A computational analysis of the oncogenic and anti-tumor immunity role of P4HA3 in human cancers
Abstract
Prolyl-4-hydroxylase subunit alpha3 (P4HA3) is a triple helical procollagen synthesis protein. The role of P4HA3 in cancer development is not well known and lacks comprehensive analyses among human cancers. This study aimed to investigate the relationship between P4HA3 expression and anti-tumor immunity and its prognostic value in pan-cancer. P4HA3 expression was analyzed from TIMER2.0, GTEx, GEPIA2.0 and TCGA databases. Genetic and DNA methylation alterations, survival analysis and proteins co-expression analysis of P4HA3 in cBio Cancer Genomics Portal, TCGA, GSCA and TIMER2.0. The correlation between P4HA3 expression and immune infiltration was analyzed by TIDE, XCELL, MCPCOUNTER, and EPIC. We performed EdU and transwell experiments to evaluate the influence of P4HA3 on the proliferation, migration and invasion abilities of different tumors. Patients derived xenograft (PDX) and subcutaneous transplantation models were utilized to explore the correlation between P4HA3 and immunotherapy response in triple-negative breast cancer (TNBC). Among 33 types of cancers, P4HA3 had generally different expression between different tumors, further analysis showed that the expression of P4HA3 was correlated with the cells infiltration of the tumor microenvironment (TME). The expression of P4HA3 was positively with the cell proliferation markers and epithelial-mesenchymal transition (EMT) markers. Moreover, P4HA3 deficiency inhibited the proliferation, migration and invasion abilities of tumor cells, and promoted anti-tumor immunotherapy of PD-1/PD-L1 inhibitor. This pan-cancer analysis of P4HA3 provides a comprehensive understanding of its oncogenic and prognosis role in different cancers, P4HA3 abnormal expression could be a useful biomarker for predicting the effectiveness of immunotherapy in cancer patients.
Affiliations: Department of Breast Surgery, Zhujiang Hospital, Southern Medical University, Guangdong, China; Department of Thoracic Surgery, Zhujiang Hospital, Southern Medical University, Guangdong, China; Department of General Surgery, Zhujiang Hospital, Southern Medical University, Guangdong, China; Department of Clinical Laboratory, The Sixth Affiliated Hospital, Sun Yat-sen University, Guangdong, China; Biomedical Innovation Center, The Sixth Affiliated Hospital, Sun Yat-sen University, Guangdong, China; Department of Gastroenterology, Shenzhen Hospital, Southern Medical University, Guangdong, China; Department of Gynaecology and Obstetrics, Southern Hospital Taihe Branch, Southern Medical University, Guangdong, China; Cellular & Molecular Diagnostics Center, Sun Yat-Sen Memorial Hospital, Sun Yat-Sen University, Guangdong, China
License: © 2024 Huang et al CC BY 4.0 This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Article links: DOI: 10.1371/journal.pcbi.1012284 | PubMed: 39504328 | PMC: PMC11573185
Relevance: Moderate: mentioned 3+ times in text
Full text: PDF (5.2 MB)
Introduction
Up to now, diagnosis and therapy of cancer have been greatly improved, but there will be about 30 million new cancer cases in 2040 according to the report of GLOBOCAN, and cancer remains a huge social problem [ref. 1]. Genetic variants and epigenetic regulation closely associated with the occurrence and development of tumors [ref. 2], TME is a multicellular system that comprise infiltrating immune cells, CAFs, and stromal cells, many therapeutic strategies targeting TME such as PD-1/PD-L1 inhibitor and immune vaccines have achieved great advances in treating cancers [ref. 3,ref. 4]. Thus, it is very meaningful to explore the function and mechanism of cancer-associated genes in cancer malignant development.
P4HA3 was firstly identified as an α-subunit of prolyl hydroxylase which is participated in triple helical procollagen synthesis [ref. 5]. Additionally, P4HA3 was shown to be high expression in obesity and contribute to the development of type 2 diabetes [ref. 6]. P4HA3 was significantly upregulated in gastric cancer (GC) [ref. 7], stomach adenocarcinoma [ref. 8], melanoma [ref. 9] and P4HA3 upregulation was involved in the metastasis and prognosis of GC patients [ref. 8]. However, the relationship between P4HA3 and TME in various of cancers is largely unknown.
In this study, we investigated the expression and function of P4HA3 based on the different datasets in various of cancers, including: adrenocortical carcinoma (ACC), bladder urothelial carcinoma (BLCA), breast cancer (BRCA), cervical squamous carcinoma (CESC), cholangiocarcinoma (CHOL), colon adenocarcinoma (COAD), lymphoid neoplasm diffuse large B-cell lymphoma (DLBC), esophageal carcinoma (ESCA), glioblastoma multiforme (GBM), head and neck squamous cell carcinoma (HNSC), kidney chromophobe (KICH), kidney renal clear cell carcinoma (KIRC), kidney renal papillary cell carcinoma (KIRP), acute myeloid leukemia (LAML), brain low grade glioma (LGG), liver hepatocellular carcinoma (LIHC), lung adenocarcinoma (LUAD), lung squamous carcinoma (LUSC), mesothelioma (MESO), ovarian serous cystadenocarcinoma (OV), pancreatic adenocarcinoma (PAAD), pheochromocytoma and paraganglioma (PCPG), prostate adenocarcinoma (PRAD), rectum adenocarcinoma (READ), sarcoma (SARC), skin cutaneous melanoma (SKCM), stomach adenocarcinoma (STAD), testicular germ cell tumor (TGCT), thyroid carcinoma (THCA), thymoma (THYM), uterine corpus endometrial carcinoma (UCEC), uterine carcinosarcoma (UCS) and uveal melanoma (UVM). We explored the relationship between P4HA3 expression profile and prognosis value, genetic alteration, epigenetic regulation and TME among different cancers. Additionally, we performed the functional role of P4HA3 in breast cancer, colon cancer and lung cancer in vitro. More importantly, we also explored the association between P4HA3 depleted and cancer immunotherapy in breast cancer in vivo experiment. Our study indicates that P4HA3 depleted can promote anti-tumor immunity and P4HA3 upregulation maybe a potential prognostic molecular.
