
GABAA receptors: structure, function, pharmacology, and related disorders
Abstract
Background:
γ-Aminobutyric acid sub-type A receptors (GABAARs) are the most prominent inhibitory neurotransmitter receptors in the CNS. They are a family of ligand-gated ion channel with significant physiological and therapeutic implications.
Main body:
GABAARs are heteropentamers formed from a selection of 19 subunits: six α (alpha1-6), three β (beta1-3), three γ (gamma1-3), three ρ (rho1-3), and one each of the δ (delta), ε (epsilon), π (pi), and θ (theta) which result in the production of a considerable number of receptor isoforms. Each isoform exhibits distinct pharmacological and physiological properties. However, the majority of GABAARs are composed of two α subunits, two β subunits, and one γ subunit arranged as γ2β2α1β2α1 counterclockwise around the center. The mature receptor has a central chloride ion channel gated by GABA neurotransmitter and modulated by a variety of different drugs. Changes in GABA synthesis or release may have a significant effect on normal brain function. Furthermore, The molecular interactions and pharmacological effects caused by drugs are extremely complex. This is due to the structural heterogeneity of the receptors, and the existence of multiple allosteric binding sites as well as a wide range of ligands that can bind to them. Notably, dysfunction of the GABAergic system contributes to the development of several diseases. Therefore, understanding the relationship between GABAA receptor deficits and CNS disorders thus has a significant impact on the discovery of disease pathogenesis and drug development.
Conclusion:
To date, few reviews have discussed GABAA receptors in detail. Accordingly, this review aims to summarize the current understanding of the structural, physiological, and pharmacological properties of GABAARs, as well as shedding light on the most common associated disorders.
Article type: Review Article
Keywords: GABA, GABA, Benzodiazepine, Barbiturates, Allosteric modulation, Autism spectrum disorder, Alzheimer’s disease, Epilepsy, Schizophrenia
Affiliations: grid.8982.b0000 0004 1762 5736Department of Biology and Biotechnology, University of Pavia, Pavia, Italy; grid.7155.60000 0001 2260 6941Department of Biotechnology, Institute of Graduate Studies and Research (IGSR), Alexandria University, Alexandria, Egypt; grid.252119.c0000 0004 0513 1456Department of Biotechnology, American University in Cairo (AUC), Cairo, Egypt; grid.7155.60000 0001 2260 6941Department of Zoology, Faculty of Science, Alexandria University, Alexandria, Egypt; grid.418376.f0000 0004 1800 7673Animal Health Research Institute (AHRI), Agricultural Research Center (ARC), Port of Alexandria, Alexandria, Egypt
License: © The Author(s) 2021 CC BY 4.0 Open AccessThis article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
Article links: DOI: 10.1186/s43141-021-00224-0 | PubMed: 34417930 | PMC: PMC8380214
Relevance: Moderate: mentioned 3+ times in text
Full text: PDF (1016 KB)
Background
γ-Aminobutyric acid (GABA), the primary inhibitory neurotransmitter in the central nervous system (CNS), is a key coordinator of brain activity. GABA’s inhibitory effects are mediated by two types of receptors, GABAA and GABAB receptors [ref. 1]. GABAergic neurotransmission is critical in neurodevelopmental disorders [ref. 2]. GABAAR is one of the most significant drug targets in the treatment of neuropsychiatric disorders such as epilepsy, insomnia, and anxiety, as well as in anesthesia in surgical operations [ref. 3]. In addition, genetic studies have documented the relationship between GABAAR subunit genes and epilepsy [ref. 4], eating disorder [ref. 5], autism [ref. 6, ref. 7], and bipolar disorders [ref. 8]. GABAB receptors are members of the C family of G protein-coupled receptors (GPCRs), which are found in the nervous system and have been linked to some neurological and psychiatric disorders [ref. 9]. They are structurally and functionally distinct from GABAA receptors and will not be covered in this article. GABAC receptors are now considered to be part of GABAA receptor isoforms that are entirely made up of rho (ρ) subunits [ref. 10]. In this review, we will try to provide a quick rundown of what we know about GABAA receptors, including their structure, function, pharmacology, and related disorders.
GABAARs structure and gene organization
GABAA receptors are ligand-gated chloride channels that consist of pentameric combinations of different subunits. A total of 19 GABAA receptor subunit genes have been identified in humans that code for six α (alpha1-6), three β (beta1-3), three γ (gamma1-3), three ρ (rho1-3), and one each of the δ (delta), ε (epsilon), π (pi), and θ (theta) (Fig. 1A; Table 1) [ref. 11–ref. 13]. The diversity of GABAA receptors is due to the alternative splicing of several genes [ref. 14]. The GABAA receptor subunit genes are mainly arranged into four clusters on the human genome’s chromosomes 4, 5, 15, and X. Four genes, α2, α4, β1, and γ1 on chromosome 4; four genes α1, α6, β2, and γ2 on chromosome 5; three genes, α5, β3, and γ3 on chromosome 15; and three genes, α3, ϵ, and θ on chromosome X (Table 1) [ref. 15]. The receptor composition and arrangement influence its functional and pharmacological properties [ref. 16, ref. 17].
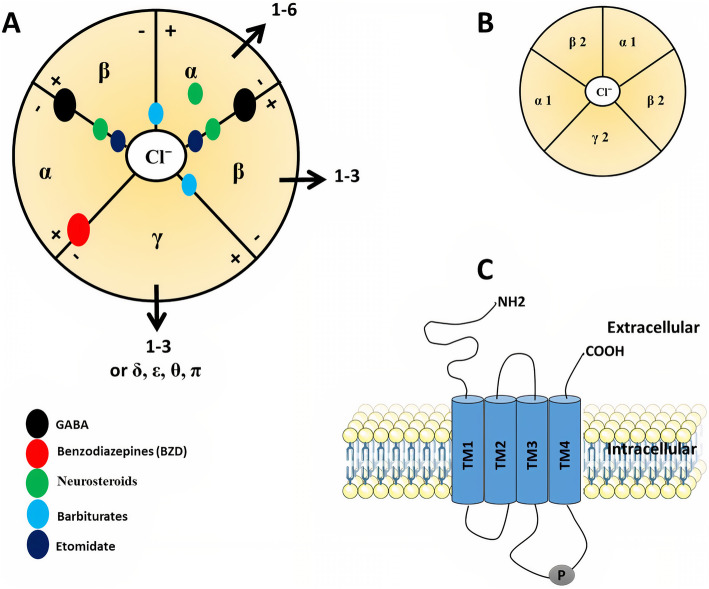
Table 1: GABAA receptor subunits
| Receptor subunit | Gene | Chromosome | Location | Reference |
|---|---|---|---|---|
| GABA-A alpha 1 (α1) | GABRA1 | 5 | 5q34 | Gene ID: 2554 |
| GABA-A alpha 2 (α2) | GABRA2 | 4 | 4p12 | Gene ID: 2555 |
| GABA-A alpha 3 (α3) | GABRA3 | X | Xq28 | Gene ID: 2556 |
| GABA-A alpha 4 (α4) | GABRA4 | 4 | 4p12 | Gene ID: 2557 |
| GABA-A alpha 5 (α5) | GABRA5 | 15 | 15q12 | Gene ID: 2558 |
| GABA-A alpha 6 (α6) | GABRA6 | 5 | 5q34 | Gene ID: 2559 |
| GABA-A beta 1 (β1) | GABRB1 | 4 | 4p12 | Gene ID: 2560 |
| GABA-A beta 2 (β2) | GABRB2 | 5 | 5q34 | Gene ID: 2561 |
| GABA-A beta 3 (β3) | GABRB3 | 15 | 15q12 | Gene ID: 2562 |
| GABA-A gamma 1 (γ1) | GABRG1 | 4 | 4p12 | Gene ID: 2565 |
| GABA-A gamma 2 (γ2) | GABRG2 | 5 | 5q34 | Gene ID: 2566 |
| GABA-A gamma 3 (γ3) | GABRG3 | 15 | 15q12 | Gene ID: 2567 |
| GABA-A delta (δ) | GABRD | 1 | 1p36.33 | Gene ID: 2563 |
| GABA-A epsilon (ε) | GABRE | X | Xq28 | Gene ID: 2564 |
| GABA-A pi (π) | GABRP | 5 | 5q35.1 | Gene ID: 2568 |
| GABA-A theta (θ) | GABRQ | X | Xq28 | Gene ID: 55879 |
| GABA-A rho 1 (ρ1) | GABRR1 | 6 | 6q15 | Gene ID: 2569 |
| GABA-A rho 2 (ρ2) | GABRR2 | 6 | 6q15 | Gene ID: 2570 |
| GABA-A rho 3 (ρ3) | GABRR3 | 3 | 3q11.2 | Gene ID: 200959 |
Data are compiled from NCBI-Gene
Each subunit has been thoroughly investigated in terms of amino acid sequence, level of expression, and localization in brain tissues, but it is still unclear the interaction between them to form many different isoforms [ref. 18]. This variety of isoforms may be present even in a single cell [ref. 10]. However, it is widely assumed that the main adult isoform is composed of α1, β2, and γ2 subunits which are arranged γ2β2α1β2α1 counterclockwise around a central pore as viewed from the cell exterior (Fig. 1B) [ref. 19].
GABAAR subunits share a common structure (Fig. 1A). The mature subunit is composed of ∼450 amino acid residues. It contains N-terminal, a large hydrophilic extracellular domain (ECD), four hydrophobic transmembrane domains (TMD: TM1–TM4) where TM2 is believed to form the pore of the chloride channel, and intracellular domain (ICD) between TM3 and TM4 which is the site of protein interactions and post-translational modifications that modulate receptor activity (Fig. 1C) [ref. 20, ref. 21]. The neurotransmitter GABA, as well as psychotropic drugs such as benzodiazepines (BZDs), bind to the N-terminal at binding sites α-β and α-γ interfaces, respectively. Neurosteroids and anesthetics like barbiturates, on the other hand, are found within the TMD of α and β subunits (Fig. 1A) [ref. 22–ref. 25].
