
Cannabis use in adolescence and young adulthood and its effects on brain structure and function: a scoping review
Abstract
Introduction:
Adolescence and young adulthood are simultaneously periods of significant brain development and the ages in which people often initiate cannabis use. This has led to significant interest in researching the effects that cannabis use in this period might have on the brains of users. This scoping review aims to summarize existing neuroimaging research on the effect of cannabis use in adolescence and/or young adulthood (ages 14-25) on brain structure, function, and metabolite concentrations.
Methods:
Following scoping review methodology, databases containing neuroimaging studies assessing the effects of cannabis use between the ages of 14 and 25 on brain structure, function, and metabolite concentrations were searched.
Results:
Our search yielded 3901 sources, of which 99 met inclusion criteria. The majority of included papers (84/99) found differences in the brain structure, function, and/or metabolite concentrations of adolescent/young adult cannabis users compared to non-using controls. Fewer studies explicitly assessed sex/gender differences, with 5 finding that sex/gender influenced the effect of cannabis use on the brain.
Conclusion:
Based on the findings of this review, there is considerable evidence to suggest that cannabis use in adolescence/young adulthood causes changes in the brains of users, however, the low quality of relevant research and scarcity of long term follow up studies, in addition to the heterogeneity of the existing research suggests that more work needs to be done to understand this relationship.
Article type: Review Article
Keywords: cannabis, adolescence, young adulthood, brain, neuroimaging
Affiliations: Department of Psychiatry, Dalhousie University, Halifax, NS, Canada; Research and Innovation, Nova Scotia Health Authority, Halifax, NS, Canada; Department of Diagnostic Radiology, Dalhousie University, Halifax, NS, Canada
License: Copyright © 2025 Nosko, Crocker and Tibbo. CC BY 4.0 This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
Article links: DOI: 10.3389/fpsyt.2025.1644105 | PubMed: 40969702 | PMC: PMC12440871
Relevance: Moderate: mentioned 3+ times in text
Full text: PDF (952 KB)
Introduction
Cannabis use is common in adolescence and young adulthood (AYA), a period which ranges from approximately 10 years old to the mid-twenties (ref. 1). School-based studies have reported, for example, that approximately 1 in 5 students used cannabis in the past year, and among these students, 1 in 10 endorsed daily cannabis use (ref. 2). Of concern is that the rates of cannabis use among youth appear to be increasing over time (ref. 3). Within a 2023 survey of Canadians’ cannabis use, it was found that 43% of people between 16–19 years old and 48% of people between 20–24 years old reported past year cannabis use, which was almost twice the amount of those over the age of 25 (ref. 4). High rates of cannabis use in AYA are also seen internationally. A 2022 survey of 12th grade students in the United States found that 30.7% of respondents endorsed past-year cannabis use (ref. 5). Similarly, a European survey reported a past-year cannabis use prevalence of 18.6% among individuals aged 15 to 24 (ref. 6). The use of cannabis during AYA is of particular interest as this time frame is known to be significant for brain maturational processes underlying important adult cognitive functions (ref. 7), such as cognitive control (ref. 8), working memory (ref. 9), risk-taking (ref. 10), and reward (ref. 11). Past research on the effects of cannabis use during this life-stage has associated it with a number of possible negative outcomes, such as a variety of cognitive deficits (ref. 12), an increased likelihood of developing an anxiety disorder (ref. 13) or a depressive disorder (ref. 14), engaging in problematic substance use (ref. 15), and increased suicidality (ref. 14, ref. 16). Cannabis use during AYA has also been associated with a greater likelihood of, and earlier onset of psychotic disorders (ref. 17), as well as worsened clinical outcomes for those young adults with current psychotic disorders (ref. 18). To understand the effect that cannabis use in AYA has on the brain, leading to changes in mental health and cognition, neuroimaging research has been applied.
Despite there being no consensus on the long-term effects of cannabis on the brain in AYA, there is a mixed body of research which suggests that such effects might exist. For example, previous research has reported that, compared to non-users, AYA cannabis users have abnormalities in brain morphology (ref. 19), resting-state brain activity (ref. 20), task-based brain activity (ref. 21), and metabolite concentrations (ref. 22). While fewer, some research has found no effect of AYA cannabis use on various brain measures (ref. 23). This body of potentially contradictory evidence, when examined more closely, has clearly differing definitions of “cannabis user,” small sample sizes, flawed study designs, and heterogenous imaging protocols severely limiting the usefulness of these findings. This makes it exceedingly difficult to meaningfully gauge the overall tenor of the results being published and as such, literature reviews are positioned to distill and synthesize these bodies of work to better inform this research area of interest.
Two recent and relevant reviews on AYA cannabis use and brain effects (ref. 24, ref. 25) have been published. One of the reviews (ref. 24) was more a narrative review, with some lack of clarity in its methodology [and adherence to established guidelines; see (ref. 26)]. The next most recent publication (ref. 25), a systematic review (n = 90 studies), included sources derived from a search conducted in 2019, thus a more recent review which includes publications completed over the last 5 years, and with consideration of brain metabolite concentrations and more refined imaging protocols, is warranted.
To summarize existing knowledge regarding the effect that AYA cannabis use has or does not have on the structure, function, and metabolite concentrations of the user’s brain, we conducted a scoping review. Scoping reviews are designed to provide a broad overview of what is- and remains to be known in a research field, as opposed to a systematic review, which focuses on answering a very specific question (ref. 27). A scoping review methodology was also chosen for this reason and because they are most appropriate for situations in which the chosen literature is highly heterogeneous and/or in situations where resources are limited (ref. 28), both of which are true for the current review.
Method
This scoping review was conducted in accordance with the Joanna Briggs Institute (JBI) methodology for scoping reviews (ref. 29). The protocol for the current review was preregistered on Open Science Framework on May 23, 2024 (ref. 30, ref. 31).
Inclusion criteria
This review included peer-reviewed studies published in English, theses/dissertations, and preprints using brain imaging and comparing cannabis users to non-users between the ages of 14 and 25 years old. This range was selected based on the WHO’s definition of AYA [10–24 years; (ref. 1)], the age ranges commonly used in research on AYA cannabis use, and the age by which puberty has typically started (ref. 32), indicating adolescence. Studies examining participants older than this age range were included if it was confirmed that participants regularly used cannabis between our age-range of interest. In addition, cannabis must have been the participants’ primary substance and its use by the participants must have been quantified. Participants were allowed to have preexisting psychiatric conditions (except substance use disorders related to a substance other than cannabis). Sex and gender effects were collected for this review when possible.
Exclusion criteria
This review did not include papers which assessed the acute effects of cannabinoid administration on the brain or animal studies.
Types of sources
This scoping review considered both experimental and quasi-experimental study designs including randomized controlled trials, non-randomized controlled trials, before and after observational studies and interrupted time-series studies. In addition, analytical observational studies including prospective and retrospective cohort studies, case-control studies and analytical cross-sectional studies were considered for inclusion. Also considered were descriptive observational study designs. Systematic reviews that meet the inclusion criteria were included with any relevant sources assessed and extracted.
Search strategy
A three-step search strategy was utilized in this review, with no limit on date of publication. First, an initial limited search of Ovid MEDLINE (NLM, Wolters Kluwer), CINAHL (EBSCO), PsycINFO (American Psychological Association), and Embase (Elsevier) was undertaken to identify relevant articles on the topic. The key words contained in the titles and abstracts of relevant articles, and the subheadings used to describe the articles were then used to develop a full search strategy for each database outlined in Appendix I. Using these terms, searches were conducted in all the listed databases as well as Google Scholar and MedRxiv.
Study selection
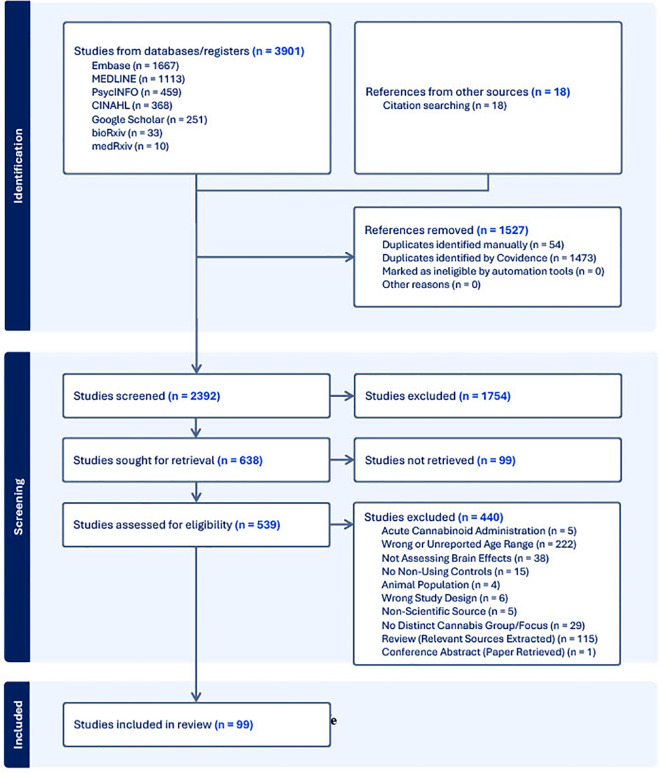
Following the search of each database, all identified citations were collected and uploaded into Covidence (ref. 33) where duplicates were removed. Following a pilot test, titles and abstracts were screened by two independent reviewers (L.N., C.H.L.). Potentially relevant sources were then retrieved in full and subsequently assessed in detail against the inclusion criteria. The reference list of all included sources were screened for additional relevant studies. Reasons for exclusion of sources were recorded and reported in the Preferred Reporting Items for Systematic reviews and Meta-Analyses extension for Scoping Reviews [PRISMA-ScR; (ref. 34)] flow diagram (f1). Any disagreements that arose between the reviewers at each stage of the selection process was resolved either through consensus, or if that was not possible, then with input from an additional reviewer (C.E.C., P.G.T.).
Data extraction
Data were extracted by two independent reviewers using a data extraction tool developed by the study team and completed in Covidence. From each paper the following was extracted: authors, year of publication, study design, sample size, mean age and age range of sample (divided into cannabis users and non-users), and relevant findings. Any disagreements that arose between the reviewers were resolved through discussion, or with input from an additional reviewer.
Sex and gender considerations
As a secondary objective, we aimed to determine if and how sex and/or gender influences the effect of AYA cannabis use on the brain. The World Health Organization’s definitions of sex and gender were used, that is, sex being an individual’s sex assigned at birth (i.e., male or female), while gender referred to an individual’s identity and societal role (i.e., man or woman) (ref. 35)). Included papers were categorized as having assessed sex/gender differences or not, with relevant sex/gender data extracted where available. Many of the studies that stated they assessed sex/gender were not explicit on the constructs they were assessing, in these cases we categorized them based on the language used (i.e., sex or gender).
Results
Characteristics of included studies
Our search yielded 99 papers that met our inclusion criteria (See f1; a number of papers included multiple imaging modalities). Out of these, 46 utilized structural neuroimaging protocols and 56 functional neuroimaging protocols. Out of the structural imaging studies, 29 used structural magnetic resonance imaging (sMRI), 12 used diffusion tensor imaging (DTI), 4 used magnetic resonance spectroscopy (MRS), and 1 used another imaging modality (CT) (See T1). Of the functional imaging studies, 39 used task-based functional magnetic resonance imaging (fMRI), 10 used resting-state fMRI, 5 used electroencephalography (EEG), and 2 used positron emission tomography (PET) (see T2).
Table 1: Proportion of different structural imaging modalities utilized in studies included in the review.
| Structural Imaging Modality | n (%) |
|---|---|
| DTI | 12 (26.1) |
| sMRI | 29 (63.0) |
| MRS | 4 (8.7) |
| Other | 1 (2.2) |
| Total | 46 |
Table 2: Proportion of different functional imaging modalities utilized in studies included in the review.
| Functional Imaging Modality | n (%) |
|---|---|
| Task-Based fMRI | 39 (69.6) |
| Resting-State fMRI | 10 (17.9) |
| EEG | 5 (8.9) |
| PET | 2 (3.6) |
| Total | 56 |
Approximately 85% (84/99) of the included studies found at least one significant difference in various aspects of brain structure/function between individuals who used cannabis between the ages of 14 and 25 and those who did not. While all studies reported the gender and/or sex of the participants, only 17% (17/99) assessed sex/gender differences related to their reported outcome(s). Five (~5%) of these studies found a significant sex/gender difference. The aim of the remainder of this review is to discuss the results of the included papers (see T3–T10 for summaries of all included studies).
