compound was first isolated, identified, and synthesized in 1964.ref. prp21122-bib-0016 Its discovery subsequently led to the identification of cannabinoid receptors and their endogenous ligands. Various therapeutic actions of these compounds have been reported and were thought to be mediated through the endocannabinoid system (ECS).ref. prp21122-bib-0017 Current legislative changes that allow cannabis for medical and/or recreational use, the progress in scientific research, and public awareness of the benefits of medicinal cannabis have all contributed to the increasing interest in the therapeutic potential of cannabinoids.ref. prp21122-bib-0018, ref. prp21122-bib-0019
TYPES OF CANNABINOIDS
As of 2020, over 150 cannabinoids have been identified.ref. prp21122-bib-0020 Cannabinoids comprise of (a) the active compounds of the Cannabis sativa plant (known as phytocannabinoids), (b) endogenous cannabinoids that are produced in our body (known as endocannabinoids), and (c) synthetic cannabinoids.ref. prp21122-bib-0021
Phytocannabinoids occur naturally in significant quantities in the cannabis plant and are concentrated in a viscous resin that is produced in glandular structures known as trichomes. Of all phytocannabinoids discovered so far, THC and CBD are the most abundant.ref. prp21122-bib-0022 Other phytocannabinoids include cannabigerol (CBG), cannabichromene (CBC), cannabicyclol (CBL), cannabivarin (CBV), tetrahydrocannabivarin (THCV), cannabidivarin (CBDV), cannabichromevarin (CBCV), cannabigerovarin (CBGV), and cannabigerol monoethyl ether (CBGM).ref. prp21122-bib-0021 As well as their analgesic, neuroprotective, and anxiolytic effects, studies have highlighted the therapeutic potential of phytocannabinoids for a variety of widespread skin conditions including acne, psoriasis, atopic dermatitis, and pruritis.ref. prp21122-bib-0023
Endocannabinoids are produced in our body and consist of lipid molecules containing long‐chain polyunsaturated fatty acids, amides, esters, and ethers that bind to cannabinoid receptors.ref. prp21122-bib-0024 Endocannabinoids act mainly as neuromodulators that affect the release of various neurotransmitters in the peripheral and neural tissues.ref. prp21122-bib-0025 They also play an important role in inflammation, insulin sensitivity, and fat and energy metabolism.ref. prp21122-bib-0026 Two of the best‐characterized endocannabinoids are N‐arachidonoylethanolamine (AEA‐anandamide) and 2‐arachidonoylglycerol (2‐AG), which are derived from arachidonic acid and affect our mood, appetite, pain sensation, inflammatory response, and memory.ref. prp21122-bib-0027, ref. prp21122-bib-0028
Synthetic cannabinoids such as the CB2 synthetic agonist, JWH‐133, and the CB1 and CB2 agonist, WIN‐55 have been widely used as a pharmacological agent, both in vitro and in vivo to obtain a more detailed understanding of cannabinoid action in order to evaluate their potential clinical use.ref. prp21122-bib-0029, ref. prp21122-bib-0030
THE ENDOCANNABINOID SYSTEM (ECS)
Historically, the two main cannabinoid receptors: cannabinoid receptor 1 (CB1) and cannabinoid receptor 2 (CB2), the main endocannabinoids: AEA‐anandamide and 2‐AG and the enzymes that produce and degrade these endocannabinoids have been known as ECS.ref. prp21122-bib-0031
In recent years, further components have widened this original definition of the ECS. These components consist of newly discovered endocannabinoid receptor ligands such as 2‐arachidonoyl glyceryl ether (noladin ether, 2‐AGE), O‐arachidonoylethanolamine (virodhamine), N‐arachidonoyldopamine (NADA) and oleic acid amide (oleamide, OA). In addition, newly discovered receptors such as G protein‐coupled receptor 55 (GPR55) and PPARs.ref. prp21122-bib-0032 However, other receptors have been recognized to participate in cannabinoid signaling. For example, it has been discovered that cannabinoids can affect a subset of transient receptor potential (TRP) channels.ref. prp21122-bib-0033 TRP subfamilies have been found to contain channels that can be modulated by endogenous, phytogenic and synthetic cannabinoids. TRP channels from these subfamilies have been reported to mediate cannabinoid activity.ref. prp21122-bib-0033 In addition to receptors and their cannabinoid ligands, the ECS encompasses several enzymes that regulate the biosynthesis and degradation of endocannabinoids. The catabolic enzyme primarily responsible for the degradation of AEA‐anandamide is fatty acid amide hydrolase (FAAH), while the main enzyme responsible for the degradation of 2‐AG is monoacylglycerol lipase (MAGL).ref. prp21122-bib-0031
The ECS plays an important role in the organism’s physiology. Dysregulation of the ECS due to variation in the expression and function of cannabinoid receptors, enzymes, or the concentration of endocannabinoids, has been associated with several diseases, such as neurodegenerative disorders, multiple sclerosis, inflammation, epilepsy, schizophrenia, glaucoma, cardiovascular diseases, obesity, and cancer.ref. prp21122-bib-0034, ref. prp21122-bib-0035
CANNABINOID RECEPTORS
Insights into the mechanism of action of phytocannabinoids led to the identification of two G protein‐coupled receptors, CB1 and CB2. ref. prp21122-bib-0036, ref. prp21122-bib-0037 CB1 is mainly expressed in the human CNS and is the main receptor responsible for the psychotropic effects of THC.ref. prp21122-bib-0038, ref. prp21122-bib-0039 Although to a lesser extent, CB1 receptors are also expressed at peripheral tissue sites where they aid in the regulation of local tissue functions.ref. prp21122-bib-0039 CB1 expression has been reported in adipose tissue, skeletal muscle, bone, skin, eye, reproductive system, and several types of cancer cells.ref. prp21122-bib-0040 A principal role for CB1 receptors is to inhibit neurotransmitter release. Strongly associated with GABAergic (inhibitory) and glutamergic (excitatory) cells, activation of CB1 receptors inhibits the release of GABA and glutamate, respectively.ref. prp21122-bib-0041 This decrease in excitability and neurotransmitter release may underlie some of the psychoactive and anti‐convulsant action of cannabinoids.
In contrast, CB2 receptors are predominantly expressed in peripheral tissues, such as the immune system, where they modulate immunological function, cell migration, and cytokine release.ref. prp21122-bib-0039, ref. prp21122-bib-0042 However, CB2 receptor expression has also been detected in the brain, however to a much lower extent compared to the immune system or the level of CB1 expression.ref. prp21122-bib-0039 CB2 activation is associated with neurodefense functions, ensuring the maintenance of bone mass and reduction of inflammation.ref. prp21122-bib-0043
In addition to the first discovered cannabinoid receptors CB1 and CB2, other cannabinoid receptors have since been identified that have responded to cannabinoid ligands, thus suggesting the existence of additional cannabinoid receptors.ref. prp21122-bib-0044 GPR55 has been identified as a novel cannabinoid receptor and has shown to interact with and be modulated by endogenous, plant, and synthetic cannabinoid ligands. AEA‐anandamide, the predominant circulating endocannabinoid, has been shown to activate GPR55 with a potency equivalent to that activating CB1 and CB2 receptors, demonstrating that this ligand has the potential to influence signaling by all three receptors equally.ref. prp21122-bib-0045 In addition, Overton et al.ref. prp21122-bib-0046 identified GPR119 as a cannabinoid receptor and it has been shown to be activated by endocannabinoids. 2‐AG has also been identified as a GPR119 agonist.ref. prp21122-bib-0047 GPR18 has been identified as a candidate cannabinoid receptor, but its classification is controversial. Several cannabinoid ligands have been described to be active as agonists or antagonists.ref. prp21122-bib-0048 N‐arachidonylglycine (NAGly) has been identified as an endogenous ligand for GPR18. However, NAGly does not have activity on the classical cannabinoid receptors CB1 and CB2. ref. prp21122-bib-0049 Therefore, it is difficult to determine whether GPR18 is a cannabinoid receptor.
