The BRCAness Landscape of Cancer
Abstract
BRCAness refers to the damaged homologous recombination (HR) function due to the defects in HR-involved non-BRCA1/2 genes. BRCAness is the important marker for the use of synthetic lethal-based PARP inhibitor therapy in breast and ovarian cancer treatment. The success provides an opportunity of applying PARP inhibitor therapy to treat other cancer types with BRCAness features. However, systematic knowledge is lack for BRCAness in different cancer types beyond breast and ovarian cancer. We performed a comprehensive characterization for 40 BRCAness-related genes in 33 cancer types with over 10,000 cancer cases, including pathogenic variation, homozygotic deletion, promoter hypermethylation, gene expression, and clinical correlation of BRCAness in each cancer type. Using BRCA1/BRCA2 mutated breast and ovarian cancer as the control, we observed that BRCAness is widely present in multiple cancer types. Based on the sum of the BRCAneass features in each cancer type, we identified the following 21 cancer types as the potential targets for PARPi therapy: adrenocortical carcinoma, bladder urothelial carcinoma, brain lower grade glioma, colon adenocarcinoma, esophageal carcinoma, head and neck squamous carcinoma, kidney chromophobe, kidney renal clear cell carcinoma, kidney renal papillary cell carcinoma, liver hepatocellular carcinoma, lung adenocarcinoma, lung squamous cell carcinoma, mesothelioma, rectum adenocarcinoma, pancreatic adenocarcinoma, prostate adenocarcinoma, sarcoma, skin cutaneous melanoma, stomach adenocarcinoma, uterine carcinosarcoma, and uterine corpus endometrial carcinoma.
Article type: Research Article
Keywords: BRCAness, genetic defects, synthetic lethal, PARP inhibitors
License: © 2022 by the authors. CC BY 4.0 Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Article links: DOI: 10.3390/cells11233877 | PubMed: 36497135 | PMC: PMC9738094
Relevance: Moderate: mentioned 3+ times in text
Full text: PDF (3.6 MB)
1. Introduction
BRCA1 and BRCA2 (BRCA1/2) play essential roles in repairing double-strand DNA breaks through homologous recombination (HR) [ref. 1,ref. 2]. Pathogenic variants in BRCA1/2 damage their function, leads to genome instability, and increased the risk of breast and ovarian cancer [ref. 3,ref. 4,ref. 5,ref. 6]. The defected BRCA1/2 is the specific marker for the use of a synthetic lethal-based PARPi (poly ADP ribose polymerase inhibitors) therapy in cancer treatment. In the process, the PARP inhibitors block PARP function of repairing single-strand breaks and cause the formation of double-strand break upon replication, which cannot be repaired by homologous recombination due to the defected BRCA1/2 function leading to the death of cancer cells [ref. 7,ref. 8,ref. 9,ref. 10,ref. 11]. BRCA1/2 defects are present in about 5% of breast cancer and 20% of ovarian cancer patients, which are the major beneficiaries for the PARP inhibitor therapy [ref. 12,ref. 13,ref. 14].
Homologous recombination pathway involves multiple genes besides BRCA1/2. Further, many genes not in HR pathway can also directly or indirectly be involved in HR [ref. 15]. In principle, defects in these non-BRCA1/2 genes could also result in the same consequences caused by the detected BRCA1/2. This is described as BRCAness [ref. 16]. Based on the studies in breast and ovarian cancer, BRCAness are present in about 20% of breast cancer and 45% of ovarian cancer [ref. 17,ref. 18], such as the mutations in RAD51C, NBS1, BRIP1, and PALB2 [ref. 19]. Cancer with defected BRCAness shared many features such as the cancer with defected BRCA1/2. For instance, BRCAness cancer genome had high abundant C-to-T transitions [ref. 20], shared the “Signature 3” of mutation signatures in cancer [ref. 21,ref. 22], and contained high level of promoter hypermethylation [ref. 23], prone to triple-negative breast cancer (TNBC) and “Basal” subtypes, and benefited from PARPi treatment [ref. 24,ref. 25,ref. 26,ref. 27,ref. 28].
The success of PARPi therapy in BRCA1/2 defected breast and ovarian cancer attracts great interests in exploring its potential to treat other cancer types with BRCAness features. Indeed, it was observed that BRCAness was present in prostate cancer, colon cancer and pancreatic cancer [ref. 29,ref. 30,ref. 31], PARP inhibitor treatment improved survival of pancreatic cancer carrying mutated HRR deficient-related genes of ATM, BRCA1/2, CHEK2, PALB2, RAD51C, RAD51D [ref. 32], and enhanced treatment response in metastatic prostate cancer carrying mutated HRR deficient-related genes ATM, BRCA1/2, BRIP1, BARD1, CDK12, CHEK1, CHEK2, FANCL, PALB2, RAD5C [ref. 33,ref. 34,ref. 35]. However, systematic knowledge is lack for BRCAness across cancer type spectrum. This largely restricts the use of synthetic lethal-based PARPi therapy as a universal option in cancer treatment.
We hypothesized that BRCAness could be a common phenomenon in cancer. To test our hypothesis, we performed a BRCAness characterization in 33 cancer types, covering pathogenic variation, homozygotic deletion, promoter hypermethylation and expression, copy number variation, and clinical correlation for BRCAness-related genes. By using BRCA1/2-mutated breast and ovarian cancer as the references, we observed the wide presence of BRCAness signatures in multiple cancer types. Our study provides a foundation to further test the potential of using PARPi therapy to treat the cancer types with BRCAness features.
2. Materials and Methods
2.1. Sources of Genome Data across 33 Cancer Types
We identified 40 BRCAness genes from literatures (ATM, ATR, AURKA, BAP1, BARD1, BLM, BRCA1, BRCA2, BRIP1, CDK12, CHD4, CHEK1, CHEK2, EMSY, ERCC1, FANCA, FANCC, FANCD2, FANCE, FANCF, FANCI, KMT2A, MRE11A, MYC, NBS1, PALB2, PARP1, PAXIP1, PLK1, PTEN, RAD50, RAD51, RAD51B, RAD51C, RAD51D, RAD52, SAMHD1, SEM1, TP53, TP53BP1, WEE1, WRN) [ref. 16,ref. 36,ref. 37]. We collected the BRCAness genomic and clinical information from these two resources: UCSC xena (http://xena.ucsc.edu/, accessed on 18 November 2020) and PanCanAtlas (https://gdc.cancer.gov/node/905/, accessed on 20 November 2020) covering 33 cancer types [ref. 38]. The details of data information are as follows: Variation data from over 10,000 cancer patients were from TCGA MAF file in PanCanAtlas; copy number variation (CNV) data detected by Affymetrix SNP 6.0 arrays were from UCSC xena; DNA methylation data detected by Illumina HumanMethylation450 BeadChip platform were from PanCanAtlas; RNA-seq data with normalized batch effects and log2 (norm_value+1) gene expression for all 33 cancer types were from PanCanAtlas; Clinical survival data were from UCSC xena. GISTIC2 was used to identify the genomic regions with significant gain or loss [ref. 39].