Table 3: Summary of DTI studies.
| Study | Study Design | Sample Size | Age of Users mean (SD), range | Age of non-users mean (SD), range | Changes associated with cannabis use* | Sex/Gender Differences* |
|---|---|---|---|---|---|---|
| (ref. 64) | Cross sectional study | 14 users, 14 non-users. | 19.3 (0.8), 18-21.2 | 18.5 (1.4), 17.3-21.5 | •Reduced FA in the left posterior internal capsule/thalamic radiation, left middle temporal gyrus, right posterior internal capsule, right superior temporal gyrus, and right arcuate.•Reduced axial diffusivity in motor tracts.•Greater radial diffusivity and trace values in bilateral arcuate. Reduced FA only in the right arcuate. | •Not assessed. |
| (ref. 23) | Cross sectional study | 25 users, 25 non-users. | 20.40 (1.94), 17-23 | 19.76 (1.90), 17-23 | •None reported. | •Not assessed. |
| (ref. 65) | Longitudinal study | 23 users, 23 non-users. | BL: 19.45 (0.66), FU: 21.31 (2.43) | BL: 19.19 (2.31),FU: 21.79 (0.82) | •Attenuated increase of FA in left superior longitudinal fasciculus, left superior frontal gyrus, the left corticospinal tract, and the right anterior thalamic radiation.•Attenuated decrease of RD in right central and posterior superior longitudinal fasciculus, posterior cingulum, and corticospinal tract. | •Not assessed. |
| (ref. 71) | Cross sectional study | 39 users, 28 non-users. | 21.5 (2.3), 18-25 | 21.4 (2.0), 18-25 | •None reported. | •Not assessed. |
| (ref. 69) | Cross sectional study | 26 users, 10 non-users | Early-onset: 20.9 (2.9), late-onset: 22.2 (2.3) | SCZ: 20.4 (2.3), non-SCZ: 21.1 (2.8) | •Greater FA and white matter density in the left posterior corpus callosum in early-onset users compared to SCZ. | •Not assessed. |
| (ref. 70) | Cross sectional study | 10 users, 10 non-users. | 21.1 (2.9), 18-27 | 23.0 (4.4), 17-30 | •Greater FA in the left anterior cingulate, left precentral gyrus, right medial frontal gyrus, left superior frontal gyrus, and right cingulate gyrus.•Greater apparent diffusion coefficient in left middle frontal gyrus and right posterior cingulate. | •Not assessed. |
| (ref. 74) | Cross sectional study | 15 users, 15 non-users | 25 (8.7) | 25.2 (8.4) | •Decreased FA in left frontal lobe.•Greater overall diffusivity in right genu. | •Not assessed. |
| (ref. 66) | Cross sectional study | 25 users, 18 non-users | 23.16 (5.87) | 23.11 (3.51) | •Decreased FA in the left internal capsule and bilateral genu of the corpus callosum. | •Not assessed. |
| (ref. 53) | Cross sectional study | 19 users, 22 non-users. | 20.58 (2.52), 18-25 | 21.59 (1.94), 18-25 | •Decreased length of uncinate fasciculus fiber bundles. | •Not assessed. |
| (ref. 67) | Longitudinal study | 158 | BL: 20, FU: 22 | BL: 20, FU: 20 | •Attenuated developmental FA increase in the cingulum. | •Not assessed. |
| (ref. 54) | Cross sectional study | 32 users, 43 non-users | 21.6 (2.2), 18-26 | 21 (2.7), 16-25 | •Decreased right uncinate fasciculus mean diffusivity. | •Greater cannabis use was associated with decreased FA in the forceps minor of males, while it was associated with increased FA in females. |
| (ref. 68) | Cross sectional study | 33 users, 34 non-users | 21.21, 18-25 | 21.12, 18-25 | •Greater mean diffusivity in the uncinate fasciculi and corpus callosum forceps minor.•Decreased FA in uncinate fasciculi. | •Not assessed. |
BL, baseline; FU, follow-up; FA, fractional anisotropy; RD, radial diffusivity; SCZ, patients with schizophrenia. *Changes included here were reported as statistically significant by the study’s author(s) (p <.05) unless otherwise stated.
Table 10: Summary of task-based fMRI studies.
| Study | Study Design | Sample Size | Age of Users mean (SD), range | Age of Non-Users mean (SD), range | Task Paradigm | Changes significantly associated with cannabis use* | Sex/Gender Differences* |
|---|---|---|---|---|---|---|---|
| (ref. 104) | Cross sectional study | 14 users, 14 non-users | 17.6 (1), 15-19 | 17.3 (1.3), 15-19 | Digital coin flip | •Increased activity in the claustrum, middle frontal gyri, and caudate when winning.•Increased activity in the claustrum, declive, right posterior and anterior cingulate, right middle frontal gyrus, and the left insula when losing. | •Not assessed. |
| (ref. 82) | Cross sectional study | 82 | 16.2 (1.2), 14-18 | 15.6 (1.37), subclinical use: 16.6 (1.34), AUDIT ≥ 4: 16.5 (1.17)14-18 | Affective Stroop task | •Greater activation in the precuneus, inferior parietal lobule, and the posterior cingulate cortex during incongruent trials. | •Not assessed. |
| (ref. 86) | Cross sectional study | 13 users, 15 non-users | 21.15 (1.9), 19-25 | 20.27 (2.3), 18-25 | Implicit association test | •Increased activity in the putamen, caudate, and right inferior frontal gyrus during compatible trials.•Decreased activity in the prefrontal cortex and right inferior frontal gyrus during incompatible trials. | •Not assessed. |
| (ref. 100) | Cross sectional study | 21 users, 21 non-users | 24.95 (3.56), 18-34 | 24.24 (4.11), 19-33 | Paired associates task | •Absence of activity change in the left posterior cingulate gyrus and right precuneus during repeated recall.•Alterations in activity trajectories across areas of the midbrain over blocks of recall. | •Not assessed. |
| (ref. 92) | Cross sectional study | 39 users, 150 non-users | 15.97 (1.06), 14-18 | 16.05 (1.18), alcohol users: 16.35 (1.11), alcohol and cannabis users: 16.31 (1.09)14-18 | Balloon analogue risk task | •None reported. | •Not assessed. |
| (ref. 106) | Longitudinal study | 32 users, 41 non-users | BL: 21.4 (2.3), FU: 21.9 (2.4) | BL: 22.2 (2.4), FU: 22.7 (2.4) | Iowa gambling task | •Increased activity in the right insula, left superior temporal gyrus, and orbitofrontal cortex during wins. | •Not assessed. |
| (ref. 117) | Cross sectional study | 53 users, 22 non-users | Frequent users: 21.3 (2.3), sporadic users: 22.1 (2.5),18-25 | 22.1 (2.4), 18-25 | Cue-reactivity task | •Increased activity in the ventral tegmental area, anterior cingulate cortex, dorsal/ventral striatum, orbitofrontal cortex, and medial frontal cortex when viewing cannabis-related images. | •Not assessed. |
| (ref. 103) | Longitudinal study | 26 users, 23 non-users | BL: 21 (2.3), 18-25, abstinent users: 22.4 (3.6), FU: 24.5 (2.6), abstinent users: 24.7 (2.9) | BL: 22.1 (2.5), 18-25, FU: 25.3 (2.5) | N-back task | •None reported. | •Not assessed. |
| (ref. 102) | Longitudinal study | 32 users, 41 non-users | BL: 21.4 (2.4), FU: 21.6 (2.4) | BL: 22 (2.3), FU: 22.5 (2.4) | N-back task | •None reported. | •Not assessed. |
| (ref. 89) | Cross sectional study | 28 users, 32 non-users | 19.3 (2), 14-23 | 18.9 (2.7), 14-23 | Flanker task | •Absence of increased activation in the ventromedial prefrontal cortex, precentral gyrus, dorsal/ventral prefrontal cortex, parietal lobe, pallidum, left paracentral lobule, occipital gyri, right putamen, thalamus, middle cingulate, and precuneus during incongruent trials. | •Not assessed. |
| (ref. 98) | Cross sectional study | 27 users, 33 non-users | 18.3 (0.5), 18-22 | 18.4 (0.6), 18-22 | Figural memory task | •Absence of activation of the hippocampus during encoding.•Absence of activation in the left inferior frontal gyrus during encoding.•Decreased activation of the parahippocampal gyri, left basal ganglia, left insula, cerebellum, and the right precentral gyrus during encoding.•Decreased activation of the hippocampus and left inferior frontal gyrus during correct responding. | •Activity in the left inferior frontal gyrus, left hippocampus, and the right inferior frontal gyrus was lower in males during responding, but unchanged in females. |
| (ref. 112) | Cross sectional study | 20 users, 22 non-users | 20.6 (2.5), 18-25 | 21.5 (1.9), 18-25 | Cyberball | •Absence of exclusion-evoked activity in the insula. | •Not assessed. |
| (ref. 52) | Cross sectional study | 20 users, 20 non-users | 21.4 (2.0), 18-25 | 20.4 (1.7), 18-25 | Social influence task | •More diffuse activation of the striatum during the social influence condition.•Increased activity of the nucleus accumbens when accepting the group’s choice. | •Not assessed. |
| (ref. 113) | Cross sectional study | 20 users, 23 non-users | 20.6 (2.5), 18-25 | 21.6 (1.9), 18-25 | Social influence task | •Increased activity in the caudate during the social influence condition.•Greater inferior frontal gyrus activation when rejecting the group’s choice. | •Not assessed. |
| (ref. 90) | Cross sectional study | 23 users, 16 non-users | 22.43 (5.29) | 22.75 (2.82) | Multi-source interference task | •Smaller region of activation within the anterior cingulate cortex in response to incongruent stimuli. | •Not assessed. |
| (ref. 85) | Cross sectional study | 21 users, 21 non-users | 36.5 (8.8) | 31 (11.7) | Multi-source interference task | •Greater functional connectivity between the anterior insular cortex, occipitoparietal cortex, dorsal anterior cingulate, and the lateral prefrontal cortex with increasing task difficulty. | •Not assessed. |
| (ref. 21) | Cross sectional study | 10 users, 14 non-users | 19-21 | 19-21 | Counting Stroop task | •Greater activity in the right postcentral gyrus, right rolandic operculum, and the right cerebellar tonsil, right supplementary motor area, cingulate gyrus, and left postcentral gyrus during incongruent trials. | •Not assessed. |
| (ref. 114) | Longitudinal study | 20 users, 20 non-users | 19.84 (1.45), 17-22 | 20.51 (1.26), 17-22 | Emotion-arousal word task | •Decreased activity in the right middle and superior temporal gyri, amygdala, right calcarine fissure + lingual gyri and cuneus, right superior temporal gyrus and insula, and the right middle frontal and dorsolateral superior frontal gyri during the presentation of negative words.•Increased activity in the right dorsolateral superior frontal gyrus and decreased activity in the right inferior parietal lobe and amygdala during the presentation of positive words. | •Not assessed. |
| (ref. 101) | Cross sectional study | 135 | 16.5 (0.13), 14-18 | 16.5 (0.14), Alcohol users: 16.9 (0.16),14-18 | Passive avoidance task | •Delayed increase in functional connectivity between frontoparietal and striatal regions as learning took place. | •Not assessed. |
| (ref. 110) | Cross sectional study | 132 | 15.79 (1.4), 14-18 | 15.76 (1.2), tobacco users: 16.29 (1.2), cannabis and tobacco users: 15.76 (1.15), alcohol users: 16.00 (1.23), cannabis, tobacco, and alcohol users: 15.94 (0.97), 14-18 | Monetary incentive delay task | •None reported. | •Not assessed. |
| (ref. 91) | Cross sectional study | 36 users, 33 non-users | Median: 21, 18-25 | Median: 21, 18-25 | N-back flanker task | •Absence of greater superior temporal gyrus activity during progressively harder trials.•Reduced activity in the postcentral gyrus, superior parietal lobe, insula, operculum, supramarginal gyrus, and thalamus during trials with cannabis-related flankers. | •Not assessed. |
| (ref. 109) | Longitudinal study | 47 users, 111 non-users | Stable-high use: 20.09 (0.35), escalating use: 20.07 (0.24) | 20.10 (0.26) | “card-guessing game” | •Inversely correlated activation patterns in the nucleus accumbens and medial prefrontal cortex during reward processing in those with increasing use during adolescence. Opposite of what was seen in consistently high users and non/low users. | •Not assessed. |
| (ref. 118) | Cross sectional study | 24 users, 24 non-users | 18.2 (0.7), 16-19 | 18 (1.9), 16-19 | Finger-tapping task | •Decreased activity in the cerebellum and right cingulate gyrus. | •Not assessed. |
| (ref. 111) | Longitudinal study | 164 users, 154 non-users | 19-22 | 19-22 | Monetary incentive delay task | •None reported. | •Not assessed. |
| (ref. 87) | Cross sectional study | 34 users, 32 non-users | female: 21.4 (20), 19-25, male: 21.7 (2.0), 18-25 | female: 21.2 (2.4), 18-25, male: 20.9 (2.7), 16-25 | Go/no-go task w/emotional stimuli | •Decreased activity in the rostral anterior cingulate cortex when inhibiting responses to fearful faces. | •None found. |
| (ref. 108) | Longitudinal study | 108 | BL: 20.1 (1.4), FU1: 22.1 (1.5), FU2: 23.8 (1.7) | BL: 20.1 (1.4), FU1: 22.1 (1.5), FU2: 23.8 (1.7) | Modified monetary incentive delay task | •Reduced nucleus accumbens activity in response to the expectation of reward. | •Not assessed. |
| (ref. 107) | Cross sectional study | 18 users, 18 non-users | 16.11 (0.41) | 16.5 (0.23) | Monetary incentive delay task | •Increased connectivity between the orbitofrontal cortex, amygdala, nucleus accumbens, lateral/medial prefrontal cortex, hippocampus, and temporal cortex while expecting reward. | •Not assessed. |