CANNABINOIDS AND THEIR ANTI‐TUMOR EFFECTS
Cannabinoids have an established role in exerting palliative effects in cancer patients and have been used to alleviate nausea, vomiting, pain and to help stimulate appetite.ref. prp21122-bib-0005, ref. prp21122-bib-0050 Besides from providing symptomatic treatment for cancer patients, cannabinoids have been shown to exert anti‐tumor actions through modulation of the intracellular signaling pathway implicated in cancer progression.ref. prp21122-bib-0004, ref. prp21122-bib-0005 The first report on anti‐proliferative properties of cannabinoids was in 1975 when Munson et al.ref. prp21122-bib-0051 demonstrated that THC inhibits lung adenocarcinoma cell growth of in vitro cell lines and in murine models after oral administration. Cannabinoids can exert anti‐tumor effects directly through the inhibition of cell proliferation and induction of apoptosis or indirectly through the inhibition of angiogenesis, invasion, and metastasis.ref. prp21122-bib-0052 Numerous studies using synthetic/endo‐/phyto‐cannabinoids and ECS regulators in various cancer cell lines support this notion.ref. prp21122-bib-0053 The anti‐tumor effects of cannabinoids have also been observed in various animal tumor models.ref. prp21122-bib-0052
Endocannabinoids such as AEA‐anandamide have been found to have anti‐proliferative effects in prostate carcinomas. Through activation of CB1, AEA‐anandamide inhibited EGF‐induced proliferation of prostate carcinoma cells by decreasing the expression of the EGF receptor (EGFR) and increasing the production of ceramide (a powerful tumor suppressor).ref. prp21122-bib-0054 Phytocannabinoids such as THC have shown to reduce tumor proliferation and lung metastases, inhibit angiogenesis and cause apoptosis in a mouse model of ErbB2‐driven metastatic breast cancer. Caffarel et al. showed that these anti‐tumor actions relied at least partially on the inhibition of the pro‐tumorigenic AKT pathway, the signal transduction pathway that promotes cell growth, cell survival, and proliferation.ref. prp21122-bib-0055 This shows that cannabinoids can modulate certain pathways involved in cancer development and exert their anti‐tumor effects at the intracellular signaling level. Synthetic cannabinoids such as WIN‐55 have also shown anti‐proliferative effects on tumor progression.ref. prp21122-bib-0056
As cannabinoids are generally well tolerated and do not produce the typical toxic effects of conventional chemotherapy, there is considerable merit in their development as potential anti‐cancer therapies. However, the presence of psychoactive effects of cannabinoids could limit their progress in this field. Unfortunately, THC‐based drugs produce both therapeutic and undesirable psychotropic actions by activating CB1 receptors in the CNS. However, other cannabinoids such as CBD are devoid of the typical psychological effects. CBD constitutes up to 40% of cannabis extracts with pharmacological effects without producing undesirable psychoactive side effects.ref. prp21122-bib-0057 Non‐psychoactive cannabinoids have gained attention due to preclinically established anti‐cancer properties and a favorable risk–benefit profile. Recent studies have indicated the value of non‐psychoactive cannabinoids such as CBD.ref. prp21122-bib-0005
NON‐PSYCHOACTIVE CANNABINOIDS AND THEIR ANTI‐TUMOR ACTIVITY
CBD has demonstrated a broad array of anti‐carcinogenic properties. In glioma cells, CBD has been shown to successfully induce tumor cell death, inhibit cell migration and invasion in vitro, decrease tumor size, vascularization, growth, and weight, and induce tumor regression in vivo.ref. prp21122-bib-0008, ref. prp21122-bib-0058, ref. prp21122-bib-0059, ref. prp21122-bib-0060, ref. prp21122-bib-0061, ref. prp21122-bib-0062, ref. prp21122-bib-0063, ref. prp21122-bib-0064 CBD induced anti‐proliferative effects on breast cancer cells through a variety of mechanisms including apoptosis, autophagy, and cell cycle arrest.ref. prp21122-bib-0006, ref. prp21122-bib-0065, ref. prp21122-bib-0066 In aggressive breast cancer in vivo and in vitro, CBD has inhibited migration, invasion, and metastasis.ref. prp21122-bib-0006, ref. prp21122-bib-0067, ref. prp21122-bib-0068, ref. prp21122-bib-0069 Moreover, in lung cancer cells, CBD has demonstrated anti‐invasive and anti‐metastatic effects.ref. prp21122-bib-0007 In leukemias/lymphomas, CBD has mediated cell death by the mechanism of apoptosis.ref. prp21122-bib-0070, ref. prp21122-bib-0071, ref. prp21122-bib-0072, ref. prp21122-bib-0073 In prostate cancer cells, CBD has induced anti‐proliferative effects and apoptosis‐mediated cell death via the intrinsic pathway.ref. prp21122-bib-0074, ref. prp21122-bib-0075
CBG, another non‐psychoactive cannabinoid has also been shown to exhibit anti‐tumor properties in carcinomas. A recent study by Lah et al.ref. prp21122-bib-0076 was the first study to report the anti‐tumor effects of CBG in glioblastoma. They found that CBG effectively impaired the relevant hallmarks of glioblastoma progression and inhibited the invasion of glioblastoma cells. Borrelli et al. investigated whether CBG protects against colon tumourigenesis. The study showed that CBG stimulated ROS production, promoted apoptosis, and reduced cell growth in colorectal cancer cells.ref. prp21122-bib-0077 Table 2 provides a summary of the anti‐tumor effects of cannabinoids in different cancers.
TABLE 2: A summary of the anti‐tumor effects of cannabinoids in different cancers.
| Cancer type | Cannabinoid | Experimental system | Effect | Reference |
|---|---|---|---|---|
| Lung carcinoma | THC | In vitro and in vivo (mouse) | Anti‐proliferative | ref. 51 |
| CBD | In vitro and in vivo (mouse) | Anti‐invasive and anti‐metastatic | ref. 7 | |
| Prostate carcinoma | AEA‐anandamide | In vitro | Anti‐proliferative | ref. 54 |
| CBD | In vitro and in vivo | Anti‐proliferative and apoptosis | ref. 74, ref. 75 | |
| Breast carcinoma | CBD | In vitro and in vivo (mouse) | Anti‐proliferative, apoptosis, cell cycle arrest, and autophagy | ref. 6, ref. 65, ref. 66 |
| Aggressive breast carcinoma | CBD | In vitro and in vivo | Inhibition of migration, invasion, and metastasis | ref. 6, ref. 67, ref. 68, ref. 69 |
| ErbB2‐driven metastatic breast carcinoma | THC | In vivo (mouse) | Anti‐proliferative, reduced lung metastases, inhibition of angiogenesis, apoptosis | ref. 55 |
| Glioma | CBD | In vitro | Apoptosis, inhibition of cell migration and invasion | ref. 8, ref. 58, ref. 59, ref. 60, ref. 61, ref. 62, ref. 63, ref. 64 |
| In vivo | Decreased tumor size, vascularization, growth and weight, induction of tumor regression | |||
| Glioblastoma | CBG | In vitro | Anti‐invasive | ref. 76 |
| Leukemias/lymphomas | CBD | In vivo and in vitro | Apoptosis | ref. 70, ref. 71, ref. 72, ref. 73 |
| Colorectal carcinoma | CBG | In vivo | Anti‐proliferative, apoptosis | ref. 77 |
The accumulated data show that non‐psychoactive cannabinoids such as CBD and CBG have illustrated a range of anti‐cancer effects in a multitude of different cancer cell lines. Therefore, CBD and CBG are attracting pharmacological interest due to their non‐psychotropic nature, ability to inhibit cancer cell proliferation, and induction of apoptosis.