GABAARs distribution
In the CNS, some GABAAR subunits possess broad expression while other subunits exhibit restricted expression. For example, the α6 subunit is expressed only in the cerebellum while the ρ subunit is expressed mainly, but not exclusively, in the retina [ref. 26]. GABAA receptors localized to postsynaptic sites in the brain are mainly composed of the α1–3, β1–3, and γ2 where GABA neurotransmitter can bind with and open chloride channels, thus increasing the anion conductance for a short period (milliseconds), leading to hyperpolarization of a depolarized membrane. This type of GABA inhibition has been termed phasic inhibition. On the other hand, GABAA receptors composed of the α4–6, β2/3 and δ subunits can localize to extrasynaptic sites where the low GABA concentration can open these receptors for a longer period which is called tonic inhibition [ref. 27]. The most popular isoforms of extrasynaptic GABAARs mediating tonic inhibition are α4βδ receptors in the forebrain, α6βδ receptors in the cerebellum and α1βδ receptors in the hippocampus [ref. 28]. It has been found that α2, α3, and β3 subunit-containing receptors are ~100 times more concentrated at synapses than in the extrasynaptic membrane [ref. 29]. Not all γ2-containing receptors are concentrated postsynaptic for example, α5βγ2 receptors are found at extrasynaptic sites involved in tonic inhibition [ref. 28]. Apart from phasic and tonic inhibition, the γ2 subunit is essential for postsynaptic clustering of GABAA receptors [ref. 30] and the γ3 subunit substitutes γ2 to contribute to the development of the postnatal brain [ref. 31]. On the other hand, outside the CNS, GABAA receptors have been found in different types of immune cells [ref. 32, ref. 33], liver cells [ref. 34], pancreatic islet β-cells [ref. 35], and airway smooth muscle [ref. 36]. Despite these observations, the laws that regulate GABAARs assembly, as well as the exact process by which GABAAR isoforms are distributed, remain unknown.
GABA neurotransmission
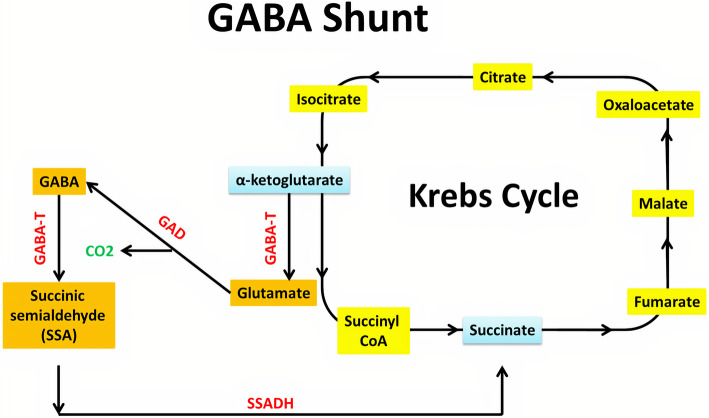
In 1950, Eugene Roberts and Sam Frankel discovered the major inhibitory neurotransmitter in the CNS of mammals, GABA [ref. 37]. Glucose is the main precursor for GABA synthesis, even though other amino acids and pyruvate act as precursors. The GABA shunt is a closed-loop system that produces and conserves GABA (Fig. 2). In GABA shunt, the first step is transamination of α-ketoglutarate produced from the metabolism of glucose in the Krebs cycle, by GABA-α ketoglutarate transaminase (GABA-T) to produce l-glutamic acid. Glutamic acid is decarboxylated to GABA by glutamic acid decarboxylase (GAD). GAD is an enzyme that uses vitamin B6 (pyridoxine) as a cofactor and is only expressed in cells that use GABA as a neurotransmitter. GABA-T metabolizes GABA to succinic semialdehyde. This transamination happens when α-ketoglutarate is present, it accepts the amino group extracted from GABA, and reforms glutamic acid. Succinic semialdehyde dehydrogenase (SSADH) oxidizes succinic semialdehyde to succinate. It can enter the Krebs cycle, thereby completing the loop [ref. 38]. A vesicular transporter helps to package newly synthesized GABA into synaptic vesicles. SNARE complexes help dock the vesicles into the plasma membrane of the cell [ref. 39]. Presynaptic neuron depolarisation releases GABA to the synaptic cleft and diffuses toward postsynaptic receptors. It can bind to post-synaptic GABA receptors (GABAA and GABAB), which modulate ion channels, hyperpolarize the cell, and prevent action potential transmission. Regardless of binding to GABAA or GABAB receptors, GABA serves as an inhibitor. In the case of GABAA ionotropic receptor, the presence of GABA increases chloride ion conductance into the cell. Consequently, the increased chloride ion influx results in membrane hyperpolarization, and neuronal excitability is reduced [ref. 40]. GABA can then be passed into three pathways. The first one is that GABA can be degraded extracellularly by GABA-T into succinate semialdehyde which then enters the citric acid cycle. The second is that the GABA can be reuptaken to nerve terminals for utilization again. The third one is that the GABA can be reuptaken to the glial cell where it undergoes metabolism to succinic semialdehyde by GABA-T or it becomes glutamine which is transported to neurones, where it is converted to glutamate by glutaminase and re-enters GABA shunt. In glia, GABA cannot be synthesized again from glutamate due to the absence of GAD [ref. 41, ref. 42].
The physiological role of GABA and GABAA receptors
Certainly, GABA/GABAARs signaling is the most prominent inhibitory pathway in the CNS. As we discussed before, there are two forms of GABA inhibition: phasic and tonic inhibition. The transient stimulation of GABAA receptors by GABA reduces postsynaptic neuron excitability, resulting in phasic inhibition [ref. 43, ref. 44]. Tonic inhibition, on the other hand, is thought to be a continuous mechanism of inhibition that regulates excitation through long-term hyperpolarization [ref. 45]. Tonic inhibition plays an important role in synaptic plasticity, neurogenesis [ref. 46, ref. 47] as well as cognitive functions [ref. 48, ref. 49]. Any disturbance in phasic or tonic inhibition is associated with many neurological and psychiatric diseases. Thus, modulating these signals has become the basis of drug therapy as well as anesthesia [ref. 50–ref. 55].
Furthermore, the GABAA receptor plays a pivotal role in neuronal cell proliferation and fate determination. A pioneering study showed that depolarizing GABA actions leads to a decrease in both DNA synthesis and the number of bromodeoxyuridine (BrdU)-labeled cells at the subventricular zone (SVZ) that mean GABA can affect the proliferation of progenitor cells in rat embryonic neocortex [ref. 56]. Furthermore, GABA or muscimol, a GABAA receptor agonist, also triggers membrane depolarization and induces proliferation of postnatal cerebellar granule progenitor cells in the developing rat cerebellum [ref. 57]. In the adult hippocampus, the neuronal progenitor cells at the subgranular zone (SGZ) show tonic GABAergic conductance. Impairment of this conductivity, as well as the increase in newly generated cells labeled by BrdU, was induced by genetic deletion of GABAARs containing α4, but not δ subunits [ref. 47, ref. 58, ref. 59]. In the postnatal subventricular zone (SVZ), GABA limits the proliferation of glial fibrillary acidic protein (GFAP)-expressing progenitors thought to be stem cells (also called Type 1 cells) [ref. 60]. Also, a recent study suggested that GABAA receptor contributes to determining the cell fate of neural stem cells [ref. 61]. These results indicate that adult neurogenesis may be influenced by multiple functions of GABAA receptors as well as ambient GABA released in an autocrine/paracrine manner [ref. 62, ref. 63].
Of note, GABAA receptors have additional physiological functions in tissues and organs outside the nervous system [ref. 64]. Such as in the pancreatic islet, β-cells synthesize huge amounts of GABA [ref. 35]. Via GABAA receptors, GABA suppresses glucagon secreted by α-cells [ref. 65], and increases insulin secreted by β-cells [ref. 66]. In addition, GABA stimulates β-cells proliferation and growth [ref. 66, ref. 67]. Therefore, targeting GABA/GABAA signaling is likely to be a part of diabetes treatment [ref. 68].
Molecular pharmacology of GABAA receptors
Apart from GABA, a variety of ligands have been discovered that bind to various locations on the GABAAR and regulate it. Binding sites are located at particular receptor subtypes, and these subtypes determine the receptors’ distinct pharmacological fingerprints [ref. 69]. The GABA-binding site, also known as the active site or orthosteric site, is where orthosteric agonists and antagonists bind. Orthosteric agonists, such as GABA, gaboxadol, isoguvacine, muscimol, and progabide [ref. 70–ref. 72], activate the receptor, resulting in increased Cl− conductance. By contrast, orthosteric antagonists, such as bicuculline and gabazine [ref. 73], compete with GABA for binding, inhibiting its effect and lowering Cl− conductance. Allosteric modulators, on the other hand, bind elsewhere on the receptor and exert their effect by causing conformational changes in the receptor either positively (PAM) such as barbiturates, benzodiazepines, z-drugs (nonbenzodiazepines) alcohol (ethanol), etomidate, glutethimide, anesthetics, and certain neurosteroids, or negatively (NAM) such as pregnenolone sulfate and zinc [ref. 54, ref. 74, ref. 75]. Non-competitive chloride channel blockers (ex., picrotoxin) are ligands that bind to or near the central pore of the GABAAR and block Cl− conductance [ref. 76]. Moreover, silent allosteric modulators (SAM) are a class of GABAAR modulators that can compete with a PAM or a NAM for the occupation of the binding site such as flumazenil [ref. 75, ref. 77]. The characteristics of ligands that contribute to receptor activation are usually used as anxiolytic, anticonvulsant, sedative, and muscle relaxant drugs. On the other side, ligands that inhibit receptor function usually have opposite pharmacological effects such as convulsion and anxiogenesis [ref. 78, ref. 79]. Interestingly, some subtypes of NAM (ex., α5IA) are being studied for their nootropic properties as well as potential therapies for GABAergic medication adverse effects [ref. 80].
GABA and GABA analogs
Cys-loop receptors typically have their neurotransmitter binding site at the extracellular interface between two neighboring subunits. The binding site’s principal face (+) is made up of three loops (A, B, and C), whereas the complementary face (−) comprises three β-strands and one loop (D, E, F, or G) [ref. 81, ref. 82]. In GABAARs, αβγ subtype (2α:2β:1γ) has two GABA binding sites at the β +/α − interfaces (Fig. 1A). When GABA occupies just one site, the channel opens; however, when both sites are occupied, the chances of channel opening rise dramatically [ref. 83]. Besides, chemicals with similar structures to GABA can attach to GABA binding sites and give different effects such as muscimol (agonist), gaboxadol (partial agonist), and bicuculline (competitive antagonist) [ref. 82].