| (ref. 115) | Cross sectional study | 37 users, 40 non-users | 19.85 (1.63), 16-23 | 19.85 (1.63), 16-23 | Emotion regulation task | •Greater left superior temporal gyrus activity in response to positive trials and reduced activity in response to negative trials. | •Not assessed. |
| (ref. 97) | Cross sectional study | 17 users, 17 non-users | 18.06 (0.75) | 17.9 (1.12) | Spatial working memory task | •Increased activity in the parietal lobes and the right basal ganglia during the spatial working memory trials.•Increasing activity in the left superior temporal gyrus and decreasing activity in the right superior temporal gyrus with more accurate responses.•Increased activity in the left anterior cingulate cortex with more accurate responses.•Decreased activity in the pulvinar and right thalamus with more accurate responses. | •Not assessed. |
| (ref. 83) | Cross sectional study | 50 users, 34 non-users | 23.98 (6.95), 17-46 | 24.53 (6.57), 17-46 | Stroop color word test | •Increased activity in the left anterior cingulate cortex during incompatible trials. | •Not assessed. |
| (ref. 94) | Cross sectional study | 15 users, 17 non-users | 18.1 (0.7), 16-18 | 17.9 (1.0), 16-18 | Spatial working memory task | •Increased activity in the inferior cuneus and right superior parietal lobule during spatial working memory trials.•Reduced activity in the medial right superior cuneus and right dorsolateral prefrontal cortex during spatial working memory trials | •Not assessed. |
| (ref. 99) | Cross sectional study | 8 users, 64 non-users | 18.1 (0.9), 16-18 | 17.6 (0.8), binge drinkers: 18.1 (0.7), binge drinker and users: 18.0 (1.0), 16-18 | Paired associates task | •Absence of hippocampus activity while encoding.•Increased activity of regions in the frontal lobe while encoding.•Decreased activity in the lingual gyrus and cuneus while encoding. | •None found. |
| (ref. 105) | Cross sectional study | 63 users, 62 non-users | Adolescents: 17.22 (0.52), 16.31-17.98, adults: 27.81 (1.49), 26.27-30.02 | Adolescents: 17.15 (0.45), 16.27-18.04, adults: 27.34 (0.86), 26.10-29.56 | Monetary incentive delay task | •Increased activity in the right frontal pole and inferior parietal cortex during reward. | •Not assessed. |
| (ref. 95) | Cross sectional study | 10 users, 14 non-users | 20, 19-21 | 20, 19-21 | Visuospatial 2-back task | •Increased activity in the right superior temporal gyrus, right inferior frontal gyrus, and the left middle frontal gyrus during 2-back trials. | •Not assessed. |
| (ref. 93) | Cross sectional study | 10 users, 14 non-users | 20, 19-21 | 20, 19-21 | Go/no-go task | •None reported. | •Not assessed. |
| (ref. 116) | Longitudinal study | 578 | BL: 14.6 (0.42) FU: 19.4 (0.96) | BL: 14.4 (0.42) matched controls: 14.5 (0.37), FU: 18.9 (0.67) matched controls: 19.3 (0.65) | Face processing task | •Increased activity in the right amygdala on trials with angry faces.•Absence of longitudinal increases in right amygdala activity in response to angry faces. | •Not assessed. |
| (ref. 88) | Cross sectional study | 34 users, 32 non-users | Female: 21.4 (2.0), 19–25 male: 21.7 (2.0), 18-25 | Female: 21.2 (2.4), 18–25 male: 20.9 (2.7), 16-25 | Go/no-go task w/emotional faces | •Reduced rostral anterior cingulate cortex activity when having to inhibit responses to fearful faces. | •Increased connectivity between the cerebellum and the right rostral anterior cingulate cortex in males. |
| (ref. 84) | Cross sectional study | 16 users, 17 non-users | 18.1 (0.7), 16-18.9 | 17.9 (1), 16-28.9 | Go/no-go task | •Increased activity in the right superior middle frontal gyrus, posterior parietal cortex, right lingual gyrus, right middle frontal gyrus, anterior insula, left anterior middle gyrus, right anterior superior gyrus, superior frontal gyrus, and medial prefrontal cortex during no-go trials.•Increased activity in the medial precuneus, right inferior frontal gyrus, right superior parietal lobule, anterior insula, right superior frontal gyrus, right inferior parietal lobule during go-trials. | •Not assessed. |
| (ref. 96) | Prospective cohort study | 22 users, 63 non-users | 15.58 (0.35) | 15.65 (0.37) | Spatial working memory task | •Reduced activity in the cuneus. | •Not assessed. |
*Changes included here were statistically significant (p <.05) unless otherwise stated.
Table 4: Summary of EEG studies.
| Study | Study Design | Sample Size | Age of Users mean (SD), range | Age of non-users mean (SD), range | Changes associated with cannabis use* | Sex/Gender Differences* |
|---|---|---|---|---|---|---|
| (ref. 128) | Cross sectional study | 21 users, 20 non-users | 26.4, 18.5-52 | 24.7, 18.1-52.6 | •Reduced P50 difference scores.•Greater P50 ratios. | •Not assessed. |
| (ref. 130) | Cross sectional study | 18 users, 36 non-users | 39.1, 20.8-56 | MMN: 40.4, 21-52.6, P50: 31.2, 20.1-52.6 | •None reported. | •Not assessed. |
| (ref. 129) | Cross sectional study | 39 users, 42 non-users | 26.4, 18.3-54.2 | 27.2, 18.1-52.6 | •Decreased frequency (and duration in long-term users) mismatch negativity. | •Not assessed. |
| (ref. 131) | Cross sectional study | 37 users, 38 smokers, 39 non-users. | 21.7 (2.10), 18-25 | Smokers: 21.4 (2.5), non-users: 22.1 (2.1), 18-25 | •Decreased P3 ERP at the Cz, Fz, and FCz electrodes during a go/no-go task. | •Not assessed. |
| (ref. 132) | Cross sectional study | 20 users, 84 non-users | F: 20.1 (1.2), M: 20.5 (1.2), 18-21.92 | Controls: F: 19.9 (1.2), M: 20.0 (1.1)Alcohol users: F: 20.0 (1.2), M: 19.7 (1.2), 18-21.92 | •Decreased amplitude of the N340 wavelength at the C1, C2, Cz, FC1, FCz, and FC2 electrodes during the Rey Auditory Verbal Learning Task. | •None found. |
*Changes included here were reported as statistically significant by the study’s author(s) (p <.05) unless otherwise stated.
Table 5: Summary of MRS studies.
| Study | Study Design | Sample Size | Age of Users mean (SD), range | Age of non-users mean (SD), range | Changes associated with cannabis use* | Sex/Gender Differences* |
|---|---|---|---|---|---|---|
| (ref. 75) | Cross sectional study | 13 users, 25 users w/bipolar disorder (BP), 15 non-users, 14 non-users with bipolar disorder. | 19.2 (1.1), BP: 18.4 (2.1) | 18.4 (2.0), BP: 17.2 (3.0) | •Increased ventral lateral prefrontal cortex N-acetyl aspartate concentrations. | •Not assessed. |
| (ref. 79) | Cross sectional study | 22 users, 21 non-users | 25.05 (3.50), 18-34 | 24.24 (4.11), 19-33 | •Decreased myoinositol in the hippocampus. | •None found. |
| (ref. 77) | Cross sectional study | 27 users, 26 non-users | 19.5 (0.6), 18-21 | 19.3(3.1), 13-24 | •Increased myoinositol in the dorsal striatum.•Decreased dorsal striatum glutamate and glutamine. | •These effects were only observed in female participants. |
| (ref. 80) | Cross sectional study | 18 users, 21 non-users | 20.39 (0.99), 18-21 | 18.43 (2.57), 14-21 | •Decreased anterior cingulate cortex GABA. | •Not assessed. |
*Changes included here were reported as statistically significant by the study’s author(s) (p <.05) unless otherwise stated.
Table 6: Summary of studies utilizing other imaging methods.
| Study | Study Design | Imaging modality | Sample Size | Age of Users mean (SD), range | Age of non-users mean (SD), range | Changes associated with cannabis use* | Sex/Gender Differences* |
|---|---|---|---|---|---|---|---|
| (ref. 81) | Cross sectional study | Computed transaxial tomography | 12 users, 34 non-users | 24.1, 20-30 | 25.8, 20-30 | •None reported. | •Not assessed. |
*Changes included here were reported as statistically significant by the study’s author(s) (p <.05) unless otherwise stated.
Table 7: Summary of PET studies.
| Study | Study Design | Sample Size | Age of Users mean (SD), range | Age of non-users mean (SD), range | Changes significantly associated with cannabis use* | Sex/Gender Differences* |
|---|---|---|---|---|---|---|
| (ref. 133) | Cross sectional study | 41 users, 18 non-users. | 18.5 (0.6), 18-20 | 18.5 (0.6), 18-20 | •Decreased [11C]ABP688 binding potential in the insula, ventral striatum, and medial orbitofrontal cortex | •Not assessed. |
| (ref. 134) | Cross sectional study | 6 users, 6 non-users | 20 (1), 18-21 | 20 (1), 18-21 | •Decreased glucose metabolism in right orbitofrontal cortex, putamen, right cerebellum, and medial posterior parietal cortex. | •Not assessed. |
*Changes included here were reported as statistically significant by the study’s author(s) (p <.05) unless otherwise stated.
Table 8: Summary of resting-state fMRI studies.
| Study | Study Design | Sample Size | Age of Users mean (SD), range | Age of non-users mean (SD), range | Changes significantly associated with cannabis use* | Sex/Gender Differences* |
|---|---|---|---|---|---|---|
| (ref. 123) | Cross sectional study | 17 users, 18 non-users | 16.5 (0.2), 15-18 | 16.1 (0.4), 14-19 | •Greater connectivity between the left cerebellum and bilateral inferior parietal lobules. | •Not assessed. |
| (ref. 124) | Longitudinal study | 23 users, 21 non-users | BL: 19.4 (0.65),FU: 21.8 (0.81)18-21 | BL: 19.3 (1.18),FU: 21.5 (1.11)18-21 | •Absence of longitudinal growth in resting-state functional connectivity between the caudal anterior cingulate cortex and the right inferior parietal lobule and right precentral gyrus.•Diminished resting-state functional connectivity between the caudal anterior cingulate cortex and the right superior frontal gyrus and dorsal/rostral anterior cingulate cortex, and the bilateral medial dorsal and anterior thalamic nucleus. | •Not assessed. |
| (ref. 20) | Cross sectional study | 70 users, 70 non-users | 16–17 years or 26–29 years | 16–17 years or 26–29 years | •Heightened resting-state connectivity in the executive control network. | •Not assessed. |
| (ref. 127) | Longitudinal study | 23 users, 23 non-users | 17.7 (0.7), 15-18 | 17.5 (0.8), 15-18 | •Diminished cerebral blood flow in the left insula, bilateral medial frontal gyrus, left superior temporal gyrus, middle temporal gyrus, and the left supra-marginal gyrus.•Elevated cerebral blood flow in the right precuneus. | •Not assessed. |
| (ref. 119) | Cross sectional study | 38 users, 37 non-users. | 24.5 (1.3), ADHD: 24.7 (1.2), 21-25 | 24.4, ADHD: 25.3, 21-25 | •Increased intrinsic functional connectivity in the left fusiform gyrus and right superior temporal sulcus. | •Not assessed. |
| (ref. 121) | Cross sectional study | 43 users, 31 non-users | 18 (1.22), 14-20 | 17.2 (1.38), 14-20 | •Increased functional connectivity between the cingulate, right middle frontal gyrus, and the orbitofrontal cortex.•Greater functional connectivity between the orbitofrontal cortex and the left precentral gyrus and right superior frontal gyrus. | •Not assessed. |
| (ref. 120) | Cross sectional study | 21 users, 21 non-users | 23 (1.4), 20.8-25.6 | 22.9 (1.9), 20.5-26.8 | •Increased effective connectivity between the left and right amygdala, left amygdala and anterior cingulate cortex, and right insula and medial prefrontal cortex. | •Not assessed. |
| (ref. 122) | Cross sectional study | 17 users, 18 non-users | 16.5 (0.2), 15-18 | 16.1 (0.4), 14-19 | •Greater fractional amplitude of the low-frequency fluctuations (fALFF) activity in the right inferior frontal gyrus, right superior parietal gyrus, right inferior semilunar lobe of the cerebellum, right inferior temporal gyrus, and the right superior frontal gyrus.•Decreased interhemispheric connectivity in the superior frontal gyrus and cerebellar pyramis.•Greater interhemispheric connectivity in the supramarginal gyrus. | •Not assessed. |
| (ref. 126) | Cross sectional study | 74 | 19.8 (1.62), 16-23 | 19.8 (1.62), 16-23 | •Decreased functional connectivity between the default mode network and the prefrontal cortex.•Increased functional connectivity between the fusiform gyrus and the culmen and the default mode network.•Increased functional connectivity between the parahippocampal and temporal gyrus, caudate, and the default mode network. | •Not assessed. |
| (ref. 125) | Cross sectional study | 8 users, 23 non-users | 20.2 (2.19), 18-25 | 19.7 (1.42), 18-25, alcohol users: 19.54 (0.97), 18-25 | •Reduced functional connectivity between the occipital fusiform gyrus and the left orbitofrontal cortex.•Greater functional connectivity between the right cerebellum and hippocampus. | •Not assessed. |
*Changes included here were statistically significant (p <.05) unless otherwise stated.