Results
Expression profiles and prognostic value of P4HA3 in different tumor types
To explore the clinical significance of P4HA3 among different tumors, we evaluated P4HA3 mRNA expression profile across TCGA and GTEx cancers by using the TIMER 2.0 and SangerBox tools. Compared to that normal tissues, P4HA3 mRNA expression was upregulated in BRCA, CHOL, COAD, ESCA, GBM, HNSC, KIRC, LUAD, LUSC, PCPG, PRAD, READ, STAD, THCA, ACC, LAML, PAAD, TCGT, and UCS tissues, suggesting that P4HA3 might function as an oncogenic role in the occurrence and development of diverse cancers (Fig 1A and 1B). Based on the GEPIA dataset, we found P4HA3 expression was positively correlated with advanced cancer stages in KIRC (P = 0.0118), LUAD (P = 0.0233), and STAD (P = 0.0396) (Fig 1C).
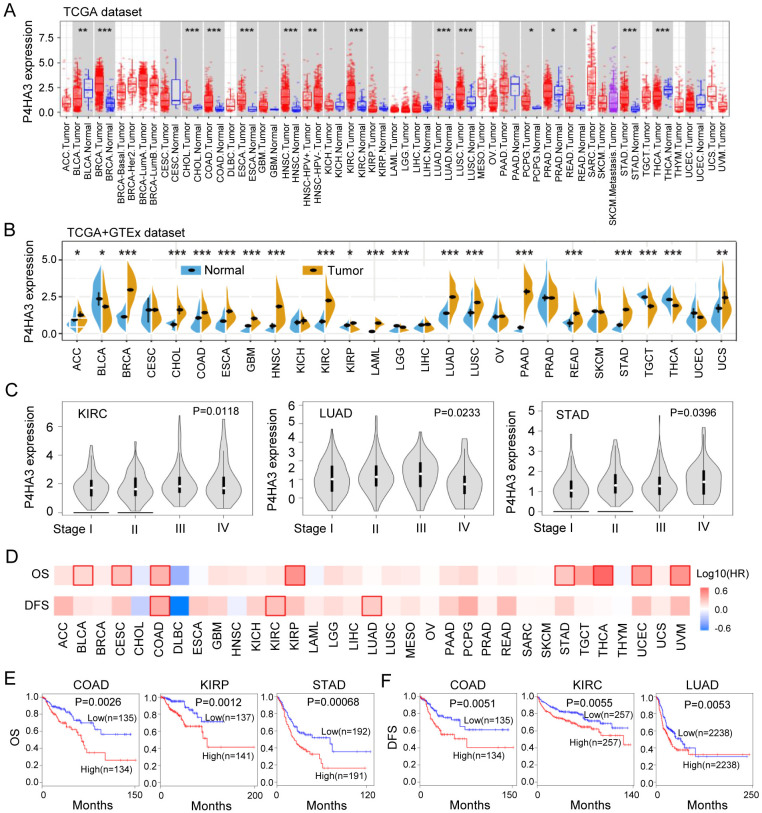
Next, we further analyzed the over survival (OS) and disease-free survival (DFS) of P4HA3 across the 33 different tumors in GEPIA datasets, the results were shown in Fig 1D and 1F. Elevated P4HA3 expression was correlated with poor OS in BLCA, CESC, COAD, KIRP, STAD, THCA, UCEC and UVM (P < 0.05) (Fig 1D), especially in COAD (P = 0.0026), KIRP (P = 0.0012), and STAD (P = 0.00068) (Fig 1E). Moreover, Elevated P4HA3 expression was correlated with poor DFS in COAD (P = 0.0051), KIRC (P = 0.0055) and LUAD (P = 0.0053) (Fig 1D and 1F). These results showed that elevated P4HA3 expression level was a great molecular significantly affecting the survival of different types of cancers.
The genetic alteration and epigenetic regulation of P4HA3 among different tumors
Based on above results that P4HA3 is generally high expression in various of cancers, we explored the genetic alteration and epigenetic regulation of P4HA3 from different databases. We firstly explored the genetic alteration of P4HA3 in TCGA pan-cancer atlas dataset by using the cBioPortal online database, we found that the overall alteration frequency of P4HA3 was widely high in various of cancers, P4HA3 had the highest genetic alteration frequency in SKCM, ESCA, OV, UCEC, HNSCC and BRCA (Fig 2A). Then, we analyzed the mutation frequency of 23 cancers by using SangerBox tool, the data showed that P4HA3 have relatively low mutation frequency in various of cancers (Fig 2B). Because of the overexpression of P4HA3 in cancers, we analyzed the copy number amplification variation of the P4HA3 DNA fragment, and found a positive correlation between the copy number amplification variation and P4HA3 expression among various of cancers, especially in UCEC, HNSCC, LUAD, OV, KIRP, ESCA, SKCM and THYM (Fig 2C) (FDR < 0.05).