Actually, it is still a mystery how amino acid residues interact with GABA. However, in a previous study based on αβγ subtype, GABA formed hydrogen bonds with α1T129 and β2T202, salt bridges with α1R66 and β2E155, and cation–pi interaction with β2Y205 [ref. 84]. On the other hand, β +/α− interface has aromatic residues formed by βY97, βY157, βF200, βY205, and αF64 which are conserved at the β +/β −, β +/γ −, and β +/δ − interfaces. Furthermore, the GABA-binding subunit residues R131, T129, and L127 are maintained at the equivalent places in the β, γ, and δ subunits [ref. 81, ref. 84, ref. 85]. Future studies will examine whether GABA and other structurally similar chemicals are attracted to these non-canonical sites, as well as how these sites may influence receptor activation.
Benzodiazepines
Benzodiazepines (BZDs) are commonly used in different treatments related to anxiety, sleep disorders, seizure disorders, muscle spasms, and some forms of depression [ref. 86]. BZD allosterically modulate GABAAR and give its therapeutic effect through binding to the α+/γ − interface (Fig. 1A) and increasing Cl− conductance [ref. 24, ref. 87]. Interestingly, amino acids involved in the binding sites of BZDs are homologous to that of the GABA binding site at the β +/α − interface [ref. 88]. Besides, mutations that converted histidine to arginine (α1H101R, α2H101R, α3H126R, and α5H105R) at the β2γ2 subtype of GABAARs eliminated diazepam activity, while reverse mutations (from R to H) elicited the diazepam response [ref. 89]. BZD-sensitive GABAARs subtypes are formed of two α subunits with two β subunits and a γ subunit (Fig. 1A) [ref. 90]. Likewise, GABAAR containing α4, α6, and γ2 subunits, potently bind many BZD ligands [ref. 91, ref. 92]. But subtypes containing δ are relative with low abundance, and the subunits replacing γ and δ, such as ε, are even rarer [ref. 93]. Of note, the GABAAR subtypes containing δ subunits are located extrasynapically inducing tonic inhibitory currents in major cell populations including cerebellar and hippocampal granule cells [ref. 43, ref. 93]. It was thought that these subtypes are not capable to bind any BZD ligands, lacking the high-affinity α+/γ− (site 1), but later it was found to bind some BZD ligands with lower affinity at distinct other sites on the GABAAR [ref. 54].
Benzodiazepines as zolpidem (an imidazopyridine) and other clinically used hypnotics like zaleplon (a pyrazolopyrimidine) and zopiclone (a cyclopyrrolone), as well as quinolones, triazolopyridazines, and beta-carbolines show a higher affinity for α1-containing receptors than for α2- or α3-containing subtypes, while they do not affect α5-containing GABAARs [ref. 93, ref. 94]. Also, imidazobenzodiazepine oxazole derivatives have shown some α2/α3 selectivity [ref. 95]. Pyrazoloquinolinones, which are examples for BZD site-active PAM in γ–containing subtypes, demonstrate a wide range of effects as well as selectivity for α and β subunits [ref. 54]. Also, BZD-site ligands have more or less efficacy than traditional BZD agonists on the traditional BZD-sensitive subtypes, and unexpected efficacy on the diazepam-insensitive subtypes like GABAAR containing α4 or α6, or α and β without γ [ref. 96, ref. 97].
Alpha5IA is selective inverse agonists that bind to the BZD site at the α5 subtype that is highly expressed in the CA1 region of the hippocampus. It has been suggested to improve cognitive functions [ref. 98]. Such α5 inverse agonists also reduce side effects of BZDs, general anesthetics [ref. 99], and alcohol [ref. 100]. They may be useful for treating Down syndrome, autism spectrum disorder, schizophrenia, and affective disorders [ref. 101].
Anesthetics
GABAARs are remarkable targets of variable volatile anesthetics, intravenous anesthetics, etomidate, and propofol, as well as steroid anesthetics, barbiturates, and ethanol [ref. 102]. Anesthetic binding sites on the GABAAR can be identified using site-directed mutagenesis [ref. 103], substituted cysteine modification protection (SCAMP) [ref. 104], or photo-affinity labeling [ref. 102, ref. 105]. At higher concentrations, some anesthetics, especially the intravenous anesthetics, etomidate, propofol, and barbiturates, could directly activate GABAARs in the absence of GABA. Such direct activation distinguished them as GABA-mimetic from benzodiazepines which lack this property. Studies that were based on site-directed mutagenesis produced several residues of interest, particularly in the trans-membrane regions of the α and β subunits, for both volatile and intravenous anesthetics [ref. 106].
Of note, methionine residues, especially αM236 and βM286 located in the M1 and M3 domains respectively, have been shown to be significant determinants of etomidate binding and function in experiments that used mutagenesis and photoreactive etomidate analogs. Based on crystal structures of GABAARs, αM236 and βM286 are expected to be found at the β +/α − interfaces in the TMD, below the GABA binding sites (Fig. 1A). Also, αT237 (M1), αI239 (M1), αL232 (M1), βV290 (M3), and βF289 (M3) are among the additional residues linked to etomidate binding and function [ref. 107, ref. 108]. Besides, in α1β3γ2 GABAARs, other anesthetic binding sites including α +/β − and γ +/β – interfaces (Fig. 1A) have been identified using photoreactive analogs of barbiturate where αA291 (M3), αY294 (M3), βM227 (M1), and γS301 (M3) were among the binding residues [ref. 82, ref. 109]. Moreover, in the TMD of β3 homomeric GABAARs at β +/β – interface, photoreactive propofol can bind to β (+) M286, β (+) F289, and β (–) M227 residues inducing functional activity of the receptor [ref. 110–ref. 112].
It has been found that β2 and β3 subunits were significant for modulation of GABAAR by i.v. anesthetics. In addition, transgenic mice that were generated through β2 (N265S) and β3 (N265M) mutations in the GABAAR became insensitive to the actions of propofol and etomidate [ref. 113, ref. 114]. The affinity and efficacy of barbiturate depend on the composition of the subunit, but the α subunit seems to be more important than β [ref. 115]. Recently, it has been suggested that the binding of barbiturate, etomidate, and propofol is predominantly at the αβ+/α−γ interface as well as the α+/β− or α+/γ− TMD interfaces in α1β2γ2 [ref. 69, ref. 116]. Other photo-affinity labeling depending studies suggested that binding sites for barbiturates and etomidate at α4β3δ GABAAR subtypes at the β+/α–, and β+/β– TMD interfaces, respectively, were not suitable for binding of delta selective compound 2 (DS2) or alphaxalone [ref. 117].
Neurosteroid
Endogenous steroids exhibit GABAAR-mediated neuroactive effects including anesthesia, anticonvulsant, analgesia, and sedation. The most common examples are allopregnanolone and its synthetic analogs [ref. 118]. Although the exact position of the neurosteroid binding sites has yet to be determined, many residues in the TMDs have been shown to impact neurosteroid activity, such as αS240 (M1), αQ241 (M1), αN407 (M4), αY410 (M4), αT236 (M1), and βY284 (M3) [ref. 119–ref. 121]. The modulatory and activation sites are located at the TMDs of α subunit and β +/α – interfaces respectively (Fig. 1A) [ref. 82, ref. 122].
Flavonoids
Flavonoids are present in most plants and a few microorganisms. They have been discovered as modulators of the BZD-site of GABAARs, but the variability of compounds within this group participated in showing their potential action at more than one additional binding site on GABAARs. Flavonoids can act as either negative, positive, or neutralizing on GABAARs or directly as allosteric agonists [ref. 123]. Flavonoids share the elementary structure of a phenylbenzopyran, most commonly of a flavan (2-phenylchromane). Subgroups contain isoflavones, flavonoles, flavones, flavanonole, flavanones, and flavanoles. Among these groups, isoflavones and flavones particularly have been found to interact with the binding site of BZD [ref. 124]. Structure-activity experiments have illustrated that flavones have higher potency on BZD radioligand binding than their flavanone or flavonol counterparts. Besides, glycosylation had a negative influence on binding [ref. 125]. Flavonoids can also interact with flumazenil-sensitive or -insensitive GABAARs [ref. 123]. Some of the flavonoids have shown subtype-selectivity like flavan-3-ol ester Fa131 [ref. 126] or 6,2′- dihydroxyflavone [ref. 127]. The flavone hispidulin showed potent activity in crossing the blood-brain barrier associated with the α6β2γ2 subtype of GABAARs, which is used to reduce the susceptibility of seizures [ref. 128].
Cannabinoids
Cannabinoids are chemical substances present in the cannabis plant. The phytocannabinoid tetrahydrocannabinol (THC) is the primary psychoactive compound in cannabis. Besides, cannabidiol (CBD) is another significant component of the plant [ref. 129]. It has been found that CBD has sedative, anxiolytic, and anticonvulsant effects and has been suggested for treating pediatric epilepsies such as Dravet syndrome [ref. 130]. CBD, also, showed a low affinity for the main cannabinoid receptor and exhibits an activity profile similar to that of GABA PAMs inducing anxiolytic and anticonvulsant effects [ref. 131].
Endocannabinoids, such as 2-Arachidonoylglycerol (2-AG), 2-Arachidonyl glyceryl ether, N-Arachidonoyl dopamine (NADA), Arachidonoylethanolamine (AEA), and Lysophosphatidylinositol (LPI) [ref. 132], are substances produced in the body activating cannabinoid receptors (CB1, CB2) [ref. 133, ref. 134]. Additionally, they have been identified as positive modulators for GABAAR subtypes [ref. 135]. Studies on recombinant receptors showed that 2-AG increases GABAAR activity at low non-saturating GABA concentrations while decreasing the activity at high saturating GABA concentrations. Therefore, the impact of endocannabinoids on GABAAR depends on the regulation of GABA inhibition [ref. 136].
Picrotoxin
Picrotoxin is a plant-derived product, with a universal efficacy as GABAAR’s chloride channel blocker. Picrotoxin is found naturally in the Anamirta Cocculus plant, although it can be synthesized chemically [ref. 137, ref. 138]. It has been utilized as a CNS stimulant, and antidote for poisoning by CNS depressants and barbiturates [ref. 139]. However, due to the toxicity of picrotoxin, it is currently used only in research. Furthermore, numerous studies indicated that a wide range of molecules from various chemical families had an affinity for picrotoxin-binding sites such as t-butylbicyclophosphorothionate (TBPS), t-butylbicycloorthobenzoate (TBOB), pentylenetetrazole, and some insecticides (ex., dieldrin and lindane) [ref. 140–ref. 142]. A study by Othman et al. (2012) [ref. 143] found that low concentrations of GABA increase picrotoxin and TBPS binding affinity to GABAAR containing α1β2γ2, while application of GABA at high concentration reduces their binding affinity to the receptor reducing channel blocking activity. This indicates that picrotoxin and ligands of picrotoxin-binding sites are highly dependent on the regulation of GABA inhibition.