Table 9: Summary of sMRI studies.
| Study | Study Design | Sample Size | Age of Users mean (SD), range | Age of non-users mean (SD), range | Changes significantly associated with cannabis use* | Sex/Gender Differences* |
|---|---|---|---|---|---|---|
| (ref. 36) | Longitudinal study | 799 | BL: 14.4 (0.4),FU: 19.0 (0.7) | BL: 14.4 (0.4),FU: 19.0 (0.7) | •Decreased prefrontal cortex cortical thickness.•Increased rate of prefrontal cortex cortical thinning. | •None found. |
| (ref. 19) | Longitudinal study | 2223 | BL: 14-19,FU: 19-22 | BL: 14-19,FU: 19-22 | •Decreased cortical thickness in dorsomedial prefrontal cortex.•Increased rate of prefrontal cortex cortical thinning.•Change in cortical thickness in left lateral temporal cortex and heteromodal and medial paralimbic cortices. | •None found. |
| (ref. 45) | Cross sectional study | 14 users, 14 non-users. | 19.3 (0.8), 18-20 | 18.5 (1.4), 18-20 | •Decreased size of hippocampus. | •Not assessed. |
| (ref. 46) | Cross sectional study | 30 users, 29 non-users | 21 (2.3), 18-30 | 22.4 (3.3), 18-30 | •Gene-dependent changes in whole hippocampus and hippocampal subregion sizes. | •Not assessed. |
| (ref. 37) | Cross sectional study | 24 users, 26 non-users | 65.4 (7.2), 57-75 | 67.7 (7.1), 57-75 | •Reduced cortical thickness of the whole hippocampus and and subregions. | •Not assessed. |
| (ref. 47) | Cross sectional study | 18 users, 18 non-users | 17.7 (0.94), 16-19 | 17.2 (0.82), 16-19 | •Lower volume of the right medial orbital prefrontal cortex. | •Not assessed. |
| (ref. 48) | Cross sectional study | 33 users, 42 non-users | 21.3 (2.4), 18-25 | 21.9 (2.4), 18-25 | •Increased volume of the cerebellar grey matter. | •None found. |
| (ref. 49) | Cross sectional study | 11 users, 13 non-users | 22 (2), 19-25 | 19-25 | •Decreased grey matter volume in the right anterior hippocampus. | •Not assessed. |
| (ref. 50) | Longitudinal study | 17 users, 17 non-users | 20.7 (2.2), 18-25 | 21.5 (2.67), 18-25 | •Greater longitudinal enlargement of- and total volumes of hippocampal subregions. | •None found. |
| (ref. 51) | Cross sectional study | 20 users, 20 non-users | 21.3 (1.9), 18-25 | 20.7 (1.9), 18-25 | •Increased grey matter density in the hypothalamus, left nucleus accumbens to subcallosal cortex, left amygdala, and sublenticular extended amygdala.•Absence of a correlation between nucleus accumbens grey matter density and morphological features.•Altered shape of left nucleus accumbens and right amygdala. | •Not assessed. |
| (ref. 52) | Cross sectional study | 20 users, 20 non-users | 21.4 (2.0), 18-25 | 20.4 (1.7), 18-25 | •Increased grey matter density in the left nucleus accumbens. | •Not assessed. |
| (ref. 62) | Longitudinal study | 20 users, 23 non-users | BL: 20.64 (2.23), 18-24, FU: 25.07 (2.64) | BL: 21.79 (2.6), 18-24, FU: 25.07 (2.64) | •None reported. | •Not assessed. |
| (ref. 53) | Cross sectional study | 19 users, 22 non-users. | 20.58 (2.52), 18-25 | 21.59 (1.94), 18-25 | •Decreased cortical thickness in the right amygdala, fusiform gyrus, and entorhinal cortex. | •Not assessed. |
| (ref. 38) | Cross sectional study | 55 users, 65 non-users | 23.6 (1.5), ADHD: 24.3 (1.3) | 23.4 (1.5), ADHD: 24.6 (1.4) | •Cortical thinning in right anterior/posterior cingulate cortex, left precentral gyrus, and bilateral superior frontal gyrus. | •Not assessed. |
| (ref. 39) | Cross sectional study | 18 users, 18 non-users | 17.8 (1.0), 16-19 | 17.3 (0.8), 16-19 | •Cortical thinning in the bilateral insula, bilateral superior frontal cortex, and the right caudal middle frontal cortex.•Cortical thickening in the right inferior and superior parietal cortex, bilateral lingual gyrus, paracentral sulcus, and the right superior temporal cortex. | •Not assessed. |
| (ref. 54) | Cross sectional study | 32 users, 43 non-users | 21.6 (2.2), 18-26 | 21 (2.7), 16-25 | •Increased volume of the right superior temporal lobe. | •Decreased volume of the left rostral anterior cingulate cortex was only observed in females. |
| (ref. 40) | Cross sectional study | 30 users, 44 non-users | 25.7 (5) | 25.8 (5.8) | •Reduced gyrification in left temporal and bilateral frontal lobes.•Absence of longitudinal reductions in frontal lobe cortical thickness and gyrification. | •Not assessed. |
| (ref. 55) | Cross sectional study | 35 users, 47 non-users | Female: 18.15 (0.86), 16-18, male: 17.92 (0.91), 16-18 | Female: 17.85 (0.73), 16-18, male: 17.65 (0.90), 16-18 | •Volume increase in the amygdala. | •This effect was only reported in females. No difference in males. |
| (ref. 61) | Cross sectional study | 16 users, 16 non-users | 18.7 (0.7), 16-18 | 18.9 (0.9), 16-18 | •None reported. | •Not assessed. |
| (ref. 63) | Cross sectional study | 16 users, 16 non-users | Females: 18.2 (0.6), 16-18, males: 18.1 (0.8), 16-18 | Females: 18.5 (0.5), 16-18, males: 17.7 (1.1), 16-18 | •None reported. | •Trend-level interaction between gender and cannabis use, such that male users had decreased prefrontal cortex volumes, while the opposite was observed in females. |
| (ref. 56) | Cross sectional study | 16 users, 16 non-users | 18.11 (0.74), 16-18 | 18.01 (0.97), 16-18 | •Higher volume of the inferior posterior vermis. | •None found. |
| (ref. 44) | Longitudinal study | 94 users, 87 non-users | Youngest cohort: 29.6 (1.23), oldest cohort: 36.2 (1.52) | Youngest cohort: 29.6 (1.23), oldest cohort: 36.2 (1.52) | •None reported. | •Not assessed. |
| (ref. 41) | Longitudinal study | 537 | BL: 14, FU: 19 | BL: 14, FU: 19 | •Increased rate of longitudinal cortical thinning. | •Not assessed. |
| (ref. 57) | Cross sectional study | 27 users, 32 non-users | 21.41 (2.21), 18-25 | 21.09 (2.32), 18-25 | •Decreased volume of the medial orbitofrontal cortex. | •None found. |
| (ref. 42) | Cross sectional study | 41 users, 41 non-users | 20.3 (2.1), 18-25, Major Depressive Disorder: 21.3 (2.4), 18-25 | 21.5 (2.0), 18-25, Major Depressive Disorder: 22.1 (2.5), 18-25 | •Cortical thinning in the right entorhinal cortex, middle temporal gyrus, medial orbitofrontal cortex, and superior frontal gyrus. | •Not assessed. |
| (ref. 43) | Cross sectional study | 147 users, 634 non-users | Occasional: 18.1 (2), 14-22, frequent: 18.5 (1.6), 15-22 | 17 (2.1), 14-22 | •Slight cortical thinning in the left frontal lobe.** | •Not assessed. |
| (ref. 58) | Cross sectional study | 33 users, 35 non-users | 21.21 (2.34), 18-25 | 21.14 (2.33), 18-25 | •Gyrification reductions in the frontal lobes and ventral-medial and medial prefrontal cortex. | •Not assessed. |
| (ref. 60) | Cross sectional study | 79 users, 79 non-users | adolescents: 16.65 (1.09), 14-18, adults: 27.4 (7.1), 18-53 | adolescents: 16.77 (0.95), 14-18, adults: 27.5 (6.8), 18-53 | •None reported. | •Not assessed. |
| (ref. 59) | Longitudinal study | 706 | BL: 14.41 (0.93), FU: 18.89 (0.66) | BL: 14.41 (0.93), FU: 18.89 (0.66) | •Attenuated longitudinal increase in the volume of the uncus. | •None found. |
*Changes included here were reported as statistically significant by the study’s author(s) (p <.05) unless otherwise stated. **Scott et al.’s (43) reported finding was significant but not reported as being reflective of cannabis-related damage.
Structural neuroimaging results
sMRI
Cortical thickness
We categorized sMRI findings based on the morphological measure examined, here being cortical thickness or volume. Of the included sMRI papers, 38% (10/26) examined cortical thickness with 80% (8/10) reporting decreased cortical thickness in cannabis using AYA compared to non-using controls (ref. 19, ref. 36–ref. 42). Cortical thinning was usually located within the frontal lobes (ref. 19, ref. 36, ref. 40, ref. 42), the hippocampus (ref. 37) and the cingulate (ref. 38). Two studies found no evidence of cortical thinning related to cannabis use (ref. 43, ref. 44).
Volume changes
Of the sMRI papers assessing brain volumes, 71% (15/21) found significant regional volume differences between users and non-users (ref. 45–ref. 59). The most common area of change reported was in the hippocampus, however, there was no consensus on change direction as both increases and decreases in volume were reported (ref. 45, ref. 46, ref. 49, ref. 50). AYA cannabis use was only associated with decreasing volumes in the prefrontal cortex (ref. 47, ref. 57, ref. 58) with the amygdala showing evidence of volume increase in two studies (ref. 51, ref. 55) but a decrease in another (ref. 53). Other brain areas reporting AYA cannabis related volume changes include the cerebellum [volume increase; (ref. 48, ref. 56)], the nucleus accumbens [volume increase; (ref. 52)], and the uncus [absence of longitudinal volume growth; (ref. 59)]. The remaining 29% of the included studies found no significant effect of AYA cannabis use on brain volumes (ref. 43, ref. 44, ref. 60–ref. 63).
Abstinence in sMRI
Medina et al. (2010) reported that cerebellar volume increases related to cannabis use remained after a month of abstinence, Ashtari et al. (2011) reported on cannabis-related changes in hippocampal volumes which persisted after an average of 6.7 months of abstinence, and Burggren et al. (2018) found changes in hippocampal cortical thickness persisted after a minimum of 22 years of abstinence from regular use (ref. 37, ref. 45, ref. 56). However, Medina et al. (2009) found no abnormalities in prefrontal cortex volumes in cannabis users who had been abstinent for 1 month (ref. 63).
Sex/gender differences in sMRI
Possible sex/gender effects have been reported by McQueeny et al. (2011), who found cannabis associated increases in amygdala volume only in women (ref. 55). Maple (2016) had a similar finding, observing that only women cannabis users showed a significant decrease in the volume of the left rostral anterior cingulate cortex (ref. 54). Medina et al. (2009) reported that cannabis use tended to be associated with increased prefrontal cortex volume in women with the opposite being found in men, however this gender effect was not statistically significant (ref. 63).
Summary of sMRI findings
Taken together, the weight of the current evidence seems to suggest that AYA cannabis use is associated with cortical thinning, particularly in the frontal lobes. Volume changes related to AYA cannabis use were inconsistent in their direction, being commonly associated with both volume increases and decreases in the hippocampus and volume decreases in the prefrontal cortex. Additionally, the weight of this small body of literature seems to suggest that structural brain changes related to cannabis use in young adults are potentially persistent. However, more research and longer periods of abstinence are needed to bolster the strength of this claim. Due to the relatively few studies which explicitly assessed sex/gender effects, research regarding sex/gender differences in the impact of AYA cannabis use on sMRI measures should be pursued.
DTI
Fractional anisotropy changes
Of the DTI papers included in this review, 9/12 assessed fractional anisotropy (FA) as the primary DTI variable, a broad measure of the integrity of neuronal pathways. Approximately 56% (5/9) of these papers found lower FA values across the brain (ref. 64–ref. 68), indicating a loss of neuronal integrity from possible neuron/myelin abnormalities, or in axon packing/coherence. Other researchers have reported increased FA in multiple regions of the brain (ref. 69, ref. 70)), hypothesized to indicate potential hyperconnectivity between brain regions. However, two studies have found no differences in FA between AYA users and non-users (ref. 23, ref. 71). Furthermore, FA has been criticized to be too broad of a measure, limiting the interpretation of these findings (ref. 72).