THE ROLE OF CANNABINOIDS IN MEDIATION OF APOPTOSIS
Apoptosis is a form of programmed cell death that is essential for the development and survival of organisms.ref. prp21122-bib-0078, ref. prp21122-bib-0079 Defects in the regulation of apoptotic cell death contribute to many diseases, including disorders in which cell accumulation occurs, such as cancer. The molecular machinery responsible for apoptosis has been revealed, uncovering a family of proteases, the caspases, which are accountable for the morphological and biochemical changes that characterize apoptosis.ref. prp21122-bib-0080, ref. prp21122-bib-0081 Regulators of the caspases have also been identified including activators and inhibitors of these cell death proteases. Through the discovery of inputs from signal transduction pathways into the core of the cell death machinery, ways of linking environmental stimuli to cell death responses or maintenance of cell survival have been demonstrated.ref. prp21122-bib-0081 The characteristics of the apoptotic cell include chromatin condensation, nuclear fragmentation, plasma membrane blebbing, and cell shrinkage. Eventually, the cell breaks into small membrane‐surrounded fragments known as apoptotic bodies which are cleared by phagocytosis without provoking an inflammatory response. Understanding the molecular mechanisms of apoptosis provides insight into the causes of pathologies where abnormal cell death regulation occurs, such as in cancer, and is beginning to provide novel approaches to the treatment of human diseases.ref. prp21122-bib-0081
Cannabinoids have been shown to activate apoptosis through CB1 or CB2 receptors. New evidence has reported that CBD promotes cell death in various gastric cancer cell lines.ref. prp21122-bib-0078, ref. prp21122-bib-0079, ref. prp21122-bib-0082 CBD has been shown to induce apoptotic cell death by suppressing X‐linked inhibitor of apoptosis (XIAP), a well‐characterized anti‐apoptotic protein, in a dose‐ and time‐dependent manner. CBD inhibited XIAP by stimulating stress‐related genes of the endoplasmic reticulum (ER) in gastric cancer cells.ref. prp21122-bib-0078, ref. prp21122-bib-0079 Zhang et al.ref. prp21122-bib-0082 showed that CBD treatment increased the protein levels of cleaved caspase‐3 and caspase‐9, subsequently inducing apoptosis cell death in gastric cancer cells. CBD increased Bax and decreased Bcl‐2 expression levels, causing a reduction of the Bcl‐2/Bax ratio. This in turn, determined an increase in mitochondrial membrane permeability and a decrease in mitochondrial transmembrane potential, thus allowing the release of cytochrome C into the cytosol and consequently, triggering apoptosis. Results from a recent study by Jeong et al.ref. prp21122-bib-0078, ref. prp21122-bib-0079 suggest that CBD can cause Noxa‐induced cell death in colorectal cancer cells. They reported that Noxa, a pro‐apoptotic member belonging to the Bcl‐2 protein family, is important for CBD‐induced apoptosis. In this study, CBD induced apoptotic cell death via ROS/ER stress‐regulated Noxa activation. Treatment with CBD increased Noxa in a dose‐ and time‐dependent manner. Noxa stimulated ROS production, which further exacerbated apoptosis. Endocannabinoids such as AEA‐anandamide and 2‐AG have shown to produce a dose‐dependent cell growth inhibitory effect in prostate cancer cells via the activation of CB1 receptors. Using Annexin V assays, it was shown that endocannabinoids induced apoptosis causing an increase in the levels of activated caspase‐3 and a reduction in the levels of Bcl‐2. In addition, endocannabinoid treatment activated the ERK pathway and simultaneously produced a decrease in the activation levels of the AKT pathway.ref. prp21122-bib-0083 Apoptosis by cannabinoids is not exclusively carried out by CB1 and CB2 receptors. Endocannabinoids such as AEA‐anandamide have been shown to induce apoptosis through transient receptor potential vanilloid subtype 1 (TRPV1) activation in human neuroblastoma and lymphoma cells. This effect occurred through oxidative stress, increased calcium influx, and activation of caspases 3 and 9.ref. prp21122-bib-0084, ref. prp21122-bib-0085 Furthermore, in cholangiocarcinoma cell lines, AEA‐anandamide exerted pro‐apoptotic activity, through the activation of GPR55 and induced apoptosis by the recruitment and activation of the death complex Fas/FasL.ref. prp21122-bib-0086
ECS IN THE OVARIES AND THEIR INTERACTIONS WITH FEMALE HORMONES
Studies have shown that CBD induced a reduction in cell proliferation in epithelial ovarian cancer cells, Kuramochi cell lines. Results from this study demonstrated that Kuramochi cell proliferation was 15% that of controls at CBD concentrations of 40 and 50 μM.ref. prp21122-bib-0087
CB1 receptors have been identified in normal, non‐cancerous ovaries.ref. prp21122-bib-0088 The presence of CB2 receptors has also been reported in the ovarian cortex, ovarian medulla, and ovarian follicles from human samples.ref. prp21122-bib-0088 Research has shown that expression of cannabinoid receptors changes as cancer progresses, specifically in ovarian cancer. Messalli et al. determined CB1 receptor expression in 66 human epithelial ovarian tumors and concluded that CB1 expression increased from benign and borderline to malignant tumors. They hypothesized that while low levels of cannabinoids may activate proliferative pathways in non‐cancerous cells, a higher cannabinoid concentration results in anti‐proliferative and apoptotic events in cancerous cells.ref. prp21122-bib-0089 Expression of the CB1 and CB2 receptors in cancerous cells, including the increase which was reported by Messalli et al. does not necessarily correlate with the expression pattern of the healthy tissue of origin.ref. prp21122-bib-0090 It is known that cannabinoid receptors and their endogenous ligands are generally upregulated in cancerous cells compared to non‐cancerous cells. Increased levels of cannabinoid receptor expression suggest that the administration of exogenous cannabinoids may impair tumor progression by inducing apoptosis.ref. prp21122-bib-0021, ref. prp21122-bib-0090 It is clear that the expression of cannabinoid receptors varies but these contradictory observations highlight the gap in the knowledge of the mechanism behind the regulation of cannabinoid receptors in malignancy. By achieving clarity in the regulation of the CB1, CB2, and GPR55 receptors at different stages and grades of the disease, this will aid in a more accurate understanding of the mechanism of action of non‐psychoactive cannabinoids on these receptors in ovarian cancer cells.
It can be argued whether the change in expression of cannabinoid receptors and their endogenous ligands such as endocannabinoids is causing cancer or if this change occurs as a result of cancer progression. A study by Hofman et al. investigated the involvement of the cancer cell‐derived GPR55 receptor agonist, L‐α‐lysophosphatidylinositol (LPI), on angiogenesis in ovarian cancer cell lines. They found that the GPR55 receptor agonist, L‐α‐lysophosphatidylinositol, mediated angiogenesis as pharmacological inhibition of GPR55 reduced the pro‐angiogenic potential of L‐α‐lysophosphatidylinositol in these cell lines. Interestingly, LPI activated only GPR55 receptors and not CB1 and CB2 receptors.ref. prp21122-bib-0091, ref. prp21122-bib-0092 Therefore, it can be concluded that inhibiting the pro‐angiogenic L‐α‐lysophosphatidylinositol/GPR55 pathway may be a promising target against angiogenesis in ovarian cancer.ref. prp21122-bib-0093 The expression of different components of the ECS is not uniform across all cancers; therefore, pharmacological manipulations of the ECS in further studies allow investigation into the link between the ECS and cancer progression.ref. prp21122-bib-0089, ref. prp21122-bib-0094
Events in the ovarian cycle are controlled by hormones secreted by the hypothalamus, the anterior pituitary, and the ovaries, collectively known as the hypothalamic–pituitary‐gonadal axis.ref. prp21122-bib-0095 The ECS has been closely linked to the hypothalamic–pituitary‐gonadal axis. CB1 receptors have been identified in the hypothalamus and anterior pituitary and CB1 and CB2 receptors are present in the ovaries.ref. prp21122-bib-0088, ref. prp21122-bib-0096, ref. prp21122-bib-0097 In multiple studies, cannabinoids such as WIN‐55, AEA‐anandamide, and THC have been shown to reduce the release of gonadotropin‐releasing hormone (GnRH) through a variety of mechanisms. For example, by indirectly modifying GnRH release by reducing the activity of neurotransmitters that facilitate GnRH release such as glutamate, whilst stimulating the activity of those known to down‐regulate GnRH secretion such as GABA, as well as by directly inhibiting hypothalamic release of GnRH.ref. prp21122-bib-0097, ref. prp21122-bib-0098, ref. prp21122-bib-0099, ref. prp21122-bib-0100, ref. prp21122-bib-0101, ref. prp21122-bib-0102, ref. prp21122-bib-0103 This reduction in GnRH, in turn, causes decreased release of luteinizing hormone (LH) and follicle‐stimulating hormone (FSH) from the anterior pituitary, resulting in decreased release of estrogen from the ovaries.ref. prp21122-bib-0104 Therefore, it is evident that endocannabinoids down‐regulate hypothalamic–pituitary‐gonadal activity and consequently, cause a reduction in estrogen levels.