Pharmacology of δ-containing GABAARs
The unique role of the δ subunit in extra-synaptic GABAARs, a group of receptors responsible for tonic GABAergic inhibition has generated immense therapeutic and research interests. However, the complicated properties of the δ subunit assembly and the rarity of δ-selective ligands are the main reasons hindering progress in pharmacological studies of these receptors. Variable compounds have been claimed to be selective for the δ subunit. The hypnotic drug THIP (4,5,6,7-tetrahydroisoxazolo[5,4-c]pyridin-3-ol) and gaboxadol are examples of compounds that are known by their direct activation of αβδ with higher efficacy and potency than αβγ but does not discriminate between αβ and αβδ receptors [ref. 144, ref. 145]. Similar to THIP, anesthetics, as well as neurosteroids, also show more pronounced action at δ-containing GABAARs, but their activity is independent of subunit composition, these compounds are not considered to be δ-selective. In contrast, 4-chloro- N-(2-thiophen-2-ylimidazo[1,2-a] pyridin-3-yl) benzamide which was found to be a positive modulator at α4/6βδ, has limited efficacy at αβγ and is inactive at αβ GABAARs [ref. 146].
GABAA receptor dysfunction and neuro-psychiatric disorders
Epilepsy
Epilepsy is a neurological disease characterized by frequent and unexpected seizures caused by abnormal brain electricity, which results in loss of consciousness and unusual behaviors [ref. 147]. Around 65 million people are affected worldwide, of all ages and genders [ref. 148]. An imbalance between excitation and inhibition induced by impaired GABAergic signaling can trigger various forms of epilepsy [ref. 149, ref. 150]. Several studies have demonstrated the importance of GABAA receptors as targets for antiepileptic drugs [ref. 45, ref. 151, ref. 152]. Mutations in GABAA receptor subunit genes have been linked to several types of idiopathic epilepsy in which the pathophysiological consequences of the mutations are impairments in the gating characteristics of the channel or receptor trafficking [ref. 4]. The severity of the disorder appears to depend on the type of mutation (nonsense, missense, or frameshift), its location in the gene (promoter or protein-coding region), the affected region of the encoded protein (intra-/extracellular or transmembrane) and the affected subunit gene [ref. 4]. Some mutations in genes encoding the α1, α6, β2, β3, γ2, or δ subunits of GABAARs have been detected in both animal models of epilepsy and patients with epilepsy [ref. 153, ref. 154]. Likewise, Dravet syndrome, also known as severe myoclonic epilepsy in infancy (SMEI), is a form of epilepsy that affects children at the age of approximately 1 year as a result of mutations in genes encoding the α1, β1, β2, and γ2 subunits of GABAARs [ref. 4, ref. 155]. Of note, several GABAAR mutations associated with epilepsy lead to abnormal trafficking of the receptors and thus partially or completely impair their expression on the synaptic plasma membrane [ref. 155, ref. 156]. Likewise, a study by Dejanovic et al. [ref. 157] discovered a missense mutation in GPHN gene, the gene encoding the gephyrin protein, in a patient with Dravet syndrome. Gephyrin is the main protein that clusters and stabilizes GABAARs at the inhibitory postsynaptic membranes of the central nervous system [ref. 158]. Moreover, during the epileptogenic period, expression of the gephyrin protein decreases gradually in the neocortex before returning to baseline during the chronic phase [ref. 159]. These findings suggest that the downregulation of GABAAR subunits or their interactors that play a functional role in receptor activity, such as gephyrin, maybe the origin of the disease and thus could be used as drug targets.
Alzheimer’s disease
Alzheimer’s disease (AD) is one of the primary diseases that cause neurodegeneration. Clinically, AD is marked by significant cognitive deficits and regarded as the most common cause of dementia. The aggregation of misfolded amyloid-beta (Aβ) protein, which forms amyloid plaques in the gray matter of the brain, is the origin of AD pathophysiology. Amyloid plaques, neuronal dysfunction, and tangles of neural fibers are major pathological features of the disease [ref. 160, ref. 161]. Several experiments, in both AD patients and mice, have shown that accumulation of misfolded Aβ interferes with GABAergic interneuron activity, causing impaired synaptic communication and loss of neural network activity, which eventually leads to cognitive dysfunction [ref. 162–ref. 165]. A recent study showed transcriptional downregulation of α1, α2, α3, α5, β1, β2, β3, δ, γ2, γ3, and θ subunits of GABAA receptors, and GAD enzyme in the middle temporal gyrus (MTG) of post-mortem brain samples from AD patients. These alterations impair the balance between excitatory and inhibitory pathways that may lead to cognitive dysfunction in AD [ref. 166]. Likewise, in biochemical studies, GABA neurotransmitter levels were substantially lower in the CSF as well as the temporal cortex of Alzheimer’s patients, implying impaired synaptic activity and neuronal transmission [ref. 44, ref. 167–ref. 169]. Also, a study by Limon et al. [ref. 170] showed that most aspects of the GABA system were impaired in the brains of AD patients, such as GABAergic neural circuit, GABA levels, and expression levels of GABAA receptors. Furthermore, in AD mice, activating GABAA receptors with baicalein (positive allosteric modulator of the benzodiazepine site of the GABAAR) for 8 weeks significantly reduced Aβ production, improved cognitive function, and decreased pathological features [ref. 171]. As a result, GABAA receptors seem to be a potential therapeutic target in the treatment of AD.
Cervical dystonia
Cervical dystonia (CD) is the most frequent type of adult-onset focal dystonia. It is a neurological disorder marked by involuntary and prolonged muscle contractions that cause irregular postures and neck tremors [ref. 172–ref. 174]. Studying the pathophysiology of isolated cervical dystonia using different methods such as magnetic resonance spectroscopy (MRS), positron emission tomography (PET), and functional magnetic resonance imaging (f-MRI) demonstrated an alteration in the GABA-mediated inhibitory signaling pathway in the cortical, cerebellar, and basal ganglia regions of the brain [ref. 175]. Similarly, a significant number of functional defects have been identified in the thalamus of patients with CD [ref. 176], and blocking GABAA receptors in the thalamus triggered CD-like symptoms in monkeys [ref. 177]. According to a recent study, GABA levels in the right thalamus were decreased in a sample of adult-onset CD patients, and the availability of GABAA receptors was negatively correlated with disease duration and the severity of dystonia [ref. 178].
Brain injury
Several studies investigated whether GABA signaling pathways are involved in several forms of brain injuries using different stroke mice models. As reported in earlier studies, increasing GABA inhibition has shown a neuroprotective role at stroke onset. In contrast, increased GABAergic tonic inhibition at extrasynaptic GABAA receptors would adversely affect and exacerbate stroke pathology. Also, these findings were in line with study results obtained from knockout mice models lacking either α5-GABAA or δ-GABAA receptors, which have revealed better recovery from stroke than healthy mice models because of GABAergic signaling remission [ref. 179, ref. 180].
Autism spectrum disorder
Autism spectrum disorder (ASD) has three characteristic behavioral features: impaired communication and social deficits, and repetitive behaviors. Several studies concluded an imbalance in the glutamatergic/GABAergic signaling pathways and neuroinflammation process were associated with ASD pathophysiology and were also detected in several ASD mice models [ref. 181]. Earlier studies reported the presence of molecular-level cortical abnormalities related to GABAergic signaling dysfunction in the brains of ASD. The excitatory and inhibitory signaling imbalance caused by variations in GABA levels represents one of the characteristic features behind behavioral deficits in autism [ref. 182]. Mendez et al. [ref. 183] conducted a PET imaging study using a radioactive ligand [11C]-Ro15–4513 VT for tracing levels of GABAA receptor α5 subunits in ASD. The results showed a reduction in GABAA receptors in the brain’s two limbic areas (amygdala and nucleus accumbens) of autism patients. Contrary to previous findings, a recent study demonstrated that the impairment in the GABAergic system in ASD mouse models and autistic patients was not associated with alterations in GABA receptor numbers between healthy and ASD controls, as concluded by an earlier study [ref. 184]. Also, a recent meta-analysis was conducted to verify earlier findings supporting the association between different genetic variants of GABAA receptor subunits and the risk of developing autism in children. In conclusion, the study showed no association between GABA receptor subunits (β3, α5, and α3) and child autism [ref. 185].
Schizophrenia
Schizophrenia is a multifactorial major psychiatric disorder whose etiology has been associated with hundreds of protein-coding genes reported by different genome-wide association studies. Changes in post-translational modifications of various proteins including GABAA receptors and their contribution to schizophrenia pathophysiology were reported [ref. 186]. A previous study showed glycosylation changes in multiple protein receptor subunits in the brains of schizophrenic patients, such as AMPA and GABAA receptor subunits [ref. 187].
Specifically, several post-mortem brain studies conducted using lectin affinity analysis and enzyme de-glycosylation of GABAA receptors of superior temporal gyrus of schizophrenic brains demonstrated a decrease in high-mannose N-glycans residues of GABA-associated proteins in individuals with schizophrenia that were specific to different GABAA receptor subunits on the ɑ1, ɑ4, β1, β2, and β3 subunits; increased high- mannose N-glycans on β1 subunit; decreased high-mannose N-glycans on ɑ1 subunit; altered total N-glycans on β2 subunits. These N-glycosylation alterations were further associated with abnormal trafficking and localization of β1/ β2 subunits leading to an aberrant inhibitory signaling system observed in schizophrenia [ref. 188, ref. 189].
Furthermore, Marques and his co-workers [ref. 190] investigated the availability of α5-GABAA receptors in the hippocampus using PET imaging for hippocampal regions schizophrenic and healthy controls. The study results demonstrated a reduction of [11C]-Ro15–4513 VT ([11C]-Ro15–4513), which is a radioactive tracer used by PET scans to assess the total volume of distribution for α5-GABAA receptors in the hippocampus of untreated schizophrenic patients versus healthy controls. In contrast, there were no differences between healthy control and the second cohort of patients treated with antipsychotics. These findings were also positively correlated with scaling using PANSS (Positive and Negative Syndrome Scale) scores (i.e., is a medical scale system that measures the severity of schizophrenic symptoms).