Other DTI metrics
DTI can generate other indices of neuronal integrity. However, within the included DTI studies, there was a great deal of heterogeneity regarding the measures used and the areas assessed. Ashtari et al. (2009) found decreased axial diffusivity in the motor pathways and increased radial diffusivity and apparent diffusion coefficient in the arcuate fasciculus (ref. 64). Becker et al. (2015) found changes in radial diffusion in multiple subcortical areas, reflecting potential cannabis-related changes to neuronal microstructure (ref. 65, ref. 73). Gruber et al. (2011) found greater overall diffusivity in the right genu, which also potentially reflects neuronal degradation (ref. 73, ref. 74). Both Maple (2016) and Shollenbarger et al. (2015b) reported that the uncinate fasciculus had decreased mean diffusivity in AYA cannabis users (ref. 54, ref. 68) and Levar et al. (2018) found that fiber bundles in the uncincate fasciculus were significantly shorter in AYA cannabis users than in non-using controls (ref. 53). Interestingly, DeLisi et al. (2006) found a greater apparent diffusion coefficient in the frontal and subcortical areas of cannabis users compared to controls, which contrasts others findings (ref. 70).
Sex/gender differences in DTI
Of the included DTI studies, only Maple (2016) examined sex/gender differences, reporting decreased FA in the forceps minor of AYA cannabis using men, while finding increased FA in AYA cannabis using women (ref. 54).
Abstinence
Ashtari et al.’s (2009) finding of cannabis use-related changes in FA were documented in users who were abstinent for at least 3 months (M = 6.7 months) (ref. 64). Shollenbarger et al. (2015b) also found that the cannabis-related change in mean diffusivity of the uncinate fasciculus persisted after a week of abstinence (ref. 68).
Summary of DTI findings
Despite the wide-ranging results included in this section, collectively they indicate that cannabis use during AYA is associated with changes to the microstructure of the brain. Many of the included studies found that AYA cannabis use was associated with changes in the FA of multiple brain areas, loosely suggesting cannabis-related neuronal hyper- and hypoconnectivity. Other, more robust measures of neuronal microstructure were also used by the included studies to support the notion that AYA cannabis use is associated with changes to neuronal microstructure. Regarding sex/gender differences and abstinence effects in the DTI literature, very little can be concluded overall due to the paucity of relevant reporting on the topic, highlighting the need for future researchers to consider these constructs in their studies of AYA cannabis use and neuronal microstructure.
MRS
Of the relatively few (n = 4) MRS studies included, there were no significant themes or overlapping results. Bitter et al. (2014) found that cannabis users had an increased concentration of N-acetyl aspartate (NAA) in the ventral lateral prefrontal cortex, a marker of neuronal integrity and potentially increased neuronal density (ref. 75, ref. 76). Muetzel et al. (2013) found increased concentrations of myoinositol in the dorsal striatum in cannabis users, potentially indicating neuroinflammation (ref. 76, ref. 77). In the same study, Muetzel et al. (2013) also found lower concentrations of glutamine and glutamate in the dorsal striatum, potentially indicating decreased energy metabolism (ref. 78) and deficits in excitatory neurotransmission (ref. 76) in AYA cannabis users compared to non-users (ref. 77). Interestingly, Blest-Hopley et al. (2020) found decreased myoinositol in the hippocampus, again potentially indicating a neuroinflammation process in this region (ref. 76, ref. 79). Finally, Subramaniam et al. (2022) found decreased GABA in the anterior cingulate cortex, indicating a deficit in inhibitory neurotransmission (ref. 78, ref. 80).
Sex/gender differences in MRS
Only 2 of the included MRS studies explicitly assessed sex and/or gender differences related to the impact of AYA cannabis use on the brain (ref. 77, ref. 79). Of these, only Muetzel et al. (2013) reported a significant effect. Specifically, their finding of increased myoinositol and decreased glutamate and glutamine in the dorsal striatum was only present in female participants (ref. 77).
Summary of MRS findings
Again, due to the few MRS studies included in this review and their lack of uniformity regarding brain areas and metabolites of interest, there is no way to meaningfully abstract across these results. The limited number of studies assessing sex/gender and the complete lack of included studies explicitly assessing abstinence also make these constructs an important area of future research.
Other structural imaging modalities
The remaining structural imaging study utilized computed transaxial tomography (CT). In this early study, Co et al. (1977) reported no effect of cannabis use on any morphological brain measure in this cohort (ref. 81).
Functional neuroimaging
Task-based fMRI
Cognitive control
Of the included task-based fMRI studies, 31% (12/39) found that cannabis users’ brains responded significantly differently from non-users during tasks requiring cognitive control or decision making. During tasks requiring greater than normal levels of cognitive control (e.g., incongruent trials in the Stroop task or “no-go” trials in a go/no-go task), 10% of the included studies found that AYA cannabis use was associated with heightened activation in a number of brain regions compared to non-using controls (ref. 21, ref. 82–ref. 84). Similarly, with increasing task difficulty, Harding et al. (2012) found greater functional connectivity between multiple brain areas (ref. 85). However, other studies found that trials requiring cognitive control elicited less activity throughout the brains of AYA cannabis users compared to non-users. (ref. 86–ref. 88). Cyr et al. (2019) found that multiple brain regions which showed elevated activation in controls during trials requiring cognitive control showed no changes in cannabis users and Gruber et al. (2012) found that a smaller area of the anterior cingulate cortex (ACC) was activated in cannabis users compared to controls on trials of this type (ref. 89, ref. 90). Adding to this, Kroon et al. (2022) found that task difficulty increases were not associated with increased activation of the superior temporal gyrus in cannabis users (ref. 91)). Only one study found an effect of cannabis use on brain activity during trials which did not require above-average cognitive exertion (i.e., congruent trials in the Stroop task or “go” trials in a go/no-go task). In this study, Ames et al. (2013) found that cannabis users had greater activity in an array of brain areas during compatible trials of an implicit association task (ref. 86). However, both Claus et al. (2018) and Smith et al. (2011) found no differences in the activation patterns of the brains of cannabis users and non-users during tasks of cognitive control (ref. 92, ref. 93).
Learning and memory
The next most common fMRI task type was those which assessed learning and/or memory in some capacity (11/39). In a variety of memory tasks, the included studies found that AYA cannabis users displayed patterns of activation that significantly differed from the brains of non-users (ref. 94–ref. 97), with no particular pattern predominating. With respect to specific stages of memory, cannabis use was found to be associated with an absence of hippocampal activation during the encoding of new memories in two studies (ref. 98, ref. 99). In addition, these studies found that cannabis use was associated significantly with altered patterns of activity across other areas of the brain during encoding, such as the frontal lobes (ref. 98, ref. 99). During the recall of previously encoded information, Blest-Hopley et al. (2021) found no related activity change in the left posterior cingulate gyrus and right precuneus alongside alterations in the patterns of activation in midbrain structures (ref. 100). Further, Dager et al. (2018) found decreased recall-related activity in cannabis users (ref. 98). Regarding learning, Hubbard et al. (2023) observed a delayed increase in functional connectivity between frontoparietal and striatal regions as learning took place in cannabis users (ref. 101). In opposition, as part of a single longitudinal study, Cousijn et al. (2014a) and Cousijn et al. (2014b) reported no cannabis-related changes in brain function during memory tasks (ref. 102, ref. 103). Considered together, these results suggest that there likely exists changes to memory functioning attributable to AYA cannabis use.
Reward
The third most common type (8/39) of task-based fMRI studies assessed risk and reward systems. The first observable trend among these studies was that cannabis users displayed significantly increased activation in response to rewarding stimuli compared to non-users (ref. 104–ref. 106). Similarly, Nestor et al. (2020) found that cannabis users had increased functional connectivity across the brain when expecting reward (ref. 107). These studies suggest that the brains of AYA cannabis users are more sensitive to reward. Unlike these studies, Martz et al. (2016) observed that cannabis users had decreased activity in the nucleus accumbens when expecting reward (ref. 108). Lichenstein et al. (2017) found that cannabis users who increased their use throughout adolescence exhibited a pattern of brain activity opposite to non/low users and consistently high users when experiencing reward (ref. 109). Specifically, cannabis users who increased their use between the ages of 14–19 displayed inversely correlated patterns of activity between the nucleus accumbens and prefrontal cortex when winning in a card guessing game at age 20, as opposed to non/low users and consistently high users which displayed a direct relationship between nucleus accumbens and prefrontal cortex activation (ref. 109). Contrary to these studies, Karoly et al. (2015) and Macedo et al. (2024) found that AYA cannabis use was not associated with any changes in reward processing (ref. 110, ref. 111).
Social cognition
Only a small number (3/39) of the included studies assessed how cannabis use impacts social cognition. Gilman et al. (2016a) found that cannabis users lacked the usual elevated activation of the insula when being socially excluded (ref. 112). Further research has found the opposite, namely that tasks of social cognition elicit greater brain activity in cannabis users: Gilman et al. (2016b) found that a greater area of the striatum was activated by cannabis users when they were being socially influenced and that cannabis users had an increase in nucleus accumbens activity when succumbing to social influence relative to non-users (ref. 52). Finally, Gilman et al. (2016c) observed increased caudate activation in cannabis users during social influence generally and greater inferior frontal gyrus activity when rejecting social influence (ref. 113).
Emotion processing
Another small portion of included fMRI studies (3/39) investigated how cannabis use impacts the processing of emotion. Heitzeg et al. (2015) found that cannabis users displayed decreased activity in many areas when viewing negatively valanced words while also displaying increased activity when viewing positively valanced words (ref. 114). Nichols et al. (2021) made a similar observation, finding that cannabis users had greater activity in the left superior temporal gyrus in response to positive stimuli and less activity in response to negative stimuli (ref. 115). Spechler et al. (2020) found that cannabis use was associated with increased activity in the right amygdala when presented with angry faces at a baseline assessment and that cannabis users displayed no longitudinal increase in this response compared to non-users (ref. 116).
Other tasks
In a cue-reactivity task, Cousijn et al. (2013a) presented cannabis users and non-users with cannabis-related images while undergoing a fMRI, finding that cannabis users displayed greater brain activity in multiple regions when viewing cannabis-related images (ref. 117). Lopez-Larson et al. (2012) had cannabis users and non-users do a finger-tapping task and found that it elicited decreased activity in the cerebellum and right cingulate gyrus of cannabis users compared to non-users (ref. 118).
Abstinence
Regarding the effects of abstinence on cannabis-related task based functional changes, Macedo et al. (2024) found no abnormalities in brain activity in <1-month abstinent users undergoing a monetary incentive delay task, but also found no effect in current users, so the effect of abstinence on changes in reward processing in this case is unclear (ref. 111). Within the included studies, abnormalities in brain activity during tasks of spatial working memory (ref. 94, ref. 97) and response inhibition (ref. 84) were still observable after 28 days of abstinence. Finally, Sullivan et al. (2022) found blunted activation in the anterior cingulate cortex in response to fearful faces in cannabis users who were abstinent for 2 weeks (ref. 88).
Sex/gender differences in task-based fMRI
Dager et al. (2018) reported a sex effect in that only AYA cannabis using males displayed reduced activity in the bilateral inferior frontal gyrus and left hippocampus when correctly responding to a figural memory task (ref. 98). Sullivan et al. (2022) found, compared to women, AYA cannabis users who were men had significantly greater connectivity between the cerebellum and right rostral anterior cingulate cortex during a go/no-go task with emotional faces, which made their brain activity more in-line with non-users (ref. 88).
Summary of fMRI findings
Abstracting across the included fMRI studies makes it clear that AYA cannabis use is likely associated with a vast array of changes in brain activity during cognitive tasks. Significant increases and decreases in activity in a variety of brain areas were found during tasks of cognitive control, learning and memory, reward, social cognition, emotion processing, cue reactivity, and motor control. The fMRI studies with explicitly abstinent AYA cannabis users suggest that cannabis-associated changes in brain activity persist after 2 weeks to a month of non-use. Regarding sex/gender differences, due to the lack of studies explicitly assessing sex/gender and the heterogenous results of those which did, very little can be concluded until further evidence comes forward.
Resting-state fMRI
60% (6/10) of the included rs-fMRI studies found increased connectivity between many regions of the brain at rest in cannabis users (ref. 20, ref. 119–ref. 123). Other researchers have found that cannabis users exhibit decreased connectivity between a variety of regions of the brain (ref. 124–ref. 126). Common areas of change include the default mode network and (ref. 119, ref. 126), and the orbitofrontal cortex (ref. 121, ref. 125).
Abstinence in rs-fMRI
Jacobus et al. (2012) found significant differences in cerebral blood flow at baseline in AYA cannabis users compared to non-users, but this difference disappeared after the users underwent 4 weeks of abstinence (ref. 127).
Summary of rs-fMRI findings
Across the included studies, it appears clear that AYA cannabis use is associated with changes to the resting-state activity of the brain, particularly the default mode network and orbitofrontal cortex where both increases and decreases in activity have been observed. Following a trend present in other imaging modalities, very little re-fMRI evidence regarding the effects of sex/gender or abstinence exists, meaning these constructs should be the focus of future research.
