
Epigenetic Studies for Evaluation of NPS Toxicity: Focus on Synthetic Cannabinoids and Cathinones
Abstract
In the recent decade, numerous new psychoactive substances (NPSs) have been added to the illicit drug market. These are synthetized to mimic the effects of classic drugs of abuse (i.e., cannabis, cocaine, etc.), with the purpose of bypassing substance legislations and increasing the pharmacotoxicological effects. To date, research into the acute pharmacological effects of new NPSs is ongoing and necessary in order to provide an appropriate contribution to public health. In fact, multiple examples of NPS-related acute intoxication and mortality have been recorded in the literature. Accordingly, several in vitro and in vivo studies have investigated the pharmacotoxicological profiles of these compounds, revealing that they can cause adverse effects involving various organ systems (i.e., cardiovascular, respiratory effects) and highlighting their potential increased consumption risks. In this sense, NPSs should be regarded as a complex issue that requires continuous monitoring. Moreover, knowledge of long-term NPS effects is lacking. Because genetic and environmental variables may impact NPS responses, epigenetics may aid in understanding the processes behind the harmful events induced by long-term NPS usage. Taken together, “pharmacoepigenomics” may provide a new field of combined study on genetic differences and epigenetic changes in drug reactions that might be predictive in forensic implications.
Article type: Review Article
Keywords: new psychoactive substances, pharmacoepigenomics, forensic science
Affiliations: Department of Neurosciences and Rehabilitation, Section of Medical Biochemistry, Molecular Biology and Genetics, University of Ferrara, 44121 Ferrara, Italy; leila.mazdai@unife.it (L.M.); michele.rubini@unife.it (M.R.); Department of Translational Medicine, Section of Legal Medicine, LTTA Center, University of Ferrara, 44121 Ferrara, Italy; fbbmtt1@unife.it (M.F.); micaela.tirri@unife.it (M.T.); giorgia.corli@unife.it (G.C.); raffaella.arfe@unife.it (R.A.); beatrice.marchetti@unife.it (B.M.); bllsrn@unife.it (S.B.); rosamaria.gaudio@unife.it (R.M.G.); Section of Legal Medicine, Department of Health Care Surveillance and Bioethics, Università Cattolica del Sacro Cuore, 00168 Rome, Italy; eb95@live.it; Fondazione IRCCS Policlinico Universitario A. Gemelli, 00168 Rome, Italy; University Center for Studies on Gender Medicine, University of Ferrara, 44121 Ferrara, Italy; Department of Anti-Drug Policies, Collaborative Center for the Italian National Early Warning System, Presidency of the Council of Ministers, 11582 Rome, Italy
License: © 2022 by the authors. CC BY 4.0 Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Article links: DOI: 10.3390/biomedicines10061398 | PubMed: 35740419 | PMC: PMC9219842
Relevance: Moderate: mentioned 3+ times in text
Full text: PDF (862 KB)
1. Introduction
In the last decade there has been a significant change in the worldwide illicit drugs market. New psychoactive substances (NPSs) have emerged as “legal alternatives” to the well-known addictive and abusive drugs (cannabis, cocaine, heroin and amphetamines) [ref. 1]. Otherwise known as “designer drugs”, these novel substances are synthesized in order to maintain and/or increase the pharmacological effect of the classic drugs of abuse but remaining outside the legal controls [ref. 2]. Therefore, NPSs represent a public health and regulatory challenge [ref. 3]. The number of NPSs monitored and seized by international organizations, such as the European Monitoring Centre for Drugs and Drug Addiction (EMCDDA) and United Nations on Drugs and Crime (UNODC), demonstrate that the NPS market is heterogeneous, rapid and dynamic [ref. 2,ref. 4]. The NPSs include a wide range of different compounds which can be classified in different categories in order to distinguish their pharmacological effects: stimulant (amphetamines, cathinones, benzofuran, indole and pyrovalerone derivatives), sedatives (synthetic opioids, designed benzodiazepines and Gamma-Hydroxybutyrate), dissociatives (phencyclidine, ketamine, diphenidine and their derivatives), synthetic cannabinoids, and psychedelics (phenethylamines, tryptamines, lysergamides) [ref. 2,ref. 3,ref. 4,ref. 5,ref. 6,ref. 7]. Among these, synthetic cannabinoids and cathinones are most often detected, and are the classes mainly used by young adults (15–35 years old) [ref. 8]. In particular, data collected in the European School Survey Project on Alcohol and Other Drugs (ESPAD) reported that students have used synthetic cannabinoids at least once in their lifetime, and it was highlighted that a higher percentage point of males than females used both synthetic cannabinoids and cathinones [ref. 9]. Moreover, Bachman and colleagues, analyzing the widespread use of synthetic cannabinoids and cathinones among adolescents in the USA, revealed that there is variability at the interindividual level, but especially between the sexes; in fact, boys are more at risk of using these NPSs than girls [ref. 10].
Relying on case reports, NPSs are often involved in polydrug use cases and can induce many and various acute adverse effects [ref. 11,ref. 12]. Symptoms such as tachycardia, restlessness, agitation, bruxism, seizures, hallucinations, psychosis, unconsciousness, respiratory failures, nausea, vomiting and death have been related to the use of synthetic cathinones [ref. 13,ref. 14] and cannabinoids [ref. 15,ref. 16]. In particular, synthetic cathinones have been associated with long-lasting renal failure [ref. 17], fetal death [ref. 18] and overdose cases typified by symptoms such as stroke, cerebral edema, myocardial infarction and subsequent death [ref. 19,ref. 20] On the other hand, dysregulation of the endocannabinoid system (ECS) seems to be involved in cardiovascular function [ref. 21]. In fact, smoking marijuana has been related to myocardial ischemia, coronary thrombosis and vasospasm [ref. 22], and synthetic cannabinoids have shown tachycardia and hypertension effects [ref. 23]. Further studies also suggest the proatherogenic role of CB1 receptor [ref. 24]. Such evidence may be therefore considered as indicating that these synthetic drugs increase the risk of cardiovascular and cerebrovascular events [ref. 25,ref. 26], confirmed by the fact that cases of acute ischemic stroke have been reported in young adults after use of synthetic cannabinoids [ref. 25]. Biopsy findings have moreover confirmed cases of acute tubular necrosis or acute interstitial nephritis in young patients [ref. 27].
In vitro and in vivo studies concerning NPSs were widely present in the literature [ref. 28,ref. 29,ref. 30,ref. 31,ref. 32,ref. 33,ref. 34,ref. 35,ref. 36,ref. 37,ref. 38]. In particular, Lenzi and colleagues have demonstrated the mutagenic capability of synthetic cathinone and cannabinoids [ref. 39,ref. 40], underlying their potential toxicity. However, in vitro tests cannot easily predict pharmacokinetics or penetration into the human brain [ref. 41,ref. 42]. Therefore, animal models have played a key role in preclinical research explaining the neurobiological, psychopathological, behavioral and etiological aspects, and studying drug dependence and acute/chronic abuse [ref. 43]. In fact, many in vivo studies show acute effects imputable to NPS-related intoxication [ref. 44,ref. 45,ref. 46,ref. 47,ref. 48].
Moreover, knowledge of long-term NPS effects is lacking [ref. 9,ref. 49] and illicit drug responses are complex traits because they may be determined by both genetic and environmental factors. Variations in the expression of drug-response-related proteins (such as drug-metabolizing enzymes (DMEs), drug transporters and therapeutic mechanisms, including drug targets and downstream signal molecules) are other important sources of interindividual variability in drug response [ref. 50,ref. 51,ref. 52,ref. 53].
Given this evidence, it may be necessary to understand a possible correlation between the use of NPSs and epigenetic changes. Indeed, data show that epigenetic modifications are linked to changes in development and behavior, but also to genetic disorders and various diseases [ref. 54,ref. 55]. It is well known that widespread epigenetic changes occur across the entire genome [ref. 56]. This potentially reversible “epigenetic” modulation of gene expression occurs through the chemical modification of DNA and histone protein tails [ref. 53], or the specific production of regulatory non-coding RNA (ncRNA) [ref. 57]. In this regard, it has been observed in patients with myocardial infarction that there are 200 differentially methylated cytosine–phosphate–guanine (CpG) sites in the gene locus involved in cardiac function [ref. 58]. In addition, a 100-fold increase was found in the level of circulating miRNAs in the plasma of patients suffering from cardiac events [ref. 59,ref. 60,ref. 61]. Studies concerning cancer prevention and innovative treatments for cancers have led to the label “pharmacoepigenomics” in reference to the emerging field of combined study on genetic variations and epigenetic modifications in drug responses [ref. 62,ref. 63]. This development may expand the scope of pharmacogenomics and better define the role of each factor involved in variable drug responses.
This narrative review aims to find potential predictive markers of organ damage, considering existing knowledge of NPS toxicology and possibly related epigenetic changes, with a focus on synthetic cathinones and cannabinoids. Our previous study suggested that in vivo synthetic stimulants, such as 4,4′-Dimethylaminorex (4,4′-DMAR), involves physiological, neurobehavioral and neurotoxicological effects, as confirmed by immunohistochemical analysis [ref. 64]. In particular, we investigated the effect of 4,4′-DMAR on the expression of specific cerebral cortex markers (oxidative/nitrosative stress, apoptosis and heat shock proteins) commonly known to be indicative of brain damage [ref. 65,ref. 66]. Therefore, considering that nerve cells require a large number of mitochondria to ensure their normal functioning, and that overproduction of ROS following the administration of stimulants may lead to oxidative damage in the mitochondria and consequently neurotoxicity [ref. 67], this could be an excellent starting point to research possible epigenetic markers of NPS-induced physiopathological damage.
Search Strategy
A systematic search was conducted using Pubmed, considering relevant data from the earliest available date up until January 2022. The following combined search terms were considered: “NPS”, “epigenetics modifications”, “DNA-methylation”, “histone modification”, “non-coding RNA”, “heart damage” and “brain damage”. All duplicates were then removed. Any title that was coherent with this narrative review main topic was included in a subsequent screening based on eligibility of abstract and full text. Studies were considered appropriate if they included the following: (1) animal model (mouse and/or rat); (2) synthetic cannabinoids and cathinones or analogous and traditional drugs of abuse; (3) connection between exposure to the substance and subsequent epigenetic changes. A total of 218 articles were included in the bibliography section (N in vivo, N in vitro). A manual search of the reference list of selected articles was also conducted.
2. Epigenetic Changes
In the last two decades, epigenetics was initially defined as “the study of mitotically and/or meiotically heritable changes in gene function that cannot be explained by changes in the DNA sequence” [ref. 68,ref. 69]. Currently, it is defined as “the study of any phenotypic diversity that is not related to genotypic differences that can be transmitted through cell divisions but are not directly traceable to the DNA sequence” [ref. 70,ref. 71].
Stretched out linearly, the negatively charged DNA hosted in the nucleus of a mammalian cell is calculated to be approximately 2 m long [ref. 72]. In order to compress all this genetic material into ~10 µm diameter of nucleus, the DNA must wrap tightly around positively charged histone proteins to form the nucleosome, the founding unit of the DNA packaging material called chromatin [ref. 73]. Chromatin structure ensures access to genetic information even in the presence of a highly protective environment, which under physiological conditions makes DNA essentially inaccessible. Nevertheless, evolution has developed enzymatic systems that make it possible to modulate chromatin, and consequently nucleosomes, to ensure “regulated” access to DNA by competent systems [ref. 74].
Tightly packed DNA is considered heterochromatin, while loosely packed DNA is considered euchromatin [ref. 75]. As opposed to euchromatin, which is freely accessible to the transcriptional machinery and actively transcribed, heterochromatin is generally transcriptionally inactive, although a recent study investigated its potential activity [ref. 76]. Changes in the components that make up chromatin (Table 1) itself make it more or less accessible to transcription, constituting epigenetic inheritance (Figure 1) [ref. 77].