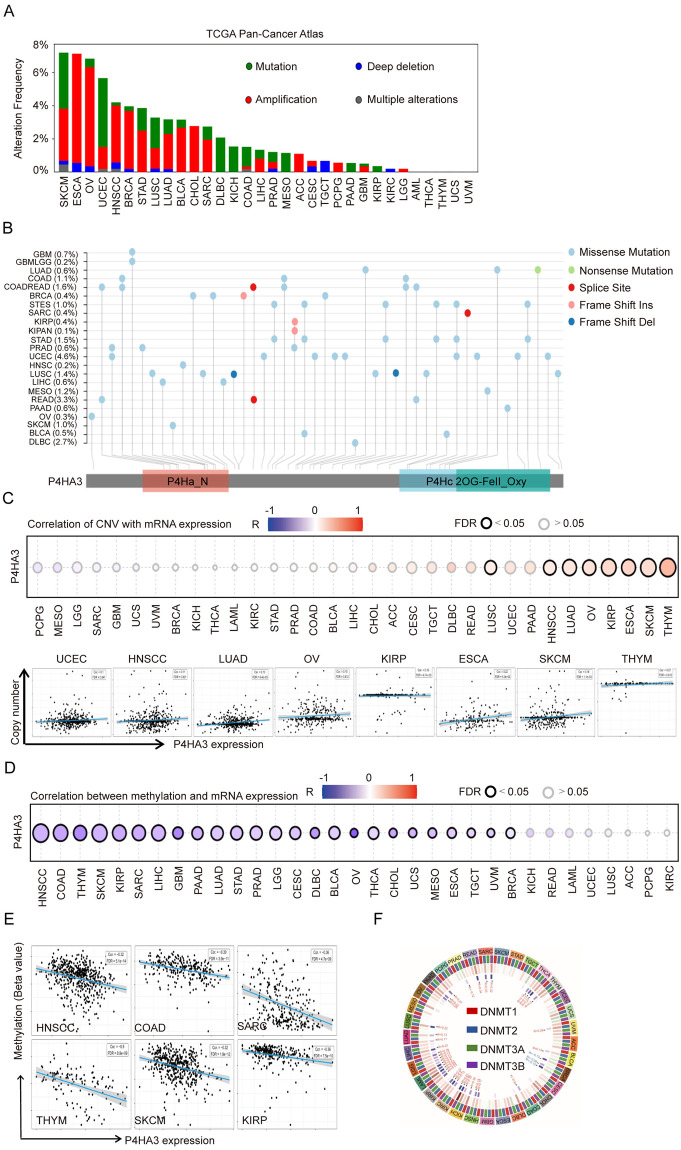
DNA methylation (DNAm) is an important epigenetic modification of chromosomes that playing a crucial role in gene regulation in cancers development [ref. 10,ref. 11]. We explored the relationship between DNA methylation and P4HA3 expression by using the gene set cancer analysis (GSCA) tool among different types of cancers. We found that there was negative correlation between DNA methylation and P4HA3 expression in cancers (Fig 2D), especially in HNSCC, COAD, OV, THYM, SKCM, KIRP and SARC (Fig 2E). The DNA methylation is regulated by methyltransferases (DNMTs) including DNMT1, DNMT2, DNMT3A and DNMT3B [ref. 12]. We found that the remarkable negative correlation with DNMTs and P4HA3 expression level in 33 types of cancers (Fig 2F). Our results showed that P4HA3 upregulation was correlated with DNA copy number amplification and methylation in various of cancers.
Genome wide association analysis of P4HA3 in different tumor types
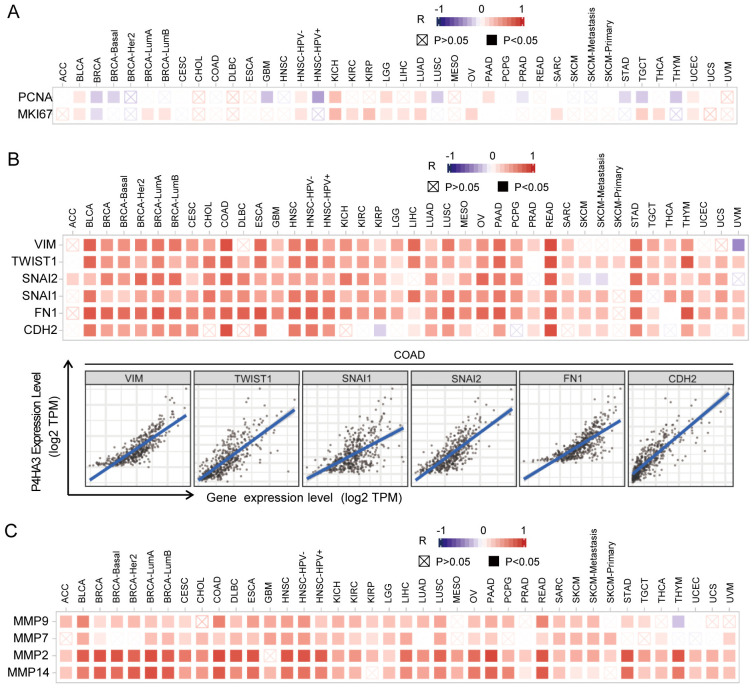
To explore the possible biological role of P4HA3 in various of cancers, we used gene co-expression analysis to evaluate P4HA3 function and regulatory mechanism. We found that P4HA3 expression was positively correlated with the typical proliferation proteins, including proliferating cell nuclear antigen (PCNA) and Ki-67 in various of cancers, such as BLCA, BRCA, HNSC, KICH by the heat map (P < 0.05) (Fig 3A). In addition, EMT is a crucial step of cancer cell migration and invasion [ref. 13], we focused on the possible associations between P4HA3 expression and classic EMT markers, such as Vimentin (VIM), TWIST1, Snail2 (SNAL2), Snail1 (SNAL1), Fibronectin1 (FN1), and N-cadherin (CDH2) [ref. 14]. The heat map showed that P4HA3 was significantly positively correlated with the expression of these EMT markers in various of cancers, especially in COAD (Fig 3B). We also analyzed the associations between P4HA3 expression and MMPs (Matrix Metalloproteinase) which also are crucial in cancer metastasis, and found that P4HA3 was significantly positively correlated with the expression of MMPs including Matrix Metalloproteinase 9 (MMP9), Matrix Metalloproteinase 7 (MMP7), Matrix Metalloproteinase 2 (MMP2) and Matrix Metalloproteinase 14 (MMP14), especially MMP2 and MMP14 (Fig 3C). All these data together suggested that high P4HA3 expression was widely associated with cancer proliferation and metastasis.
Correlation between P4HA3 expression and stromal and immune infiltration in cancers
The vital role of the TME including various immune, stromal and tumor cell types in regulating tumor growth, metastasis and prognosis is now generally understood [ref. 15,ref. 16]. We explored the possible correlation between P4HA3 expression level and the infiltrating immune and stromal cells by using the ESTIMATE algorithm [ref. 17], and found that P4HA3 expression was prominently correlated with immune score, stromal score and ESTIMATE score in various of cancers (Fig 4A). It is notably that P4HA3 expression level is positively correlated with immune score, stromal score and ESTIMATE score in BRCA and COAD (Fig 4A). Previous studies revealed that high immune/stromal/ESTIMATE scores were significantly associated with metastasis and poor prognosis in BRCA and COAD [ref. 18], suggesting that P4HA3 as an oncogene via increasing immune and stromal cell infiltration in BRCA and COAD. These data indicate that P4HA3 plays a crucial role in regulating the TME.