EEG
Broyd et al. (2013) found that AYA cannabis users displayed diminished sensory gating and Greenwood et al. (2014) found significant abnormalities in EEG signals related to mismatch negativity (MMN) in AYA cannabis users (ref. 128, ref. 129). Broyd et al. (2016) replicated their sensory gating study, although findings were only different at a trend level (p>.05) (ref. 130). Apart from these findings, Maij et al. (2017) found that AYA cannabis users exhibited a significantly lower average amplitude of the P3 ERP at the Cz, Fz, and FCz electrodes during a go/no-go task, which the authors interpret as indicating deficient response inhibition (ref. 131). Finally, regarding memory, Smith et al. (2017) reported that AYA cannabis users had significantly lower N340 amplitude at electrodes C1, C2, Cz, FC1, FCz, and FC2 during the Rey Auditory Verbal Learning Task, indicating deficient memory (ref. 132).
Abstinence studies with EEG
The only EEG study to explicitly examine abstinent users was Broyd et al. (2016), reporting no significant abnormalities in ERPs relating to sensory processing in AYA cannabis users who had been abstinent for >1 month [M = 3.5 years; (ref. 130)]. Since Broyd et al. (2016) did not also make these measurements prior to abstinence nor compare these users to current cannabis users, the strength of this finding alone is questionable. However, when viewed in the context of Broyd et al.’s (2013) finding of deficiencies in sensory gating in current cannabis users, Broyd et al.’s (2016) findings more strongly suggest that a period of >1 month may remediate some of the sensory deficits related to AYA cannabis use (ref. 128, ref. 130).
Summary of EEG findings
The included EEG studies found that AYA cannabis use was associated with brain activity changes linked to sensory gating, executive function, and memory. Due to the small number of these studies overall as well as studies assessing sex/gender or abstinence, more research on the brain effects of AYA cannabis use using EEG is warranted. However, when comparing the included studies which assessed abstinence to each other, their results suggest that abstinence may reverse AYA cannabis use-related changes to brain activity detected by EEG.
PET
Of all studies included in the current review, only 2 used PET. Cox et al. (2020) reported an interaction between adolescent cannabis use and level of externalizing traits, such that participants who scored highly on their measure of externalizing traits and used cannabis regularly had a significantly decreased availability of glutamate receptors in their orbitofrontal cortex, striatum, and insula (ref. 133). Sevy et al. (2008) found that male adolescents who were former cannabis users had decreased glutamate metabolism in the putamen, right cerebellum, precuneus, right orbitofrontal cortex, and the right medial posterior parietal cortex (ref. 134).
Abstinence and PET
Sevy et al.’s (2008) finding of deficient glutamate metabolism in cannabis users was found in participants who were abstinent for at least one month [M = 15 weeks; (ref. 134)].
Summary of PET findings
Due to only 2 studies in the current review using PET to assess the brains of AYA cannabis users, no sweeping conclusions can be made generally or regarding sex/gender and abstinence.
Discussion
The purpose of this scoping review was to summarize the existing evidence regarding the effect that cannabis use during AYA (ages 14-25) may have on the structure, function, and metabolic function of the brain. Our summary of existing literature suggests that AYA cannabis use is associated with changes in cortical thickness, brain volumes, neuronal white matter microstructure, metabolite concentrations, task-based and resting-state functional brain activity, and neurotransmitter metabolism/receptor density. However, these results must be considered in light of the limitations of a scoping review, namely being the absence of evidence quality assessments and meta-analyses. These findings should also be considered alongside the goal of the current review, which was not to provide an answer to the question of AYA cannabis use’s effect on the brain, but to collate and summarize relevant research.
Regarding the effects of abstinence from cannabis use on the brain, we found a limited number of conflicting findings. The majority of included studies which assessed abstinent AYA cannabis users found that cannabis-related changes persisted over time (ref. 45, ref. 56, ref. 64, ref. 68, ref. 84, ref. 88, ref. 94, ref. 97, ref. 128, ref. 134), with one study finding that changes in the thickness of the layers of the hippocampus persisted for decades (ref. 37). The remaining studies instead found that abstinent AYA cannabis users did not differ from non-users on their particular neuroimaging measure (ref. 63, ref. 111, ref. 127, ref. 130). However, only one of these latter studies assessed the participants pre- and post-abstinence (ref. 127), limiting the usefulness of these results. Overall, these findings suggest that abstinence does not necessarily reverse cannabis-related brain changes when use is initiated in adolescence or emerging adulthood, which is concerning for this age group considering the brain development that occurs during this time (ref. 7).
Regarding sex and/or gender differences related to the brain effects of cannabis in the included studies, only 17/99 explicitly assessed how sex/gender impacted their results. Of these 17, only 5 found a significant sex or gender difference (ref. 54, ref. 55, ref. 77, ref. 88, ref. 98). McQueeny et al. (2011), Maple (2016), and Sullivan et al. (2022) assessed gender, while Muetzel et al. (2013) and Dager et al. (2018) assessed sex. In addition to these significant findings, Medina et al. (2009) found a trend-level effect of gender on AYA cannabis use-related brain volume changes (ref. 63). These findings are salient enough to elicit AYA sex/gender considerations as well as concern over how rare the practice of assessing sex/gender differences in conjunction with cannabis use in neuroimaging studies is. The potential interaction between sex/gender and cannabis brain effects with this AYA cohort is important in our further biological understanding of these interactions, as well as any sex/gender specific knowledge translation activity. It is thus important that future neuroimaging research investigating the effects of AYA cannabis use on the brain incorporates sex/gender analyses into their designs.
The current review has multiple strengths: we had our searches developed with the help of evidence synthesis experts and topic experts, we had two independent reviewers screen and extract data from all included studies, and we searched through multiple databases and preprint servers. However, the aforementioned results need to be considered in light of the limitations of scoping review methodology (ref. 34), We did not include an assessment of bias or assessment of the quality of the included studies. While a systematic review and meta-analysis could evaluate the quality of the evidence, a scoping review aggregates the results to a more easily actionable state. Systematic reviews and meta-analyses necessitate a more specific question and more restrictive inclusion criteria which would not allow us to properly assess the range of evidence that exists regarding the effect of adolescent/young adult cannabis use on the brain (ref. 27). In short, although a systematic review and meta-analysis would provide more rigor than a scoping review, it would require far more resources and limit the amount of evidence available for review. Another limitation of our review is the fact that it included mostly cross-sectional studies, which does not allow the establishment of temporal precedence, meaning that it is just as likely that brain abnormalities reported in this review could have existed prior to- or even led to AYA cannabis use. Finally, with this scoping review, cannabis use patterns, as well as preferred cannabis product used, were not a primary focus. Further research should consider these and other variables to inform the literature.
There are many potential implications of this review, including at the policy level. A prudent review and assessment of age of access to cannabis products, in those jurisdictions where cannabis is legalized, is warranted. Governments and local authorities should also invest in continued age-specific education around the potential brain effects resulting from AYA cannabis use and AYA specific interventions/harm reduction techniques. Finally, research needs to not only continue in this area, but it needs to do so with more consistency and rigor.
Conclusion
Here we show that there is evidence that cannabis use during AYA is associated with changes to brain structure and function, that these effects can still be seen after varying periods of abstinence, and that the sex/gender of the user might influence the relationship between AYA cannabis use and brain changes. Even though it is not the goal of the current review, it is worth noting that coming to a strong conclusion about the relationship between AYA cannabis use and brain changes would be exceedingly difficult due to the heterogeneity of the existing literature. Despite this, there are some consistent signals that are coming out of the literature showing, on balance, a negative impact on brain structure and function with cannabis use in this age group. What is difficult to comment on at this time, is the potential varying effects of amounts of cannabis used and potency of product on these brain imaging variables, and effects of longer period of abstinence. With few studies examining sex/gender, little can be said on these specific effects as well, so we recommend that future research meaningfully consider detailed cannabis use patterns/products used and sex/gender in their designs and analyses.
References
- 1 World Health Organization. Adolescent and young adult health (2024). Available online at: https://www.who.int/news-room/fact-sheets/detail/adolescents-health-risks-and-solutions (Accessed March 03, 2025).
- H Sampasa-Kanyinga, HA Hamilton, AG LeBlanc, J-P Chaput. Cannabis use among middle and high school students in Ontario: a school-based cross-sectional study.. CMAJ Open. (, 2018. [DOI | PubMed]
- AD Hamilton, JB Jang, ME Patrick, JE Schulenberg, KM Keyes. Age, period and cohort effects in frequent cannabis use among US students: 1991–2018.. Addiction. (, 2019. [DOI | PubMed]
- Canadian cannabis survey 2023: Summary (. 2024
- RA Miech, LD Johnston, ME Patrick, PM O’Malley, JG Bachman, JE Schulenberg. Monitoring the Future national survey results on drug use, 1975-2022: Secondary school students.. 2023
- Cannabis – the current situation in Europe (European Drug Report 2025) (. 2025
- K Konrad, C Firk, PJ Uhlhaas. Brain development during adolescence.. Dtsch Arztebl Int. (, 2013. [DOI | PubMed]
- K Veroude, J Jolles, G Croiset, L Krabbendam. Changes in neural mechanisms of cognitive control during the transition from late adolescence to young adulthood.. Dev Cognit Neurosci. (, 2013. [DOI | PubMed]
- CM Gómez, CI Barriga-Paulino, EI Rodríguez-Martínez, MÁ Rojas-Benjumea, A Arjona, J Gómez-González. The neurophysiology of working memory development: from childhood to adolescence and young adulthood.. Rev Neurosci. (, 2018. [DOI | PubMed]
- L Steinberg. A social neuroscience perspective on adolescent risk-taking.. Dev Rev. (, 2008. [DOI | PubMed]
- L Van Leijenhorst, K Zanolie, CS Van Meel, PM Westenberg, SARB Rombouts, EA Crone. What motivates the adolescent? brain regions mediating reward sensitivity across adolescence.. Cereb Cortex. (, 2010. [DOI | PubMed]
- L Dellazizzo, S Potvin, S Giguère, A Dumais. Evidence on the acute and residual neurocognitive effects of cannabis use in adolescents and adults: a systematic meta-review of meta-analyses.. Addiction. (, 2022. [DOI | PubMed]
- L Degenhardt, C Coffey, H Romaniuk, W Swift, JB Carlin, WD Hall. The persistence of the association between adolescent cannabis use and common mental disorders into young adulthood.. Addiction. (, 2013. [DOI | PubMed]
- G Gobbi, T Atkin, T Zytynski, S Wang, S Askari, J Boruff. Association of cannabis use in adolescence and risk of depression, anxiety, and suicidality in young adulthood: a systematic review and meta-analysis.. JAMA Psychiatry. (, 2019. [DOI | PubMed]
- M Taylor, SM Collin, MR Munafò, J MacLeod, M Hickman, J Heron. Patterns of cannabis use during adolescence and their association with harmful substance use behaviour: findings from a UK birth cohort.. J Epidemiol Community Health. (, 1978. [DOI | PubMed]
- K Schmidt, I Tseng, A Phan, T Fong, J Tsuang. A systematic review: adolescent cannabis use and suicide.. Addict Disord Their Treat. (, 2020. [DOI]
- DT Myran, M Pugliese, LD Harrison, M Solmi, KK Anderson, JG Fiedorowicz. changes in incident schizophrenia diagnoses associated with cannabis use disorder after cannabis legalization.. JAMA Netw Open. (, 2025. [DOI | PubMed]
- M Athanassiou, A Dumais, G Gnanhoue, A Abdel-Baki, D Jutras-Aswad, S Potvin. A systematic review of longitudinal studies investigating the impact of cannabis use in patients with psychotic disorders.. Expert Rev Neurother. (, 2021. [DOI | PubMed]
- MD Albaugh, MM Owens, A Juliano, J Ottino-Gonzalez, R Cupertino, Z Cao. Differential associations of adolescent versus young adult cannabis initiation with longitudinal brain change and behavior.. Mol Psychiatry. (, 2023. [DOI | PubMed]
- N Ertl, W Lawn, C Mokrysz, TP Freeman, N Alnagger, A Borissova. Associations between regular cannabis use and brain resting-state functional connectivity in adolescents and adults.. J Psychopharmacol. (, 2023. [DOI | PubMed]
- T Hatchard, PA Fried, MJ Hogan, I Cameron, AM Smith. Marijuana use impacts cognitive interference: an fMRI investigation in young adults performing the counting Stroop task.. J Addict Res Ther. (, 2014. [DOI]
- AP Prescot, AE Locatelli, PF Renshaw, DA Yurgelun-Todd. Neurochemical alterations in adolescent chronic marijuana smokers: a proton MRS study.. Neuroimage. (, 2011. [DOI | PubMed]
- HE Atmaca. Effects of recreational cannabis use and subclinical psychosis risk on brain white matter integrity and structural connectivity in adolescence., 2022
- JC Scott. Impact of adolescent cannabis use on neurocognitive and brain development.. Child Adolesc Psychiatr Clin N Am. (, 2023. [DOI | PubMed]
- SD Lichenstein, N Manco, LM Cope, L Egbo, KA Garrison, J Hardee. Systematic review of structural and functional neuroimaging studies of cannabis use in adolescence and emerging adulthood: evidence from 90 studies and 9441 participants.. Neuropsychopharmacology. (, 2022. [DOI | PubMed]
- AC Tricco, E Lillie, W Zarin, K O’Brien, H Colquhoun, M Kastner. A scoping review on the conduct and reporting of scoping reviews.. BMC Med Res Methodol. (, 2016. [DOI | PubMed]
- Z Munn, MDJ Peters, C Stern, C Tufanaru, A McArthur, E Aromataris. Systematic review or scoping review? Guidance for authors when choosing between a systematic or scoping review approach.. BMC Med Res Methodol. (, 2018. [DOI | PubMed]
- J Grimshaw. A knowledge synthesis chapter . Available online at:
- MD Peters, C Godfrey, P McInerney, Z Munn, AC Tricco, H Khalil. Scoping reviews. In:. JBI Manual for Evidence Synthesis., 2024. [DOI]
- 30 Open Science Framework. OSF. Washington DC, Center for Open Science.