Table 1: Summary of all epigenetic changes.
| Epigenetic Mechanisms | Epigenetic Modification | Effects on Gene Expression |
|---|---|---|
| Histone modification | Addition of an acetyl group (Ac) to the amine group of the lysine residues of histones H2B, H3 and H4; addition of one or more methyl groups (Me) on lysine or arginine residues preferentially found on histone tails. | Activation or repression of gene transcription. |
| DNA methylation | Addition of a methyl (Me) group that occurs preferentially in C- and G-rich genomic region CpG islands. | Gene repression. |
| Non-coding RNA | Different RNAs—not translated into proteins—that influence gene expression at the transcriptional and translational level. | Regulation of gene expression (miRNAs binds mRNAs in the 3’UTR region, which promotes their degradation or prevents their translation in protein). |
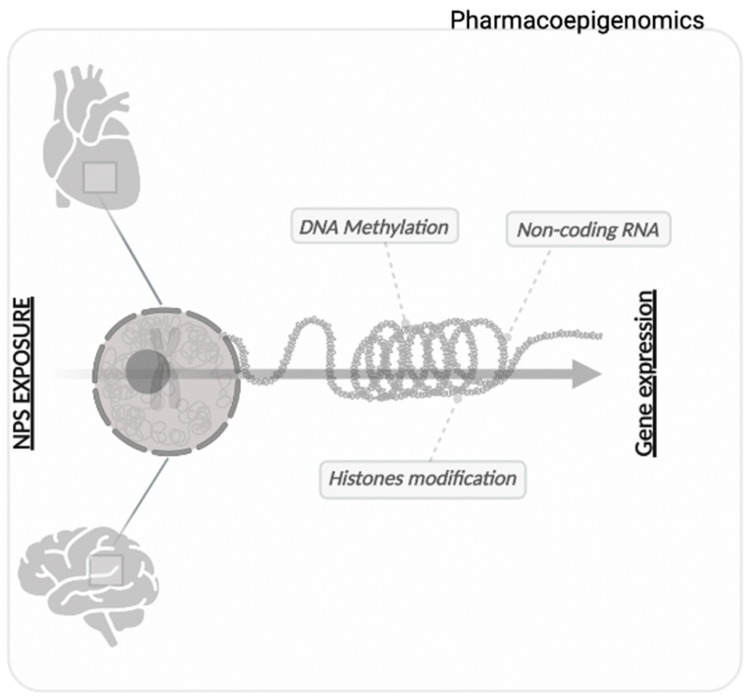
2.1. Histones Modification
Post-translational modifications on the residues of the histone tails of nucleosomes allow the relaxing of chromatin and its condensation, which enables regulation of gene expression, DNA replication, recombination and repair processes [ref. 62]. The first identified histone modifications were acetylation and methylation, followed by ubiquitination and phosphorylation [ref. 78]. All histone modifications (which are directed by enzymatic reactions) exert their action by means of two mechanisms: through a local or remote alteration of the whole chromatin structure, and through a positive or negative modification of the binding to effector molecules [ref. 79]. The functional consequences of histone modifications occur through “reader” proteins that bind to specific modified residues and make transcriptional changes. The enzymes (acetyltransferases (HAT) catalyse acetylation and deacetylases (HDAC) deacetylation, while methyltransferases (HMT) catalyze methylation and demethylases (HDM) demethylation])involved in these various covalent histone modifications can be labeled as “writers” and “erasers” [ref. 79]. Among this, the best-known epigenetic mechanism of histone alteration in the brain is the post-translational covalent modification of the N-terminal tails of histones in different amino acid residues [ref. 80].
The methods used to study histone modifications are mainly based on systems using monoclonal and polyclonal antibodies for target recognition, such as chromatin immunoprecipitation [ref. 81] and proteomic methods, have only recently been developed [ref. 74].
2.2. DNA Methylation
DNA methylation is a process that induces gene silencing and heterochromatin formation [ref. 82]. It is involved in the regulation of gene expression, genomic imprinting, X-chromosome inactivation and silencing of centromeric regions [ref. 83,ref. 84].
The most common DNA modification is the covalent transfer of the methyl group (-CH3) to cytosine located within the CpG island, long over 500 base pairs (bp) [ref. 85,ref. 86]. This produces 5-methylCytosine (5-mC) in the gene promoter region [ref. 87,ref. 88]. The level of methylation of the CpG islands of the regulatory region of a gene is associated with the transcription levels of that gene [ref. 89]. Hypomethylation typically enhances gene expression [ref. 90], while hypermethylation is associated with gene silencing (as opposed to histone methylation) [ref. 91]. There is of course also genetic variability between individuals in the density of CpG sites, which influences potential methylation levels and thus affects the regulatory activities of reference genes [ref. 91].
Compared with readily reversible histone tail modifications, DNA methylation is considered a more stable epigenetic change, altering the chromatin structure from an opened—transcriptionally active—to closed—transcriptionally inactive—state [ref. 92].
Historically, methods for studying DNA methylation can be specific or non-specific. In particular, specific methods provided a global assessment of methylation levels in cells. On the other hand, non-specific methods make a qualitative assessment of changes occurring in a given DNA segment in different cells after treatment with bioactive molecules [ref. 93]. Such detection methods are based on three principles: DNA digestion with restriction enzymes, use of anti-methylcytosine antibodies or methyl-binding domain proteins to enrich methylated genomic DNA fragments and DNA conversion by bisulfite treatment and sequencing [ref. 94].
In recent years, DNA methylation analysis has been increasingly available for their potential use in biomedical research, and has been performed through genome-wide and high-throughput methods [ref. 84]. These advances have significantly accelerated epigenomic research and opened up new perspectives, in particular through the development and application of massively parallel sequencing technologies [ref. 95].
2.3. Non-Coding RNA
Non-coding RNAs (ncRNAs) have been defined as RNAs having information and functions but not having the ability to encode proteins [ref. 96], and they have been shown to play important roles in the regulation of cellular functions [ref. 97]. NcRNAs are epigenetic markers for prognosis, diagnosis and treatment detectable in both tissues and biological fluids that can influence gene expression [ref. 98]. Generally, ncRNAs can be divided into housekeeping or regulatory types [ref. 99]. Among the housekeeping transfer RNA (tRNA), rRNA and small nuclear RNA (snRNA) have been identified. Regulatory ncRNAs include various RNA types distinguishable in long non-coding RNA (lncRNA) and small non-coding RNA (sncRNAs) [ref. 100]. LncRNAs have been placed into six different categories relative to their proximity to the protein-coding genes, and categorized as sense or antisense [ref. 99]. Moreover, an increasing number of lncRNAs have been shown to have crucial roles in several biological processes, such as X-chromosome inactivation and imprinting [ref. 101,ref. 102,ref. 103], maintenance of nuclear architecture [ref. 104], epigenetic control of gene expression and promoter-specific gene regulation [ref. 105]. On the other hand, sncRNAs include microRNA (miRNA), P-element-induced Wimpy testis (PIWI)-interacting RNAs (piRNAs) and short interfering RNA (siRNA) [ref. 106]. Furthermore, circular RNA (circRNA) has been labeled as a special class of non-coding RNA [ref. 107].
The analysis techniques for these sequences are similar to those conventionally used to study other RNAs. They are techniques based on quantitative PCR, sequencing and bioinformatic analysis [ref. 108].
3. Addiction and Epigenetic Mechanisms Related to Drugs of Abuse
Addiction is described as compulsive use, seeking and craving of drugs, and seems to be related to pathological neurobiological changes in neural processes that normally serve reward-related learning [ref. 109].
Furthermore, early life experiences and environmental factors together with genetic susceptibility result in addiction development [ref. 110]. Drug abuse is characterized by two different stages [ref. 111]. In the initial period, after an occasional intake of drugs, the dopaminergic system in NAc triggers pleasurable feelings that results in the uncontrolled pursuit and use of such substances [ref. 112]. In the second stage, different clinical processes occur, which are related to more complex and long-lasting mechanisms implicating alterations in other signal neurotransmitters [ref. 113] (e.g., outcomes on glutamate receptors after exposure to methamphetamine; Table 2 [ref. 114]). Recent studies suggest that different drugs of abuse induce epigenetic changes [ref. 115,ref. 116,ref. 117] in gene expression and post-transcriptional regulation (see Table 2 [ref. 118,ref. 119,ref. 120,ref. 121]). Indeed, the role of microRNAs in drug addiction and neuroplasticity [ref. 115,ref. 116,ref. 117] due to their ability to regulate gene expression has been widely demonstrated [ref. 122,ref. 123]. This suggests that different epigenetic changes are potentially related to drug addiction [ref. 124,ref. 125,ref. 126].
Table 2: Summary of information obtained from the available literature about the main epigenetic changes observed in rodents after intake of traditional stimulants. We report: the substances, the effects found after epigenetic modification, information on the animal model used (by specifying genotype, gender and age), and which detection method is the most appropriate to identify the previously highlighted epigenetic modification.
| Substance | Target | Epigenetic Modification | Effect | Animal Information | Tissue/Cell Type | DNA Methylation Method | References | ||
|---|---|---|---|---|---|---|---|---|---|
| Genotype | Gender | Age | |||||||
| Methamphetamine | GluA1 | Hypoacetylation H4 | Downregulation | Sprague–Dawley rats | Male | Adult | Striatum | Immunoblot/ChIP/Antibody | [ref. 114] |
| GluA2 | |||||||||
| Cocaine | fosB | Increased CBP acetylation H4 | Upregulation | C57BL/6J | Crossed C57BL6J mutant males with BALBc females to generate the F1 hybrids | Adult | Striatum | ChIP/qPCR/Immunoblotting | [ref. 118] |
| BALBc mice | |||||||||
| Cocaine | BDNF | Increased | Upregulation | Sprague–Dawley rats | Male | Adult 10–14 weeks old | Striatum | ChIP/qPCR/RT-PCR/immunohistochemistry | [ref. 119] |
| Cdk5 | Acetylation H4 | Bl6/C57 mice | |||||||
| Methamphetamine | 5 of IEGs | Increased DNA methylation | Downregulation | CD-1 mice | Male | Adult | PFC | RT2 Profiler PCR/qPCR/Pyrosequencing | [ref. 120] |
| Grm1 | Hippocampus | ||||||||
| Methamphetamine | FosB | Increased acetylation H4 | Upregulation | C57BL/6 mice | Male | Adult | Striatum | ChIP/Western Blot | [ref. 121] |
| Increase CREB phosphorylation | |||||||||
| Cocaine | N/A | DNMT3A | Downregulation | C57BL/6J mice and Long Evans rats | Male | Adult | NAc | ChIP promoter analysis/Global DNA methylation analysis | [ref. 126] |
| Increased DNA methylation | |||||||||
| Cocaine | 55 genes | TET1 | Downregulation Alternative splicing | C57BL/6J mice | Male | Adult 8–10 weeks old | NAc | qPCR/western blotting/immunohistochemistry/stereotaxic viral manipulations/ChIP-seq/RNA-seq | [ref. 129] |
| Increased DNA methylation | |||||||||
| Cocaine | PP1c | Increased DNA methylation DNMT3A and DNMT3B | Downregulation and Upregulation of FosB | Mice C57BL/6 | Male | Adult | NAc | qPCR/ChIP/MeDIP/western Blotting | [ref. 140] |
| Cocaine | Cdkl5 | Increased DNA methylation (MeCP2) | Downregulation | Wistar rats | Male | Adult 8–9 weeks old | NAc | RT-PCR—qPCR/Bisulfite sequencing/Chromatin immunoprecipitation/Immunohistochemistry | [ref. 141] |
| Cocaine | cFos, FosB, BDNF e Cdk5 | Increased Acetylation H3 and H4 | Upregulation | Sprague–Dawley rats | Male | Adult | Striatum | ChIP | [ref. 119] |
| Cocaine | ΔΦοσ | Increased Acetylation H3 and H4 | Upregulation: sirt1 sirt2 | C57BL/6 mice | Male | 10–12 weeks | NAc | ChIP/Array | [ref. 143] |
| CREB | |||||||||
| Cocaine | 172 genes | Decrease Histone Deacetylation HDAC5 | Upregulation | Mice C57BL/6 | N/A | Not adult | NAc | Western Blotting/Immunohistochemistry/ChIP/qPCR/RT-PCR/Microarrays | [ref. 142] |
| Cocaine | N/A | DNMT3B | Upregulation | C57/BL6 mice | Male | Adult | PFC | Global DNA methylation analysis, qPCR, Western blotting | [ref. 145] |
| Decrease DNA methylation | |||||||||
| Cocaine | 57 genes | DNMT3A Increased DNA methylation | Up-/downregulation | C57BL/6 mice | Male | Adult aged 5–6 months | NAc | MicroArray Illumina/qPCR/MeDIP | [ref. 146] |
| Cocaine | PKCε | Increased DNA methylation | Downregulation | Sprague–Dawley rats | Pregnant | Fetal (21 days) | Heart | Quantitative Methylation-Specific PCR/ChIP | [ref. 147] |
Over the years, attempts have been made to define the mechanisms by which exposure to a drug of abuse alters mRNA levels through chromatin regulation mechanisms or by activating/inactivating additional genes for altered expression after a period of abstinence [ref. 127].