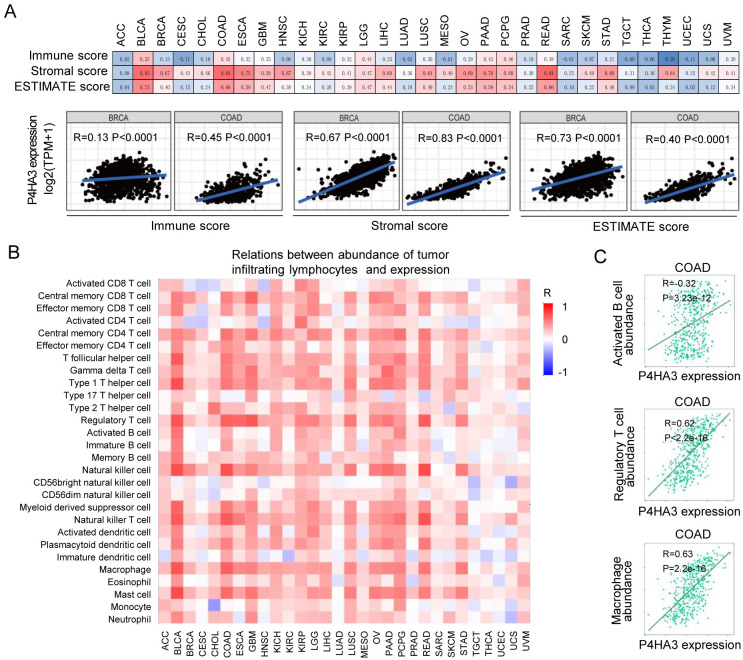
In order to further explore the relationship between P4HA3 expression and immune cells, we analyzed the association between P4HA3 expression and infiltration of 28 kinds of immune cells in the TISIDB platform [ref. 19]. In various of cancers, P4HA3 expression was positively correlated with infiltrated central memory CD8+ T cell, central memory CD4+ T cell, Type 1 T helper cell, regulatory T cell, natural killer cell, macrophage and mast cell (Fig 4B). In the previous studies, infiltrating immune cells play different role and different clinical significance in various of cancers [ref. 20]. In COAD, the infiltration of immune cells populations including activated CD8+ T cell, immature dendritic cell, CD56 bright/dim natural killer cell showed anti-tumor activity. By contrast, CD8+ T cell, activated B cell, regulatory T cell, M2 macrophage cell, regulatory dendritic cell and neutrophile cell had the characteristic of cancer-promoting [ref. 21, ref. 22]. In our present research, we found that P4HA3 expression was positively correlated with activated B cell, regulatory T cell and macrophage cell infiltration in COAD (Fig 4C). These results possibly indicate that P4HA3 promotes tumor development by regulating activated B cell, regulatory T cell and macrophage cell infiltration in COAD.
Correlation between P4HA3 expression and immune cell markers in cancers
CAFs are closely involved with cancer progression and therapeutic resistance, are the most abundant stromal cells in TME [ref. 3]. In order to explore the relationship between P4HA3 expression and infiltrating CAFs, we used four different algorithms (TIDE, XCELL, MCPCOUNTER and EPIC) in our present work. We found that P4HA3 expression level was positively associated with the number of infiltrating CAFs in various of cancers (appeared in at least 3 out 4 algorithms) (Fig 5A). especially in BRCA and COAD, P4HA3 expression level was positively associated with the number of infiltrating CAFs in the 4 algorithms (P < 0.0001) (Fig 5B). Next, we further analyzed the correlation between P4HA3 expression and immune marker genes, including CSF1R and CD86 (representing monocyte cells), IL10, CD68 and CCL2 (representing tumor-associated macrophage cells; TAM), PTGS2, NOS2 and IRF5 (representing M1 macrophage cells), VSIG4, MS4A4A and CD163 (representing M2 macrophage cells). The results showed that the expression of monocyte, TAM, M1 macrophage and M2 macrophage markers were significantly correlated with P4HA3 expression in BLCA, BRCA, COAD, ESCA, GBM, HNSC, KICH, KIRC, KIRP, LGG, LIHC, LUAD, LUSC, OV, PAAD, PCPG, PRAD, READ, SKCM, STAD, THYM and UVM (P < 0.05) (Fig 5C). Specifically, it was found that CSF1R and CD86 of monocyte were positively correlated with P4HA3 expression in BLCA, COAD, ESCA, READ and THYM; IL10, CD68 and CCL2 of TAMs were positively correlated with P4HA3 expression in BLCA, BRCA, COAD, ESCA, GBM, HNSC, KICH, KIRC, KIRP, LGG, LIHC, LUAD, LUSC, OV, PAAD, PCPG, PRAD, READ, SKCM, STAD, THYM and UVM; PTGS2, NOS2 and IRF5 of M1 phenotype were positively correlated with P4HA3 expression in BLCA, BRCA, LIHC, LUAD, LUSC, OV, PAAD, SKCM, UVM; VSIG4, MS4A4A and CD163 of M2 phenotype were positively correlated with P4HA3 expression in BLCA, BRCA, CHOL, COAD, ESCA, GBM, HNSC, KICH, KIRC, KIRP, LGG, LIHC, LUSC, OV, PAAD, PCPG, PRAD, READ, SKCM, STAD, THYM and UVM (P < 0.05) (Fig 5C). The role of P4HA3 in regulating TME and cancer progression need more research to confirm.