2.2. Pathogenic Variant Data in BRCAness Genes
To identify pathogenic and likely pathogenic variants (PLPs) for each BRCAness gene across each cancer type, we first extracted the variants that passed filtering and belonged to non-silent subtypes from TCGA pan-cancer variation data. We further downloaded all variations from ClinVar (https://www.ncbi.nlm.nih.gov/clinvar/, accessed on 25 January 2021) and COSMIC (the Catalogue of Somatic Mutations in Cancer, https://cancer.sanger.ac.uk/cosmic, accessed on 25 January 2021) databases and extracted the “Pathogenic” and “Likely Pathogenic” variants from ClinVar and “Pathogenic” variants from COSMIC (PLP). We compared the TCGA variants to ClinVar/COSMIC PLP variants to identify germline and somatic PLPs for the BRCAness genes in each cancer type. We calculated variation frequency for the PLP variants in each BRCAness gene in each cancer type and tested the correlation of PLP variation frequencies between BRCAness genes and BRCA1/2 in all 33 cancer types by Pearson’s correlation coefficient analysis. p-value < 0.05 was considered as statistically significant.
2.3. CNV Data in BRCAness Genes
We used GISTIC2 to identify CNV at gene-level [ref. 39], with gene count of “−2” defined as homozygotic mutation. We retained homozygous deletion for CNV analysis across all 33 cancer types. We calculated homozygous deletion frequency for each BRCAness gene in each cancer type, and clustered homozygous deletion frequencies for all 33 cancer types using R package “ComplexHeatmap” [ref. 40]. We also tested the correlation of frequencies for homozygous deletions between BRCA1/2 and BRCAness genes in all 33 cancer types by Pearson’s correlation coefficient analysis. p-value < 0.05 was used as statistically significant in correlation analyses.
2.4. DNA Methylation Data in BRCAness Genes
We used DNA methylation data from the HumanMethylation450 arrays. We mapped the probes to each BRCAness genes using the Illumina GPL13534 platform (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GPL13534, accessed on 23 May 2021). Promoter for BRCA1/2 and BRCAness genes was defined as within 10 kb surrounding transcription start site (TSS) for each gene. We obtained hypermethylated data with mean gene-level promoter methylation values > 0.3. We clustered promoter methylation levels (mean promoter methylation values) for BRCA1/2 and BRCAness genes in 33 cancer types using R package “ComplexHeatmap” [ref. 40].
2.5. Gene Expression and Functional Enrichment Analysis
We used the Wilcox’s rank sum test to identify differentially expressed genes (DEGs) in each cancer type. We adjusted the p-values by Benjamini–Hochberg (BH) method [ref. 41]. We defined the differentially expressed genes with adjusted p-values < 0.01 and at least two-fold changes in expression level. We also performed Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) functional enrichment annotation using the DAVID tool (http://david.abcc.ncifcrf.gov/, accessed on 23 May 2021, version 6.8), with Benjamini p.adjust < 0.05 as statistically significance [ref. 42].
2.6. Clinical Relevance of BRCAness Genes
We divided the patients into high-risk and low-risk groups based on the median expression of each BRCAness gene. We used the Kaplan–Meier method to generate survival curves, and two-sided log-rank tests to assess the differences in overall survival between the high-risk and low-risk groups by using the R package “survival” and p-value < 0.05 as significant difference. We calculated the hazard ratio and 95% confidence level and plotted the results by R “forestplot” package.
2.7. Identifying Candidate Cancer Types for PARPi Therapy
Taking breast cancer and ovarian cancer as the references, we ranked the 33 cancer types based on following six integrated features: for somatic and germline pathogenic variation, and homozygous deletion, we calculated the frequencies of all BRCAness genes in each cancer type; for promoter methylation, we calculated the total promoter methylation level of all BRCAness genes in each cancer type; for gene expression, we calculated the total number of differentially expressed BRCAness genes in each cancer type; for prognostics, we located the number of risky BRCAness genes in each cancer type. We set BRCA/OV = 1 to identify BRCAness cancer types that only the cancer type with >=1 qualified as BRCAness cancer types.
3. Results
3.1. Overview of the Study
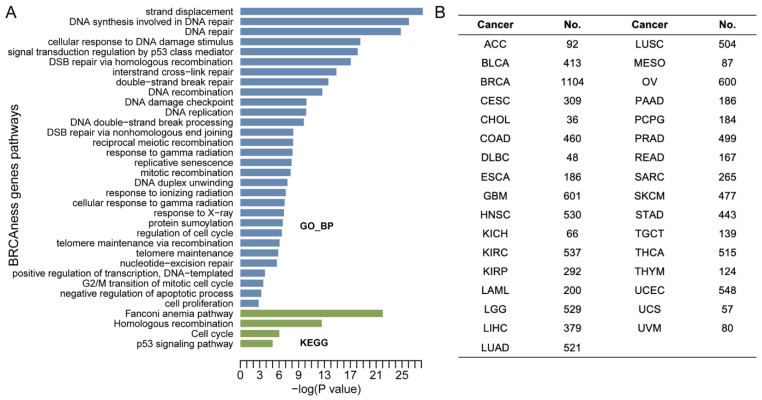
Functional enrichment analysis showed that the 40 BRCAness genes were involved in homologous recombination, DNA repair, cell cycle, and Fanconi anemia pathways (Figure 1A). Using the TCGA multi-omics data from 33 cancer types derived from over 10,000 cancer cases, we characterized BRCAness genes and their clinical relevance (Figure 1B, Table S1) by focusing on six characters: pathogenic variation, CNV, homozygosity, DNA methylation and gene expression, and clinical prognosis. In each analysis, BRCA1/2 status was used as gene-level controls, and BRCA1/2 mutated-breast and ovarian cancer were used as the cancer type-level controls.