However, previous studies have pointed out that traditional drugs of abuse, such as cannabis (Table 2 [ref. 128]) and cocaine (Table 3 [ref. 129]), may influence gene expression by inducing epigenetic changes. Indeed, the role of the endocannabinoid system (ES) in reward-related learning and addiction was already investigated [ref. 130,ref. 131]. Specifically, it has been shown that the use of cannabis may interfere with endocannabinoid signalling and affect the neural pathways that regulate reward-related learning and cognition [ref. 131], thus strengthening the hypothesis that synthetic cannabinoid may affect the same neural pathways. Therefore, the ES is possibly sensitive to environmental epigenetic cues. In fact, there is evidence of genetically related variations in reactivity to stress and negative emotional states, and drug craving as a result of an altered endocannabinoid system feature [ref. 132]. In particular, the endocannabinoid system undergoes dynamic changes during adolescence, which is when anxiety disorders often emerge [ref. 132]. This could possibly explain data showing that cannabis use among adolescents has been associated with an increased risk of subsequent cocaine intake [ref. 133], and abuse of synthetic cannabinoids has been associated with multiple drug use, including stimulants [ref. 134], and induces a behavioral change in exposed animals (see Table 3, [ref. 135]). To confirm this, a recent study has shown that cannabinoid exposure in adolescent can change the behavioral and epigenetic response to cocaine in rodents [ref. 136,ref. 137]. In addition, attention was paid to the proenkephalin opioid neuropeptide (Penk) gene in NAc that directly regulates heroin addiction. Adolescent rats exposed to THC overexpressed Penk and reported reduced methylation of histone H3 lysine 9 in NAc (Table 3, [ref. 138]).
Table 3: Summarize of information obtained from the available literature about the main epigenetic changes observed in rodents after intake of traditional and new synthetic cannabinoids. We reported: the substances, the effects found after epigenetic modification, information on the animal model used (by specifying genotype, gender and age), and which detection method is the most appropriate to identify the previously highlighted epigenetic modification.
| Substance | Target | Epigenetic Modification | Effect | Animal Information | Tissue/Cell Type | DNA Methylation Method | References | ||
|---|---|---|---|---|---|---|---|---|---|
| Genotype | Gender | Age | |||||||
| Δ⁹–THC | Appbp2 CD27 | LncRNA | Downregulation of miR-17/92 cluster and miR-374b/421 cluster Up-regulation of miR-146 | C57BL/6J mice | Female | 6–7 weeks old | Lymph node cells CD4+ | RNA-seq | [ref. 128] |
| WIN55212.2 | Rgs7 | Hypermethylation | Downregulation | C57Bl6/J mice | Male | Adolescent aged 4 weeks | Hippocampal CA regions | qPCR/RT–PCR | [ref. 135] |
| Δ⁹–THC | Penk | Decrease H3K9 Methylation | Upregulation | Long Evans rats | Male | Adolescents 21-day-old | NAc | ChIP | [ref. 138] |
| Δ⁹–THC | N/A | H3K9me2 H3K9me3 H3K9ac H3K14ac | Both | Sprague–Dawley rats | Female | Adolescent (35–45 postnatal day) and adult (75–85 postnatal day) | Hippo NAc Amy | Western Blot | [ref. 148] |
| Δ⁹–THC | 177 genes | DNA Methylation | Both | Sprague–Dawley rats | Male | Nine-week-old, sexually mature | Semen | Bisulfite Sequencing/Pyrosequencing | [ref. 149] |
| JWH133 | CB2 | Upregulation of H3K4me3 Downregulating H3K9me2 | Upregulation Prdm9 c-Kit Stra8 | Swiss CD-1 mice | Male | Seven-day-old | SPG cells (Spermatogonia) | qPCR/RT–PCR/ChIP/Western Blot | [ref. 150] |
| Δ⁹–THC | Drd2 | Increased 2meH3K9 | Decrease 3meH3K4 | Long Evans rat | Male and female | Adult | NAc | In Situ Hybridization Histochemistr/ChIP/ | [ref. 151] |
| and mRNA level | |||||||||
| Δ⁹–THC | 1027 | DNA Methylation | Alterated | Long Evan rats | Male and female | Adolescent | NAc | Enhanced Reduced Representation Bisulfite Sequencing | [ref. 152] |
| genes | |||||||||
| Δ⁹–THC | Genes associated with plasticity | Increase Suv39H1 and H3K9me3 | Downregulation | Sprague-Dawley rats | Female | Adolescent and adult | PFC | RT2 Profile PCR Array/ChIP | [ref. 153] |
| HU-210 | Dlk1-Dio3-imprinted domain | Differential miRNA expression | Both | Wistar rats | FemaleMale | PregnantOffspring | Left and right hemispheres of the entorhinal cortex (EC) | qPCR/RT–PCR | [ref. 154] |
Furthermore, studies on exposure to psychostimulants, such as cocaine, have revealed the occurrence of epigenetic changes previously attributed to psychiatric disorders (drug addiction, depression or Rett syndrome) since the 2000s [ref. 61,ref. 62,ref. 139]; (Table 2, [ref. 140,ref. 141]). In particular, chronic cocaine exposure has been shown to recruit histone acetyltransferases and regulate histone acetylation/deacetylation in NAc [ref. 142]; (Table 2, [ref. 114,ref. 115,ref. 116,ref. 117,ref. 118,ref. 119,ref. 120,ref. 121,ref. 122,ref. 123,ref. 124,ref. 125,ref. 126,ref. 127,ref. 128,ref. 129,ref. 130,ref. 131,ref. 132,ref. 133,ref. 134,ref. 135,ref. 136,ref. 137,ref. 138,ref. 139,ref. 140,ref. 141,ref. 142,ref. 143]). The same study has shown that the main effect of chronic cocaine exposure is gene activation, with more genes showing H3 or H4 hyperacetylation than hypoacetylation (Table 2, [ref. 143]). Moreover, changes at the epigenetic level occur in mice repeatedly administered with 3,4-methylenedioxymethamphetamine (MDMA), and this could be related to the cardiotoxicity induced by such drugs of abuse [ref. 144]. The knowledge obtained in the context of abuse (e.g., use of methionine as a therapeutic target for cocaine addiction (Table 2, [ref. 145,ref. 146]) and pathological information could be used to investigate markers that define therapeutic targets or intoxication antidotes to avoid permanent damage to the organism.
4. Overview of Epigenetic Factors and Their Clinical-Toxicological Use
Recent studies have shown that ageing is a complex multifactorial mechanism shared by all living organisms and is expressed by the gradual decline of physiological functions [ref. 155] and influenced by various genetic, lifestyle and environmental factors [ref. 156]. Therefore, it is important to differentiate chronological age from biological age [ref. 157]. Individuals of similar chronological age may show very different susceptibilities to age-related diseases and death, which presumably reflects differences in biological ageing processes [ref. 155]. In particular, DNA methylation patterns change in the process of aging and contribute to age-related disease development [ref. 158]. Diverse epigenetic changes occur during the lifetimes of mammals [ref. 159]. In particular, studies have shown a correlation between age and DNA methylation at the level of single CpG sites [ref. 160,ref. 161]. On this basis, predictive models have been built in order to quantify age-related phenotypes or outcomes, such as diseases (cardiovascular and neurodegenerative conditions) or mortality [ref. 161,ref. 162]; such models are labelled as “epigenetic clocks”. Epigenetic age deceleration has been associated with longevity [ref. 163], strengthening the above-mentioned assumption. Taken together, these notions suggest that chronic use of certain drugs could lead to neurotoxicity effects proportionate to increasing subject age—being amplified in adult and elderly animals compared to young ones [ref. 164,ref. 165,ref. 166]. In this context, differences have been observed in terms of epigenetic modifications that interact differentially in adult and adolescent subjects under the same conditions of exposure to substances of abuse (Table 3, [ref. 148]).
Furthermore, a possible gender-related difference among such epigenetic changes has been studied. Specifically, a recent study has shown sex-specific variations in the DNA methylation patterns of two distinct genes (FIGN and PRR4) [ref. 167]. These sex disparities have been previously confirmed by studies on the mouse hippocampus and the human frontal cortex [ref. 168]. According to Global Health Observatory (GHO) data, nowadays global life expectancy at birth is 76 years for females and 71 years for males [ref. 169]. In agreement with this evidence, data have shown that male’ mortality rates due to cardiovascular, cancer and Parkinson’s disease are higher than those of females at a given age. On the other hand, females show an increased risk of Alzheimer’s and autoimmune diseases [ref. 167].
Furthermore, gender can influence subjective effects and pharmacotoxicological responses to drugs [ref. 170] and can also present differences in the activity of specific enzymes involved in drug metabolism [ref. 171]. In particular, CYP450 family enzymes play a crucial role [ref. 172]. The isoenzymes CYP2D6, CYP3A4, CYP1A1, CYP1A2 and CYP2C19 are involved in the metabolism of most psychiatric drugs and many other drugs commonly used and prescribed in daily practice (beta-blockers, opioid analgesics, anticonvulsants, antihistamines, cortisones and the macrolide antibiotics) [ref. 172,ref. 173]. Studies have also confirmed the involvement of the above-mentioned enzymes in the metabolism of synthetic cannabinoids [ref. 174,ref. 175] and cathinones [ref. 176]. For example, the cytochrome P450 3A4 is abundantly expressed in the liver, but its activity is higher in women than in men [ref. 177]. This could be due to female-specific issues, such as pregnancy, menopause, oral contraceptive use and menstruation, suggesting a possible role played by sexual hormone levels [ref. 177]. Other features, such as body weight and the amount of adipose tissue, depending on the subjects themselves, should be considered [ref. 171].
Compelling studies have shown that NPSs, such as synthetic cannabinoids and cathinone, can induce gender-related effects on animals [ref. 170]. In certain respects, the effects of 3,4-Methylenedioxypyrovalerone (MDPV) on cardiovascular parameters are deeper and long-lasting in males than females [ref. 178]. However, Fattore and colleagues have reported that females are more susceptible to the reward-seeking and anxiety effects respectively induced by synthetic cannabinoids and cathinones than males [ref. 179]. Different studies have also investigated effects of prenatal exposure to cocaine [ref. 147,ref. 180,ref. 181,ref. 182,ref. 183,ref. 184]. Specifically, it has been demonstrated that cocaine can induce sex-dependent epigenetic changes during the gestational period [ref. 180] and may increase the heart’s vulnerability to ischemic damage during adulthood [ref. 183]. Moreover, mice prenatal cocaine exposure has been shown to be related to alteration of the genes involved in Wnt and the cadherin system [ref. 184]. This suggests that prenatal exposure to cocaine leads to an increased susceptibility of the heart to ischaemic damage in the adult offspring (F1), due to decreased PKC gene expression. Thus, programming of PKC gene expression patterns in the heart already occurs in utero (F0) (Table 2, [ref. 147]).
These considerations reflect the danger of synthetic cannabinoids, precisely because Δ9-THC can cross the placental barrier and come into contact with the fetus [ref. 185]. This can cause defective development of the child’s brain and produce neurobehavioral toxicity [ref. 186]. In addition, studies on the effects of synthetic cannabinoids such as JWH-018 report neurobehavioral alterations and in particular evidence of substance dependence [ref. 187].