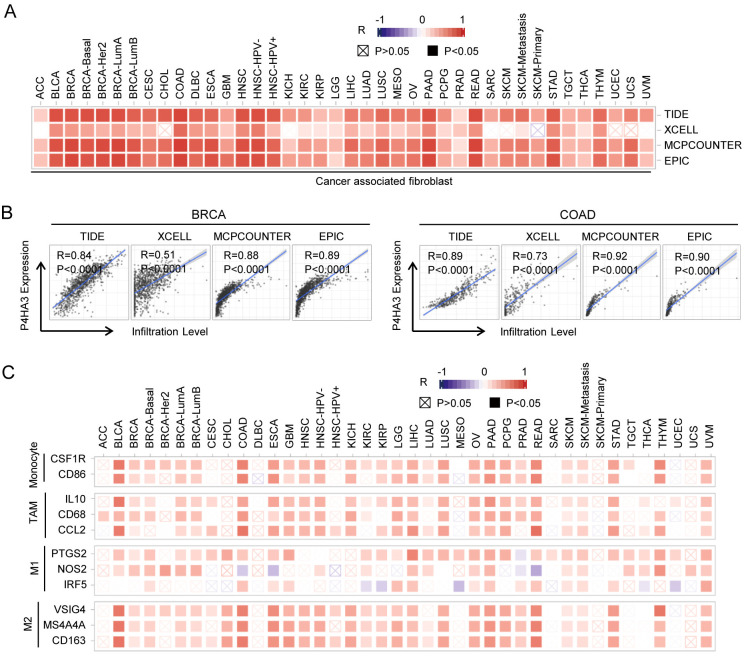
Correlations between P4HA3 expression and Microsatellite (MSI) and Tumor mutation burden (TMB) in cancers
P4HA3 was positively correlated with MSI in UCS, COAD, MESO and PRAD, and negatively correlated with MSI in STAD and LUSC (Fig 6A). P4HA3 was positively correlated with TMB in THYM, PRAD, LUAD, LGG and LAML, and negatively correlated with TMB in TGCT, LUSC, LIHC, KIRP, HNSC, and BRCA (Fig 6B). In addition, it is very crucial that cancer cells escape the immunosurveillance of host in the prognosis of cancers [ref. 23]. We analyzed the correlation between P4HA3 expression and immune checkpoint genes, including PD-1, PD-L1, CD276, and CTLA-4. In BRCA, P4HA3 expression was positively correlated with the expression of CD200, NRP1, LAIR1, TNFSF4, HAVCR2, CD276, VSIR and TNFRSF4, and negatively correlated with the expression of MSH2, MSH6, EPCAM, BTLA, LAG3, ICOS, CTLA-4, PD-L1, TMIGD2, IDO1, TIGIT. In COAD, P4HA3 expression was positively correlated with the expression of PMS2, BTLA, CD200, NRP1, LAIR1, TNFSF4, ICOS, CTLA-4, CD48, CD28, CD200R1, HAVCR2, ADORA2A, CD276, CD80, TNFSF14, PD-1, TNFSF18, TNFRSF8, CD27, VSIR, TNFRSF4, CD40, TIGIT, PD-L1, CD86 and TNFRSF9, and negatively correlated with the EPCAM. In LGG, P4HA3 expression was positively correlated with the expression of BTLA, TNFRSF14, NRP1, LAIR1, TNFSF4, CD244, ICOS, CD40LG, CTLA4, CD48, CD28, CD200R1, HAVCR2, CD276, KIR3DL1, CD80, PDCD1, LGALS9, CD160, TNFSF14, IDO2, IDO1, PDCD1LG2, CD27, TNFRSF25, TNFRSF4, CD40, TNFRSF18, TNFSF15, CD274, CD86, CD44, TNFRSF9 (Fig 6C). Significantly, P4HA3 expression was negatively correlated with the expression of PD-1 in BRCA, and positively correlated with the expression of PD-1 in LGG and PCPG. P4HA3 expression was positively correlated with the expression of PD-L1 in CHOL, COAD, KIRC, LGG, LIHC and PAAD. P4HA3 expression was positively correlated with the expression of CTLA-4 in BLCA, COAD, GBM, LGG, OV and PRAD, and negatively correlated with the expression of CTLA-4 in BRCA (Fig 6C). These results showed that high expression of P4HA3 played a crucial role in cancer progression by regulating immune checkpoint gene, such as PD-1/PD-L1/CTLA-4. Further, we analyzed the functional enrichment of high P4HA3 expression and low P4HA3 expression by GSEA platform. KEGG and Hallmark enrichment plots showed that high expression of P4HA3 was associated with metabolic-related pathways, including metabolism of nitrogen and bile acid (Fig 6D).
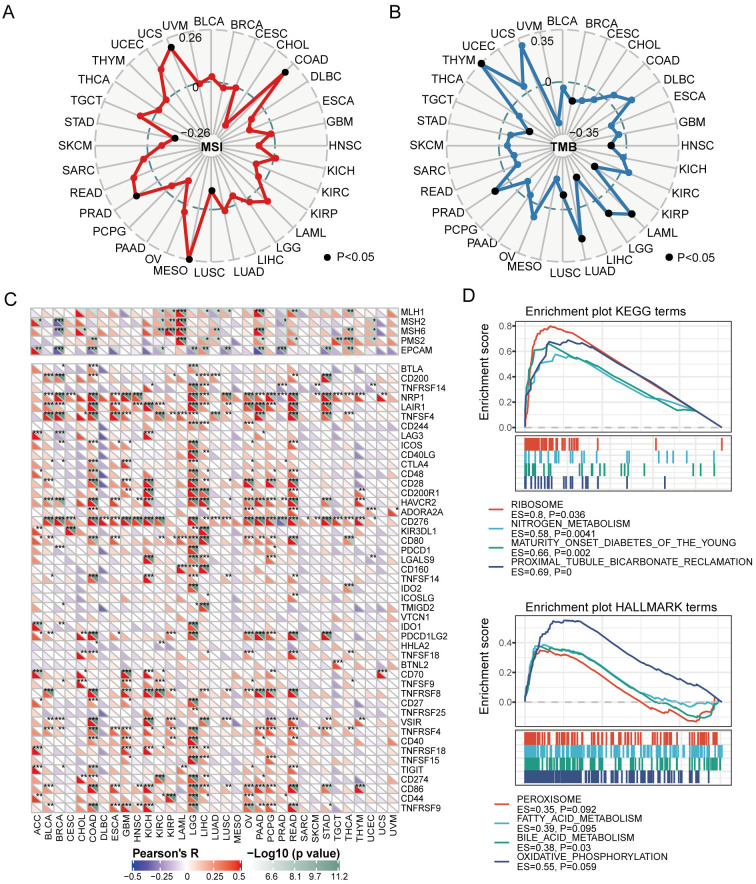
Confirmation by in vitro and in vivo experiments
To confirm the anti-tumor role of P4HA3 deficiency by enhancing anti-tumor immune procedure in cancers, we performed migration, invasion and EdU experiments in vitro and mouse tumor models in vivo. We found that P4HA3 loss significantly inhibited migration, invasion and proliferation abilities in MDA-MB-231, HCT116 and A549 cell lines (Fig 7A and 7B), the related statistical results were shown in S1A and S1B Fig. The knockdown efficiency of P4HA3 in MDA-MB-231, HCT116 and A549 cell lines were detected by using qRT-PCR and western-blot, and the knockdown efficiency was up to 80% (S1C and S1D Fig). In PDX mouse model of TNBC, we found P4HA3-knocked down markedly inhibited PDX tumor growth (Fig 7C and S1E Fig), P4HA3-knocked down also inhibited the expression of P4HA3 and Ki-67 in IHC sections (Fig 7D). More importantly, we found that the volume of the P4HA3-knocked out tumors markedly decreased relative to that of the P4HA3-wildtype tumors after using BMS-1 in c57 mouse model of inoculating 4T1 cells with or without BMS-1, a small-molecular inhibitor of PD-1/PD-L1 (Fig 7E and S1F Fig). We also found P4HA3-knocked out decreased Ki-67 expression and increased PD-1 expression by western-blot assay (Fig 7F). These results indicated higher sensitivity of the P4HA3-knocked out tumor to the PD-1/PD-L1 inhibitor in triple-negative breast cancer.