3.2. Pathogenic Variation in BRCAness Genes in Different Cancer Types
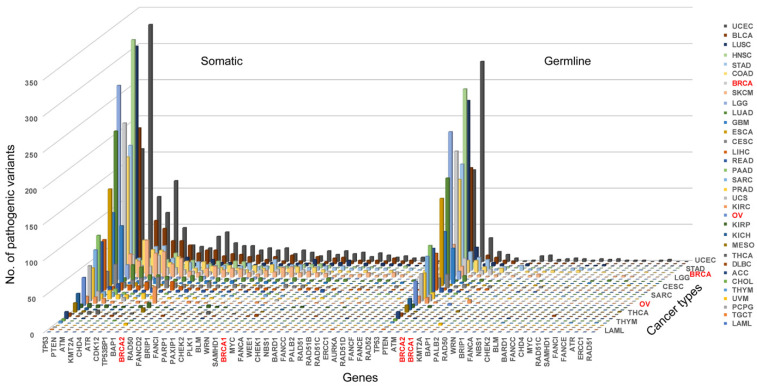
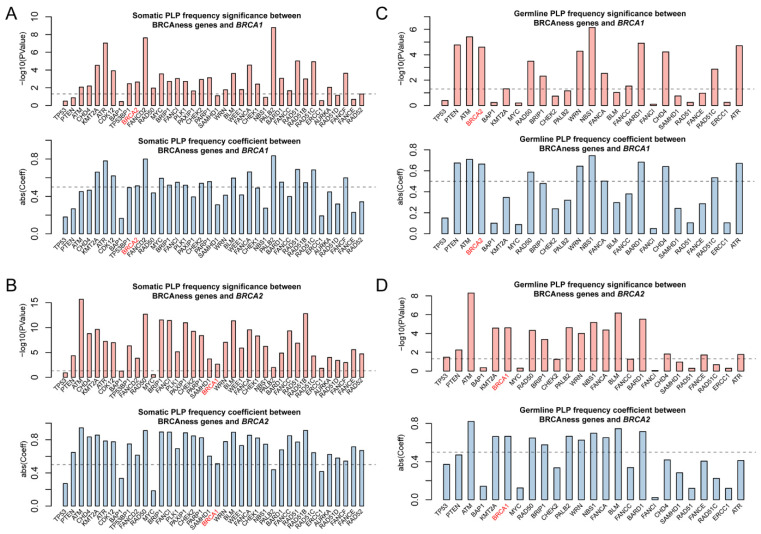
Pathogenic variation in BRCAness gene is the direct indication for the presence of BRCAness in cancer. We searched for both germline and somatic pathogenic variants in BRCA1/2 and BRCAness genes in each of the 33 cancer types and identified a total of 808 germline and 4017 somatic pathogenic mutations in BRCA1, BRCA2 and 37 of the 38 BRCAness genes distributed in 33 cancer types. On average, there were 24 germline and 122 somatic mutations per cancer type distributed at different frequencies in different cancer types (Tables S2–S4). As expected, BRCA1/2 had both germline and somatic pathogenic variation in breast and ovarian cancer, whereas BRCAness genes had high prevalence of somatic pathogenic variation distributed in cancer type-specific manners (Figure 2, Table S2). The prevalence of somatic pathogenic variation in BRCAness genes ranged between 0.14% and 39.81%, with TP53 as the highest among all BRCAness genes, followed by PTEN, ATM, CHD4, KMT2A, ATR, CDK12, BAP1 and TP53BP1 higher than BRCA2, and FANCD2, RAD50, MYC, BRIP1, FANCI, PLK1, PAXIP1, CHEK2, PARP1 and SAMHD1, all of which were higher than BRCA1 (Figure 2, Table S2). Of the 33 cancer types, 6 had higher prevalence of BRCAness pathogenic variation than breast cancer (BRCA) and 19 higher than ovarian cancer (OV), with UCEC as the highest of 1787 among all 33 cancer types (Figure 2, Table S2). LAML, TGCT, PCPG, UVM, CHOL, and THYM had very low prevalence of BRCAness somatic and germline pathogenic variation. For example, LAML (acute myeloid leukemia) had neither somatic nor germline pathogenic variation in BRCAness genes, although AML has consistent cytogenetic abnormality (Figure 2, Table S2). Therefore, these cancer types were unlikely relevant with BRCAness. Then, we performed the variation frequency correlation analysis to explore whether similar variant patterns exist between BRCAness genes and BRCA1/2. We performed Pearson correlation analysis to test the correlation between BRCAness pathogenic variation and BRCA1/2 genes. Except TP53, BAP1, PALB2, ERCC1 with the somatic pathogenic variation, nearly all BRCAness genes had significant correlation with BRCA1 and/or BRCA2 (Figure 3A,B), and about a third of BRCAness genes with germline pathogenic variation had significant correlation with BRCA1 and/or BRCA2 (Figure 3C,D).