- JA Nosko, CE Crocker, C Hambly Lapointe, PG Tibbo. Cannabis use in adolescence and young adulthood and its effects on brain structure and function: a scoping review. (. 2024. [DOI]
- 32 The National Health Service. Early or delayed puberty. NHS. Available online at: https://www.nhs.uk/conditions/early-or-delayed-puberty/ (Accessed April 29, 2025).
- Covidence Systematic Review Software. Available online at:
- AC Tricco, E Lillie, W Zarin, KK O’Brien, H Colquhoun, D Levac. PRISMA extension for scoping reviews (PRISMA-ScR): checklist and explanation.. Ann Intern Med. (, 2018. [DOI | PubMed]
- 35 World Health Organization. Gender and health. Available online at: https://www.who.int/health-topics/gender#tab=tab_1 (Accessed March 03, 2025).
- MD Albaugh, J Ottino-Gonzalez, A Sidwell, C Lepage, A Juliano, MM Owens. Association of cannabis use during adolescence with neurodevelopment.. JAMA Psychiatry. (, 2021. [DOI | PubMed]
- AC Burggren, P Siddarth, Z Mahmood, ED London, TM Harrison, DA Merrill. Subregional hippocampal thickness abnormalities in older adults with a history of heavy cannabis use.. Cannabis Cannabinoid Res. (, 2018. [DOI | PubMed]
- KM Lisdahl, L Tamm, JN Epstein, T Jernigan, BSG Molina, SP Hinshaw. The impact of ADHD persistence, recent cannabis use, and age of regular cannabis use onset on subcortical volume and cortical thickness in young adults.. Drug Alcohol Depend. (, 2016. [DOI | PubMed]
- MP Lopez-Larson, P Bogorodzki, J Rogowska, E McGlade, JB King, J Terry. Altered prefrontal and insular cortical thickness in adolescent marijuana users.. Behav Brain Res. (, 2011. [DOI | PubMed]
- I Mata, R Perez-Iglesias, R Roiz-Santianez, D Tordesillas-Gutierrez, A Pazos, A Gutierrez. Gyrification brain abnormalities associated with adolescence and early-adulthood cannabis use.. Brain Res. (, 2010. [DOI | PubMed]
- MM Owens, MD Albaugh, N Allgaier, D Yuan, G Robert, RB Cupertino. Bayesian causal network modeling suggests adolescent cannabis use accelerates prefrontal cortical thinning.. Transl Psychiatry. (, 2022. [DOI | PubMed]
- M Radoman, SS Hoeppner, RM Schuster, AE Evins, JM Gilman. Marijuana use and major depressive disorder are additively associated with reduced verbal learning and altered cortical thickness.. Cognit Affect Behav Neurosci. (, 2019. [DOI | PubMed]
- JC Scott, AFG Rosen, TM Moore, DR Roalf, TD Satterthwaite, ME Calkins. Cannabis use in youth is associated with limited alterations in brain structure.. Neuropsychopharmacology. (, 2019. [DOI | PubMed]
- MH Meier, RA Schriber, J Beardslee, J Hanson, D Pardini. Associations between adolescent cannabis use frequency and adult brain structure: a prospective study of boys followed to adulthood.. Drug Alcohol Depend. (, 2019. [DOI | PubMed]
- M Ashtari, B Avants, L Cyckowski, KL Cervellione, D Roofeh, P Cook. Medial temporal structures and memory functions in adolescents with heavy cannabis use.. J Psychiatr Res. (, 2011. [DOI | PubMed]
- A Batalla, V Lorenzetti, Y Chye, M Yücel, C Soriano-Mas, S Bhattacharyya. The Influence of DAT1, COMT, and BDNF genetic polymorphisms on total and subregional hippocampal volumes in early onset heavy cannabis users.. Cannabis Cannabinoid Res. (, 2018. [DOI | PubMed]
- JC Churchwell, M Lopez-Larson, DA Yurgelun-Todd. Altered frontal cortical volume and decision making in adolescent cannabis users.. Front Psychol. (, 2010. [DOI | PubMed]
- J Cousijn, RW Wiers, KR Ridderinkhof, W van den Brink, DJ Veltman, AE Goudriaan. Grey matter alterations associated with cannabis use: results of a VBM study in heavy cannabis users and healthy controls.. Neuroimage. (, 2012. [DOI | PubMed]
- T Demirakca, A Sartorius, G Ende, N Meyer, H Welzel, G Skopp. Diminished gray matter in the hippocampus of cannabis users: possible protective effects of cannabidiol.. Drug Alcohol Depend. (, 2011. [DOI | PubMed]
- A Garimella, S Rajguru, UK Singla, V Alluri. Marijuana and the hippocampus: a longitudinal study on the effects of marijuana on hippocampal subfields.. Prog Neuropsychopharmacol Biol Psychiatry. (, 2020. [DOI | PubMed]
- JM Gilman, JK Kuster, S Lee, MJ Lee, BW Kim, N Makris. Cannabis use is quantitatively associated with nucleus accumbens and amygdala abnormalities in young adult recreational users.. J Neurosci. (, 2014. [DOI | PubMed]
- JM Gilman, S Lee, JK Kuster, MJ Lee, BW Kim, A van der Kouwe. Variable activation in striatal subregions across components of a social influence task in young adult cannabis users.. Brain Behav. (, 2016. [DOI | PubMed]
- N Levar, AN Francis, MJ Smith, WC Ho, JM Gilman. Verbal memory performance and reduced cortical thickness of brain regions along the uncinate fasciculus in young adult cannabis users.. Cannabis Cannabinoid Res. (, 2018. [DOI | PubMed]
- KE Maple. Cannabis use and affective processing: a brain structure analysis.. 2016
- T McQueeny, CB Padula, J Price, KL Medina, P Logan, SF Tapert. Gender effects on amygdala morphometry in adolescent marijuana users.. Behav Brain Res. (, 2011. [DOI | PubMed]
- KL Medina, BJ Nagel, SF Tapert. Abnormal cerebellar morphometry in abstinent adolescent marijuana users.. Psychiatry Res Neuroimaging. (, 2010. [DOI | PubMed]
- JS Price, T McQueeny, S Shollenbarger, EL Browning, J Wieser, KM Lisdahl. Effects of marijuana use on prefrontal and parietal volumes and cognition in emerging adults.. Psychopharmacol (Berl). (, 2015. [DOI | PubMed]
- SG Shollenbarger, J Price, J Wieser, K Lisdahl. Impact of cannabis use on prefrontal and parietal cortex gyrification and surface area in adolescents and emerging adults.. Dev Cognit Neurosci. (, 2015. [DOI | PubMed]
- T Yu, T Jia, L Zhu, S Desrivières, C Macare, Y Bi. Cannabis-associated psychotic-like experiences are mediated by developmental changes in the parahippocampal gyrus.. J Am Acad Child Adolesc Psychiatry. (, 2020. [DOI | PubMed]
- BJ Weiland, RE Thayer, BE Depue, A Sabbineni, AD Bryan, KE Hutchison. Daily marijuana use is not associated with brain morphometric measures in adolescents or adults.. J Neurosci. (, 2015. [DOI | PubMed]
- KL Medina, BJ Nagel, A Park, T McQueeny, SF Tapert. Depressive symptoms in adolescents: associations with white matter volume and marijuana use.. J Child Psychol Psychiatry. (, 2007. [DOI | PubMed]
- L Koenders, V Lorenzetti, L de Haan, C Suo, WAM Vingerhoets, W van den Brink. Longitudinal study of hippocampal volumes in heavy cannabis users.. J Psychopharmacol. (, 2017. [DOI | PubMed]
- KL Medina, T McQueeny, BJ Nagel, KL Hanson, TT Yang, SF Tapert. Prefrontal cortex morphometry in abstinent adolescent marijuana users: subtle gender effects.. Addict Biol. (, 2009. [DOI | PubMed]
- M Ashtari, K Cervellione, J Cottone, BA Ardekani, S Kumra. Diffusion abnormalities in adolescents and young adults with a history of heavy cannabis use.. J Psychiatr Res. (, 2009. [DOI | PubMed]
- MP Becker, PF Collins, KO Lim, RL Muetzel, M Luciana. Longitudinal changes in white matter microstructure after heavy cannabis use.. Dev Cognit Neurosci. (, 2015. [DOI | PubMed]
- SA Gruber, MK Dahlgren, KA Sagar, A Gonenc, SE Lukas. Worth the wait: effects of age of onset of marijuana use on white matter and impulsivity.. Psychopharmacol (Berl). (, 2014. [DOI | PubMed]
- SD Lichenstein, DS Shaw, EE Forbes. Cannabis, connectivity, and coming of age: Associations between cannabis use and anterior cingulate cortex connectivity during the transition to adulthood.. Front Hum Neurosci. (, 2022. [DOI | PubMed]
- SG Shollenbarger, J Price, J Wieser, K Lisdahl. Poorer frontolimbic white matter integrity is associated with chronic cannabis use, FAAH genotype, and increased depressive and apathy symptoms in adolescents and young adults.. NeuroImage Clin. (, 2015. [DOI | PubMed]
- N Dekker, N Schmitz, BD Peters, TA van Amelsvoort, DH Linszen, L de Haan. Cannabis use and callosal white matter structure and integrity in recent-onset schizophrenia.. Psychiatry Res Neuroimaging. (, 2010. [DOI | PubMed]
- LE DeLisi, HC Bertisch, KU Szulc, M Majcher, K Brown, A Bappal. A preliminary DTI study showing no brain structural change associated with adolescent cannabis use.. Harm Reduct J. (, 2006. [DOI | PubMed]
- J Cousijn, YJ Toenders, LS Velzen, AM Kaag. The relation between cannabis use, dependence severity and white matter microstructure: a diffusion tensor imaging study.. Addict Biol. (, 2022. [DOI | PubMed]
- CR Figley, MN Uddin, K Wong, J Kornelsen, J Puig, TD Figley. Potential pitfalls of using fractional anisotropy, axial diffusivity, and radial diffusivity as biomarkers of cerebral white matter microstructure.. Front Neurosci. (, 2022. [DOI | PubMed]
- N Solowij, A Zalesky, V Lorenzetti, M Yücel. Chapter 40 – chronic cannabis use and axonal fiber connectivity. In:. Handbook of Cannabis and Related Pathologies., 2017. [DOI]
- SA Gruber, MM Silveri, MK Dahlgren, D Yurgelun-Todd. Why so impulsive? White matter alterations are associated with impulsivity in chronic marijuana smokers.. Exp Clin Psychopharmacol. (, 2011. [DOI | PubMed]
- SM Bitter, WA Weber, W-J Chu, CM Adler, JC Eliassen, SM Strakowski. N-acetyl aspartate levels in adolescents with bipolar and/or cannabis use disorders.. J Dual Diagn. (, 2014. [DOI | PubMed]
- M Kulich, LM Fisher, C Voelker. Imaging findings in mild traumatic brain injury. In:. Neurosensory Disorders in Mild Traumatic Brain Injury., 2019. [DOI]
- RL Muetzel, M Marjańska, PF Collins, MP Becker, R Valabrègue, EJ Auerbach. In vivo 1H magnetic resonance spectroscopy in young-adult daily marijuana users.. NeuroImage Clin. (, 2013. [DOI | PubMed]
- SC Licata, PF Renshaw. Neurochemistry of drug action: insights from proton magnetic resonance spectroscopic imaging and their relevance to addiction.. Ann N Y Acad Sci. (, 2010. [DOI | PubMed]
- G Blest-Hopley, A O’Neill, R Wilson, V Giampietro, D Lythgoe, A Egerton. Adolescent-onset heavy cannabis use associated with significantly reduced glial but not neuronal markers and glutamate levels in the hippocampus.. Addict Biol. (, 2020. [DOI | PubMed]
- P Subramaniam, A Prescot, E McGlade, P Renshaw, D Yurgelun-Todd. Examination of gamma-aminobutyric acid and glutamate-glutamine levels in association with impulsive behavior in adolescent marijuana users.. Drug Alcohol Depend. (, 2022. [DOI | PubMed]
- BT Co, DW Goodwin, M Gado, M Mikhael, SY Hill. Absence of cerebral atrophy in chronic cannabis users. Evaluation by computerized transaxial tomography.. J Am Med Assoc. (, 1977. [DOI | PubMed]
- J Aloi, KS Blair, KI Crum, H Meffert, SF White, PM Tyler. Adolescents show differential dysfunctions related to Alcohol and Cannabis Use Disorder severity in emotion and executive attention neuro-circuitries.. NeuroImage Clin. (, 2018. [DOI | PubMed]
- KA Sagar, MK Dahlgren, A Gonenc, MT Racine, MW Dreman, SA Gruber. The impact of initiation: Early onset marijuana smokers demonstrate altered Stroop performance and brain activation.. Dev Cognit Neurosci. (, 2015. [DOI | PubMed]