Epigenetic Inheritance
Multigenerational epigenetic inheritance is defined as the germline-mediated heritage of epigenetic information between generations after direct environmental influences, which results in phenotypic variation in the offspring [ref. 188,ref. 189]. However, some literature considers transgenerational epigenetic inheritance in the absence of continued direct environmental influences [ref. 190]. Specifically, exposure to a variety of environmental factors in F0 of both genders before gestation may have a direct impact on the germ cells, which will affect the F1 generation. Therefore, the phenotypes found in F0 and F1 animals are considered multigenerational. In contrast, only those traits that persist in the F2 generation and beyond are considered examples of transgenerational inheritance (Figure 2) [ref. 190]. This phenomenon has been investigated in detail with regard to X-chromosome inactivation [ref. 191] and exposure to drugs of abuse [ref. 27]. Drug exposure during embryonic development simultaneously exposes the developing fetus (F1) and germ cells (F2) to the effects of the drug. Following the same pattern, drug-using parents expose their germ cells and consequently the F1 generation [ref. 192]. Furthermore, it was found that perinatal exposure to CBD induced changes in DNA methylation levels in F1 generation associated with increased anxiety and improved memory behavior in a sex-specific manner [ref. 193]. These changes have been associated with exposure to environmental factors that could also induce epigenetic changes in sperm or ova (Table 3 [ref. 125,ref. 191,ref. 192]). Previous studies have shown that prenatal THC exposure can alter cognitive function, emotional reactivity [ref. 151] and responses to drugs of abuse, such as methamphetamine [ref. 194] and opiates (Table 3, [ref. 193]). Direct exposure to drugs of abuse in the embryo (F3) and/or in the parents (F2) has been also proven to cause transgenerationally inherited alterations in DNA methylation (Figure 2) [ref. 195]. In support of this, the NAc epigenome was analyzed in a Long Evans rat model, identifying 1027 differentially methylated regions associated with parental exposure to THC in F1 adults (Table 3, [ref. 152]). These studies provide new information on drug-related intergenerational epigenetic effects and are a starting point for the study of neurobiological effects underlying drug abuse vulnerability.
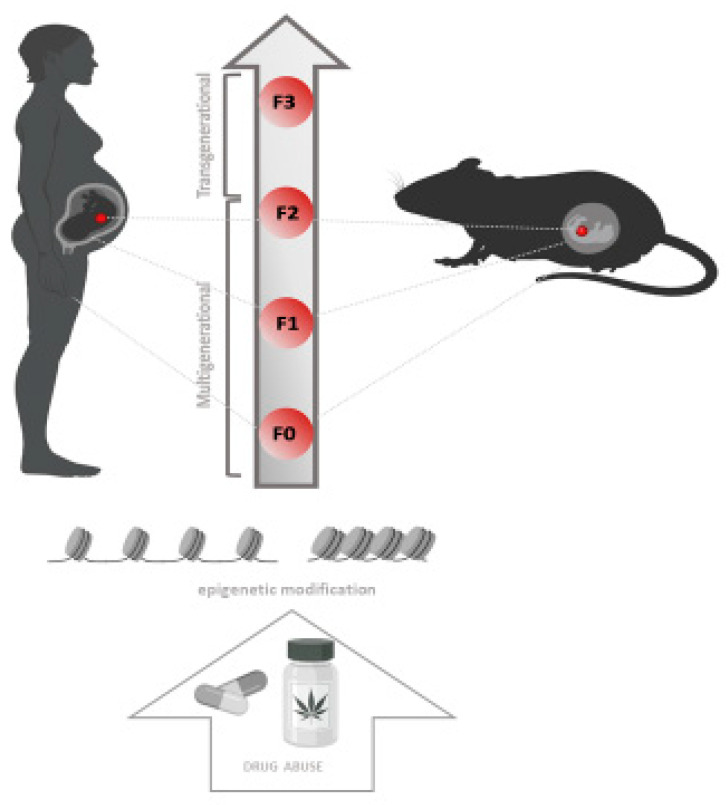
Concerning the use of synthetic cannabinoids, scientific evidence shows that there is a predisposition to neurodevelopmental disorders (i.e., schizophrenia and autism spectrum disorders) in the offspring of women exposed to this class of substance during pregnancy [ref. 196]; (Table 3, [ref. 135,ref. 153,ref. 154]). Moreover, a recent study has connected cannabis exposure during the perinatal period to epigenetic changes in animal models [ref. 197]. Epigenetic changes possibly induced by NPS abuse have not yet been investigated. Despite this, recent study has revealed that THJ-2201 and 5F-PB22 affected neuronal differentiation, and this could result in neurodevelopmental disorders if synthetic cannabinoids use occurs before or during pregnancy [ref. 198].
5. New Perspectives in the Forensic Field
From the perspective of forensic medicine, several reports concerning the effects of NPSs can be found in the literature [ref. 32,ref. 199]. Indeed, there is no doubt that the availability and consumption of these substances has become increasingly important in recent decades, with the number of deaths attributed to NPSs growing year by year [ref. 200].
However, detailed data on the number of substances marketed and consumed, and on the extent and type of organ damage caused by them, are still insufficient [ref. 201]. The advent of NPSs has called into question traditional methods of drug detection, monitoring, surveillance, and control [ref. 200]; for instance, in the clinical setting, rapid immunoassay screening tests typically cannot detect NPSs [ref. 202]. Similarly, in the domain of forensic toxicology, NPSs may be difficult to distinguish from more well-known illicit substances and other psychotropic chemicals due to a lack of available data on their pharmacology, toxicology, and health impacts [ref. 203]. Indeed, NPS research is typically overlooked in regular screens, and traditional approaches might not be as effective for detecting NPSs [ref. 200]. Clinicians are thus required to be familiar with the many classes of these compounds, as well as their effects, in order to apply this information to the specific case and carry out focused toxicological assessments [ref. 204]. Given the numerous challenges associated with NPS detection and characterization, as well as the relative lack of pertinent data in the literature, it is obvious that identifying as many methods as possible to obtain information on these specific substances of abuse is becoming increasingly important. Forensic epigenetics is a relatively new field of research with a wide range of potential applications. In forensic medicine, DNA methylation is typically chosen over other epigenetic alterations because of its in vitro stability and great sensitivity in terms of the amount of DNA required [ref. 205]. Based on the notion that environmental stimuli can cause individual epigenomic variations [ref. 205,ref. 206], forensic investigations apply DNA methylation to the identification of tissues and cells [ref. 207], to the determination of both sex and age of individuals [ref. 208,ref. 209], and to the differentiation between monozygotic twins [ref. 210]. Furthermore, DNA methylation can disclose information about a person’s socioeconomic status, diet, physical activity, alcohol use [ref. 211], smoking status and drug use [ref. 212], and thus may aid in the discovery of specific markers that can provide more detailed information about the characteristics and effects of NPSs.
Hence, in order to identify various effective methods of prevention and treatment, future preclinical, clinical and forensic research on synthetic cannabinoids and cathinones could benefit from the support of epigenetics. Moreover, given the complexity of the NPS issue, there is no doubt that combining different methods of investigation with integrated pharmacological and psychological strategies aimed at treating intoxication symptoms will yield significant benefits. Potential investigation methods for this purpose include forensic investigation, pre-clinical in vitro research and in vivo models of mice and other animal species to provide important tools for analyzing human drug metabolism, as well as clinical-level investigations. In addition, the modification of treatment regimens for the more well-known drugs of abuse can help address the rising incidence of intoxication reported following the use of synthetic cathinones and cannabinoids.
6. Conclusions
This review aims to provide an overview of epigenetic modulation and possible hyper- or hypo-expression of epigenetic markers of tissue damage. In fact, the epigenetic structure can be studied in animal models throughout the life cycle of an organism, from conception to adulthood and old age [ref. 213]. An ideal model needs to be accessible at all stages of prenatal and postnatal life, and to have a reasonable time span to enable assessment of all possible outcomes of an experiment [ref. 214]. Thus, animal models allow the investigation of the probable interplay between epigenetic changes and effects caused by the intake of synthetic cannabinoids and cathinone, focusing on gender related differences. Given the great potential toxicological and forensic value of understanding epigenetic changes induced by exposure to drugs of abuse, the overall strength of the present narrative review is the suggestion of a translational evaluation of the pharmacotoxicological effects of NPSs widely reported by preclinical and clinical literature. However, this aspect also represents a weakness, because of the great variety of environmental and non-environmental factors that can influence epigenomic changes.