3.3. Homozygotic Deletion Patterns between BRCA1/2 and BRCAness Genes in Different Cancer Types
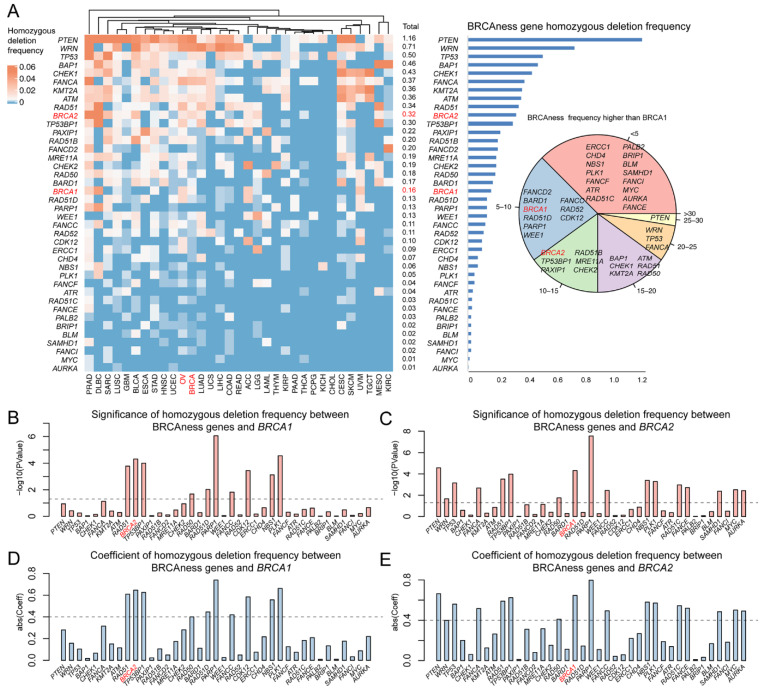
Pathogenic variation in human BRCA is predominantly heterozygotic due to the embryo lethal effects of BRCA1/2 homozygotic variation. Therefore, homozygosity provides an important indicator to test the similarity between BRCAness and BRCA1/2 pathogenic variation. From the homozygotic deletion identified in BRCAness genes, we observed low frequency of 0.01–1.16% homozygotic deletion in BRCAness genes, comparing to 0.16% in BRCA1 and 0.32% in BRCA2 (Figure 4A). Of the 40 BRCAness genes, 21 (RAD51D, PARP1, WEE1, FANCC, RAD52, CDK12, ERCC1, CHD4, NBS1, PLK1, FANCF, ATR, RAD51C, FANCE, PALB2, BRIP1, BLM, SAMHD1, FANCI, MYC, AURKA) had lower than the 0.16% in BRCA1 and 29 (TP53BP1, PAXIP1, RAD51B, FANCD2, MRE11A, CHEK2, RAD50, BARD1, RAD51D, PARP1, WEE1, FANCC, RAD52, CDK12, ERCC1, CHD4, NBS1, PLK1, FANCF, ATR, RAD51C, FANCE, PALB2, BRIP1, BLM, SAMHD1, FANCI, MYC, AURKA) had lower than the 0.32% in BRCA2 (Figure 4A). Nine BRCAness genes had significant correlation with BRCA1 and 15 with BRCA2 (Figure 4B–E). Of the 33 cancer types, 23 (LUAD, KIRC, STAD, ESCA, UVM, TGCT, ACC, COAD, LIHC, HNSC, UCEC, LGG, GBM, UCS, LAML, READ, KIRP, THYM, PAAD, THCA, PCPG, KICH, CHOL) were higher than BRCA cancer and OV cancer (Table S5). The results showed that like BRCA and the related cancer types, homozygotic variation was insignificantly present in BRCAness genes and their related cancer types.
3.4. Methylation and Expression Patterns between BRCA1/2 and BRCAness Genes in Different Cancer Types
Promoter methylation plays important roles in de-regulation of BRCA1/2 expression in breast and ovarian cancer [ref. 43]. We investigated promoter methylation in BRCAness genes across all 33 cancer types. By using promoter methylation level in BRCA2 as the cutoff, 16 of the 40 BRCAness gene promoters were hypermethylated across nearly all cancer types (Figure 5A, Table S6). We then tested the effects of promoter hypermethylation on BRCAness gene expression. In the 19 cancer types with expression data available from at least 5 normal samples as control, 10 cancer types of PRAD, READ, KIRP, COAD, STAD, LUAD, THCA, LIHC, ESCA, PAAD had lower expression of BRCAness genes than their normal controls by referring to breast cancer (ovarian cancer was not included due to the lack of expression data from normal ovarian control) (Figure 5B). Of the 33 cancer types, BRCA cancer had total levels of BRCA1/2 and BRCAness gene promoter hypermethylation of 14.244 and OV cancer had 13.926. By using OV cancer as the cutoff, 20 of the 33 cancer types had promoter hypermethylation (Table S6). The hypermethylation in OV had distinct features from other cancer types: a group of BRCAness genes of MER11A, RAD51, PALB2, CHD4, BRIP1, MYC, and FANCE were increasingly expressed whereas another group of WRN, CDK12, CHEK2, FANCA, and TP53 was decreasingly expressed (Figure 5C). Each cancer type also had specific hypermethylated BRCAness genes (Figure 5C). For example, FANCF promoter was hypermethylated and had decreased expression in eight cancer types of BLCA, CESC, COAD, DLBC, ESCA, HNSC, UCEC, and UCS (Figure 5C). The results highlighted that BRCA1/2 and BRCAness genes shared high similarity of promoter hypermethylation across multiple cancer types.
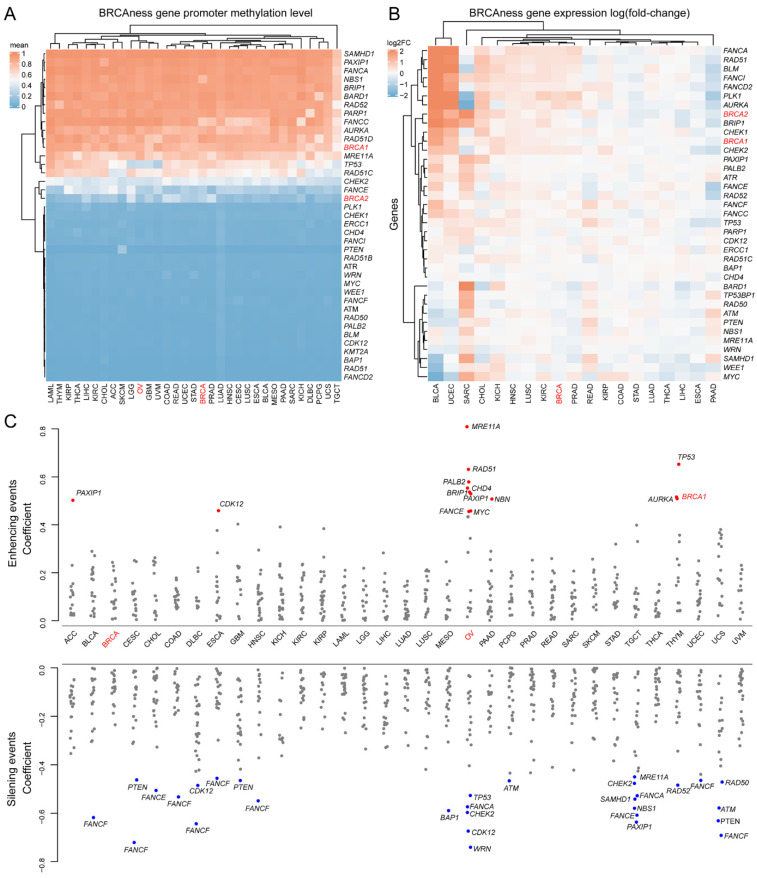
3.5. Prognostics of BRCAness Gene Expression across Different Cancer Types
We analyzed the prognostic potential of altered BRCAness gene expression across different cancer types. We observed that the altered expression of each BRCAness gene was significantly correlated with the overall survival in at least one cancer type except TGCT, THCA, and UCS (Figure 6A). For example, increased expression of AURKA was correlated with worse survival in 16 of the 33 cancer types including KIRP (log-rank p = 3.41 × 10−8, KICH (log-rank p = 5.20 × 10−6), and KIRC (log-rank p = 1.11 × 10−10) (Figure 6B); increased expression of PTEN was correlated with better survival in 6 cancer types including LGG (log-rank p = 4.44 × 10−9), KIRC (log-rank p = 4.21 × 10−3), LIHC (log-rank p = 2.64 × 10−2), UCEC (log-rank p = 4.25 × 10−5), PRAD (log-rank p = 7.07 × 10−2) and LAML (log-rank p = 3.64 × 10−2) (Figure 6B). Altered expression of BRCAness genes can classify cancer patients into high- and low-risk groups. For example, based on altered expression of BRCAness genes, KIRC patients were divided into the lower expression group of 112 patients and the higher expression group of 422 patients (Figure 6C), in which the lower expression subgroup had significantly better survival than the higher expression subgroup (Figure 6D and Figure S1). The results showed that similar to BRCA1/2 in breast and ovarian cancer, BRCAness genes were the prognostic markers for multiple cancer types.