Not only has research highlighted the effects of cannabinoids on gonadal hormones such as estrogen, but studies have also demonstrated the effects of estrogen on the ECS. In a study by MacCarrone et al., it was shown that in mouse uterus, estrogen decreased the activity of FAAH, the major enzyme responsible for the degradation of the endocannabinoid AEA‐anandamide, and, therefore, led to increased concentrations of cannabinoid.ref. prp21122-bib-0084, ref. prp21122-bib-0085 In another study, the administration of estradiol in ovariectomized female rats increased the levels of synthesized AEA‐anandamide in the medial basal hypothalamus indicating that estradiol could also increase endocannabinoid levels by directly interacting with endocannabinoid synthesis.ref. prp21122-bib-0102 Furthermore, another study showed that there was a positive correlation between the plasma peak levels of AEA‐anandamide and peak plasma 17β‐estradiol and gonadotrophin levels at ovulation.ref. prp21122-bib-0105 A possible underlying mechanism responsible for this could be that elevated levels of estrogens at ovulation inhibit FAAH activity, and as a consequence, increase plasma levels of endocannabinoids. Taken together, these studies show that, unlike the down‐regulation of hypothalamic–pituitary‐gonadal activity by endocannabinoids, it is evident that estrogen plays a role in modulating endocannabinoid signaling by up‐regulating cannabinoid content.ref. prp21122-bib-0106
Considering the well‐documented interactions between the ECS and gonadal hormones such as estrogens and the fact that these studies show that estrogen can modify cannabinoid levels and vice versa, it is not evident that the manipulation of the level of estrogen and, therefore, modification of the level of endocannabinoids may contribute to the progression of cancer. For example, in women who develop cancer while taking hormone‐replacement therapy or contraceptive pills, it could be argued that the manipulation of estrogen levels and therefore modification in the level of endocannabinoids could contribute to the progression of cancer. Although there is evidence that shows an interaction between estrogen and the ECS, more studies are required to investigate the link between estrogen manipulation and modification of endocannabinoid levels and the contribution of this to the development of gynecological cancers. This would ultimately enable us to establish the therapeutic potential of targeting the estrogen and ECS interaction as an approach for the treatment of gynecological cancers.
CANNABINOIDS IMPLICATION IN CELL DEATH THROUGH APOPTOSIS, CELL CYCLE ARREST, AND AUTOPHAGY IN OVARIAN CANCER
Research has highlighted key proteins and pathways involved in apoptosis that have been altered to evade cell death in ovarian cancer. Through comparing caspase‐3 and caspase‐8 levels in normal ovary, benign mass, and ovarian cancer, in vitro studies showed that caspase‐3 and caspase‐8 levels were lower in the benign mass and malignant group compared to the normal ovary group.ref. prp21122-bib-0107 Similarly, another study also showed that ovarian cancer tumors had low levels of caspase‐8 and were associated with shorter overall survival compared to tumors from patients that had high levels of caspase‐8.ref. prp21122-bib-0108 Therefore, since ovarian tumors have been shown to exhibit lower levels of these caspases, activating this pathway and thus increasing the levels of these caspases could provide a therapeutic strategy for inducing apoptosis in ovarian cancer cells. Survivin, a member of the Inhibitors of Apoptosis Proteins (IAP) family, blocks apoptosis by inhibiting caspase‐3 and caspase‐7. The over‐expression of survivin and, therefore, increased inhibition of caspase‐3 and caspase‐7 in ovarian cancer cell lines, IGROV‐1 and OAW42, highlighted its role in influencing cell sensitivity to taxanes.ref. prp21122-bib-0109 Histone deacetylase (HDAC) enzymes are a group of enzymes that are known to silence genes via catalyzing the removal of acetyl groups from histones as well as non‐histone proteins.ref. prp21122-bib-0110 In ovarian cancer, HDAC6 is often elevated leading to the inactivation of p53 apoptotic function. This was reversed when Bitler et al. used a small molecule HDAC6 inhibitor, ACY1215.ref. prp21122-bib-0111 The PI3K/AKT pathway is a key intracellular signal transduction pathway and has an important role in the regulation of apoptosis and cell survival. The loss of PTEN together with other mutations causes this pathway to be over‐expressed, resulting in reduced apoptosis.ref. prp21122-bib-0112, ref. prp21122-bib-0113 Enhanced expression of the PI3K/AKT pathway has been recognized as a hallmark of many cancers, including ovarian cancer.ref. prp21122-bib-0114
Key proteins involved in the regulation of cell cycle arrest have been identified to be exploited in ovarian cancer to promote cell cycle progression and inhibit cell cycle arrest. Cyclin E1 mainly coordinates with cyclin‐dependent kinase 2 (CDK2) to facilitate the progression of the G1/S cell cycle.ref. prp21122-bib-0115 In ovarian cancer cells, enforcing cyclin E1 expression stimulates cell proliferation,ref. prp21122-bib-0116 and over‐expression of cyclin E1 has been linked to the development of chemo‐resistance in ovarian cancer.ref. prp21122-bib-0117, ref. prp21122-bib-0118 It has been shown that ovarian tumors with elevated cyclin E1 levels often exhibit higher CDK2 expression.ref. prp21122-bib-0119, ref. prp21122-bib-0120 Studies show that the abundance of cyclin E1 correlates with tumor progression in patients with ovarian cancer.ref. prp21122-bib-0121, ref. prp21122-bib-0122, ref. prp21122-bib-0123, ref. prp21122-bib-0124 Mutations in the tumor suppressor gene, p53, have been detected in all histological types of epithelial ovarian cancer, and serous carcinomas, have been detected at higher frequencies. Alterations in the p53 network represent up to 96% of patients with high‐grade serous ovarian carcinoma.ref. prp21122-bib-0125 The loss of p53 function is another exploited mechanism that ovarian cancer cells deploy to inhibit cell cycle arrest and apoptosis. Dysregulation of the cell cycle signaling pathway CDK4/6‐cyclin D/p16‐Rb is one of the most common abnormalities in human cancer.ref. prp21122-bib-0126 Studies have identified that in ovarian cancer, p16 expression is most commonly altered due to promoter methylation.ref. prp21122-bib-0127, ref. prp21122-bib-0128, ref. prp21122-bib-0129 Overexpression of cyclin D1 has been described in ovarian cancer tumors and has been associated with a more aggressive tumor phenotype and poor prognosis.ref. prp21122-bib-0130 Mutations of the Rb gene have been reported in ovarian cancers.ref. prp21122-bib-0131, ref. prp21122-bib-0132 Together, these altered proteins and signaling pathways involved in the cell cycle regulation contribute to uncontrolled cell proliferation in ovarian cancer by avoiding cell cycle arrest.