Depression
Major depression is one of the debilitating diseases that leads to neurons’ anatomical and functional changes in the brain’s prefrontal cortex and is induced by chronic stress. Earlier studies had concluded that dysfunction in monoaminergic signaling was the main contribution to depression pathophysiology. Lately, accumulating evidence has suggested the potential role of GABAergic signaling dysfunction in predispositions of depression as it has been reported that both depression and chronic stress are associated with an imbalance in inhibition, and excitation of neuronal signaling resulted from a deficiency in neuronal transmission onto the brain’s prefrontal cortex (PFC). This imbalance resulted from the deficient transmission of GABAergic inhibitory signals onto the brain’s excitatory glutamate interneurons. In this context, several studies were conducted to demonstrate the correlation between GABAergic dysfunction and depression. For instance, a study showed using magnetic resonance imaging established decreased GABA and GAD67 levels and alterations in distinct types of GABA receptor subunits in the brains of depressed patients and stressed mice models. Studies conducted on genetically modified depressed mice models lacking specific GABA receptors showed depressive mice behaviors [ref. 191].
Data from magnetic resonance imaging MRI studies reported a reduction in hippocampal volume of the brain of depressed patients, which leads to alterations in neural circuits of different areas of the brain related to emotionality, such as amygdala and prefrontal cortex. Interestingly, study results using depressed mice models lacking GABAA receptors showed that any alterations in the brain’s GABAergic system were presented by cognitive, neuroanatomical, and behavioral deficits like significant depression disorder symptoms presented by depressed animal models. Accordingly, it is now presumed that the GABAergic system plays a vital role in controlling neuronal transmission in neuronal maturation in the hippocampus. Therefore, it is considered a therapeutic target for potential antidepressant drugs [ref. 28, ref. 192].
Attention and social behavior
Several studies have shown that inhibiting cortical GABAA receptors causes impaired attention [ref. 16, ref. 193–ref. 197], social behavior [ref. 198], and decision-making [ref. 199]. Recently, it has been demonstrated that mice models having impaired 5-alpha GABAA receptors were presented with behavioral deficits like symptoms associated with attention and social disorders [ref. 194].
Conclusion
Deep insights into the different GABAA receptor isoforms’ composition, arrangement, subunit interactors, and molecular pharmacology will give us a clear vision to understand alterations that may lead to CNS disorders. In our view, these discussions are of vital importance in drug discovery and development in the future.
References
- TA Simeone, SD Donevan, JM Rho. Molecular biology and ontogeny of gamma-aminobutyric acid (GABA) receptors in the mammalian central nervous system. J Child Neurol, 2003. [DOI | PubMed]
- K Ramamoorthi, Y Lin. The contribution of GABAergic dysfunction to neurodevelopmental disorders. Trends Mol Med, 2011. [DOI | PubMed]
- ER Korpi, ST Sinkkonen. GABA(A) receptor subtypes as targets for neuropsychiatric drug development. Pharmacol Ther, 2006. [DOI | PubMed]
- RL Macdonald, J-Q Kang, MJ Gallagher. Mutations in GABAA receptor subunits associated with genetic epilepsies. J Physiol, 2010. [DOI | PubMed]
- CS Bloss, W Berrettini, AW Bergen. Genetic association of recovery from eating disorders: the role of GABA receptor SNPs. Neuropsychopharmacol Off Publ Am Coll Neuropsychopharmacol, 2011. [DOI]
- AL Collins, D Ma, PL Whitehead. Investigation of autism and GABA receptor subunit genes in multiple ethnic groups. Neurogenetics, 2006. [DOI | PubMed]
- DQ Ma, PL Whitehead, MM Menold. Identification of significant association and gene-gene interaction of GABA receptor subunit genes in autism. Am J Hum Genet, 2005. [DOI | PubMed]
- SA Ament, S Szelinger, G Glusman. Rare variants in neuronal excitability genes influence risk for bipolar disorder. Proc Natl Acad Sci U S A, 2015. [DOI | PubMed]
- B Bettler, K Kaupmann, J Mosbacher, M Gassmann. Molecular structure and physiological functions of GABA(B) receptors. Physiol Rev, 2004. [DOI | PubMed]
- RW Olsen, W Sieghart. International Union of Pharmacology. LXX. Subtypes of γ-aminobutyric acidA receptors: classification on the basis of subunit composition, pharmacology, and function. Update. Pharmacol Rev, 2008. [DOI | PubMed]
- JL Steiger, SJ Russek. GABAA receptors: building the bridge between subunit mRNAs, their promoters, and cognate transcription factors. Pharmacol Ther, 2004. [DOI | PubMed]
- W Sieghart, K Fuchs, V Tretter. Structure and subunit composition of GABA(A) receptors. Neurochem Int, 1999. [DOI | PubMed]
- KH Backus, M Arigoni, U Drescher. Stoichiometry of a recombinant GABAA receptor deduced from mutation-induced rectification. Neuroreport, 1993. [DOI | PubMed]
- C Daniel, M Öhman. RNA editing and its impact on GABAA receptor function. Biochem Soc Trans, 2009. [DOI | PubMed]
- MG Darlison, I Pahal, C Thode. Consequences of the evolution of the GABA(A) receptor gene family. Cell Mol Neurobiol, 2005. [DOI | PubMed]
- E Sigel, R Baur, G Trube. The effect of subunit composition of rat brain GABAA receptors on channel function. Neuron, 1990. [DOI | PubMed]
- F Minier, E Sigel. Positioning of the alpha-subunit isoforms confers a functional signature to gamma-aminobutyric acid type A receptors. Proc Natl Acad Sci U S A, 2004. [DOI | PubMed]
- E Sigel, ME Steinmann. Structure, function, and modulation of GABAA receptors. J Biol Chem, 2012. [DOI | PubMed]
- R Baur, F Minier, E Sigel. A GABAA receptor of defined subunit composition and positioning: concatenation of five subunits. FEBS Lett, 2006. [DOI | PubMed]
- ZW Chen, RW Olsen. GABAA receptor associated proteins: a key factor regulating GABAA receptor function. J Neurochem, 2007. [DOI | PubMed]
- TC Jacob, SJ Moss, R Jurd. GABAA receptor trafficking and its role in the dynamic modulation of neuronal inhibition. Nat Rev Neurosci, 2008. [DOI | PubMed]
- S-H Chuang, DS Reddy. Genetic and molecular regulation of extrasynaptic GABA-A Receptors in the Brain: Therapeutic Insights for Epilepsy. J Pharmacol Exp Ther, 2018. [DOI | PubMed]
- U Rudolph, B Antkowiak. Molecular and neuronal substrates for general anaesthetics. Nat Rev Neurosci, 2004. [DOI | PubMed]
- E Sigel, BP Lüscher. A closer look at the high affinity benzodiazepine binding site on GABAA receptors. Curr Top Med Chem, 2011. [DOI | PubMed]
- M Wang. Neurosteroids and GABA-A receptor function. Front Endocrinol, 2011. [DOI]
- W Sieghart, G Sperk. Subunit composition, distribution and function of GABA(A) receptor subtypes. Curr Top Med Chem, 2002. [DOI | PubMed]
- JL Maguire, BM Stell, M Rafizadeh, I Mody. Ovarian cycle-linked changes in GABA(A) receptors mediating tonic inhibition alter seizure susceptibility and anxiety. Nat Neurosci, 2005. [DOI | PubMed]
- B Luscher, T Fuchs, CL Kilpatrick. GABAA receptor trafficking-mediated plasticity of inhibitory synapses. Neuron, 2011. [DOI | PubMed]
- Y Kasugai, JD Swinny, JDB Roberts. Quantitative localisation of synaptic and extrasynaptic GABAA receptor subunits on hippocampal pyramidal cells by freeze-fracture replica immunolabelling. Eur J Neurosci, 2010. [DOI | PubMed]
- C Essrich, M Lorez, JA Benson. Postsynaptic clustering of major GABAA receptor subtypes requires the gamma 2 subunit and gephyrin. Nat Neurosci, 1998. [DOI | PubMed]
- K Baer, C Essrich, JA Benson. Postsynaptic clustering of gamma-aminobutyric acid type A receptors by the gamma3 subunit in vivo. Proc Natl Acad Sci U S A, 1999. [DOI | PubMed]
- H Bjurstöm, J Wang, I Ericsson. GABA, a natural immunomodulator of T lymphocytes. J Neuroimmunol, 2008. [DOI | PubMed]
- S Alam, DL Laughton, A Walding, AJ Wolstenholme. Human peripheral blood mononuclear cells express GABAA receptor subunits. Mol Immunol, 2006. [DOI | PubMed]
- GY Minuk, M Zhang, Y Gong. Decreased hepatocyte membrane potential differences and GABAA-beta3 expression in human hepatocellular carcinoma. Hepatology, 2007. [DOI | PubMed]
- E Adeghate, AS Ponery. GABA in the endocrine pancreas: cellular localization and function in normal and diabetic rats. Tissue Cell, 2002. [DOI | PubMed]
- K Mizuta, D Xu, Y Pan. GABAA receptors are expressed and facilitate relaxation in airway smooth muscle. Am J Phys Lung Cell Mol Phys, 2008. [DOI]
- E Roberts, S Frankel. gamma-Aminobutyric acid in brain: its formation from glutamic acid. J Biol Chem, 1950. [DOI | PubMed]
- AW Bown, BJ Shelp. The metabolism and functions of [gamma]-Aminobutyric acid. Plant Physiol, 1997. [DOI | PubMed]
- TC Südhof. Neurotransmitter release: the last millisecond in the life of a synaptic vesicle. Neuron, 2013. [DOI | PubMed]