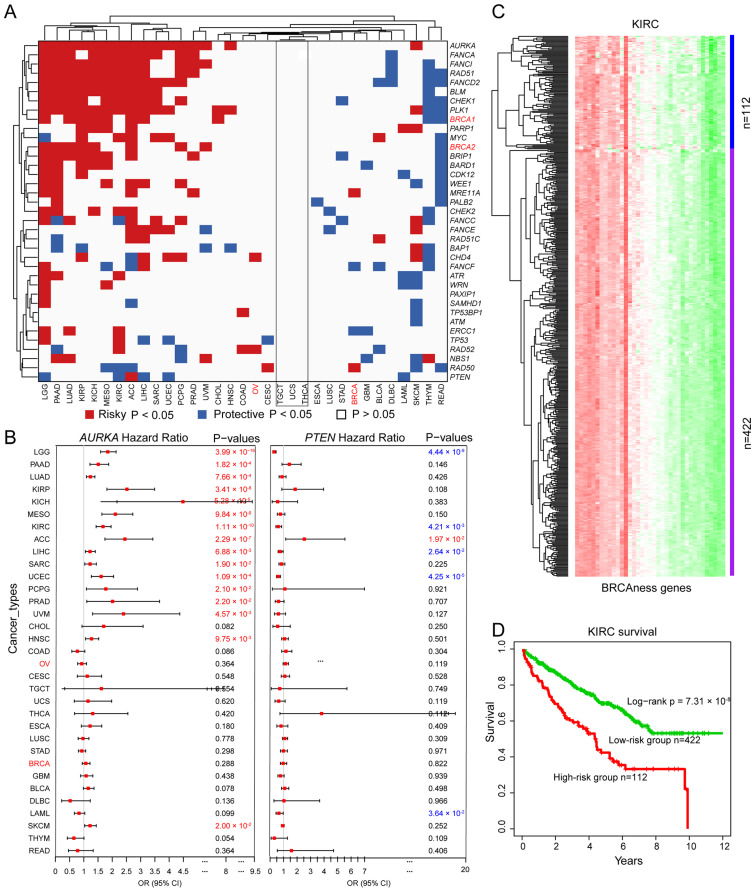
3.6. BRCAness Cancer Types Sharing High Similarity with BRCA and OV Cancer
Data from above analyses identified multiple cancer types enriched with BRCAness features. We ranked the 33 cancer types based on the sum of BRCAneass features in the six features of somatic pathogenic variation, germline pathogenic variation, homozygous deletion, expression, and clinical prognosis. By using the sum in both BRCA cancer and OV cancer values = 1 as the cut-off, we observed that the following 21 cancer types had BRCAness features higher than BRCA cancer and OV cancer: UCEC, BLCA, PAAD, LGG, SARC, LUAD, KICH, UCS, ACC, COAD, LIHC, ESCA, HNSC, READ, STAD, SKCM, LUSC, MESO, KIRC, KIRP, and PRAD (Table 1). Of the 21 cancer types, UCEC (uterine corpus endometrial carcinoma) was the highest as referred by BRCA cancer and OV cancer.
Table 1: The 21 candidate cancer types for PARPi test as referred by both BRCA cancer and OV cancer.