Autophagy, from the Greek, meaning self‐eating refers to a cellular process committed to the lysosomal degradation of self‐constituents.ref. prp21122-bib-0133 Dysregulation of autophagy plays a significant role in the pathogenesis and resistance to radiotherapy and chemotherapy in ovarian cancer. A plethora of signaling pathways and proteins whose expression has been found altered in ovarian cancers have an impact on autophagy.ref. prp21122-bib-0134 For example, alterations in LC3, a component of the autophagy machinery. Compared to benign tissues and borderline ovarian tumors, highly malignant ovarian cancer cells were shown to express very low levels of LC3.ref. prp21122-bib-0135 Another example is an alteration in the signaling pathway, PI3K‐AKT‐mammalian target of rapamycin (mTOR). In ovarian cancer patients, a hyperactive state of mTOR has been associated with a poor prognosis.ref. prp21122-bib-0136 There is also sustained up‐regulation of the PI3K‐AKT–mTOR pathway in ovarian cancers that results in increased suppression of the autophagic process.ref. prp21122-bib-0137, ref. prp21122-bib-0138 Moreover, deletion of the gene that encodes Beclin‐1, another component of the autophagy machinery, has been identified in ovarian cancers.ref. prp21122-bib-0139 The expression of Beclin‐1 has been shown to be dramatically higher in benign and borderline ovarian tumors than those in malignant ovarian cancers.ref. prp21122-bib-0135 Inhibition of autophagy contributed to ovarian cancer development and was required to suppress Beclin‐1 and up‐regulate Bcl‐2.ref. prp21122-bib-0140
From the accumulated literature it is evident that cannabinoids affect many of the pathways and proteins involved in the evasion of apoptosis, cell cycle arrest, and autophagy in ovarian cancer (Figure 1). For example, in ovarian cancer, there are reduced levels of caspase‐3 and caspase‐8 contributing to inhibition of apoptosis. Cannabinoids can activate these caspases and thus activate apoptosis.ref. prp21122-bib-0004 The increased expression of the PI3K/AKT pathway contributing to inhibition of apoptosis in ovarian cancer cells is a pathway that is affected by cannabinoids. Cannabinoids can inhibit this pathway leading to activation of apoptosis. Some of the key proteins and pathways involved in cell cycle arrest that have been exploited in ovarian cancer to enable cell survival have also been shown to be affected by cannabinoids. For example, the increased activation of cyclin D leads to loss of Rb‐E2F, and thus inhibition of cell cycle arrest in ovarian cancer is a pathway that is affected by cannabinoids. Through activation of p27, cannabinoids can inhibit cyclin D that leads to activation of Rb‐E2F and therefore activation of cell cycle arrest. Similarly, the increased activation of cyclin E and, consequently, increased activation of CDK2 in ovarian cancer are proteins that are affected by cannabinoids. Through multiple mechanisms, cannabinoids can inhibit the activity of cyclin E and CDK2 and therefore allow activation of cell cycle arrest. It has also been shown that cannabinoids can affect some of the key proteins and pathways involved in autophagy that have been exploited in ovarian cancer to enable cell survival. One of these proteins being LC3. The reduced levels of LC3 in ovarian cancer inhibit autophagy, however, cannabinoids such as CBD and AEA‐anandamide have been shown to activate this protein, resulting in the activation of autophagy. Another pathway that cannabinoids can affect is the PI3K‐AKT–mTOR pathway. In ovarian cancer, this pathway is upregulated as a mechanism of inhibiting autophagy, however, it has been shown that cannabinoids inhibit this pathway leading to activation of autophagy. Therefore, the accumulated data indicate that cannabinoids have a role in the mediation of cell death through apoptosis, cell cycle arrest, and autophagy in ovarian cancer and could provide a therapeutic strategy that targets the ability to evade cell death in ovarian cancer (Figure 1).
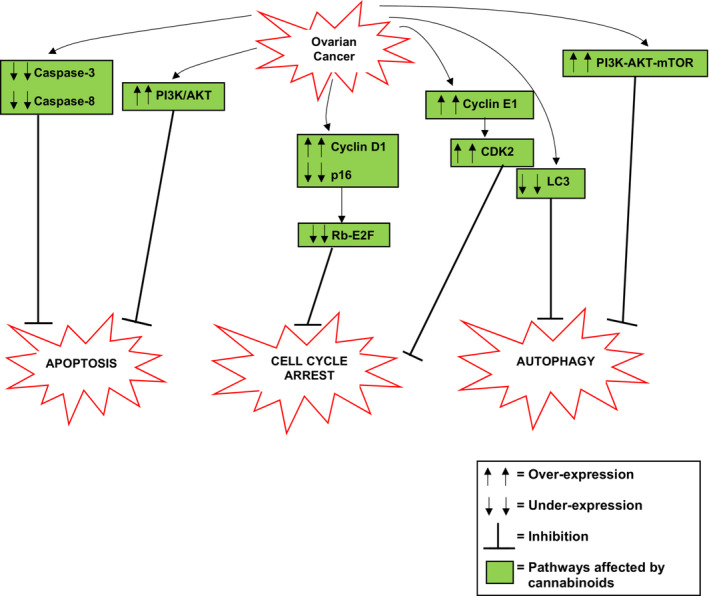
CONCLUSION AND FUTURE DIRECTION
The current clinical status of gynecological cancers inevitably motivates researchers to identify and explore new and improved therapies for patients. Pre‐clinical studies suggest that cannabinoids may exert favorable outcomes in the treatment of cancers. Although the presence of psychoactive effects limits the progression of cannabinoids into clinical practice, the use of non‐psychoactive cannabinoids that are devoid of these adverse effects such as CBD provides a solution.
The anti‐proliferative and pro‐apoptotic effects of cannabinoids are well established. However, the relationship between the effects of cannabinoids and the expression of cannabinoid receptors in gynecological malignancy needs further investigation. The introduction of cannabinoid‐based medications such as sativex and epidiolex (CBD) is encouraging. Currently, sativex and epidiolex are the only cannabinoid‐based medications that are licensed for use in the UK (British National Formulary, 2022).ref. prp21122-bib-0141 Sativex is available in the form of an oromucosal spray and contains both THC and CBD in an approximate 1:1 ratio. Sativex is indicated for adjunctive therapy in the treatment of moderate to severe spasticity in multiple sclerosis (Electronic Medicines Compendium, 2022).ref. prp21122-bib-0142 Epidiolex is in the form of an oral solution and contains CBD. Epidiolex is indicated for adjunctive therapy in the treatment of seizures associated with Lennox–Gastaut syndrome or Dravet syndrome to be used in conjunction with clobazam. Epidiolex is also indicated for adjunctive therapy in the treatment of seizures associated with the tuberous sclerosis complex (Electronic Medicines Compendium, 2022).ref. prp21122-bib-0143 Both drugs were well tolerated with minimum and manageable side effects.ref. prp21122-bib-0144, ref. prp21122-bib-0145, ref. prp21122-bib-0146, ref. prp21122-bib-0147, ref. prp21122-bib-0148, ref. prp21122-bib-0149, ref. prp21122-bib-0150, ref. prp21122-bib-0151 This has encouraged a UK randomized clinical trial to test the use of sativex in patients with glioblastoma. The trial is aimed at measuring whether adding sativex to chemotherapy extends overall patient survival, delays the progression of the disease, or improves quality of life.
With the encouraging pre‐clinical results on anti‐tumor effects of cannabinoids in ovarian cancer, further research is needed to complement the existing knowledge and enable the translation of cannabinoids for cancer treatment into the clinic. This should be followed by the initiation of clinical trials to re‐purpose sativex and epidiolex in ovarian cancer patients.
In addition, future work should aim to understand the change in the level of estrogen on the level of the cannabinoid system and vice versa in gynecological cancers. Nevertheless, the accumulated data support further studies on the use of cannabinoids as a potential candidate in the treatment strategy for ovarian cancer. This review serves as a platform upon which existing knowledge and research can be built upon to ultimately establish if non‐psychoactive cannabinoids have the potential of becoming a part of an effective ovarian cancer strategy.