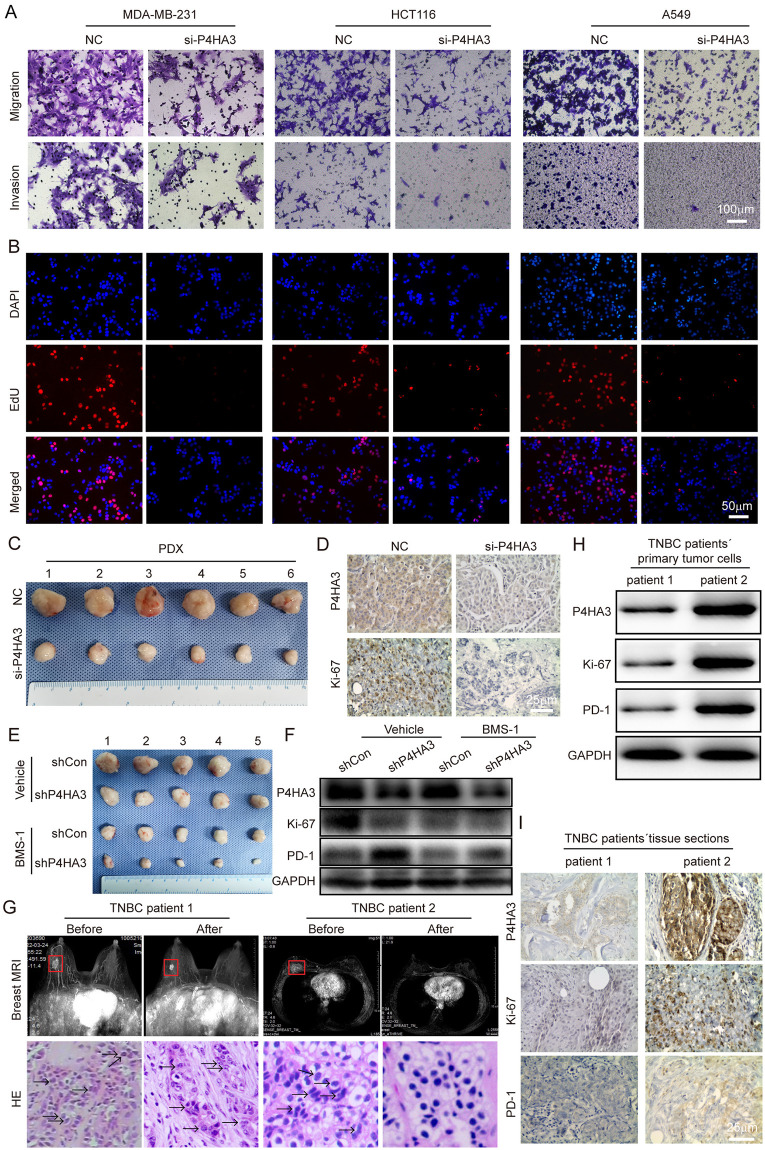
P4HA3 expression could reflect the immunotherapy response of TNBC
To determine whether P4HA3 expression could represent the TNBC patients´ immunotherapy response, we retrospectively selected two TNBC patients with neoadjuvant immunotherapy (PD-1 inhibitor: Camrelizumab). Breast MRI scan images showed that the tumor size of the two patients was significantly reduced after immunotherapy, and the tumor of second TNBC patient was totally didn’t find. Meanwhile, we used hematoxylin and eosin staining (HE) in needle biopsy or intact tissues of tumors before or after immunotherapy to show immunotherapy effect, we found that the tumor of second TNBC patient was achieved a pathological complete response (pcr) after immunotherapy (Fig 7G). More importantly, the tumor tissues from needle biopsy of two TNBC patients before immunotherapy were conducted for WB and IHC staining to detect the expression of P4HA3, Ki-67 and PD-1, we found that the expression of P4HA3, Ki-67 and PD-1 in second TNBC patient was significantly higher than the first TNBC patient (Fig 7H and 7I). Above results indicated that TNBC patients with high expression of P4HA3 and PD-1 will possibly benefit from immunotherapy, P4HA3 could be a predictor for immunotherapy response of TNBC patients like PD-1.
Discussion
In the previous study, P4HA3 was aberrantly expressed in gastric cancer [ref. 7], colon cancer and squamous cell carcinomas [ref. 24]. In our study, the expression of P4HA3 was investigated in the 33 types of tumors in the databases, the expression of P4HA3 was generally upregulated among various of tumors. The biological function of P4HA3 was proved to be involved in EMT, migration and invasion in colon cancer [ref. 24]. We integrated analyzed the function of P4HA3 among 33 types of cancers, and found that P4HA3 was associated with proliferation markers (PCNA and MKI67), EMT markers (VIM, TWIST1, SNAI1, SNAI2, FN1 and CDH2), extracellular matrix (ECM) markers (MMP9, MMP7, MMP2 and MMP14). In the study, we found that P4HA3 significantly promoted the migration, invasion and proliferation abilities of MDA-MB-231, HCT116 and A549 cells. According to these results, we conducted that P4HA3 conferred malignant phenotypes to various of cancer cells via activating EMT, in the further, we will perform more experiments to verify this conjecture. Enrichment analysis results indicated that high expression of P4HA3 was associated with metabolic-related pathways, the metabolic disorder of host and cancer cells are crucial for understanding tumor origin and development [ref. 25]. In NSCLC, TGF-β pathway altered the amino acid metabolism and induced EMT happen by increasing P4HA3 expression, consequently contributed to cancer cell malignant progression [ref. 26]. We supposed that P4HA3 was associated with EMT, ECM malignant phenotypes to various of cancer cells via amino acid metabolism activation, these results provided us some new insights of P4HA3 and amino acid metabolism to search for effective therapeutic targets, biological experiments are needed to verify these new findings and promote clinical transformation.