| Cancer | Rank by BRCA Cancer | Rank by OV Cancer | Sum | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Somatic | Germline | Homozygotic | Methylation | Expression | Prognosis | Somatic | Germline | Homozygotic | Methylation | Expression | Prognosis | ||
| UCEC | 5.9 | 4.7 | 0.7 | 1.0 | 7.0 | 3.0 | 11.0 | 17.2 | 0.5 | 1.0 | 7.0 | 3.0 | 62.1 |
| BLCA | 3.8 | 2.0 | 1.3 | 1.0 | 10.5 | 1.0 | 4.8 | 14.6 | 1.0 | 1.0 | 10.5 | 1.0 | 52.6 |
| PAAD | 1.5 | 1.9 | 0.2 | 1.0 | 5.5 | 9.0 | 4.5 | 6.1 | 0.1 | 1.0 | 5.5 | 9.0 | 45.3 |
| LGG | 1.5 | 1.9 | 0.7 | 1.0 | – | 12.0 | 4.6 | 5.7 | 0.5 | 1.1 | – | 12.0 | 41.0 |
| SARC | 1.1 | 1.2 | 1.8 | 1.0 | 8.5 | 4.5 | 2.9 | 4.0 | 1.4 | 1.0 | 8.5 | 4.5 | 40.5 |
| LUAD | 1.8 | 1.5 | 1.0 | 1.0 | 0.0 | 7.0 | 3.5 | 6.9 | 0.8 | 1.0 | 0.0 | 7.0 | 31.6 |
| KICH | 1.2 | 1.2 | 0.1 | 1.0 | 3.0 | 6.5 | 2.9 | 4.2 | 0.1 | 1.0 | 3.0 | 6.5 | 30.5 |
| UCS | 3.2 | 4.0 | 0.7 | 0.9 | – | 0.0 | 9.6 | 10.1 | 0.5 | 0.9 | – | 0.0 | 30.0 |
| ACC | 0.4 | 0.7 | 0.8 | 1.0 | – | 10.0 | 1.6 | 1.4 | 0.6 | 1.0 | – | 10.0 | 27.4 |
| COAD | 2.9 | 2.2 | 0.7 | 1.0 | 0.0 | 1.0 | 5.3 | 11.0 | 0.6 | 1.0 | 0.0 | 1.0 | 26.8 |
| LIHC | 1.1 | 0.8 | 0.7 | 1.0 | 0.5 | 7.0 | 1.8 | 4.4 | 0.6 | 1.0 | 0.5 | 7.0 | 26.4 |
| ESCA | 2.0 | 3.3 | 1.0 | 1.0 | 1.0 | 0.0 | 7.7 | 7.7 | 0.8 | 1.0 | 1.0 | 0.0 | 26.4 |
| HNSC | 2.5 | 2.4 | 0.7 | 1.0 | 0.5 | 1.0 | 5.8 | 9.5 | 0.5 | 1.0 | 0.5 | 1.0 | 26.4 |
| READ | 2.3 | 2.4 | 0.5 | 1.0 | 2.0 | 0.0 | 5.8 | 8.5 | 0.4 | 1.0 | 2.0 | 0.0 | 25.9 |
| STAD | 2.8 | 2.6 | 1.0 | 1.0 | 0.0 | 0.0 | 6.0 | 10.5 | 0.8 | 1.0 | 0.0 | 0.0 | 25.6 |
| SKCM | 2.5 | 0.8 | 1.1 | 1.0 | – | 3.5 | 1.9 | 9.3 | 0.9 | 1.0 | – | 3.5 | 25.5 |
| LUSC | 2.7 | 2.6 | 1.1 | 1.0 | 0.0 | 0.0 | 6.3 | 9.9 | 0.8 | 1.0 | 0.0 | 0.0 | 25.3 |
| MESO | 0.7 | 0.8 | 1.2 | 1.0 | – | 7.0 | 1.9 | 2.4 | 0.9 | 1.0 | – | 7.0 | 23.9 |
| KIRC | 0.5 | 0.3 | 1.0 | 1.0 | 0.5 | 7.0 | 0.8 | 1.8 | 0.8 | 1.0 | 0.5 | 7.0 | 22.3 |
| KIRP | 0.4 | 0.1 | 0.5 | 1.0 | 0.0 | 7.5 | 0.3 | 1.5 | 0.4 | 1.1 | 0.0 | 7.5 | 20.3 |
| PRAD | 0.5 | 0.5 | 3.1 | 1.0 | 1.0 | 3.0 | 1.1 | 2.0 | 2.4 | 1.0 | 1.0 | 3.0 | 19.6 |
| BRCA&OV | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 12.0 |
4. Discussion
By characterizing pathogenic variants, somatic variants, homozygotic deletion, promoter methylation, gene expression, and clinical prognosis, our study portraits a comprehensive view for BRCAness landscape in cancer and reveals that BRCAness is widely present in different cancer types. We observed that genetic variation was widely presence in BRCAness genes in multiple cancer types, homozygotic variation was rare event in BRCAness genes as in BRCA, promoter methylation was common in BRCAness genes and caused alterative expression, the defects in BRCAness genes were strong prognostics markers as BRCA. By referring to the sum of BRCAness features higher than BRCA defected breast and ovarian cancer, we identified 21 BRCAness cancer types as the candidate targets for PARPi trial to further determine the efficacy of PARPi therapy in each cancer type.
By targeting multiple oncogenic components, synthetic lethal has shown promising potential as best exemplified by using PARPi to treat BRCA1/2 mutated breast and ovarian cancer. In our current study, we analyzed the potential of using PARPi therapy to treat other cancer types with BRCAness features. Through analyzing multiple features in 33 cancer types, our study provided the following evidence showing high similarity between BRCAness and BRCA1/2 mutation in multiple cancer types: (1) Genetic variation was widely present in BRCAness genes in multiple cancer types as represented by UCEC, BLCA, LUSC, HNSC, STAD, and COAD [ref. 44]; (2) homozygotic variation was a rare event in BRCAness genes as in BRCA1/2 mutation. Similar to the embryonic lethal effects in BRCA1/2, homozygous variation in BRCAness genes was not common across different cancer types although homozygous deletions in some BRCAness genes (FANCD2, PTEN, TP53, BRCA2) could be present [ref. 45]; (3) promoter methylation was common in many BRCAness genes and caused alterative expression [ref. 46]. Similar to the promoter methylation occurred in BRCA1/2, promoter methylation was present in nearly half of the BRCAness genes, and their expression were silenced in many cancer types in cancer type-specific manner; (4) the defects in BRCAness genes were strong prognostics markers as BRCA1/2 mutation, as shown in the BRCAness genes of CHEK2, ATM, RAD51D, EMSY, PALB2, BRIP1, ERCC1, RAD50, ATR, RAD51C in ovarian cancer [ref. 47].
Our study reveals that BRCAness defects are commonly present in multiple cancer types as BRCA1/2 defects in breast and ovarian cancer. Therefore, it opens a possibility to further test the potential of expanding PARPi therapy from breast and ovarian cancer to more cancer types with BRCAness features.