AUTHOR CONTRIBUTIONS
Both authors contributed to all aspects of writing.
CONFLICT OF INTEREST STATEMENT
The authors declare that they have no conflict of interest.
ETHICS STATEMENT
Not applicable.
References
- Gynaecological cancers .. 2021
- Cancer statistics, 2018.. Cancer J Clin., 2018. [DOI]
- Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries.. Cancer J Clin., 2018. [DOI]
- Molecular mechanism of cannabinoids in cancer progression.. Int J Mol Sci., 2021. [DOI | PubMed]
- Cannabinoid pharmacology in cancer research: a new hope for cancer patients?. Eur J Pharmacol., 2016. [DOI]
- Antitumor activity of plant cannabinoids with emphasis on the effect of cannabidiol on human breast carcinoma.. J Pharmacol Exp Ther., 2006. [DOI | PubMed]
- Cannabidiol inhibits lung cancer cell invasion and metastasis via intercellular adhesion molecule‐1.. FASEB J., 2012. [DOI | PubMed]
- Antitumor effects of cannabidiol, a nonpsychoactive cannabinoid, on human glioma cell lines.. J Pharmacol Exp Ther., 2004. [DOI | PubMed]
- Ovarian Cancer: Recognition and Initial Management .. 2011
- Current ovarian cancer maintenance strategies and promising new developments.. J Cancer., 2021. [DOI | PubMed]
- Ovarian cancer: current status and strategies for improving therapeutic outcomes.. Cancer Med., 2019. [DOI | PubMed]
- The religious and medicinal uses of cannabis in China, India and Tibet.. J Psychoactive Drugs., 1981. [DOI | PubMed]
- The classification of substance use disorders: historical, contextual, and conceptual considerations.. Behav Sci., 2016. [DOI | PubMed]
- History of cannabis as a medicine: a review.. Rev Bras Psiquiatr., 2006. [DOI | PubMed]
- Biomedical benefits of cannabinoids?. Addict Biol., 1999. [DOI | PubMed]
- Isolation, structure and partial synthesis of an active constituent of hashish.. J Am Chem Soc., 1964. [DOI]
- The role of endocannabinoids system in fatty liver disease and therapeutic potentials.. Saudi J Gastroenterol., 2013. [DOI | PubMed]
- Medicinal use of cannabis in Europe: the fact that more countries legalize the medicinal use of cannabis should not become an argument for unfettered and uncontrolled use.. EMBO Rep., 2015. [DOI | PubMed]
- Medical cannabis ‐ the Canadian perspective.. J Pain Res., 2016. [DOI | PubMed]
- Analysis of impurities of cannabidiol from hemp. Isolation, characterization and synthesis of cannabidibutol, the novel cannabidiol butyl analog.. J Pharm Biomed Anal., 2019. [DOI]
- Cannabinoids as therapeutic agents in cancer: current status and future implications.. Oncotarget., 2014. [DOI | PubMed]
- The evolving science of phytocannabinoids.. Nat Rev Chem., 2017. [DOI]
- Cannabis sativa and skin health: dissecting the role of Phytocannabinoids.. Planta Med., 2021. [DOI | PubMed]
- Cannabimimetic fatty acid derivatives in cancer and inflammation.. Prostaglandins Other Lipid Mediat., 2000. [DOI | PubMed]
- Neurobiology cannabinoids act backwards.. Nature., 2001. [DOI | PubMed]
- The endocannabinoid system and its role in regulation of metabolism in peripheral tissues.. Postepy Biochem., 2012. [PubMed]
- Isolation and structure of a brain constituent that binds to the cannabinoid receptor.. Science., 1992. [DOI | PubMed]
- Identification of an endogenous 2‐monoglyceride, present in canine gut, that binds to cannabinoid receptors.. Biochem Pharmacol., 1995. [DOI | PubMed]
- Crosstalk between chemokine receptor CXCR4 and cannabinoid receptor CB2 in modulating breast cancer growth and invasion.. PLoS One., 2011. [DOI | PubMed]
- Synthetic cannabinoid receptor agonists inhibit tumor growth and metastasis of breast cancer.. Mol Cancer Ther., 2009. [DOI | PubMed]
- The endocannabinoid system: a target for cancer treatment.. Int J Mol Sci., 2020. [DOI | PubMed]
- Modulation of the endocannabinoid system as a potential anticancer strategy.. Front Pharmacol., 2019. [DOI | PubMed]
- Cannabinoid ligands targeting TRP channels.. Front Mol Neurosci., 2019. [DOI | PubMed]
- The therapeutic aspects of the endocannabinoid system (ECS) for cancer and their development: from nature to laboratory.. Curr Pharm Des., 2016. [DOI | PubMed]
- Therapeutic applications of cannabinoids.. Chem Biol Interact., 2018. [DOI | PubMed]
- Structure of a cannabinoid receptor and functional expression of the cloned cDNA.. Nature., 1990. [DOI | PubMed]
- Molecular characterization of a peripheral receptor for cannabinoids.. Nature., 1993. [DOI | PubMed]
- Cannabinoids and the expanded endocannabinoid system in neurological disorders.. Nat Rev Neurol., 2020. [DOI | PubMed]
- Cannabinoid receptors and the endocannabinoid system: signaling and function in the central nervous system.. Int J Mol Sci., 2018. [DOI | PubMed]
- Endocannabinoid signaling at the periphery: 50 years after THC.. Trends Pharmacol Sci., 2015. [DOI | PubMed]
- The therapeutic potential of cannabis.. Lancet Neurol., 2003. [DOI | PubMed]
- Progress in brain cannabinoid CB2 receptor research: from genes to behaviour.. Neurosci Biobehav Rev., 2019. [DOI | PubMed]
- The endocannabinoid pathway in Huntington’s disease: a comparison with other neurodegenerative diseases.. Prog Neurobiol., 2007. [DOI | PubMed]
- Evidence for novel cannabinoid receptors.. Pharmacol Ther., 2005. [DOI | PubMed]
- The orphan receptor GPR55 is a novel cannabinoid receptor.. Br J Pharmacol., 2007. [DOI | PubMed]
- Deorphanization of a G protein‐coupled receptor for oleoylethanolamide and its use in the discovery of small‐molecule hypophagic agents.. Cell Metab., 2006. [DOI | PubMed]
- 2‐Oleoyl glycerol is a GPR119 agonist and signals GLP‐1 release in humans.. J Clin Endocrinol Metab., 2011. [DOI | PubMed]
- Δ(9)‐tetrahydrocannabinol and N‐arachidonyl glycine are full agonists at GPR18 receptors and induce migration in human endometrial HEC‐1B cells.. Br J Pharmacol., 2012. [DOI | PubMed]
- Structural requirements for binding of anandamide‐type compounds to the brain cannabinoid receptor.. J Med Chem., 1997. [DOI | PubMed]
- Cannabinoids for symptom management and cancer therapy: the evidence.. J Natl Compr Cancer Netw., 2016. [DOI]
- Antineoplastic activity of cannabinoids.. J Natl Cancer Inst., 1975. [DOI | PubMed]
- Towards the use of cannabinoids as antitumour agents.. Nat Rev Cancer., 2012. [DOI | PubMed]
- The endocannabinoid signaling system in cancer.. Trends Pharmacol Sci., 2013. [DOI | PubMed]
- Anti‐proliferative and apoptotic effects of anandamide in human prostatic cancer cell lines: implication of epidermal growth factor receptor down‐regulation and ceramide production.. Prostate., 2003. [DOI | PubMed]
- Cannabinoids reduce ErbB2‐driven breast cancer progression through Akt inhibition.. Mol Cancer., 2010. [DOI | PubMed]
- An introduction to the endocannabinoid system: from the early to the latest concepts.. Best Pract Res Clin Endocrinol Metab., 2009. [DOI | PubMed]