In this study, we firstly explored the significant role of P4HA3 deficiency in promoting anti-tumor immunity and immunotherapeutic limitation in various of cancers. The rapid development of cancer immunotherapy has significantly extended the life of tumor patients especially advanced NSCLC. Basing on the successful development of immune checkpoint blockade inhibitors (ICBs) against PD-1 (e.g., pembrolizumab, nivolumab), PD-L1 (e.g., atezolizumab, avelumab) and CTLA4 (e.g., ipilimumab), ICBs have produced great responses in many tumors [ref. 27]. Various of creative immunotherapies are currently undergoing clinical trial for cancer treatment such as chimeric antigen receptor-T cell immunotherapies (CAR-T) [ref. 28]. but for BRCA as an immune-cold (non-T cell infiltration) solid tumor, these patients could not get considerable clinical benefit from ICBs, therefore, the diversity of immune evasion remains a key challenge in converting “cold” tumors into “hot” (T cell infiltration) ones [ref. 29]. In the present study, P4HA3 deficiency can strengthen anti-tumor immunity by increasing PD-1 expression and decreasing Ki-67 expression in BRCA. It would be logical that P4HA3 deficiency enhanced susceptibility to ICB therapy for breast cancer. This inference was proved in breast cancer mouse model receiving BMS-1 (PD-1/PD-L1 inhibitor) therapy (Fig 6E). Therefore, these results indicated the potential role of P4HA3 in tumor immunology and prognostic biomarker of cancers.
CAFs are the most abundant and hyper-activated in cancer progression within TME [ref. 3]. All in all, CAFs are involved in tumor initiation, angiogenesis, metastasis, chemoresistance and immunosuppression via cell-cell interaction and the production of multiple regulatory molecules [ref. 30, ref. 31]. Previous studies showed that CD10+GPR77+ CAF subset provided a survival niche for cancer stem cells and induced drug resistance in breast and lung cancer [ref. 32], they found that CCL18-PITPNM3 could induce the information of CD10+GPR77+ CAFs, blocking CCL18-PITPNM3 signal axis could not only reverse chemoresistance and immunosuppression and inhibit cancer progression but also prevent the development of pro-tumor CAFs in the early stage of cancer progression [ref. 33]. In our study, we found that the infiltration of CAFs was notably associated with P4HA3 expression in various of tumors, especially BRCA, suggesting that P4HA3 may involve in the generation and evolution of CAFs. In the further, we will conduct more experiments to verify the viewpoint that P4HA3 may be an attractive therapeutic target by blocking pro-tumor CAFs and reversing immunosuppression and chemoresistance in the TME.
Materials and methods
Ethics statement
The study protocol and patients’ samples were approved by The Ethics Committee of the Zhujiang Hospital, Southern Medical University (Guangzhou, China). All patients provided informed written consent. All clinical tissue specimens from patients were acquired with patients’ written informed consents and all experiments were conducted in accordance with the Declaration of Helsinki. The animal experiments were approved by the Animal Care and Use Committee of Zhujiang Hospital, south medical university (Guangzhou, China), the committee number is LAEC-2021-103. The maximal tumor size/burden was not exceeded the permissible tolerance in institutional guidelines.
P4HA3 expression and survival analysis
The expression of P4HA3 in tumor and adjacent para-cancer tissues, as well as in various tumor stages across 33 types of cancers was analyzed using the TIMER2.0 online tool (http://timer.comp-genomics.org/) [ref. 34]. The expression of P4HA3 in normal tissues was obtained from Genotype-Tissue Expression Project (GTEx) [ref. 35]. To explore the association between P4HA3 expression and patient survival, the GEPIA2.0 database was used (http://gepia2.cancerpku.cn/#index) [ref. 36]. The median expression of P4HA3 was defined as the cutoff value to determine the high or low expression of P4HA3 by using the “Survival Map” module.
Genetic and DNA methylation alteration of P4HA3
The mutation frequency of P4HA3 in tumor tissues across 33 types of cancers was present by the cBio Cancer Genomics Portal (http://cbioportal.org) [ref. 37]. The specific genetic alteration information of P4HA3 was conducted by the “TCGA Pan-Cancer Atlas Studies” dataset. To evaluate the effect of copy number variation and DNA methylation on P4HA3 expression, the GSCA (Gene Set Cancer Analysis) (http://bioinfo.life.hust.edu.cn/GSCA/#/) was used [ref. 38]. The correlation analyses for P4HA3 expression and copy number variation, DNA methylation levels as well as the expression of methylation regulators (DNMT1, DNMT2, DNMT3A, and DNMT3B) across various cancers were conducted.
Co-expression analysis of P4HA3
We utilized the “Gene_Corr” module of TIMER2.0 to explore the association between the expression of P4HA3 and tumor malignancy and immune related genes in various cancers [ref. 34]. Several tumor malignancy and immune related genes were analyzed, including proliferation markers (PCNA and MKI67), epithelial-mesenchymal transition markers (VIM, TWIST1, and CDH2, etc.), microsatellite instability markers (MLH1, MSH2, MSH6, and PMS2, etc.), macrophage markers (CD86, CSF1R, and IL10, etc.), and immune checkpoint markers (PDCD1, CTLA4, and LAG3, etc.). The p-value and correlation coefficient were conducted by the Spearman correlation analysis method. p < 0.05 was considered significant.
Immune microenvironment analysis
In order to understand the potential effect of P4HA3 on immune microenvironment, the stromal/immune/estimate score and immune cell infiltration levels were evaluated. The stromal/ immune/estimate score of each tumor sample were evaluated by the R package “estimate” [ref. 17]. Spearman correlation analysis was used to reveal the associations between P4HA3 expression and stromal/immune/estimate scores. We used the TISIDB online platform to evaluate the correlation between P4HA3 expression and immune cell abundance [ref. 19]. As for cancer-associated fibroblast infiltration, four algorithms (TIDE, XCELL, MCPCOUNTER, and EPIC) were used. The purity-adjusted p-value and correlation coefficient were conducted by the Spearman’s test. p < 0.05 was considered significant.