References
- P.J. O’Donovan, D.M. Livingston. BRCA1 and BRCA2: Breast/ovarian cancer susceptibility gene products and participants in DNA double-strand break repair. Carcinogenesis, 2010. [DOI | PubMed]
- X. Li, W.D. Heyer. Homologous recombination in DNA repair and DNA damage tolerance. Cell Res., 2008. [DOI | PubMed]
- C.J. Lord, A. Ashworth. The DNA damage response and cancer therapy. Nature, 2012. [DOI | PubMed]
- A. Tutt, D. Bertwistle, J. Valentine, A. Gabriel, S. Swift, G. Ross, C. Griffin, J. Thacker, A. Ashworth. Mutation in Brca2 stimulates error-prone homology-directed repair of DNA double-strand breaks occurring between repeated sequences. EMBO J., 2001. [DOI | PubMed]
- F. Xia, D.G. Taghian, J.S. DeFrank, Z.C. Zeng, H. Willers, G. Iliakis, S.N. Powell. Deficiency of human BRCA2 leads to impaired homologous recombination but maintains normal nonhomologous end joining. Proc. Natl. Acad. Sci. USA, 2001. [DOI | PubMed]
- M.E. Moynahan, A.J. Pierce, M. Jasin. BRCA2 is required for homology-directed repair of chromosomal breaks. Mol. Cell, 2001. [DOI | PubMed]
- K. Moore, N. Colombo, G. Scambia, B.G. Kim, A. Oaknin, M. Friedlander, A. Lisyanskaya, A. Floquet, A. Leary, G.S. Sonke. Maintenance Olaparib in Patients with Newly Diagnosed Advanced Ovarian Cancer. N. Engl. J. Med., 2018. [DOI | PubMed]
- J.K. Litton, H.S. Rugo, J. Ettl, S.A. Hurvitz, A. Goncalves, K.H. Lee, L. Fehrenbacher, R. Yerushalmi, L.A. Mina, M. Martin. Talazoparib in Patients with Advanced Breast Cancer and a Germline BRCA Mutation. N. Engl. J. Med., 2018. [DOI | PubMed]
- M. Robson, S.A. Im, E. Senkus, B. Xu, S.M. Domchek, N. Masuda, S. Delaloge, W. Li, N. Tung, A. Armstrong. Olaparib for Metastatic Breast Cancer in Patients with a Germline BRCA Mutation. N. Engl. J. Med., 2017. [DOI | PubMed]
- A. Tutt, M. Robson, J.E. Garber, S.M. Domchek, M.W. Audeh, J.N. Weitzel, M. Friedlander, B. Arun, N. Loman, R.K. Schmutzler. Oral poly(ADP-ribose) polymerase inhibitor olaparib in patients with BRCA1 or BRCA2 mutations and advanced breast cancer: A proof-of-concept trial. Lancet, 2010. [DOI | PubMed]
- P.C. Fong, D.S. Boss, T.A. Yap, A. Tutt, P. Wu, M. Mergui-Roelvink, P. Mortimer, H. Swaisland, A. Lau, M.J. O’Connor. Inhibition of poly(ADP-ribose) polymerase in tumors from BRCA mutation carriers. N. Engl. J. Med., 2009. [DOI | PubMed]
- M.R. Stratton, N. Rahman. The emerging landscape of breast cancer susceptibility. Nat. Genet., 2008. [DOI | PubMed]
- E.M. John, A. Miron, G. Gong, A.I. Phipps, A. Felberg, F.P. Li, D.W. West, A.S. Whittemore. Prevalence of pathogenic BRCA1 mutation carriers in 5 US racial/ethnic groups. JAMA, 2007. [DOI | PubMed]
- K.E. Malone, J.R. Daling, D.R. Doody, L. Hsu, L. Bernstein, R.J. Coates, P.A. Marchbanks, M.S. Simon, J.A. McDonald, S.A. Norman. Prevalence and predictors of BRCA1 and BRCA2 mutations in a population-based study of breast cancer in white and black American women ages 35 to 64 years. Cancer Res., 2006. [DOI | PubMed]
- K. Schlacher, N. Christ, N. Siaud, A. Egashira, H. Wu, M. Jasin. Double-strand break repair-independent role for BRCA2 in blocking stalled replication fork degradation by MRE11. Cell, 2011. [DOI | PubMed]
- N. Turner, A. Tutt, A. Ashworth. Hallmarks of ‘BRCAness’ in sporadic cancers. Nat. Rev. Cancer, 2004. [DOI | PubMed]
- H. Davies, D. Glodzik, S. Morganella, L.R. Yates, J. Staaf, X. Zou, M. Ramakrishna, S. Martin, S. Boyault, A.M. Sieuwerts. HRDetect is a predictor of BRCA1 and BRCA2 deficiency based on mutational signatures. Nat. Med., 2017. [DOI | PubMed]
- N.M. Tung, M.E. Robson, S. Ventz, C.A. Santa-Maria, R. Nanda, P.K. Marcom, P.D. Shah, T.J. Ballinger, E.S. Yang, S. Vinayak. TBCRC 048: Phase II Study of Olaparib for Metastatic Breast Cancer and Mutations in Homologous Recombination-Related Genes. J. Clin. Oncol., 2020. [DOI | PubMed]
- E.N. Imyanitov. Ovarian cancer genome. Methods Mol. Biol., 2013. [DOI | PubMed]
- S. Nik-Zainal, L.B. Alexandrov, D.C. Wedge, P. Van Loo, C.D. Greenman, K. Raine, D. Jones, J. Hinton, J. Marshall, L.A. Stebbings. Mutational processes molding the genomes of 21 breast cancers. Cell, 2012. [DOI | PubMed]
- P. Polak, J. Kim, L.Z. Braunstein, R. Karlic, N.J. Haradhavala, G. Tiao, D. Rosebrock, D. Livitz, K. Kubler, K.W. Mouw. A mutational signature reveals alterations underlying deficient homologous recombination repair in breast cancer. Nat. Genet., 2017. [DOI | PubMed]
- L.B. Alexandrov, S. Nik-Zainal, D.C. Wedge, S.A. Aparicio, S. Behjati, A.V. Biankin, G.R. Bignell, N. Bolli, A. Borg, A.L. Borresen-Dale. Signatures of mutational processes in human cancer. Nature, 2013. [DOI | PubMed]
- Comprehensive molecular portraits of human breast tumours. Nature, 2012. [DOI | PubMed]
- E. Lee, R. McKean-Cowdin, H. Ma, D.V. Spicer, D. Van Den Berg, L. Bernstein, G. Ursin. Characteristics of triple-negative breast cancer in patients with a BRCA1 mutation: Results from a population-based study of young women. J. Clin. Oncol., 2011. [DOI | PubMed]