- R Miles. A homeostatic switch. Nature, 1999. [DOI | PubMed]
- AL Pehrson, C Sanchez. Altered γ-aminobutyric acid neurotransmission in major depressive disorder: a critical review of the supporting evidence and the influence of serotonergic antidepressants. Drug Des Devel Ther, 2015. [DOI]
- NM Rowley, KK Madsen, A Schousboe, H Steve White. Glutamate and GABA synthesis, release, transport and metabolism as targets for seizure control. Neurochem Int, 2012. [DOI | PubMed]
- M Farrant, Z Nusser. Variations on an inhibitory theme: phasic and tonic activation of GABA(A) receptors. Nat Rev Neurosci, 2005. [DOI | PubMed]
- Y Li, H Sun, Z Chen. Implications of GABAergic neurotransmission in Alzheimer’s disease. Front Aging Neurosci, 2016. [DOI | PubMed]
- S Schipper, MW Aalbers, K Rijkers. Tonic GABAA receptors as potential target for the treatment of temporal lobe epilepsy. Mol Neurobiol, 2016. [DOI | PubMed]
- S Ge, ELK Goh, KA Sailor. GABA regulates synaptic integration of newly generated neurons in the adult brain. Nature, 2006. [DOI | PubMed]
- V Duveau, S Laustela, L Barth. Spatiotemporal specificity of GABAA receptor-mediated regulation of adult hippocampal neurogenesis. Eur J Neurosci, 2011. [DOI | PubMed]
- V Lee, G MacKenzie, A Hooper, J Maguire. Reduced tonic inhibition in the dentate gyrus contributes to chronic stress-induced impairments in learning and memory. Hippocampus, 2016. [DOI | PubMed]
- LJ Martin, AA Zurek, JF MacDonald. Alpha5GABAA receptor activity sets the threshold for long-term potentiation and constrains hippocampus-dependent memory. J Neurosci, 2010. [DOI | PubMed]
- KA Yamada, WP Norman, P Hamosh, RA Gillis. Medullary ventral surface GABA receptors affect respiratory and cardiovascular function. Brain Res, 1982. [DOI | PubMed]
- JB Hammond, F Ahmad. Hepatic encephalopathy and role of antibenzodiazepines. Am J Ther, 1998. [DOI | PubMed]
- P Termsarasab, T Thammongkolchai, SJ Frucht. Medical treatment of dystonia. J Clin Mov Disord, 2016. [DOI | PubMed]
- D Kondziella. The top 5 neurotransmitters from a clinical neurologist’s perspective. Neurochem Res, 2017. [DOI | PubMed]
- RW Olsen. GABA(A) receptor: positive and negative allosteric modulators. Neuropharmacology, 2018. [DOI | PubMed]
- VT Pedrón, AP Varani, B Bettler, GN Balerio. GABA(B) receptors modulate morphine antinociception: pharmacological and genetic approaches. Pharmacol Biochem Behav, 2019. [DOI | PubMed]
- JJ LoTurco, DF Owens, MJ Heath. GABA and glutamate depolarize cortical progenitor cells and inhibit DNA synthesis. Neuron, 1995. [DOI | PubMed]
- ML Fiszman, LN Borodinsky, JH Neale. GABA induces proliferation of immature cerebellar granule cells grown in vitro. Brain Res Dev Brain Res, 1999. [DOI | PubMed]
- G Deidda, IF Bozarth, L Cancedda. Modulation of GABAergic transmission in development and neurodevelopmental disorders: investigating physiology and pathology to gain therapeutic perspectives. Front Cell Neurosci, 2014. [DOI | PubMed]
- J Song, C Zhong, MA Bonaguidi. Neuronal circuitry mechanism regulating adult quiescent neural stem-cell fate decision. Nature, 2012. [DOI | PubMed]
- X Liu, Q Wang, TF Haydar, A Bordey. Nonsynaptic GABA signaling in postnatal subventricular zone controls proliferation of GFAP-expressing progenitors. Nat Neurosci, 2005. [DOI | PubMed]
- 61.Song J, Olsen RHJ, Sun J et al (2016) Neuronal circuitry mechanisms regulating adult mammalian neurogenesis. Cold Spring Harb Perspect Biol 8. 10.1101/cshperspect.a018937
- SZ Young, J-C Platel, JV Nielsen. GABA(A) Increases calcium in subventricular zone astrocyte-like cells through L- and T-type voltage-gated calcium channels. Front Cell Neurosci, 2010. [DOI | PubMed]
- SZ Young, CA Lafourcade, J-C Platel. GABAergic striatal neurons project dendrites and axons into the postnatal subventricular zone leading to calcium activity. Front Cell Neurosci, 2014. [DOI | PubMed]
- M Watanabe, K Maemura, K Kanbara. GABA and GABA receptors in the central nervous system and other organs. Int Rev Cytol, 2002. [DOI | PubMed]
- E Xu, M Kumar, Y Zhang. Intra-islet insulin suppresses glucagon release via GABA-GABAA receptor system. Cell Metab, 2006. [DOI | PubMed]
- I Purwana, J Zheng, X Li. GABA promotes human β-cell proliferation and modulates glucose homeostasis. Diabetes, 2014. [DOI | PubMed]
- J Tian, H Dang, Z Chen. γ-Aminobutyric acid regulates both the survival and replication of human β-cells. Diabetes, 2013. [DOI | PubMed]
- SV Korol, Z Jin, Y Jin. Functional characterization of native, high-affinity GABAA receptors in human pancreatic β cells. EBioMedicine, 2018. [DOI | PubMed]
- RW Olsen. Allosteric ligands and their binding sites define γ-aminobutyric acid (GABA) type A receptor subtypes. Adv Pharmacol, 2015. [DOI | PubMed]
- G Bartholini. Pharmacology of the GABAergic system: effects of progabide, a GABA receptor agonist. Psychoneuroendocrinology, 1984. [DOI | PubMed]
- E Vashchinkina, A Panhelainen, OY Vekovischeva. GABA site agonist gaboxadol induces addiction-predicting persistent changes in ventral tegmental area dopamine neurons but is not rewarding in mice or baboons. J Neurosci, 2012. [DOI | PubMed]
- A Wahab, U Heinemann, K Albus. Effects of gamma-aminobutyric acid (GABA) agonists and a GABA uptake inhibitor on pharmacoresistant seizure like events in organotypic hippocampal slice cultures. Epilepsy Res, 2009. [DOI | PubMed]
- GAR Johnston. Advantages of an antagonist: bicuculline and other GABA antagonists. Br J Pharmacol, 2013. [DOI | PubMed]
- M-D Wang, M Rahman, D Zhu, T Bäckström. Pregnenolone sulphate and Zn2+ inhibit recombinant rat GABA(A) receptor through different channel property. Acta Physiol (Oxford), 2006. [DOI]
- 75.Vega Alanis BA, Iorio MT, Silva LL et al (2020) Allosteric GABA(A) receptor modulators-a review on the most recent heterocyclic chemotypes and their synthetic accessibility. Molecules 25. 10.3390/molecules25040999
- M Alqazzaz, AJ Thompson, KL Price. Cys-loop receptor channel blockers also block GLIC. Biophys J, 2011. [DOI | PubMed]
- EJ Hoffman, EW Warren. Flumazenil: a benzodiazepine antagonist. Clin Pharm, 1993. [PubMed]
- U Rudolph, F Knoflach. Beyond classical benzodiazepines: novel therapeutic potential of GABAA receptor subtypes. Nat Rev Drug Discov, 2011. [DOI | PubMed]
- O Henschel, KE Gipson, A Bordey. GABAA receptors, anesthetics and anticonvulsants in brain development. CNS Neurol Disord Drug Targets, 2008. [DOI | PubMed]
- GR Dawson, KA Maubach, N Collinson. An inverse agonist selective for alpha5 subunit-containing GABAA receptors enhances cognition. J Pharmacol Exp Ther, 2006. [DOI | PubMed]
- T Lynagh, SA Pless. Principles of agonist recognition in Cys-loop receptors. Front Physiol, 2014. [DOI | PubMed]
- HC Chua, M Chebib. GABA(A) Receptors and the diversity in their structure and pharmacology. Adv Pharmacol, 2017. [DOI | PubMed]
- SW Baumann, R Baur, E Sigel. Individual properties of the two functional agonist sites in GABA(A) receptors. J Neurosci, 2003. [DOI | PubMed]
- R Bergmann, K Kongsbak, PL Sørensen. A unified model of the GABAA receptor comprising agonist and benzodiazepine binding sites. PLoS One, 2013. [DOI | PubMed]
- KT Laha, PN Tran. Multiple tyrosine residues at the GABA binding pocket influence surface expression and mediate kinetics of the GABAA receptor. J Neurochem, 2013. [DOI | PubMed]
- H Möhler, JM Fritschy, U Rudolph. A new benzodiazepine pharmacology. J Pharmacol Exp Ther, 2002. [DOI | PubMed]
- U Rudolph, H Möhler. Analysis of GABAA receptor function and dissection of the pharmacology of benzodiazepines and general anesthetics through mouse genetics. Annu Rev Pharmacol Toxicol, 2004. [DOI | PubMed]
- M Ernst, D Brauchart, S Boresch, W Sieghart. Comparative modeling of GABA(A) receptors: limits, insights, future developments. Neuroscience, 2003. [DOI | PubMed]
- JA Benson, K Löw, R Keist. Pharmacology of recombinant gamma-aminobutyric acidA receptors rendered diazepam-insensitive by point-mutated alpha-subunits. FEBS Lett, 1998. [DOI | PubMed]
- JM Fritschy, H Mohler. GABAA-receptor heterogeneity in the adult rat brain: differential regional and cellular distribution of seven major subunits. J Comp Neurol, 1995. [DOI | PubMed]
- RM McKernan, TW Rosahl, DS Reynolds. Sedative but not anxiolytic properties of benzodiazepines are mediated by the GABA(A) receptor alpha1 subtype. Nat Neurosci, 2000. [DOI | PubMed]
- W Hevers, H Lüddens. The diversity of GABAA receptors. Pharmacological and electrophysiological properties of GABAA channel subtypes. Mol Neurobiol, 1998. [DOI | PubMed]
- RW Olsen, W Sieghart. International Union of Pharmacology. LXX. Subtypes of gamma-aminobutyric acid (A) receptors: classification on the basis of subunit composition, pharmacology, and function. Update. Pharmacol Rev, 2008. [DOI | PubMed]
- RW Olsen, W Sieghart. GABA A receptors: subtypes provide diversity of function and pharmacology. Neuropharmacology, 2009. [DOI | PubMed]