References
- A. Roman-Urrestarazu, R. Robertson, J. Yang. European Monitoring Centre for Drugs and Drug Addiction, European drug report 2018: Trends and developments. Bmj, 2018
- D. Luethi, M.E. Liechti. Designer drugs: Mechanism of action and adverse effects. Arch. Toxicol., 2020. [DOI | PubMed]
- 3. EMCDDA European Monitoring Centre for Drugs and Drug Addiction European Drug Report 2020: Trends and Devel-OpmentsPublications Office of the European UnionLisbon, Portugal2020
- 4. United Nations Office on Drugs and Labor World Drug Report 2020United NationsVienna, Austria2021
- L.D. Simmler, A. Rickli, Y. Schramm, M.C. Hoener, M.E. Liechti. Pharmacological profiles of aminoindanes, piperazines, and pipradrol derivatives. Biochem. Pharmacol., 2014. [DOI | PubMed]
- L. Simmler, T. Buser, M. Donzelli, Y. Schramm, L.-H. Dieu, J. Huwyler, S. Chaboz, M. Hoener, M. Liechti. Pharmacological characterization of designer cathinones in vitro Br. J. Pharmacol., 2013. [DOI]
- S. Bilel, J.A. Neto, R. Arfè, M. Tirri, R.M. Gaudio, A. Fantinati, T. Bernardi, F. Boccuto, B. Marchetti, G. Corli. In vitro and in vivo pharmaco-dynamic study of the novel fentanyl derivatives: Acrylfentanyl, Ocfentanyl and Furanylfentanyl. Neuropharmacology, 2022. [DOI | PubMed]
- 8. EMCDDA European Monitoring Centre for Drugs and Drug Addiction European Drug Report 2021: Trends and Devel-OpmentsPublications Office of the European UnionLisbon, Portugal2021
- ESPAD Report 2019
- M.E. Patrick, P. O’Malley, D.D. Kloska, J. Schulenberg, L. Johnston, R. Miech, J.G. Bachman. Novel psychoactive substance use by US adolescents: Characteristics associated with use of synthetic cannabinoids and synthetic cathinones. Drug Alcohol Rev., 2015. [DOI | PubMed]
- S.L. Hill, M. Dunn, C. Cano, S.J. Harnor, I.R. Hardcastle, J. Grundlingh, P.I. Dargan, D.M. Wood, S. Tucker, T. Bartram. Human Toxicity Caused by Indole and Indazole Carboxylate Synthetic Cannabinoid Receptor Agonists: From Horizon Scanning to Notification. Clin. Chem., 2018. [DOI | PubMed]
- J. Klavž, M. Gorenjak, M. Marinšek. Suicide attempt with a mix of synthetic cannabinoids and synthetic cathinones: Case report of non-fatal intoxication with AB-CHMINACA, AB-FUBINACA, alpha-PHP, alpha-PVP and 4-CMC. Forensic Sci. Int., 2016. [DOI | PubMed]
- Y. Fujita, T. Mita, K. Usui, Y. Kamijo, S. Kikuchi, M. Onodera, Y. Fujino, Y. Inoue. Toxicokinetics of the Synthetic Cathinone α-Pyrrolidinohexanophenone. J. Anal. Toxicol., 2018. [DOI | PubMed]
- F. Dragogna, L. Oldani, M. Buoli, A.C. Altamura. A case of severe psychosis induced by novel recreational drugs. F1000Research, 2014. [DOI | PubMed]
- V. Angerer, S. Jacobi, F. Franz, V. Auwärter, J. Pietsch. Three fatalities associated with the synthetic cannabinoids 5F-ADB, 5F-PB-22, and AB-CHMINACA. Forensic Sci. Int., 2017. [DOI | PubMed]
- L.J. Schep, R. Slaughter, S. Hudson, R. Place, M. Watts. Delayed seizure-like activity following analytically confirmed use of previously unreported synthetic cannabinoid analogues. Hum. Exp. Toxicol., 2015. [DOI | PubMed]
- M. Levine, R. Levitan, A. Skolnik. Compartment Syndrome After “Bath Salts” Use: A Case Series. Ann. Emerg. Med., 2013. [DOI | PubMed]
- P. Adamowicz, P. Hydzik. Fetal death associated with the use of 3,4-MDPHP and α-PHP. Clin. Toxicol., 2019. [DOI]
- P.J. Nicholson, M.J. Quinn, J.D. Dodd. Headshop heartache: Acute mephedrone ‘meow’ myocarditis. Heart, 2010. [DOI | PubMed]
- D. James, R.D. Adams, R. Spears, G. Cooper, D.J. Lupton, J.P. Thompson, S.H.L. Thomas. Clinical characteristics of mephedrone toxicity reported to the U.K. National Poisons Information Service. Emerg. Med. J., 2011. [DOI | PubMed]
- M.S. Castaneto, D.A. Gorelick, N.A. Desrosiers, R.L. Hartman, S. Pirard, M.A. Huestis. Synthetic cannabinoids: Epidemiology, pharmacodynamics, and clinical implications. Drug Alcohol Depend., 2014. [DOI | PubMed]
- S.E. O’Sullivan. Endocannabinoids and the Cardiovascular System in Health and Disease. Handb. Exp. Pharmacol., 2015. [DOI | PubMed]
- P. Kumar, S. Zaleta, S. Miller. Chest pain, troponin rise, and ST-elevation in an adolescent boy following the use of the synthetic cannabis product K2. Ann. Pediatr. Cardiol., 2016. [DOI | PubMed]
- P. Pacher. Cannabinoid CB 1 Receptor Antagonists for Atherosclerosis and Cardiometabolic Disorders. Arter. Thromb. Vasc. Biol., 2009. [DOI]
- R. Le Boisselier, J. Alexandre, V. Lelong-Boulouard, D. Debruyne. Focus on cannabinoids and synthetic cannabinoids. Clin. Pharmacol. Ther., 2017. [DOI | PubMed]
- P. Pacher, S. Steffens, G. Haskó, T.H. Schindler, G. Kunos. Cardiovascular effects of marijuana and synthetic cannabinoids: The good, the bad, and the ugly. Nat. Rev. Cardiol., 2018. [DOI | PubMed]
- Acute kidney injury associated with synthetic cannabinoid use—Multiple states, 2012. MMWR Morb. Mortal. Wkly. Rep., 2013. [PubMed]
- C. Camuto, A. Guglielmelli, F. De-Giorgio, X. de la Torre, M. Mazzarino, M. Marti, F. Botrè. In vitro metabolic profile of mexedrone, a mephedrone analog, studied by high- and low-resolution mass spectrometry. Drug Test. Anal., 2022. [DOI | PubMed]
- S. Bilel, M. Tirri, R. Arfè, C. Sturaro, A. Fantinati, V. Cristofori, T. Bernardi, F. Boccuto, M. Cavallo, A. Cavalli. In Vitro and In Vivo Pharmaco-Toxicological Characterization of 1-Cyclohexyl-x-methoxybenzene Derivatives in Mice: Comparison with Tramadol and PCP. Int. J. Mol. Sci., 2021. [DOI | PubMed]
- R. Arfè, S. Bilel, M. Tirri, P. Frisoni, G. Serpelloni, M. Neri, F. Boccuto, T. Bernardi, F. Foti, F. De-Giorgio. Comparison of N-methyl-2-pyrrolidone (NMP) and the “date rape” drug GHB: Behavioral toxicology in the mouse model. Psychopharmacology, 2021. [DOI | PubMed]
- F. Foti, S. Bilel, M. Tirri, R. Arfè, F. Boccuto, T. Bernardi, G. Serpelloni, F. De-Giorgio, M. Marti. Low-normal doses of methiopropamine induce aggressive behaviour in mice. Psychopharmacology, 2021. [DOI | PubMed]
- G. Martinotti, A. Negri, S. Schiavone, C. Montemitro, C. Vannini, G. Baroni, M. Pettorruso, F. de Giorgio, R. Giorgetti, V. Verrastro. Club Drugs: Psychotropic Effects and Psychopathological Characteristics of a Sample of Inpatients. Front. Psychiatry, 2020. [DOI | PubMed]
- C. Camuto, S. Pellegrini, F. De-Giorgio, X. de la Torre, M. Marti, M. Mazzarino, F. Botrè. Urinary excretion profile of methiopropamine in mice following intraperitoneal administration: A liquid chromatography–tandem mass spectrometry investigation. Drug Test. Anal., 2021. [DOI | PubMed]
- A. Miuli, M. Pettorruso, E. Romanelli, G. Stigliano, D. Di Giuda, F. De-Giorgio, G. Martinotti, M. Di Giannantonio. Does DRD2 Taq1A Mediate Aripiprazole-Induced Gambling Disorder? A Pharmacogenetic Hypothesis. Front. Psychiatry, 2020. [DOI | PubMed]
- S. Bilel, N. Azevedo, R. Arfè, M. Tirri, A. Gregori, G. Serpelloni, F. De-Giorgio, P. Frisoni, M. Neri, G. Calò. In vitro and in vivo pharmacological characterization of the synthetic opioid MT-45. Neuropharmacology, 2020. [DOI | PubMed]
- A. Ossato, S. Bilel, A. Gregori, A. Talarico, C. Trapella, R.M. Gaudio, F. De Giorgio, F. Tagliaro, M. Neri, L. Fattore. Neurological, sensorimotor and cardiorespiratory alterations induced by methoxetamine, ketamine and phencyclidine in mice. Neuropharmacology, 2018. [DOI | PubMed]
- A. Ossato, L. Uccelli, S. Bilel, I. Canazza, G. Di Domenico, M. Pasquali, G. Pupillo, M.A. De Luca, A. Boschi, F. Vincenzi. Psychostimulant Effect of the Synthetic Cannabinoid JWH-018 and AKB48: Behavioral, Neurochemical, and Dopamine Transporter Scan Imaging Studies in Mice. Front. Psychiatry, 2017. [DOI | PubMed]
- A. Fantinati, A. Ossato, S. Bianco, I. Canazza, F. de Giorgio, C. Trapella, M. Marti. 1-cyclohexyl-x-methoxybenzene derivatives, novel psychoactive substances seized on the internet market. Synthesis and in vivo pharmacological studies in mice. Hum. Psychopharmacol. Clin. Exp., 2017. [DOI]
- M. Lenzi, V. Cocchi, S. Gasperini, R. Arfè, M. Marti, P. Hrelia. Evaluation of Cytotoxic and Mutagenic Effects of the Synthetic Cathinones Mexedrone, α-PVP and α-PHP. Int. J. Mol. Sci., 2021. [DOI | PubMed]
- M. Lenzi, V. Cocchi, L. Cavazza, S. Bilel, P. Hrelia, M. Marti. Genotoxic Properties of Synthetic Cannabinoids on TK6 Human Cells by Flow Cytometry. Int. J. Mol. Sci., 2020. [DOI | PubMed]
- R. Santos-Toscano, A. Guirguis, C. Davidson. How preclinical studies have influenced novel psychoactive substance legislation in the UK and Europe. Br. J. Clin. Pharmacol., 2020. [DOI | PubMed]
- D. Luethi, M.C. Hoener, M.E. Liechti. Effects of the new psychoactive substances diclofensine, diphenidine, and methoxphenidine on monoaminergic systems. Eur. J. Pharmacol., 2018. [DOI | PubMed]
- A. Belin-Rauscent, D. Belin. Animal Models of Drug Addiction. Addictions: From Pathophysiology to Treatment, 2012. [DOI]
- J.A. Marusich, K.R. Grant, B.E. Blough, J. Wiley. Effects of synthetic cathinones contained in “bath salts” on motor behavior and a functional observational battery in mice. NeuroToxicology, 2012. [DOI | PubMed]
- W.E. Fantegrossi, B.M. Gannon, S.M. Zimmerman, K.C. Rice. In vivo Effects of Abused ‘Bath Salt’ Constituent 3,4-methylenedioxypyrovalerone (MDPV) in Mice: Drug Discrimination, Thermoregulation, and Locomotor Activity. Neuropsychopharmacology, 2013. [DOI | PubMed]
- S. Bilel, M. Tirri, R. Arfè, S. Stopponi, L. Soverchia, R. Ciccocioppo, P. Frisoni, S. Strano-Rossi, C. Miliano, F. De-Giorgio. Pharmacological and Behavioral Effects of the Synthetic Cannabinoid AKB48 in Rats. Front. Neurosci., 2019. [DOI | PubMed]
- S. Bilel, M. Tirri, R. Arfè, A. Ossato, C. Trapella, G. Serpelloni, M. Neri, L. Fattore, M. Marti. Novel halogenated synthetic cannabinoids impair sensorimotor functions in mice. NeuroToxicology, 2020. [DOI | PubMed]
- N. Simola, G. Costa, M.A. De Luca, G. Piras, J. Marongiu, L. Fattore. Neuronal and peripheral damages induced by synthetic psychoactive substances: An update of recent findings from human and animal studies. Neural Regen. Res., 2020. [DOI | PubMed]