- Cannabidiol—Recent advances.. Chem Biodivers., 2007. [DOI | PubMed]
- Local delivery of cannabinoid‐loaded microparticles inhibits tumor growth in a murine xenograft model of glioblastoma multiforme.. PLoS One., 2013. [DOI | PubMed]
- Targeting glioma initiating cells with a combined therapy of cannabinoids and temozolomide.. Biochem Pharmacol., 2018. [DOI | PubMed]
- Optimization of a preclinical therapy of cannabinoids in combination with temozolomide against glioma.. Biochem Pharmacol., 2018. [DOI | PubMed]
- 5‐lipoxygenase and anandamide hydrolase (FAAH) mediate the antitumor activity of cannabidiol, a non‐psychoactive cannabinoid.. J Neurochem., 2008. [DOI | PubMed]
- The combination of cannabidiol and Δ9‐tetrahydrocannabinol enhances the anticancer effects of radiation in an orthotopic murine glioma model.. Mol Cancer Ther., 2014. [DOI | PubMed]
- Cannabidiol, a non‐psychoactive cannabinoid compound, inhibits proliferation and invasion in U87‐MG and T98G glioma cells through a multitarget effect.. PLoS One., 2013. [DOI | PubMed]
- A combined preclinical therapy of cannabinoids and temozolomide against glioma.. Mol Cancer Ther., 2011. [DOI | PubMed]
- Cannabidiol induces programmed cell death in breast cancer cells by coordinating the cross‐talk between apoptosis and autophagy.. Mol Cancer Ther., 2011. [DOI | PubMed]
- Novel mechanism of cannabidiol‐induced apoptosis in breast cancer cell lines.. Breast., 2018. [DOI | PubMed]
- Modulation of the tumor microenvironment and inhibition of EGF/EGFR pathway: novel anti‐tumor mechanisms of Cannabidiol in breast cancer.. Mol Oncol., 2015. [DOI | PubMed]
- Pathways mediating the effects of cannabidiol on the reduction of breast cancer cell proliferation, invasion, and metastasis.. Breast Cancer Res Treat., 2011. [DOI | PubMed]
- Targeting multiple cannabinoid anti‐tumour pathways with a resorcinol derivative leads to inhibition of advanced stages of breast cancer.. Br J Pharmacol., 2014. [DOI | PubMed]
- γ‐Irradiation enhances apoptosis induced by Cannabidiol, a non‐psychotropic cannabinoid, in cultured HL‐60 myeloblastic leukemia cells.. Leuk Lymphoma., 2003. [DOI | PubMed]
- Cannabidiol‐induced apoptosis in human leukemia cells: a novel role of Cannabidiol in the regulation of p22phox and Nox4 expression.. Mol Pharmacol., 2006. [DOI | PubMed]
- Cannabidiol directly targets mitochondria and disturbs calcium homeostasis in acute lymphoblastic leukemia.. Cell Death Dis., 2019. [DOI]
- Anticancer effects of phytocannabinoids used with chemotherapy in leukaemia cells can be improved by altering the sequence of their administration.. Int J Oncol., 2017. [DOI | PubMed]
- Non‐THC cannabinoids inhibit prostate carcinoma growth in vitro and in vivo: pro‐apoptotic effects and underlying mechanisms.. Br J Pharmacol., 2013. [DOI | PubMed]
- In vitro anticancer activity of plant‐derived cannabidiol on prostate cancer cell lines.. Pharmacol Pharm., 2014. [DOI]
- Cannabigerol is a potential therapeutic agent in a novel combined therapy for glioblastoma.. Cell., 2021. [DOI]
- Colon carcinogenesis is inhibited by the TRPM8 antagonist cannabigerol, a cannabis‐derived non‐psychotropic cannabinoid.. Carcinogenesis., 2014. [DOI | PubMed]
- Cannabidiol promotes apoptosis via regulation of XIAP/Smac in gastric cancer.. Cell Death Dis., 2019. [DOI]
- Cannabidiol‐induced apoptosis is mediated by activation of Noxa in human colorectal cancer cells.. Cancer Lett., 2019. [DOI | PubMed]
- Human ICE/CED‐3 protease nomenclature.. Cell., 1996. [DOI | PubMed]
- Mechanisms of apoptosis.. Am J Pathol., 2000. [DOI | PubMed]
- Cannabidiol induces cell cycle arrest and cell apoptosis in human gastric cancer SGC‐7901 cells.. Biomolecules., 2019. [DOI | PubMed]
- Proapoptotic effect of endocannabinoids in prostate cancer cells.. Oncol Rep., 2015. [DOI | PubMed]
- Down‐regulation of anandamide hydrolase in mouse uterus by sex hormones.. Eur J Biochem., 2000. [DOI | PubMed]
- Anandamide induces apoptosis in human cells via vanilloid receptors. Evidence for a protective role of cannabinoid receptors.. J Biol Chem., 2000. [DOI | PubMed]
- Anandamide exerts its antiproliferative actions on cholangiocarcinoma by activation of the GPR55 receptor.. Lab Investig., 2011. [DOI | PubMed]
- Cannabidiol: Assessing preclinical safety in ovarian and endometrial carcinoma cell lines.. J Clin Oncol., 2020. [DOI]
- Localisation and function of the endocannabinoid system in the human ovary.. PLoS One., 2009. [DOI | PubMed]
- Cannabinoid receptor type 1 immunoreactivity and disease severity in human epithelial ovarian tumors.. Am J Obstet Gynecol., 2014. [DOI]
- Anticancer mechanisms of cannabinoids.. Curr Oncol., 2016. [DOI]
- The GPR55 ligand L‐alpha‐lysophosphatidylinositol promotes RhoA‐dependent Ca2+ signaling and NFAT activation.. FASEB J., 2009. [PubMed]
- Identification of GPR55asalysophosphatidylinositolreceptor.. Biochem Biophys Res Commun., 2007. [PubMed]
- The GPR 55 agonist, L‐α‐lysophosphatidylinositol, mediates ovarian carcinoma cell‐induced angiogenesis.. Br J Pharmacol., 2015. [DOI | PubMed]
- Endocannabinoid system as a regulator of tumor cell malignancy ‐ biological pathways and clinical significance.. Onco Targets Ther., 2016. [DOI | PubMed]
- 95 Meccariello R , Chianese R , Fasano S , Pierantoni R . Endocannabinoids and kisspeptins: two modulators in fight for the regulation of GnRH activity. In: Vizcarra J , ed. Gonadotropin. IntechOpen; 2013:57‐88. doi:10.5772/48443
- Endocannabinoid signaling in mammalian ovary.. Eur J Obstet Gynecol Reprod Biol., 2014. [DOI | PubMed]
- Regulation of gonadotropin‐releasing hormone secretion by cannabinoids.. Endocrinology., 2005. [DOI | PubMed]
- Mode of action of delta‐9‐tetrahydrocannabinol on hypothalamo‐pituitary function in adult female rats.. J Reprod Fertil., 1979. [DOI | PubMed]
- Male‐female differences in the effects of cannabinoids on sexual behaviour and gonadal hormone function.. Horm Behav., 2010. [DOI | PubMed]
- Dual‐phenotype GABA/glutamate neurons in adult preoptic area: sexual dimorphism and function.. J Neurosci., 2004. [DOI | PubMed]
- Endocannabinoid control of glutamate NMDA receptors: the therapeutic potential and consequences of dysfunction.. Oncotarget., 2016. [DOI | PubMed]
- The inhibitory effect of anandamide on luteinizing hormone‐releasing hormone secretion is reversed by estrogen.. Proc Natl Acad Sci U S A., 2004. [DOI | PubMed]
- The influence of anandamide on the anterior pituitary hormone secretion in ewes‐ex vivo study.. Animals., 2020. [DOI | PubMed]
- The role of the endocannabinoid system in female reproductive tissues.. J Ovarian Res., 2019. [DOI | PubMed]