- MM Poe, KR Methuku, G Li. Synthesis and characterization of a novel γ-aminobutyric acid type A (GABA(A)) receptor ligand that combines outstanding metabolic stability, pharmacokinetics, and anxiolytic efficacy. J Med Chem, 2016. [DOI | PubMed]
- Z Varagic, J Ramerstorfer, S Huang. Subtype selectivity of α+β- site ligands of GABAA receptors: identification of the first highly specific positive modulators at α6β2/3γ2 receptors. Br J Pharmacol, 2013. [DOI | PubMed]
- X Simeone, DCB Siebert, K Bampali. Molecular tools for GABA(A) receptors: high affinity ligands for β1-containing subtypes. Sci Rep, 2017. [DOI | PubMed]
- F Sternfeld, RW Carling, RA Jelley. Selective, orally active gamma-aminobutyric acidA alpha5 receptor inverse agonists as cognition enhancers. J Med Chem, 2004. [DOI | PubMed]
- B Antkowiak, U Rudolph. New insights in the systemic and molecular underpinnings of general anesthetic actions mediated by γ-aminobutyric acid A receptors. Curr Opin Anaesthesiol, 2016. [DOI | PubMed]
- DJ Nutt, M Besson, SJ Wilson. Blockade of alcohol’s amnestic activity in humans by an alpha5 subtype benzodiazepine receptor inverse agonist. Neuropharmacology, 2007. [DOI | PubMed]
- U Rudolph, H Möhler. GABAA receptor subtypes: therapeutic potential in Down syndrome, affective disorders, schizophrenia, and autism. Annu Rev Pharmacol Toxicol, 2014. [DOI | PubMed]
- RW Olsen, G-D Li. GABA(A) receptors as molecular targets of general anesthetics: identification of binding sites provides clues to allosteric modulation. Can J Anaesth, 2011. [DOI | PubMed]
- SJ Mihic, Q Ye, MJ Wick. Sites of alcohol and volatile anaesthetic action on GABA(A) and glycine receptors. Nature, 1997. [DOI | PubMed]
- SA Forman, KW Miller. Mapping general anesthetic sites in heteromeric γ-aminobutyric acid type A receptors reveals a potential for targeting receptor subtypes. Anesth Analg, 2016. [DOI | PubMed]
- SA Forman, KW Miller. Anesthetic sites and allosteric mechanisms of action on Cys-loop ligand-gated ion channels. Can J Anaesth, 2011. [DOI | PubMed]
- PS Garcia, SE Kolesky, A Jenkins. General anesthetic actions on GABA(A) receptors. Curr Neuropharmacol, 2010. [DOI | PubMed]
- DC Chiara, Z Dostalova, SS Jayakar. Mapping general anesthetic binding site(s) in human α1β3 γ-aminobutyric acid type A receptors with [3H]TDBzl-etomidate, a photoreactive etomidate analogue. Biochemistry, 2012. [DOI | PubMed]
- DS Stewart, M Hotta, R Desai, SA Forman. State-dependent etomidate occupancy of its allosteric agonist sites measured in a cysteine-substituted GABAA receptor. Mol Pharmacol, 2013. [DOI | PubMed]
- DC Chiara, SS Jayakar, X Zhou. Specificity of intersubunit general anesthetic-binding sites in the transmembrane domain of the human α1β3γ2 γ-aminobutyric acid type A (GABAA) receptor. J Biol Chem, 2013. [DOI | PubMed]
- MM Eaton, LQ Cao, Z Chen. Mutational analysis of the putative high-affinity propofol binding site in human β3 homomeric GABAA receptors. Mol Pharmacol, 2015. [DOI | PubMed]
- NP Franks. Structural comparisons of ligand-gated ion channels in open, closed, and desensitized states identify a novel propofol-binding site on mammalian γ-aminobutyric acid type A receptors. Anesthesiology, 2015. [DOI | PubMed]
- GMS Yip, Z-W Chen, CJ Edge. A propofol binding site on mammalian GABAA receptors identified by photolabeling. Nat Chem Biol, 2013. [DOI | PubMed]
- R Jurd, M Arras, S Lambert. General anesthetic actions in vivo strongly attenuated by a point mutation in the GABA(A) receptor beta3 subunit. FASEB J Off Publ Fed Am Soc Exp Biol, 2003. [DOI]
- DS Reynolds, TW Rosahl, J Cirone. Sedation and anesthesia mediated by distinct GABA(A) receptor isoforms. J Neurosci, 2003. [DOI | PubMed]
- SA Thompson, PJ Whiting, KA Wafford. Barbiturate interactions at the human GABAA receptor: dependence on receptor subunit combination. Br J Pharmacol, 1996. [DOI | PubMed]
- MC Maldifassi, R Baur, E Sigel. Functional sites involved in modulation of the GABAA receptor channel by the intravenous anesthetics propofol, etomidate and pentobarbital. Neuropharmacology, 2016. [DOI | PubMed]
- DC Chiara, Y Jounaidi, X Zhou. General anesthetic binding sites in human α4β3δ γ-aminobutyric acid type A receptors (GABAARs). J Biol Chem, 2016. [DOI | PubMed]
- D Belelli, JJ Lambert. Neurosteroids: endogenous regulators of the GABA(A) receptor. Nat Rev Neurosci, 2005. [DOI | PubMed]
- G Akk, P Li, J Bracamontes. Mutations of the GABA-A receptor alpha1 subunit M1 domain reveal unexpected complexity for modulation by neuroactive steroids. Mol Pharmacol, 2008. [DOI | PubMed]
- AM Hosie, ME Wilkins, HMA da Silva, TG Smart. Endogenous neurosteroids regulate GABAA receptors through two discrete transmembrane sites. Nature, 2006. [DOI | PubMed]
- P Li, AK Bandyopadhyaya, DF Covey. Hydrogen bonding between the 17beta-substituent of a neurosteroid and the GABA(A) receptor is not obligatory for channel potentiation. Br J Pharmacol, 2009. [DOI | PubMed]
- AM Hosie, ME Wilkins, TG Smart. Neurosteroid binding sites on GABA(A) receptors. Pharmacol Ther, 2007. [DOI | PubMed]
- JR Hanrahan, M Chebib, GAR Johnston. Interactions of flavonoids with ionotropic GABA receptors. Adv Pharmacol, 2015. [DOI | PubMed]
- JR Hanrahan, M Chebib, GAR Johnston. Flavonoid modulation of GABA(A) receptors. Br J Pharmacol, 2011. [DOI | PubMed]
- G Wang, CG Clark, FG Rodgers. Detection in Escherichia coli of the genes encoding the major virulence factors, the genes defining the O157:H7 serotype, and components of the type 2 Shiga toxin family by multiplex PCR. J Clin Microbiol, 2002. [DOI | PubMed]
- SP Fernandez, KN Mewett, JR Hanrahan. Flavan-3-ol derivatives are positive modulators of GABA(A) receptors with higher efficacy for the alpha(2) subtype and anxiolytic action in mice. Neuropharmacology, 2008. [DOI | PubMed]
- F Wang, Z Xu, CT Yuen. 6,2’-Dihydroxyflavone, a subtype-selective partial inverse agonist of GABAA receptor benzodiazepine site. Neuropharmacology, 2007. [DOI | PubMed]
- D Kavvadias, P Sand, KA Youdim. The flavone hispidulin, a benzodiazepine receptor ligand with positive allosteric properties, traverses the blood-brain barrier and exhibits anticonvulsive effects. Br J Pharmacol, 2004. [DOI | PubMed]
- Z Atakan. Cannabis, a complex plant: different compounds and different effects on individuals. Ther Adv Psychopharmacol, 2012. [DOI | PubMed]
- G Ruffolo, P Cifelli, C Roseti. A novel GABAergic dysfunction in human Dravet syndrome. Epilepsia, 2018. [DOI | PubMed]
- 131.An D, Peigneur S, Hendrickx LA, Tytgat J (2020) Targeting cannabinoid receptors: current status and prospects of natural products. Int J Mol Sci 21. 10.3390/ijms21145064
- DM Lambert, CJ Fowler. The endocannabinoid system: drug targets, lead compounds, and potential therapeutic applications. J Med Chem, 2005. [DOI | PubMed]
- I Katona, TF Freund. Multiple functions of endocannabinoid signaling in the brain. Annu Rev Neurosci, 2012. [DOI | PubMed]
- E Sigel, R Baur, I Rácz. The major central endocannabinoid directly acts at GABA(A) receptors. Proc Natl Acad Sci U S A, 2011. [DOI | PubMed]
- R Baur, J Gertsch, E Sigel. Do N-arachidonyl-glycine (NA-glycine) and 2-arachidonoyl glycerol (2-AG) share mode of action and the binding site on the β2 subunit of GABAA receptors?. PeerJ, 2013. [DOI | PubMed]
- T Golovko, R Min, N Lozovaya. Control of inhibition by the direct action of cannabinoids on GABAA receptors. Cereb Cortex, 2015. [DOI | PubMed]
- SWM Crossley, G Tong, MJ Lambrecht. Synthesis of (-)-picrotoxinin by late-stage strong bond activation. J Am Chem Soc, 2020. [DOI | PubMed]
- J Cao, W Thor, S Yang. Synthesis of the tricyclic picrotoxane motif by an oxidative cascade cyclization. Org Lett, 2019. [DOI | PubMed]
- E Nilsson, B Eyrich. On treatment of barbiturate poisoning. Acta Med Scand, 1950. [DOI | PubMed]
- D Pericić, K Mirković, M Jazvinsćak, F Besnard. [3H]t-butylbicycloorthobenzoate binding to recombinant alpha1beta2gamma2s GABA(A) receptor. Eur J Pharmacol, 1998. [DOI | PubMed]
- AV Kalueff. Mapping convulsants’ binding to the GABA-A receptor chloride ionophore: a proposed model for channel binding sites. Neurochem Int, 2007. [DOI | PubMed]
- MJ Jembrek, J Vlainic. GABA receptors: pharmacological potential and pitfalls. Curr Pharm Des, 2015. [DOI | PubMed]
- NA Othman, M Gallacher, TZ Deeb. Influences on blockade by t-butylbicyclo-phosphoro-thionate of GABA(A) receptor spontaneous gating, agonist activation and desensitization. J Physiol, 2012. [DOI | PubMed]
- N Brown, J Kerby, TP Bonnert. Pharmacological characterization of a novel cell line expressing human alpha(4)beta(3)delta GABA(A) receptors. Br J Pharmacol, 2002. [DOI | PubMed]
- SI Stórustovu, B Ebert. Pharmacological characterization of agonists at delta-containing GABAA receptors: functional selectivity for extrasynaptic receptors is dependent on the absence of gamma2. J Pharmacol Exp Ther, 2006. [DOI | PubMed]