- H.R. Sumnall, M. Evans-Brown, J. McVeigh. Social, policy, and public health perspectives on new psychoactive substances. Drug Test. Anal., 2011. [DOI | PubMed]
- K. Westbrook, V. Stearns. Pharmacogenomics of breast cancer therapy: An update. Pharmacol. Ther., 2013. [DOI | PubMed]
- C.L. Miranda Furtado, M.C. Dos Santos Luciano, R. da Silva Santos, G.P. Furtado, M.O. Moraes, C. Pessoa. Epidrugs: Targeting epigenetic marks in cancer treatment. Epigenetics, 2019. [DOI | PubMed]
- C. López-Camarillo, D. Gallardo-Rincón, M.E. Álvarez-Sánchez, L.A. Marchat. Pharmaco-epigenomics: On the Road of Translation Medicine. Adv. Exp. Med. Biol., 2019. [DOI | PubMed]
- A. Ravaei, M. Rubini. Rescuing effect of folates on methotrexate cytotoxicity in human trophoblast cells. Clin. Exp. Rheumatol., 2021. [DOI]
- D.S. Moore. Behavioral epigenetics. WIREs Syst. Biol. Med., 2017. [DOI]
- E.J. Kyzar, J.P. Bohnsack, S.C. Pandey. Current and Future Perspectives of Noncoding RNAs in Brain Function and Neuropsychiatric Disease. Biol. Psychiatry, 2022. [DOI | PubMed]
- R.R. Kanherkar, N. Bhatia-Dey, A.B. Csoka. Epigenetics across the human lifespan. Front. Cell Dev. Biol., 2014. [DOI | PubMed]
- J.K. Kim, M. Samaranayake, S. Pradhan. Epigenetic mechanisms in mammals. Exp., 2009. [DOI | PubMed]
- M. Rask-Andersen, D. Martinsson, M. Ahsan, S. Enroth, W.E. Ek, U. Gyllensten, A. Johansson. Epigenome-wide association study reveals differential DNA methylation in individuals with a history of myocardial infarction. Hum. Mol. Genet., 2016. [DOI | PubMed]
- Y. Devaux, M. Vausort, E. Goretti, P. Nazarov, F. Azuaje, G. Gilson, M.F. Corsten, B. Schroen, M.-L. Lair, S. Heymans. Use of Circulating MicroRNAs to Diagnose Acute Myocardial Infarction. Clin. Chem., 2012. [DOI | PubMed]
- S. Dimmeler, A.M. Zeiher. Circulating microRNAs: Novel biomarkers for cardiovascular diseases?. Eur. Heart J., 2010. [DOI | PubMed]
- Y. D’Alessandra, P. Devanna, F. Limana, S. Straino, A. Di Carlo, P.G. Brambilla, M. Rubino, M.C. Carena, L. Spazzafumo, M. De Simone. Circulating microRNAs are new and sensitive biomarkers of myocardial infarction. Eur. Heart J., 2010. [DOI | PubMed]
- A. Lopomo, F. Coppedè. Pharmacoepigenetics and pharmacoepigenomics of gastrointestinal cancers. Expert Rev. Gastroenterol. Hepatol., 2018. [DOI | PubMed]
- K.M. Smits, A.H. Cleven, M.P. Weijenberg, L.A. Hughes, J.G. Herman, A.P. de Bruïne, M. van Engeland. Pharmacoepigenomics in colorectal cancer: A step forward in predicting prognosis and treatment response. Pharmacogenomics, 2008. [DOI | PubMed]
- M. Tirri, P. Frisoni, S. Bilel, R. Arfè, C. Trapella, A. Fantinati, G. Corli, B. Marchetti, F. De-Giorgio, C. Camuto. Worsening of the Toxic Effects of (±) Cis-4,4′-DMAR Following Its Co-Administration with (±) Trans-4,4′-DMAR: Neuro-Behavioural, Physiological, Immunohistochemical and Metabolic Studies in Mice. Int. J. Mol. Sci., 2021. [DOI | PubMed]
- S. Schiavone, M. Neri, A.B. Maffione, P. Frisoni, M.G. Morgese, L. Trabace, E. Turillazzi. Increased iNOS and Nitrosative Stress in Dopaminergic Neurons of MDMA-Exposed Rats. Int. J. Mol. Sci., 2019. [DOI]
- D. Rudin, M.E. Liechti, D. Luethi. Molecular and clinical aspects of potential neurotoxicity induced by new psychoactive stimulants and psychedelics. Exp. Neurol., 2021. [DOI | PubMed]
- M. Carvalho, H. Carmo, V.M. Costa, J.P. Capela, H. Pontes, F. Remião, F. Carvalho, M.D.L. Bastos. Toxicity of amphetamines: An update. Arch. Toxicol., 2012. [DOI | PubMed]
- C.-T. Wu, J.R. Morris. Genes, Genetics, and Epigenetics: A Correspondence. Science, 2001. [DOI | PubMed]
- C. Dupont, D.R. Armant, C.A. Brenner. Epigenetics: Definition, Mechanisms and Clinical Perspective. Semin. Reprod. Med., 2009. [DOI | PubMed]
- R. Holliday. Epigenetics: A Historical Overview. Epigenetics, 2006. [DOI | PubMed]
- A. Ashe, V. Colot, B.P. Oldroyd. How does epigenetics influence the course of evolution?. Philos. Trans. R. Soc. B Biol. Sci., 2021. [DOI | PubMed]
- B. Alberts, A. Johnson, J. Lewis, M. Raff, K. Roberts, P. Walter. Chromosomal DNA and Its Packaging in the Chromatin Fiber. Molecular Biology of the Cell, 2002
- D.M. Walker, H.M. Cates, E.A. Heller, E.J. Nestler. Regulation of chromatin states by drugs of abuse. Curr. Opin. Neurobiol., 2015. [DOI | PubMed]
- M. Romani. Epigenetica, 2021
- I. Solovei, K. Thanisch, Y. Feodorova. How to rule the nucleus: Divide et impera. Curr. Opin. Cell Biol., 2016. [DOI | PubMed]
- J. Liu, M. Ali, Q. Zhou. Establishment and evolution of heterochromatin. Ann. N. Y. Acad. Sci., 2020. [DOI | PubMed]
- G. Felsenfeld, M. Groudine. Controlling the double helix. Nature, 2003. [DOI | PubMed]
- A.J. Bannister, T. Kouzarides. Regulation of chromatin by histone modifications. Cell Res., 2011. [DOI | PubMed]
- K. Hyun, J. Jeon, K. Park, J. Kim. Writing, erasing and reading histone lysine methylations. Exp. Mol. Med., 2017. [DOI | PubMed]
- I. Maze, K.-M. Noh, A. Soshnev, C.D. Allis. Every amino acid matters: Essential contributions of histone variants to mammalian development and disease. Nat. Rev. Genet., 2014. [DOI | PubMed]
- S. Tiwari. Recent Advancement in Methodology for Understanding Epigenetic Modifications. J. Clin. Epigenetics, 2017. [DOI]
- E. Li, Y. Zhang. DNA Methylation in Mammals. Cold Spring Harb. Perspect. Biol., 2014. [DOI | PubMed]
- J.I. Martín-Subero. How epigenomics brings phenotype into being. Pediatr. Endocrinol. Rev., 2011. [PubMed]
- M. Umer, Z. Herceg. Deciphering the Epigenetic Code: An Overview of DNA Methylation Analysis Methods. Antioxid. Redox Signal., 2013. [DOI | PubMed]
- A. Bird. DNA methylation patterns and epigenetic memory. An excellent review of mammalian DNA methylation. Genes Dev., 2002. [DOI | PubMed]
- P.A. Jones, S.B. Baylin. The fundamental role of epigenetic events in cancer. Nat. Rev. Genet., 2002. [DOI | PubMed]
- G.R. Wyatt. Recognition and estimation of 5-methylcytosine in nucleic acids. Biochem. J., 1951. [DOI | PubMed]
- D. Takai, P.A. Jones. Comprehensive analysis of CpG islands in human chromosomes 21 and 22. Proc. Natl. Acad. Sci. USA, 2002. [DOI | PubMed]
- K. Chen, B.S. Zhao, C. He. Nucleic Acid Modifications in Regulation of Gene Expression. Cell Chem. Biol., 2016. [DOI | PubMed]
- E. Li, C. Beard, R. Jaenisch. Role for DNA methylation in genomic imprinting. Nature, 1993. [DOI | PubMed]
- M. Curradi, A. Izzo, G. Badaracco, N. Landsberger. Molecular Mechanisms of Gene Silencing Mediated by DNA Methylation. Mol. Cell. Biol., 2002. [DOI | PubMed]
- J.L. Miller, P.A. Grant. The Role of DNA Methylation and Histone Modifications in Transcriptional Regulation in Humans. Epigenetics: Development and Disease, 2013. [DOI]
- A. Eharrison, A. Eparle-Mcdermott. DNA Methylation: A Timeline of Methods and Applications. Front. Genet., 2011. [DOI | PubMed]
- J. Tang, F. Fang, D.F. Miller, J.M. Pilrose, D. Matei, T.H.-M. Huang, K.P. Nephew. Global DNA Methylation Profiling Technologies and the Ovarian. Cancer Epigenetics, 2015. [DOI]
- M. Bibikova, J. Fan. Genome-wide DNA methylation profiling. WIREs Syst. Biol. Med., 2010. [DOI]
- J.S. Mattick, I.V. Makunin. Non-coding RNA. Hum. Mol. Genet., 2006. [DOI | PubMed]
- J.S. Mattick. RNA regulation: A new genetics?. Nat. Rev. Genet., 2004. [DOI | PubMed]
- J. Mattick, I.V. Makunin. Small regulatory RNAs in mammals. Hum. Mol. Genet., 2005. [DOI | PubMed]
- M.-M. Wei, G.-B. Zhou. Long Non-coding RNAs and Their Roles in Non-small-cell Lung Cancer. Genom. Proteom. Bioinform., 2016. [DOI | PubMed]
- H.M. Awan, A. Shah, F. Rashid, G. Shan. Primate-specific Long Non-coding RNAs and MicroRNAs. Genom. Proteom. Bioinform., 2017. [DOI]
- J.T. Lee, M.S. Bartolomei. X-Inactivation, Imprinting, and Long Noncoding RNAs in Health and Disease. Cell, 2013. [DOI | PubMed]
- C. Kanduri. Long noncoding RNAs: Lessons from genomic imprinting. Biochim. Biophys. Acta (BBA) Bioenerg. Mech., 2016. [DOI]
- J.M. Autuoro, S.P. Pirnie, G.G. Carmichael. Long Noncoding RNAs in Imprinting and X Chromosome Inactivation. Biomolecules, 2014. [DOI | PubMed]
- Y.S. Mao, H. Sunwoo, B. Zhang, D.L. Spector. Direct visualization of the co-transcriptional assembly of a nuclear body by noncoding RNAs. Nat. Cell Biol., 2011. [DOI | PubMed]
- J.-W. Wei, K. Huang, C. Yang, C.-S. Kang. Non-coding RNAs as regulators in epigenetics (Review). Oncol. Rep., 2017. [DOI | PubMed]
- S. Choudhuri. Small noncoding RNAs: Biogenesis, function, and emerging significance in toxicology. J. Biochem. Mol. Toxicol., 2010. [DOI | PubMed]
- Y. Shi, X. Jia, J. Xu. The new function of circRNA: Translation. Clin. Transl. Oncol., 2020. [DOI | PubMed]
- K. Saliminejad, H.R. Khorram Khorshid, S. Soleymani Fard, S.H. Ghaffari. An overview of microRNAs: Biology, functions, therapeutics, and analysis methods. J. Cell. Physiol., 2019. [DOI | PubMed]
- S.E. Hyman, R.C. Malenka, E.J. Nestler. Neural mechanisms of addiction: The Role of Reward-Related Learning and Memory. Annu. Rev. Neurosci., 2006. [DOI | PubMed]
- F. Vassoler, G. Sadri-Vakili. Mechanisms of transgenerational inheritance of addictive-like behaviors. Neuroscience, 2014. [DOI | PubMed]
- T.J. Gould. Addiction and cognition. Addict. Sci. Clin. Pract., 2010. [PubMed]
- M.W. Feltenstein, R.E. See. The neurocircuitry of addiction: An overview. Br. J. Pharmacol., 2008. [DOI | PubMed]
- P.W. Kalivas, N.D. Volkow. The Neural Basis of Addiction: A Pathology of Motivation and Choice. Am. J. Psychiatry, 2005. [DOI | PubMed]
- S. Jayanthi, M.T. McCoy, B. Chen, J.P. Britt, S. Kourrich, H.-J. Yau, B. Ladenheim, I.N. Krasnova, A. Bonci, J.L. Cadet. Methamphetamine Downregulates Striatal Glutamate Receptors via Diverse Epigenetic Mechanisms. Biol. Psychiatry, 2014. [DOI | PubMed]
- T. Heinbockel, A.B. Csoka. Epigenetic Effects of Drugs of Abuse. Int. J. Environ. Res. Public Health, 2018. [DOI]