- SF Tapert, AD Schweinsburg, SPA Drummond, MP Paulus, SA Brown, TT Yang. Functional MRI of inhibitory processing in abstinent adolescent marijuana users.. Psychopharmacol (Berl). (, 2007. [DOI | PubMed]
- IH Harding, N Solowij, BJ Harrison, M Takagi, V Lorenzetti, DI Lubman. Functional connectivity in brain networks underlying cognitive control in chronic cannabis users.. Neuropsychopharmacology. (, 2012. [DOI | PubMed]
- SL Ames, JL Grenard, AW Stacy, L Xiao, Q He, SW Wong. Functional imaging of implicit marijuana associations during performance on an Implicit Association Test (IAT).. Behav Brain Res. (, 2013. [DOI | PubMed]
- KE Maple. Cannabis-using youth demonstrated blunted rostral anterior cingulate cortex activation, but normal functional connectivity, during an emotional go/no-go task.. 2019
- RM Sullivan, KE Maple, AL Wallace, AM Thomas, KM Lisdahl. Examining inhibitory affective processing within the rostral anterior cingulate cortex among abstinent cannabis-using adolescents and young adults.. Front Psychiatry. (, 2022. [DOI | PubMed]
- M Cyr, GZ Tau, M Fontaine, FR Levin, R Marsh. Deficient functioning of frontostriatal circuits during the resolution of cognitive conflict in cannabis-using youth.. J Am Acad Child Adolesc Psychiatry. (, 2019. [DOI | PubMed]
- SA Gruber, MK Dahlgren, KA Sagar, A Gönenc, WDS Killgore. Age of onset of marijuana use impacts inhibitory processing.. Neurosci Lett. (, 2012. [DOI | PubMed]
- E Kroon, L Kuhns, J Cousijn. Context dependent differences in working memory related brain activity in heavy cannabis users.. Psychopharmacol (Berl). (, 2022. [DOI | PubMed]
- ED Claus, SW Feldstein Ewing, RE Magnan, E Montanaro, KE Hutchison, AD Bryan. Neural mechanisms of risky decision making in adolescents reporting frequent alcohol and/or marijuana use.. Brain Imaging Behav. (, 2018. [DOI | PubMed]
- AM Smith, RAL Zunini, CD Anderson, CA Longo, I Cameron, MJ Hogan. Impact of marijuana on response inhibition: an fMRI study in young adults.. J Behav Brain Sci. (, 2011. [DOI]
- AD Schweinsburg, BJ Nagel, BC Schweinsburg, A Park, RJ Theilmann, SF Tapert. Abstinent adolescent marijuana users show altered fMRI response during spatial working memory.. Psychiatry Res Neuroimaging. (, 2008. [DOI | PubMed]
- AM Smith, CA Longo, PA Fried, MJ Hogan, I Cameron. Effects of marijuana on visuospatial working memory: an fMRI study in young adults.. Psychopharmacol (Berl). (, 2010. [DOI | PubMed]
- B Tervo-Clemmens, D Simmonds, FJ Calabro, DF Montez, JA Lekht, NL Day. Early cannabis use and neurocognitive risk: a prospective functional neuroimaging study.. Biol Psychiatry Cognit Neurosci Neuroimaging. (, 2018. [DOI | PubMed]
- CB Padula, AD Schweinsburg, SF Tapert. Spatial working memory performance and fmri activation interactions in abstinent adolescent marijuana users.. Psychol Addict Behav. (, 2007. [DOI | PubMed]
- AD Dager, MR Tice, GA Book, H Tennen, SA Raskin, CS Austad. Relationship between fMRI response during a nonverbal memory task and marijuana use in college students.. Drug Alcohol Depend. (, 2018. [DOI | PubMed]
- AD Schweinsburg, BC Schweinsburg, BJ Nagel, LT Eyler, SF Tapert. Neural correlates of verbal learning in adolescent alcohol and marijuana users.. Addiction. (, 2011. [DOI | PubMed]
- G Blest-Hopley, A O’Neill, R Wilson, V Giampietro, S Bhattacharyya. Disrupted parahippocampal and midbrain function underlie slower verbal learning in adolescent-onset regular cannabis use.. Psychopharmacol (Berl). (, 2021. [DOI | PubMed]
- NA Hubbard, KB Miller, J Aloi, S Bajaj, KT Wakabayashi, RJR Blair. Evaluating instrumental learning and striatal–cortical functional connectivity in adolescent alcohol and cannabis use.. Addict Biol. (, 2023. [DOI | PubMed]
- J Cousijn, RW Wiers, KR Ridderinkhof, W van den Brink, DJ Veltman, AE Goudriaan. Effect of baseline cannabis use and working-memory network function on changes in cannabis use in heavy cannabis users: a prospective fMRI study.. Hum Brain Mapp. (, 2014. [DOI | PubMed]
- J Cousijn, WAM Vingerhoets, L Koenders, L de Haan, W van den Brink, RW Wiers. Relationship between working-memory network function and substance use: a 3-year longitudinal fMRI study in heavy cannabis users and controls.. Addict Biol. (, 2014. [DOI | PubMed]
- A Acheson, KL Ray, CS Hines, K Li, MA Dawes, CW Mathias. Functional activation and effective connectivity differences in adolescent marijuana users performing a simulated gambling task.. J Addict. (, 2015. [DOI | PubMed]
- M Skumlien, C Mokrysz, TP Freeman, MB Wall, M Bloomfield, R Lees. Neural responses to reward anticipation and feedback in adult and adolescent cannabis users and controls.. Neuropsychopharmacology. (, 2022. [DOI | PubMed]
- J Cousijn, RW Wiers, KR Ridderinkhof, W van den Brink, DJ Veltman, LJ Porrino. Individual differences in decision making and reward processing predict changes in cannabis use: a prospective functional magnetic resonance imaging study.. Addict Biol. (, 2013. [DOI | PubMed]
- LJ Nestor, B Behan, J Suckling, H Garavan. Cannabis-dependent adolescents show differences in global reward-associated network topology: a functional connectomics approach.. Addict Biol. (, 2020. [DOI | PubMed]
- ME Martz, EM Trucco, LM Cope, JE Hardee, JM Jester, RA Zucker. Association of marijuana use with blunted nucleus accumbens response to reward anticipation.. JAMA Psychiatry. (, 2016. [DOI | PubMed]
- SD Lichenstein, S Musselman, DS Shaw, S Sitnick, EE Forbes. Nucleus accumbens functional connectivity at age 20 is associated with trajectory of adolescent cannabis use and predicts psychosocial functioning in young adulthood.. Addiction. (, 2017. [DOI | PubMed]
- HC Karoly, AD Bryan, BJ Weiland, A Mayer, A Dodd, SW Feldstein Ewing. Does incentive-elicited nucleus accumbens activation differ by substance of abuse? An examination with adolescents.. Dev Cognit Neurosci. (, 2015. [DOI | PubMed]
- I Macedo, TO Paiva, R Pasion, L Daedelow, A Heinz, A Magalhaes. Light cannabis use and the adolescent brain: an 8-years longitudinal assessment of mental health, cognition, and reward processing.. Psychopharmacol (Berl). (, 2024. [DOI | PubMed]
- JM Gilman, MT Curran, V Calderon, RM Schuster, AE Evins. Altered neural processing to social exclusion in young adult marijuana users.. Biol Psychiatry Cognit Neurosci Neuroimaging. (, 2016. [DOI | PubMed]
- JM Gilman, RM Schuster, MT Curran, V Calderon, A van der Kouwe, AE Evins. Neural mechanisms of sensitivity to peer information in young adult cannabis users.. Cognit Affect Behav Neurosci. (, 2016. [DOI | PubMed]
- MM Heitzeg, LM Cope, ME Martz, JE Hardee, RA Zucker. Brain activation to negative stimuli mediates a relationship between adolescent marijuana use and later emotional functioning.. Dev Cognit Neurosci. (, 2015. [DOI | PubMed]
- ES Nichols, J Penner, KA Ford, M Wammes, RWJ Neufeld, DGV Mitchell. Emotion regulation in emerging adults with major depressive disorder and frequent cannabis use.. NeuroImage Clin. (, 2021. [DOI | PubMed]
- PA Spechler, B Chaarani, C Orr, MD Albaugh, NR Fontaine, ST Higgins. Longitudinal associations between amygdala reactivity and cannabis use in a large sample of adolescents.. Psychopharmacol (Berl). (, 2020. [DOI | PubMed]
- J Cousijn, AE Goudriaan, KR Ridderinkhof, W van den Brink, DJ Veltman, RW Wiers. Neural responses associated with cue-reactivity in frequent cannabis users.. Addict Biol. (, 2013. [DOI | PubMed]
- MP Lopez-Larson, J Rogowska, P Bogorodzki, CE Bueler, EC McGlade, DA Yurgelun-Todd. Cortico-cerebellar abnormalities in adolescents with heavy marijuana use.. Psychiatry Res Neuroimaging. (, 2012. [DOI | PubMed]
- C Kelly, FX Castellanos, O Tomaselli, K Lisdahl, L Tamm, T Jernigan. Distinct effects of childhood ADHD and cannabis use on brain functional architecture in young adults.. NeuroImage Clin. (, 2017. [DOI | PubMed]
- L Ma, JM Hettema, J Cousijn, JM Bjork, JL Steinberg, L Keyser-Marcus. Resting-state directional connectivity and anxiety and depression symptoms in adult cannabis users.. Biol Psychiatry Cognit Neurosci Neuroimaging. (, 2021. [DOI | PubMed]
- MP Lopez-Larson, J Rogowska, D Yurgelun-Todd. Aberrant orbitofrontal connectivity in marijuana smoking adolescents.. Dev Cognit Neurosci. (, 2015. [DOI | PubMed]
- C Orr, R Morioka, B Behan, S Datwani, M Doucet, J Ivanovic. Altered resting-state connectivity in adolescent cannabis users.. Am J Drug Alcohol Abuse. (, 2013. [DOI | PubMed]
- B Behan, CG Connolly, S Datwani, M Doucet, J Ivanovic, R Morioka. Response inhibition and elevated parietal-cerebellar correlations in chronic adolescent cannabis users.. Neuropharmacology. (, 2014. [DOI | PubMed]
- J Camchong, PF Collins, MP Becker, KO Lim, M Luciana. Longitudinal alterations in prefrontal resting brain connectivity in non-treatment-seeking young adults with cannabis use disorder.. Front Psychiatry. (, 2019. [DOI | PubMed]
- A Wilson, K Gicas, WD Stevens, L Sergio, M Wojtowicz. Substance use is associated with worse mental health and altered resting state functional connectivity in female university athletes at baseline: a pilot study.. PloS One. (, 2021. [DOI | PubMed]
- EA Osuch, K Manning, RA Hegele, J Theberge, R Neufeld, D Mitchell. Depression, marijuana use and early-onset marijuana use conferred unique effects on neural connectivity and cognition.. Acta Psychiatr Scand. (, 2016. [DOI | PubMed]
- J Jacobus, D Goldenberg, CE Wierenga, NJ Tolentino, TT Liu, SF Tapert. Altered cerebral blood flow and neurocognitive correlates in adolescent cannabis users.. Psychopharmacol (Berl). (, 2012. [DOI | PubMed]
- SJ Broyd, L-M Greenwood, RJ Croft, A Dalecki, J Todd, PT Michie. Chronic effects of cannabis on sensory gating.. Int J Psychophysiol. (, 2013. [DOI | PubMed]
- L-M Greenwood, SJ Broyd, R Croft, J Todd, PT Michie, S Johnstone. Chronic effects of cannabis use on the auditory mismatch negativity.. Biol Psychiatry. (, 2014. [DOI | PubMed]
- SJ Broyd, L Greenwood, HH van Hell, RJ Croft, H Coyle, B Lee-Bates. Mismatch negativity and P50 sensory gating in abstinent former cannabis users.. Neural Plast. (, 2016. [DOI | PubMed]
- DLR Maij, BJM van de Wetering, IHA Franken, DL Maij, BJ van de Wetering, IH Franken. Cognitive control in young adults with cannabis use disorder: An event-related brain potential study.. J Psychopharmacol. (, 2017. [DOI | PubMed]
- JL Smith, FM De Blasio, JM Iredale, AJ Matthews, R Bruno, M Dwyer. Verbal learning and memory in cannabis and alcohol users: an event-related potential investigation.. Front Psychol. (, 2017. [DOI | PubMed]
- SML Cox, M Tippler, N Jaworska, K Smart, N Castellanos-Ryan, F Durand. mGlu5 receptor availability in youth at risk for addictions: effects of vulnerability traits and cannabis use.. Neuropsychopharmacology. (, 2020. [DOI | PubMed]
- S Sevy, GS Smith, Y Ma, V Dhawan, T Chaly, PB Kingsley. Cerebral glucose metabolism and D2/D3 receptor availability in young adults with cannabis dependence measured with positron emission tomography.. Psychopharmacol (Berl). (, 2008. [DOI | PubMed]