- The relationship between plasma levels of the endocannabinoid, anandamide, sex steroids, and gonadotrophins during the menstrual cycle.. Fertil Steril., 2010. [DOI | PubMed]
- Minireview: endocannabinoids and gonadal hormones: bidirectional interactions in physiology and behavior.. Endocrinology., 2012. [DOI | PubMed]
- Comparative expression of caspases and annexin V in benign and malignant ovarian tumors.. J Cancer Res Ther., 2018. [DOI | PubMed]
- Caspase 8 expression may determine the survival of women with ovarian cancer.. Cell Death Dis., 2016. [DOI | PubMed]
- Expression of the anti‐apoptotic gene survivin correlates with taxol resistance in human ovarian cancer.. Cell Mol Life Sci., 2002. [DOI | PubMed]
- Histone deacetylase inhibitors: molecular mechanisms of action and clinical trials as anti‐cancer drugs.. Am J Transl Res., 2011. [PubMed]
- ARID1A‐mutated ovarian cancers depend on HDAC6 activity.. Nat Cell Biol., 2017. [DOI | PubMed]
- The Akt/PKB pathway: molecular target for cancer drug discovery.. Oncogene., 2005. [DOI | PubMed]
- The role of genetic abnormalities of PTEN and the phosphatidylinositol 3‐kinase pathway in breast and ovarian tumorigenesis, prognosis, and therapy.. Semin Oncol., 2001. [DOI | PubMed]
- AKT and mTOR phosphorylation is frequently detected in ovarian cancer and can be targeted to disrupt ovarian tumor cell growth.. Oncogene., 2004. [DOI | PubMed]
- An integrated view of cyclin E function and regulation.. Cell Cycle., 2012. [DOI | PubMed]
- Gene amplification CCNE1 is related to poor survival and potential therapeutic target in ovarian cancer.. Cancer., 2010. [DOI | PubMed]
- Integrated genome‐wide DNA copy number and expression analysis identifies distinct mechanisms of primary chemoresistance in ovarian carcinomas.. Clin Cancer Res., 2009. [DOI | PubMed]
- Amplicon‐dependent CCNE1 expression is critical for clonogenic survival after cisplatin treatment and is correlated with 20q11 gain in ovarian cancer.. PLoS One., 2010. [DOI | PubMed]
- Cyclin E1 deregulation occurs early in secretory cell transformation to promote formation of fallopian tube‐derived high‐grade serous ovarian cancers.. Cancer Res., 2014. [DOI | PubMed]
- Analysis of cyclin E and CDK2 in ovarian cancer: gene amplification and RNA overexpression.. Int J Cancer., 1998. [DOI | PubMed]
- Polyploid giant cancer cells with budding and the expression of cyclin E, S‐phase kinase‐associated protein 2, stathmin associated with the grading and metastasis in serous ovarian tumor.. BMC Cancer., 2014. [DOI | PubMed]
- Cyclin E1 (CCNE1) as independent positive prognostic factor in advanced stage serous ovarian cancer patients ‐ a study of the OVCAD consortium.. Eur J Cancer., 2014. [DOI | PubMed]
- Cyclin E expression is correlated with tumor progression and predicts a poor prognosis in patients with ovarian carcinoma.. Cancer., 2006. [DOI | PubMed]
- Implication of malignancy and prognosis of p27(kip1), cyclin E, and Cdk2 expression in epithelial ovarian tumors.. Gynecol Oncol., 2001. [DOI | PubMed]
- The hallmarks of ovarian cancer: proliferation and cell growth.. EJC Suppl., 2020. [DOI | PubMed]
- The hallmarks of cancer.. Cell., 2000. [DOI | PubMed]
- Methylation of tumor suppressor gene p16 and prognosis of epithelial ovarian cancer.. Gynecol Oncol., 2004. [DOI | PubMed]
- Inactivation of p16/CDKN2 and p15/MTS2 is associated with prognosis and response to chemotherapy in ovarian cancer.. Int J Cancer., 2002. [DOI | PubMed]
- Characterization of chromosome 9 in human ovarian neoplasia identifies frequent genetic imbalance on 9q and rare alterations involving 9p, including CDKN2.. Cancer Res., 1995. [PubMed]
- Cyclin D1, p53, and p21Waf1/Cip1 expression is predictive of poor clinical outcome in serous epithelial ovarian cancer.. Clin Cancer Res., 2004. [DOI | PubMed]
- Evidence of functional RB protein in epithelial ovarian carcinomas despite loss of heterozygosity at the RB locus.. Cancer Res., 1994. [PubMed]
- Loss of heterozygosity at the RB‐1 locus and pRB immunostaining in epithelial ovarian tumors: a molecular, immunohistochemical, and clinicopathologic study.. Int J Gynecol Pathol., 2001. [DOI | PubMed]
- Eaten alive: a history of macroautophagy.. Nat Cell Biol., 2010. [DOI | PubMed]
- Involvement of autophagy in ovarian cancer: a working hypothesis.. J Ovarian Res., 2012. [DOI | PubMed]
- Decreased expression of autophagy‐related proteins in malignant epithelial ovarian cancer.. Autophagy., 2008. [DOI | PubMed]
- Mammalian target of rapamycin is a biomarker of poor survival in metastatic serous ovarian carcinoma.. Hum Pathol., 2010. [DOI | PubMed]
- Sequence mutations and amplification of PIK3CA and AKT2 genes in purified ovarian serous neoplasms.. Cancer Biol Ther., 2006. [DOI | PubMed]
- The oncogenic mutation in the pleckstrin homology domain of AKT1 in endometrial carcinomas.. Br J Cancer., 2009. [DOI | PubMed]
- Cloning and genomic organization of beclin 1, a candidate tumor suppressor gene on chromosome 17q21.. Genomics., 1999. [DOI | PubMed]
- Autophagy inhibitor LRPPRC suppresses mitophagy through interaction with mitophagy initiator Parkin.. PLoS One., 2014. [DOI | PubMed]
- 141 Joint Formulary Committee . British National Formulary (online). BMJ Group and Pharmaceutical Press. 2022. Accessed June 7, 2022. http://www.medicinescomplete.com
- Sativex Oromucosal Spray GW Pharma Ltd .. 2022
- Epidyolex 100mg/ml Oral Solution GW Pharma Ltd .. 2022. Accessed June 06, 2022.
- Long‐term safety and efficacy of cannabidiol in children and adults with treatment resistant Lennox‐Gastaut syndrome or Dravet syndrome: expanded access program results.. Epilepsy Res., 2019. [DOI | PubMed]
- Cannabidiol drugs clinical trial outcomes and adverse effects.. Front Pharmacol., 2020. [DOI | PubMed]
- Safety and tolerability of nabiximols oromucosal spray: a review of more than 15 years accumulated evidence from clinical trials.. Expert Rev Neurother., 2021. [DOI | PubMed]
- THC and CBD oromucosal spray (Sativex®) in the management of spasticity associated with multiple sclerosis.. Expert Rev Neurother., 2011. [DOI | PubMed]
- Abuse potential assessment of cannabidiol (CBD) in recreational polydrug users: a randomized, double‐blind, controlled trial.. Epilepsy Behav., 2018. [DOI | PubMed]
- Sativex long‐term use: an open‐label trial in patients with spasticity due to multiple sclerosis.. J Neurol., 2013. [DOI | PubMed]
- Sativex in the treatment of pain of neurological origin or symptoms of multiple sclerosis: interim analysis of a long‐term, open label, safety and tolerability study.. J Pain., 2005. [DOI]
- Cannabidiol in patients with seizures associated with Lennox‐Gastaut syndrome (GWPCARE4): a randomised, double‐blind, placebo‐controlled phase 3 trial.. Lancet., 2018. [DOI | PubMed]