- G. Schratt. microRNAs at the synapse. Nat. Rev. Neurosci., 2009. [DOI | PubMed]
- J.-L. Dreyer. New insights into the roles of microRNAs in drug addiction and neuroplasticity. Genome Med., 2010. [DOI | PubMed]
- A.A. Levine, Z. Guan, A. Barco, S. Xu, E.R. Kandel, J.H. Schwartz. CREB-binding protein controls response to cocaine by acetylating histones at the fosB promoter in the mouse striatum. Proc. Natl. Acad. Sci. USA, 2005. [DOI | PubMed]
- A. Kumar, K.-H. Choi, W. Renthal, N.M. Tsankova, D.E. Theobald, H.-T. Truong, S. Russo, Q. LaPlant, T.S. Sasaki, K.N. Whistler. Chromatin Remodeling Is a Key Mechanism Underlying Cocaine-Induced Plasticity in Striatum. Neuron, 2005. [DOI | PubMed]
- M.-C. Cheng, S.-H. Hsu, C.-H. Chen. Chronic methamphetamine treatment reduces the expression of synaptic plasticity genes and changes their DNA methylation status in the mouse brain. Brain Res., 2015. [DOI | PubMed]
- H.-Y. Shen, A. Kalda, L. Yu, J. Ferrara, J. Zhu, J.-F. Chen. Additive effects of histone deacetylase inhibitors and amphetamine on histone H4 acetylation, cAMP responsive element binding protein phosphorylation and Delta FosB expression in the striatum and locomotor sensitization in mice. Neuroscience, 2008. [DOI | PubMed]
- S. Vasudevan, Y. Tong, J.A. Steitz. Switching from Repression to Activation: MicroRNAs Can Up-Regulate Translation. Science, 2007. [DOI | PubMed]
- P. Rajasethupathy, F. Fiumara, R. Sheridan, D. Betel, S.V. Puthanveettil, J.J. Russo, C. Sander, T. Tuschl, E. Kandel. Characterization of Small RNAs in Aplysia Reveals a Role for miR-124 in Constraining Synaptic Plasticity through CREB. Neuron, 2009. [DOI | PubMed]
- N. Tsankova, W. Renthal, A. Kumar, E.J. Nestler. Epigenetic regulation in psychiatric disorders. Nat. Rev. Neurosci., 2007. [DOI | PubMed]
- A.J. Robison, E.J. Nestler. Transcriptional and epigenetic mechanisms of addiction. Nat. Rev. Neurosci., 2011. [DOI | PubMed]
- Q. LaPlant, V. Vialou, H.E.C. Iii, D. Dumitriu, J. Feng, B.L. Warren, I. Maze, D. Dietz, E.L. Watts, S. Iñiguez. Dnmt3a regulates emotional behavior and spine plasticity in the nucleus accumbens. Nat. Neurosci., 2010. [DOI | PubMed]
- A. Kalda, A. Zharkovsky. Epigenetic Mechanisms of Psychostimulant-Induced Addiction. Int. Rev. Neurobiol., 2015. [DOI | PubMed]
- X. Yang, M. Bam, P.S. Nagarkatti, M. Nagarkatti. RNA-seq Analysis of δ9-Tetrahydrocannabinol-treated T Cells Reveals Altered Gene Expression Profiles That Regulate Immune Response and Cell Proliferation. J. Biol. Chem., 2016. [DOI | PubMed]
- J. Feng, N.-Y. Shao, K.E. Szulwach, V. Vialou, J. Huynh, C. Zhong, T. Le, D. Ferguson, M. Cahill, Y. Li. Role of Tet1 and 5-hydroxymethylcytosine in cocaine action. Nat. Neurosci., 2015. [DOI | PubMed]
- L.H. Parsons, Y.L. Hurd. Endocannabinoid signalling in reward and addiction. Nat. Rev. Neurosci., 2015. [DOI | PubMed]
- R. Mechoulam, L.A. Parker. The Endocannabinoid System and the Brain. Annu. Rev. Psychol., 2013. [DOI | PubMed]
- H.C. Meyer, F.S. Lee, D.G. Gee. The Role of the Endocannabinoid System and Genetic Variation in Adolescent Brain Development. Neuropsychopharmacology, 2018. [DOI | PubMed]
- D.M. Fergusson, J.M. Boden, L.J. Horwood. Cannabis use and other illicit drug use: Testing the cannabis gateway hypothesis. Addiction, 2006. [DOI | PubMed]
- J. Orihuel, L. Gómez-Rubio, C. Valverde, R. Capellán, D. Roura-Martínez, M. Ucha, E. Ambrosio, A. Higuera-Matas. Cocaine-induced Fos expression in the rat brain: Modulation by prior Δ9-tetrahydrocannabinol exposure during adolescence and sex-specific effects. Brain Res., 2021. [DOI | PubMed]
- J. Tomas-Roig, E. Benito, R.C. Agis-Balboa, F. Piscitelli, S. Hoyer-Fender, V. Di Marzo, U. Havemann-Reinecke. Chronic exposure to cannabinoids during adolescence causes long-lasting behavioral deficits in adult mice. Addict. Biol., 2017. [DOI | PubMed]
- P.H. Gobira, A.L. Roncalho, N.R. Silva, G.P. Silote, A.J. Sales, S.R. Joca. Adolescent cannabinoid exposure modulates the vulnerability to cocaine-induced conditioned place preference and DNMT3a expression in the prefrontal cortex in Swiss mice. Psychopharmacology, 2021. [DOI | PubMed]
- M. Scherma, J.S. Qvist, A. Asok, S.-S.C. Huang, P. Masia, M. Deidda, Y.B. Wei, R.K. Soni, W. Fratta, P. Fadda. Cannabinoid exposure in rat adolescence reprograms the initial behavioral, molecular, and epigenetic response to cocaine. Proc. Natl. Acad. Sci. USA, 2020. [DOI | PubMed]
- H.C. Tomasiewicz, M.M. Jacobs, M.B. Wilkinson, S.P. Wilson, E.J. Nestler, Y.L. Hurd. Proenkephalin Mediates the Enduring Effects of Adolescent Cannabis Exposure Associated with Adult Opiate Vulnerability. Biol. Psychiatry, 2012. [DOI | PubMed]
- A.M. Gowen, K.E. Odegaard, J. Hernandez, S. Chand, S. Koul, G. Pendyala, S.V. Yelamanchili. Role of microRNAs in the pathophysiology of addiction. Wiley Interdiscip. Rev. RNA, 2021. [DOI | PubMed]
- K. Anier, K. Malinovskaja, A. Aonurm-Helm, A. Zharkovsky, A. Kalda. DNA Methylation Regulates Cocaine-Induced Behavioral Sensitization in Mice. Neuropsychopharmacology, 2010. [DOI | PubMed]
- D. Carouge, L. Host, D. Aunis, J. Zwiller, P. Anglard. CDKL5 is a brain MeCP2 target gene regulated by DNA methylation. Neurobiol. Dis., 2010. [DOI | PubMed]
- W. Renthal, I. Maze, V. Krishnan, H.E. Covington, G. Xiao, A. Kumar, S. Russo, A. Graham, N. Tsankova, T.E. Kippin. Histone Deacetylase 5 Epigenetically Controls Behavioral Adaptations to Chronic Emotional Stimuli. Neuron, 2007. [DOI | PubMed]
- W. Renthal, A. Kumar, G. Xiao, M. Wilkinson, H.E. Covington, I. Maze, D. Sikder, A. Robison, Q. LaPlant, D. Dietz. Genome-wide Analysis of Chromatin Regulation by Cocaine Reveals a Role for Sirtuins. Neuron, 2009. [DOI | PubMed]
- C.A. Koczor, I. Ludlow, R.S. Hight, Z. Jiao, E. Fields, T. Ludaway, R. Russ, R.A. Torres, W. Lewis. Ecstasy (MDMA) Alters Cardiac Gene Expression and DNA Methylation: Implications for Circadian Rhythm Dysfunction in the Heart. Toxicol. Sci., 2015. [DOI | PubMed]
- W. Tian, M. Zhao, M. Li, T. Song, M. Zhang, L. Quan, S. Li, Z. Sun. Reversal of Cocaine-Conditioned Place Preference through Methyl Supplementation in Mice: Altering Global DNA Methylation in the Prefrontal Cortex. PLoS ONE, 2012. [DOI | PubMed]
- K. Anier, A. Zharkovsky, A. Kalda. S-adenosylmethionine modifies cocaine-induced DNA methylation and increases locomotor sensitization in mice. Int. J. Neuropsychopharmacol., 2013. [DOI | PubMed]
- H. Zhang, A. Darwanto, T.A. Linkhart, L.C. Sowers, L. Zhang. Maternal Cocaine Administration Causes an Epigenetic Modification of Protein Kinase Cϵ Gene Expression in Fetal Rat Heart. Mol. Pharmacol., 2007. [DOI | PubMed]
- P. Prini, F. Penna, E. Sciuccati, T. Alberio, T. Rubino. Chronic Δ9-THC Exposure Differently Affects Histone Modifications in the Adolescent and Adult Rat Brain. Int. J. Mol. Sci., 2017. [DOI]
- S.K. Murphy, N. Itchon-Ramos, Z. Visco, Z. Huang, C. Grenier, R. Schrott, K. Acharya, M.-H. Boudreau, T. Price, D.J. Raburn. Cannabinoid exposure and altered DNA methylation in rat and human sperm. Epigenetics, 2018. [DOI | PubMed]
- D. Di Giacomo, E. De Domenico, C. Sette, R. Geremia, P. Grimaldi. Type 2 cannabinoid receptor contributes to the physiological regulation of spermatogenesis. FASEB J., 2016. [DOI | PubMed]
- J.A. DiNieri, X. Wang, H. Szutorisz, S.M. Spano, J. Kaur, P. Casaccia, D. Dow-Edwards, Y.L. Hurd. Maternal Cannabis Use Alters Ventral Striatal Dopamine D2 Gene Regulation in the Offspring. Biol. Psychiatry, 2011. [DOI | PubMed]
- C.T. Watson, H. Szutorisz, P. Garg, Q. Martin, J.A. Landry, A.J. Sharp, Y.L. Hurd. Genome-Wide DNA Methylation Profiling Reveals Epigenetic Changes in the Rat Nucleus Accumbens Associated With Cross-Generational Effects of Adolescent THC Exposure. Neuropsychopharmacology, 2015. [DOI | PubMed]
- P. Prini, F. Rusconi, E. Zamberletti, M. Gabaglio, F. Penna, M. Fasano, E. Battaglioli, D. Parolaro, T. Rubino. Adolescent THC exposure in female rats leads to cognitive deficits through a mechanism involving chromatin modifications in the prefrontal cortex. J. Psychiatry Neurosci., 2018. [DOI | PubMed]
- S. Hollins, K. Zavitsanou, F.R. Walker, M.J. Cairns. Alteration of imprinted Dlk1-Dio3 miRNA cluster expression in the entorhinal cortex induced by maternal immune activation and adolescent cannabinoid exposure. Transl. Psychiatry, 2014. [DOI | PubMed]
- M.E. Levine, A.T. Lu, A. Quach, B.H. Chen, T.L. Assimes, S. Bandinelli, L. Hou, A.A. Baccarelli, J.D. Stewart, Y. Li. An epigenetic biomarker of aging for lifespan and healthspan. Aging, 2018. [DOI | PubMed]
- A. Vidaki, D. Ballard, A. Aliferi, T. Miller, L.P. Barron, D.S. Court. DNA methylation-based forensic age prediction using artificial neural networks and next generation sequencing. Forensic Sci. Int. Genet., 2017. [DOI | PubMed]
- A.E. Field, N. Robertson, T. Wang, A. Havas, T. Ideker, P.D. Adams. DNA Methylation Clocks in Aging: Categories, Causes, and Consequences. Mol. Cell, 2018. [DOI | PubMed]
- S. Bocklandt, W. Lin, M.E. Sehl, F.J. Sánchez, J.S. Sinsheimer, S. Horvath, E. Vilain. Epigenetic Predictor of Age. PLoS ONE, 2011. [DOI | PubMed]
- V.V. Ashapkin, L.I. Kutueva, B.F. Vanyushin. Epigenetic Clock: Just a Convenient Marker or an Active Driver of Aging?. Adv. Exp. Med. Biol., 2019. [DOI | PubMed]
- R. Slieker, M. Van Iterson, R. Luijk, M. Beekman, D.V. Zhernakova, M.H. Moed, H. Mei, M. Van Galen, P. Deelen, M.J. Bonder. Age-related accrual of methylomic variability is linked to fundamental ageing mechanisms. Genome Biol., 2016. [DOI | PubMed]
- C.G. Bell, R. Lowe, P.D. Adams, A.A. Baccarelli, S. Beck, J.T. Bell, B.C. Christensen, V.N. Gladyshev, B.T. Heijmans, S. Horvath. DNA methylation aging clocks: Challenges and recommendations. Genome Biol., 2019. [DOI | PubMed]
- S. Horvath, K. Raj. DNA methylation-based biomarkers and the epigenetic clock theory of ageing. Nat. Rev. Genet., 2018. [DOI | PubMed]