- ML Jensen, KA Wafford, AR Brown. A study of subunit selectivity, mechanism and site of action of the delta selective compound 2 (DS2) at human recombinant and rodent native GABA(A) receptors. Br J Pharmacol, 2013. [DOI | PubMed]
- MP Jacobs, GG Leblanc, A Brooks-Kayal. Curing epilepsy: progress and future directions. Epilepsy Behav, 2009. [DOI | PubMed]
- DC Hesdorffer, V Beck, CE Begley. Research implications of the Institute of Medicine report, epilepsy across the spectrum: promoting health and understanding. Epilepsia, 2013. [DOI | PubMed]
- Y Ben-Ari, J-L Gaiarsa, R Tyzio, R Khazipov. GABA: a pioneer transmitter that excites immature neurons and generates primitive oscillations. Physiol Rev, 2007. [DOI | PubMed]
- K Kaila, E Ruusuvuori, P Seja. GABA actions and ionic plasticity in epilepsy. Curr Opin Neurobiol, 2014. [DOI | PubMed]
- E Palma, G Ruffolo, P Cifelli. Modulation of GABAA receptors in the treatment of epilepsy. Curr Pharm Des, 2017. [DOI | PubMed]
- SM Janković, M Dješević, SV Janković. Experimental GABA A receptor agonists and allosteric modulators for the treatment of focal epilepsy. J Exp Pharmacol, 2021. [DOI | PubMed]
- S Braat, RF Kooy. The GABAA receptor as a therapeutic target for neurodevelopmental disorders. Neuron, 2015. [DOI | PubMed]
- S Hirose. Mutant GABA(A) receptor subunits in genetic (idiopathic) epilepsy. Prog Brain Res, 2014. [DOI | PubMed]
- M Mele, RO Costa, CB Duarte. Alterations in GABA(A)-receptor trafficking and synaptic dysfunction in brain disorders. Front Cell Neurosci, 2019. [DOI | PubMed]
- J-Q Kang, W Shen, C Zhou. The human epilepsy mutation GABRG2(Q390X) causes chronic subunit accumulation and neurodegeneration. Nat Neurosci, 2015. [DOI | PubMed]
- B Dejanovic, T Djémié, N Grünewald. Simultaneous impairment of neuronal and metabolic function of mutated gephyrin in a patient with epileptic encephalopathy. EMBO Mol Med, 2017. [DOI | PubMed]
- G Choii, J Ko. Gephyrin: a central GABAergic synapse organizer. Exp Mol Med, 2015. [DOI | PubMed]
- M Fang, L Shen, H Yin. Downregulation of gephyrin in temporal lobe epilepsy neurons in humans and a rat model. Synapse, 2011. [DOI | PubMed]
- IW Hamley. The amyloid beta peptide: a chemist’s perspective. Role in Alzheimer’s and fibrillization. Chem Rev, 2012. [DOI | PubMed]
- Y Xu, M Zhao, Y Han, H Zhang. GABAergic inhibitory interneuron deficits in Alzheimer’s disease: implications for treatment. Front Neurosci, 2020. [DOI | PubMed]
- L Verret, EO Mann, GB Hang. Inhibitory interneuron deficit links altered network activity and cognitive dysfunction in Alzheimer model. Cell, 2012. [DOI | PubMed]
- N Craddock, L Jones, IR Jones. Strong genetic evidence for a selective influence of GABA A receptors on a component of the bipolar disorder phenotype. Mol Psychiatry, 2010. [DOI | PubMed]
- S Frere, I Slutsky. Alzheimer’s disease: from firing instability to homeostasis network collapse. Neuron, 2018. [DOI | PubMed]
- DJ Selkoe. Early network dysfunction in Alzheimer’s disease. Science, 2019. [DOI | PubMed]
- 166.Govindpani K, Turner C, Waldvogel HJ et al (2020) Impaired expression of GABA signaling components in the Alzheimer’s disease middle temporal gyrus. Int J Mol Sci 21. 10.3390/ijms21228704
- MC Gueli, G Taibi. Alzheimer’s disease: amino acid levels and brain metabolic status. Neurol Sci Off J Ital Neurol Soc Ital Soc Clin Neurophysiol, 2013. [DOI]
- SR Bareggi, M Franceschi, L Bonini. Decreased CSF concentrations of homovanillic acid and gamma-aminobutyric acid in Alzheimer’s disease. Age- or disease-related modifications?. Arch Neurol, 1982. [DOI | PubMed]
- D Grouselle, R Winsky-Sommerer, JP David. Loss of somatostatin-like immunoreactivity in the frontal cortex of Alzheimer patients carrying the apolipoprotein epsilon 4 allele. Neurosci Lett, 1998. [DOI | PubMed]
- A Limon, JM Reyes-Ruiz, R Miledi. Loss of functional GABA(A) receptors in the Alzheimer diseased brain. Proc Natl Acad Sci U S A, 2012. [DOI | PubMed]
- S-Q Zhang, D Obregon, J Ehrhart. Baicalein reduces β-amyloid and promotes nonamyloidogenic amyloid precursor protein processing in an Alzheimer’s disease transgenic mouse model. J Neurosci Res, 2013. [DOI | PubMed]
- 172.Defazio G, Jankovic J, Giel JL, Papapetropoulos S (2013) Descriptive epidemiology of cervical dystonia. Tremor Other Hyperkinet Mov 3. 10.7916/D80C4TGJ
- HA Jinnah, A Berardelli, C Comella. The focal dystonias: current views and challenges for future research. Mov Disord, 2013. [DOI | PubMed]
- HA Jinnah, SA Factor. Diagnosis and treatment of dystonia. Neurol Clin, 2015. [DOI | PubMed]
- BD Berman, RT Pollard, E Shelton. GABA(A) Receptor availability changes underlie symptoms in isolated cervical dystonia. Front Neurol, 2018. [DOI | PubMed]
- C Dresel, Y Li, V Wilzeck. Multiple changes of functional connectivity between sensorimotor areas in focal hand dystonia. J Neurol Neurosurg Psychiatry, 2014. [DOI]
- D Guehl, P Burbaud, T Boraud, B Bioulac. Bicuculline injections into the rostral and caudal motor thalamus of the monkey induce different types of dystonia. Eur J Neurosci, 2000. [DOI | PubMed]
- CL Groth, M Brown, JM Honce. Cervical dystonia is associated with aberrant inhibitory signaling within the thalamus. Front Neurol, 2021. [DOI]
- AN Clarkson, BS Huang, SE Macisaac. Reducing excessive GABA-mediated tonic inhibition promotes functional recovery after stroke. Nature, 2010. [DOI | PubMed]
- C Wu, D Sun. GABA receptors in brain development, function, and injury. Metab Brain Dis, 2015. [DOI | PubMed]
- A El-Ansary, L Al-Ayadhi. GABAergic/glutamatergic imbalance relative to excessive neuroinflammation in autism spectrum disorders. J Neuroinflammation, 2014. [DOI | PubMed]
- R Pizzarelli, E Cherubini. Alterations of GABAergic signaling in autism spectrum disorders. Neural Plast, 2011. [DOI | PubMed]
- MA Mendez, J Horder, J Myers. The brain GABA-benzodiazepine receptor alpha-5 subtype in autism spectrum disorder: a pilot [(11)C]Ro15-4513 positron emission tomography study. Neuropharmacology, 2013. [DOI | PubMed]
- 184.Horder J, Andersson M, Mendez MA et al (2018) GABA(A) receptor availability is not altered in adults with autism spectrum disorder or in mouse models. Sci Transl Med 10. 10.1126/scitranslmed.aam8434
- M Mahdavi, M Kheirollahi, R Riahi. Meta-analysis of the association between GABA receptor polymorphisms and autism spectrum disorder (ASD). J Mol Neurosci, 2018. [DOI | PubMed]
- TM Mueller, JH Meador-Woodruff. Post-translational protein modifications in schizophrenia. NPJ Schizophr, 2020. [DOI | PubMed]
- SE Williams, RG Mealer, EM Scolnick. Aberrant glycosylation in schizophrenia: a review of 25 years of post-mortem brain studies. Mol Psychiatry, 2020. [DOI | PubMed]
- TM Mueller, CE Remedies, V Haroutunian, JH Meador-Woodruff. Abnormal subcellular localization of GABAA receptor subunits in schizophrenia brain. Transl Psychiatry, 2015. [DOI | PubMed]
- TM Mueller, V Haroutunian, JH Meador-Woodruff. N-Glycosylation of GABAA receptor subunits is altered in schizophrenia. Neuropsychopharmacol Off Publ Am Coll Neuropsychopharmacol, 2014. [DOI]
- 190.Marques TR, Ashok AH, Angelescu I et al (2020) GABA-A receptor differences in schizophrenia: a positron emission tomography study using [(11)C]Ro154513. Mol Psychiatry. 10.1038/s41380-020-0711-y
- MV Fogaça, RS Duman. Cortical GABAergic Dysfunction in stress and depression: new insights for therapeutic interventions. Front Cell Neurosci, 2019. [DOI | PubMed]
- YS Kim, B-E Yoon. Altered GABAergic signaling in brain disease at various stages of life. Exp Neurobiol, 2017. [DOI | PubMed]
- ML Auger, J Meccia, SB Floresco. Regulation of sustained attention, false alarm responding and implementation of conditional rules by prefrontal GABA(A) transmission: comparison with NMDA transmission. Psychopharmacology, 2017. [DOI | PubMed]
- TA Paine, S Chang, R Poyle. Contribution of GABA(A) receptor subunits to attention and social behavior. Behav Brain Res, 2020. [DOI | PubMed]
- TA Paine, LE Slipp, WAJ Carlezon. Schizophrenia-like attentional deficits following blockade of prefrontal cortex GABAA receptors. Neuropsychopharmacol Off Publ Am Coll Neuropsychopharmacol, 2011. [DOI]
- SK Asinof, TA Paine. Inhibition of GABA synthesis in the prefrontal cortex increases locomotor activity but does not affect attention in the 5-choice serial reaction time task. Neuropharmacology, 2013. [DOI | PubMed]
- AL Pehrson, CO Bondi, NKB Totah, B Moghaddam. The influence of NMDA and GABA(A) receptors and glutamic acid decarboxylase (GAD) activity on attention. Psychopharmacology, 2013. [DOI | PubMed]
- TA Paine, N Swedlow, L Swetschinski. Decreasing GABA function within the medial prefrontal cortex or basolateral amygdala decreases sociability. Behav Brain Res, 2017. [DOI | PubMed]
- PT Piantadosi, S Khayambashi, MG Schluter. Perturbations in reward-related decision-making induced by reduced prefrontal cortical GABA transmission: relevance for psychiatric disorders. Neuropharmacology, 2016. [DOI | PubMed]