- W.D. Foulkes, J.S. Brunet, I.M. Stefansson, O. Straume, P.O. Chappuis, L.R. Begin, N. Hamel, J.R. Goffin, N. Wong, M. Trudel. The prognostic implication of the basal-like (cyclin E high/p27 low/p53+/glomeruloid-microvascular-proliferation+) phenotype of BRCA1-related breast cancer. Cancer Res., 2004. [DOI | PubMed]
- W.D. Foulkes, I.M. Stefansson, P.O. Chappuis, L.R. Begin, J.R. Goffin, N. Wong, M. Trudel, L.A. Akslen. Germline BRCA1 mutations and a basal epithelial phenotype in breast cancer. J. Natl. Cancer Inst., 2003. [DOI | PubMed]
- L. Liu, W. Zhou, C.T. Cheng, X. Ren, G. Somlo, M.Y. Fong, A.R. Chin, H. Li, Y. Yu, Y. Xu. TGFbeta induces “BRCAness” and sensitivity to PARP inhibition in breast cancer by regulating DNA-repair genes. Mol. Cancer Res., 2014. [DOI | PubMed]
- W.R. Bodily, B.H. Shirts, T. Walsh, S. Gulsuner, M.C. King, A. Parker, M. Roosan, S.R. Piccolo. Effects of germline and somatic events in candidate BRCA-like genes on breast-tumor signatures. PLoS ONE, 2020. [DOI | PubMed]
- D.C. Wedge, G. Gundem, T. Mitchell, D.J. Woodcock, I. Martincorena, M. Ghori, J. Zamora, A. Butler, H. Whitaker, Z. Kote-Jarai. Sequencing of prostate cancers identifies new cancer genes, routes of progression and drug targets. Nat. Genet., 2018. [DOI | PubMed]
- R. Yaeger, W.K. Chatila, M.D. Lipsyc, J.F. Hechtman, A. Cercek, F. Sanchez-Vega, G. Jayakumaran, S. Middha, A. Zehir, M.T.A. Donoghue. Clinical Sequencing Defines the Genomic Landscape of Metastatic Colorectal Cancer. Cancer Cell, 2018. [DOI | PubMed]
- B.J. Raphael, R.H. Hruban, A.J. Aguirre, R.A. Moffitt, J.J. Yeh, C. Stewart, A.G. Robertson, A.D. Cherniack, M. Gupta, G. Gad Getz. Integrated Genomic Characterization of Pancreatic Ductal Adenocarcinoma. Cancer Cell, 2017. [DOI | PubMed]
- H.M. Singh, P. Bailey, D. Hubschmann, A.K. Berger, J.P. Neoptolemos, D. Jager, J. Siveke, C. Springfeld. Poly(ADP-ribose) polymerase inhibition in pancreatic cancer. Genes Chromosom. Cancer, 2021. [DOI | PubMed]
- J. de Bono, J. Mateo, K. Fizazi, F. Saad, N. Shore, S. Sandhu, K.N. Chi, O. Sartor, N. Agarwal, D. Olmos. Olaparib for Metastatic Castration-Resistant Prostate Cancer. N. Engl. J. Med., 2020. [DOI | PubMed]
- W. Abida, D. Campbell, A. Patnaik, J.D. Shapiro, B. Sautois, N.J. Vogelzang, E.G. Voog, A.H. Bryce, R. McDermott, F. Ricci. Non-BRCA DNA Damage Repair Gene Alterations and Response to the PARP Inhibitor Rucaparib in Metastatic Castration-Resistant Prostate Cancer: Analysis From the Phase II TRITON2 Study. Clin. Cancer Res., 2020. [DOI | PubMed]
- J. Mateo, N. Porta, D. Bianchini, U. McGovern, T. Elliott, R. Jones, I. Syndikus, C. Ralph, S. Jain, M. Varughese. Olaparib in patients with metastatic castration-resistant prostate cancer with DNA repair gene aberrations (TOPARP-B): A multicentre, open-label, randomised, phase 2 trial. Lancet Oncol., 2020. [DOI | PubMed]
- A.K. Byrum, A. Vindigni, N. Mosammaparast. Defining and Modulating ‘BRCAness’. Trends Cell Biol., 2019. [DOI | PubMed]
- C.J. Lord, A. Ashworth. BRCAness revisited. Nat. Rev. Cancer, 2016. [DOI | PubMed]
- N. Cancer Genome Atlas Research, J.N. Weinstein, E.A. Collisson, G.B. Mills, K.R. Shaw, B.A. Ozenberger, K. Ellrott, I. Shmulevich, C. Sander, J.M. Stuart. The Cancer Genome Atlas Pan-Cancer analysis project. Nat. Genet., 2013. [DOI | PubMed]
- C.H. Mermel, S.E. Schumacher, B. Hill, M.L. Meyerson, R. Beroukhim, G. Getz. GISTIC2.0 facilitates sensitive and confident localization of the targets of focal somatic copy-number alteration in human cancers. Genome Biol., 2011. [DOI | PubMed]
- Z. Gu, R. Eils, M. Schlesner. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics, 2016. [DOI | PubMed]
- X. Chen. False discovery rate control for multiple testing based on discrete p-values. Biom. J., 2020. [DOI | PubMed]
- D.W. Huang, B.T. Sherman, R.A. Lempicki. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc., 2009. [DOI | PubMed]
- Y. Hu, M. Guo. Synthetic lethality strategies: Beyond BRCA1/2 mutations in pancreatic cancer. Cancer Sci., 2020. [DOI | PubMed]
- T.A. Knijnenburg, L. Wang, M.T. Zimmermann, N. Chambwe, G.F. Gao, A.D. Cherniack, H. Fan, H. Shen, G.P. Way, C.S. Greene. Genomic and Molecular Landscape of DNA Damage Repair Deficiency across The Cancer Genome Atlas. Cell Rep., 2018. [DOI | PubMed]
- J. Cheng, J. Demeulemeester, D.C. Wedge, H.K.M. Vollan, J.J. Pitt, H.G. Russnes, B.P. Pandey, G. Nilsen, S. Nord, G.R. Bignell. Pan-cancer analysis of homozygous deletions in primary tumours uncovers rare tumour suppressors. Nat. Commun., 2017. [DOI | PubMed]
- S. Saghafinia, M. Mina, N. Riggi, D. Hanahan, G. Ciriello. Pan-Cancer Landscape of Aberrant DNA Methylation across Human Tumors. Cell Rep., 2018. [DOI | PubMed]
- M. Hjortkjaer, M. Malik Aagaard Jorgensen, M. Waldstrom, D. Ornskov, E. Sogaard-Andersen, A. Jakobsen, K. Dahl-Steffensen. The clinical importance of BRCAness in a population-based cohort of Danish epithelial ovarian cancer. Int. J. Gynecol. Cancer, 2019. [DOI | PubMed]