- S. Horvath, C. Pirazzini, M.G. Bacalini, D. Gentilini, A.M. Di Blasio, M. Delledonne, D. Mari, B. Arosio, D. Monti, G. Passarino. Decreased epigenetic age of PBMCs from Italian semi-supercentenarians and their offspring. Aging, 2015. [DOI | PubMed]
- M.E. Reveron, T.J. Monks, C.L. Duvauchelle. Age-dependent (+)MDMA-mediated Neurotoxicity in Mice. Neurotoxicology, 2005. [DOI | PubMed]
- L. Frau, G. Costa, P.F. Porceddu, A. Khairnar, M.P. Castelli, M.G. Ennas, C. Madeddu, J. Wardas, M. Morelli. Influence of caffeine on 3,4-methylenedioxymethamphetamine-induced dopaminergic neuron degeneration and neuroinflammation is age-dependent. J. Neurochem., 2016. [DOI | PubMed]
- R. Feio-Azevedo, V.M. Costa, D.J. Barbosa, A. Teixeira-Gomes, I. Pita, S. Gomes, F.C. Pereira, M. Duarte-Araújo, J.A. Duarte, F. Marques. Aged rats are more vulnerable than adolescents to “ecstasy”-induced toxicity. Arch. Toxicol., 2018. [DOI | PubMed]
- I. Yusipov, M.G. Bacalini, A. Kalyakulina, M. Krivonosov, C. Pirazzini, N. Gensous, F. Ravaioli, M. Milazzo, C. Giuliani, M. Vedunova. Age-related DNA methylation changes are sex-specific: A comprehensive assessment. Aging, 2020. [DOI | PubMed]
- D.R. Masser, N. Hadad, H.L. Porter, C.A. Mangold, A. Unnikrishnan, M.M. Ford, C.B. Giles, C. Georgescu, M.G. Dozmorov, J.D. Wren. Sexually divergent DNA methylation patterns with hippocampal aging. Aging Cell, 2017. [DOI | PubMed]
- Life Expectancy at Birth (Years)
- R.C. Slieker, C.L. Relton, T.R. Gaunt, P.E. Slagboom, B.T. Heijmans. Age-related DNA methylation changes are tissue-specific with ELOVL2 promoter methylation as exception. Epigenetics Chromatin, 2018. [DOI | PubMed]
- H. Whitley, W. Lindsey. Sex-based differences in drug activity. Am. Fam. Physician, 2009. [PubMed]
- T. Chu. Gender Differences in Pharmacokinetics. US Pharm., 2014
- T. Lynch, A. Price. The effect of cytochrome P450 metabolism on drug response, interactions, and adverse effects. Am. Fam. Physician., 2007. [PubMed]
- A.L. Patton, K.A. Seely, A.L. Yarbrough, W. Fantegrossi, L.P. James, K.R. McCain, R. Fujiwara, P.L. Prather, J.H. Moran, A. Radominska-Pandya. Altered metabolism of synthetic cannabinoid JWH-018 by human cytochrome P450 2C9 and variants. Biochem. Biophys. Res. Commun., 2018. [DOI | PubMed]
- W.E. Fantegrossi, J.H. Moran, A. Radominska-Pandya, P.L. Prather. Distinct pharmacology and metabolism of K2 synthetic cannabinoids compared to Delta(9)-THC: Mechanism underlying greater toxicity?. Life Sci., 2014. [DOI | PubMed]
- A.J. Pedersen, T.H. Petersen, K. Linnet. In Vitro Metabolism and Pharmacokinetic Studies on Methylone. Drug Metab. Dispos., 2013. [DOI | PubMed]
- R.Z. Harris, L.Z. Benet, J.B. Schwartz. Gender Effects in Pharmacokinetics and Pharmacodynamics. Drugs, 1995. [DOI | PubMed]
- S.J. McClenahan, M.D. Hambuchen, C.M. Simecka, M.G. Gunnell, M.D. Berquist, S.M. Owens. Cardiovascular effects of 3,4-methylenedioxypyrovalerone (MDPV) in male and female Sprague-Dawley rats. Drug Alcohol Depend., 2019. [DOI | PubMed]
- L. Fattore, M. Marti, R. Mostallino, M.P. Castelli. Sex and Gender Differences in the Effects of Novel Psychoactive Substances. Brain Sci., 2020. [DOI]
- S. Bae, R.D. Gilbert, C.A. Ducsay, L. Zhang. Prenatal cocaine exposure increases heart susceptibility to ischaemia–reperfusion injury in adult male but not female rats. J. Physiol., 2005. [DOI | PubMed]
- N. De Giovanni, D. Marchetti. Cocaine and its metabolites in the placenta: A systematic review of the literature. Reprod. Toxicol., 2012. [DOI | PubMed]
- S. Bae, L. Zhang. Prenatal cocaine exposure increases apoptosis of neonatal rat heart and heart susceptibility to ischemia-reperfusion injury in 1-month-old rat. J. Cereb. Blood Flow Metab., 2005. [DOI | PubMed]
- K.D. Meyer, H. Zhang, L. Zhang. Prenatal cocaine exposure abolished ischemic preconditioning-induced protection in adult male rat hearts: Role of PKCε. Am. J. Physiol. Circ. Physiol., 2009. [DOI]
- S.I. Novikova, F. He, J. Bai, M.S. Lidow. Neuropathology of the cerebral cortex observed in a range of animal models of prenatal cocaine exposure may reflect alterations in genes involved in the Wnt and cadherin systems. Synapse, 2005. [DOI | PubMed]
- R. Thompson, K. DeJong, J. Lo. Marijuana Use in Pregnancy: A Review. Obstet. Gynecol. Surv., 2019. [DOI | PubMed]
- J.C. Sadeu, C.L. Hughes, S. Agarwal, W.G. Foster. Alcohol, drugs, caffeine, tobacco, and environmental contaminant exposure: Reproductive health consequences and clinical implications. Crit. Rev. Toxicol., 2010. [DOI | PubMed]
- N. Pintori, M.P. Castelli, C. Miliano, N. Simola, P. Fadda, L. Fattore, M. Scherma, M.G. Ennas, R. Mostallino, G. Flore. Repeated exposure to JWH-018 induces adaptive changes in the mesolimbic and mesocortical dopaminergic pathways, glial cells alterations, and behavioural correlates. J. Cereb. Blood Flow Metab., 2021. [DOI | PubMed]
- M.K. Skinner. Environmental epigenetic transgenerational inheritance and somatic epigenetic mitotic stability. Epigenetics, 2011. [DOI | PubMed]
- M.K. Skinner. What is an epigenetic transgenerational phenotype? F3 or F2. Reprod. Toxicol., 2008. [DOI | PubMed]
- E.E. Nilsson, I. Sadler-Riggleman, M.K. Skinner. Environmentally induced epigenetic transgenerational inheritance of disease. Environ. Epigenetics, 2018. [DOI | PubMed]
- M. Cloutier, C. Harris, S. Gayen, E. Maclary, S. Kalantry. Experimental Analysis of Imprinted Mouse X-Chromosome Inactivation. Methods Mol. Biol., 2018. [DOI | PubMed]
- J. Escher, S. Robotti. Pregnancy drugs, fetal germline epigenome, and risks for next-generation pathology: A call to action. Environ. Mol. Mutagen., 2019. [DOI | PubMed]
- N.M. Wanner, M. Colwell, C. Drown, C. Faulk. Developmental cannabidiol exposure increases anxiety and modifies genome-wide brain DNA methylation in adult female mice. Clin. Epigenetics, 2021. [DOI | PubMed]
- L. Silva, N. Zhao, S. Popp, D. Dow-Edwards. Prenatal tetrahydrocannabinol (THC) alters cognitive function and amphetamine response from weaning to adulthood in the rat. Neurotoxicology Teratol., 2012. [DOI | PubMed]
- A.N. Brown, J. Feng. Drug Addiction and DNA Modifications. Neuroepigenomics in Aging and Disease, 2017
- J. Alexandre, H. Carmo, F. Carvalho, J.P. Silva. Synthetic cannabinoids and their impact on neurodevelopmental processes. Addict. Biol., 2020. [DOI | PubMed]
- A.F. Scheyer, M. Melis, V. Trezza, O.J. Manzoni. Consequences of Perinatal Cannabis Exposure. Trends Neurosci., 2019. [DOI | PubMed]
- J. Alexandre, R. Malheiro, D.D. Da Silva, H. Carmo, F. Carvalho, J.P. Silva. The Synthetic Cannabinoids THJ-2201 and 5F-PB22 Enhance In Vitro CB1 Receptor-Mediated Neuronal Differentiation at Biologically Relevant Concentrations. Int. J. Mol. Sci., 2020. [DOI | PubMed]
- F. De-Giorgio, S. Bilel, A. Ossato, M. Tirri, R. Arfè, F. Foti, G. Serpelloni, P. Frisoni, M. Neri, M. Marti. Acute and repeated administration of MDPV increases aggressive behavior in mice: Forensic implications. Int. J. Legal. Med., 2019. [DOI | PubMed]
- A. Peacock, R. Bruno, N. Gisev, L. Degenhardt, W. Hall, R. Sedefov, J. White, K.V. Thomas, M. Farrell, P. Griffiths. New psychoactive substances: Challenges for drug surveillance, control, and public health responses. Lancet, 2019. [DOI | PubMed]
- F. Foti, M. Marti, A. Ossato, S. Bilel, E. Sangiorgi, F. Botrè, B. Cerbelli, A. Baldi, F. De-Giorgio. Phenotypic effects of chronic and acute use of methiopropamine in a mouse model. Int. J. Legal Med., 2019. [DOI | PubMed]
- K.E. Grafinger, M.E. Liechti, E. Liakoni. Clinical value of analytical testing in patients presenting with new psychoactive substances intoxication. Br. J. Clin. Pharmacol., 2020. [DOI | PubMed]
- F. De-Giorgio, S. Bilel, M. Tirri, R. Arfè, C. Trapella, C. Camuto, F. Foti, P. Frisoni, M. Neri, F. Botrè. Methiopropamine and its acute behavioral effects in mice: Is there a gray zone in new psychoactive substances users?. Int. J. Legal Med., 2020. [DOI | PubMed]
- F. De-Giorgio, E. Bergamin, S. Bilel, M. Tirri, R. Arfè, B. Marchetti, G. Corli, G. Serpelloni, M. Marti. Ethanol enhanced MDPV- and cocaine-induced aggressive behavior in mice: Forensic implications. Drug Alcohol Depend., 2021. [DOI | PubMed]
- A. Vidaki, M. Kayser. From forensic epigenetics to forensic epigenomics: Broadening DNA investigative intelligence. Genome Biol., 2017. [DOI | PubMed]
- Sabeeha, S.E. Hasnain. Forensic Epigenetic Analysis: The Path Ahead. Med. Princ. Pract., 2019. [DOI | PubMed]
- H.Y. Lee, S.-E. Jung, E.H. Lee, W.I. Yang, K.-J. Shin. DNA methylation profiling for a confirmatory test for blood, saliva, semen, vaginal fluid and menstrual blood. Forensic Sci. Int. Genet., 2016. [DOI | PubMed]
- M. Tachibana. Epigenetic regulation of mammalian sex determination. J. Med. Investig., 2015. [DOI | PubMed]
- S. Horvath. DNA methylation age of human tissues and cell types. Genome Biol., 2013. [DOI | PubMed]
- M.F. Fraga, E. Ballestar, M.F. Paz, S. Ropero, F. Setien, M.L. Ballestar, D. Heine-Suñer, J.C. Cigudosa, M. Urioste, J. Benitez. From The Cover: Epigenetic differences arise during the lifetime of monozygotic twins. Proc. Natl. Acad. Sci. USA, 2005. [DOI | PubMed]
- C. Liu, R.E. Marioni, Å.K. Hedman, L. Pfeiffer, P.-C. Tsai, L.M. Reynolds, A.C. Just, Q. Duan, C.G. Boer, T. Tanaka. A DNA methylation biomarker of alcohol consumption. Mol. Psychiatry, 2018. [DOI | PubMed]
- K.W. Lee, Z. Pausova. Cigarette smoking and DNA methylation. Front. Genet., 2013. [DOI | PubMed]
- C. Harris. Animal Models in Epigenetic Research: Institutional Animal Care and Use Committee Considerations across the Lifespan. ILAR J., 2012. [DOI | PubMed]
- N.L.H. Phillips, T.L. Roth. Animal Models and Their Contribution to Our Understanding of the Relationship Between Environments, Epigenetic Modifications, and Behavior. Genes, 2019. [DOI]