Tumor mutational burden (TMB) and microsatellite instability (MSI) analysis
TMB was defined as the number of mutations (including somatic, coding, base substitution and indel) detected per million genomes. We obtained the TCGA genomic data across 33 types of cancers from the cBio Cancer Genomics Portal and calculated the TMB of each sample [ref. 37]. We obtained TCGA MSI score (MANTIS) across 33 types of cancers by “cBioPortalData” the R package [ref. 39]. The associations between P4HA3 expression and TMB/MSI scores were calculated by Spearman’s test. p < 0.05 was considered significant.
Gene set enrichment analysis (GSEA)
To explore the underlying mechanism of P4HA3, the GSEA was performed, which Kyoto Encyclopedia of Genes and Genomes (KEGG) and Hallmark module of the Molecular Signatures Database were selected [ref. 40]. Each of the top four pathway was present.
Cell migration and invasion assays
In this study, to explore the migration and metastasis abilities of breast cancer, colon cancer and lung cancer cells, we used transwell chambers for building the cell barrier. In invasion assay, a layer of Matrigel (BD Bioscience) was placed above the chamber. Indicated cells were pretreated with siP4HA3 (RiboBio, Guangzhou) for 48 h, and was subsequently trypsinized and washed by PBS. Two thousand cells were seeded into the upper chambers in DMEM with 3% FBS, and the lower chambers were added into DMEM with 10% FBS. About 12 h, the chambers were collected and quantified by photographing in 3 random fields.
EdU assay
In this study, we used an EdU (5-ethynyl-2´-deoxyuridine; Invitrogen, California, USA) assay to detect the proliferation abilities of breast cancer, colon cancer and lung cancer cells. Indicated cells were pretreated with siP4HA3 (RiboBio, Guangzhou) for 48 h, and 2 × 103 cells were plated in 96-well plates for 24 h. The cells were incubated with 10 μM EdU solution for 24 h, and sequent fixation, permeabilization and EdU staining. The results were determined with fluorescence microscopy and were collected and quantified by photographing in 3 random fields.
In vivo experiments
To explore the influence of P4HA3 on TNBC tumorigenesis, we established one case PDX model as follows: TNBC clinical specimens were obtained from TNBC patient (the tumor size was about 3.5 cm, the patient age´s age was 48 years old) who experienced tumor excised at Zhujiang Hospital, south medical university (Guangzhou, China) in 2022. The tumor was cut into small incision and putted into the fourth pair mammary fat pads of anaesthetized four-week-old NOSCID. When the maximum diameter of PDX was up to 5mm about two months, the siRNA against to P4HA3 were intravenous injection of NOSCID one time every 3 days about 6 weeks. Tumor volumes did not exceed the maximum volumes according to the requirement of Animal Care and Use Committee. The tumors were measured every one week and was calculated as the formula: V = 1/2 × width2 × length.
To explore the relationship between P4HA3 and immunotherapy of TNBC, we established subcutaneous tumor model. In 4T1 cells, we steady knocked out P4HA3 by transducing shCon or shP4HA3 lentivirus selected by using puromycin. 1 × 106 4T1 cells with shCon or shP4HA3 were injected into the third right mammary fat pads with C57/BL6 background. Once tumors reached 5 mm in diameter according to the standard modified formula Volume (mm3) = (length × height2)/2, half of the shCon and shP4HA3 mice were treated with 100 U/L PD-1/PD-L1 inhibitor BMS-1 (Selleck Cas: No. 1675201-83-8) every 3 days. After 21 days of treatment, the tumors were excised, tumor tissues were individually preserved for IHC, ISH, RNA and protein extraction.
Statistical analysis
All statistical analyses were conducted by student t-test by using Graph pad prism 5.0, the results in vitro and in vivo were repeated three independent experiments. The statistical analyses of results in vitro and in vivo were separately presented as mean ± SD and mean ± SEM.
Supplementary Materials
- P4HA3 deficiency inhibited malignant biological abilities of cancer cells.The statistical analysis from Fig 7. (A) P4HA3 deficiency inhibited migration and invasion abilities of MDA-MB-231, HCT116 and A549 cell lines by transwell assay, (***p < 0.001 among the different groups by paired Student´s t tests, mean ± SD). (B) P4HA3 deficiency inhibited proliferation ability of MDA-MB-231, HCT116 and A549 cell lines by EdU assay, (***p < 0.001 among the different groups by paired Student´s t tests, mean ± SD). (C) The knockdown efficiency of P4HA3 in MDA-MB-231, HCT116 and A549 cell lines were detected by using qRT-PCR, (***p < 0.001 among the different groups by paired Student´s t tests, mean ± SD). (D) The knockdown efficiency of P4HA3 in MDA-MB-231, HCT116 and A549 cell lines were detected by using WB. (E) Tumor size in different groups were calculated every 7 days over 2 months. (***p < 0.001 between the NC group and siRNA group by paired Student´s t tests, mean ± SD). (F) Tumor size in different groups were calculated every one week over one month. (***p < 0.001 among the different groups by paired Student´s t tests, mean ± SD).(TIF) (TIF)
- Submitted filename: Authors response to reviewers tracked change.docx (DOCX)
References
- F Bray, J Ferlay, I Soerjomataram, RL Siegel, LA Torre, A Jemal. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries.. CA: a cancer journal for clinicians., 2018. [DOI | PubMed]
- SL Topalian, FS Hodi, JR Brahmer, SN Gettinger, DC Smith, DF McDermott. Safety, activity, and immune correlates of anti-PD-1 antibody in cancer.. The New England journal of medicine., 2012. [DOI | PubMed]
- X Chen, E Song. Turning foes to friends: targeting cancer-associated fibroblasts.. Nature reviews Drug discovery., 2019. [DOI | PubMed]
- WX Xu, J Zhang, YT Hua, SJ Yang, DD Wang, JH Tang. An Integrative Pan-Cancer Analysis Revealing LCN2 as an Oncogenic Immune Protein in Tumor Microenvironment.. Frontiers in oncology., 2020. [DOI | PubMed]
- Y Luo, W Xu, H Chen, D Warburton, R Dong, B Qian. A novel profibrotic mechanism mediated by TGFbeta-stimulated collagen prolyl hydroxylase expression in fibrotic lung mesenchymal cells.. The Journal of pathology., 2015. [DOI | PubMed]
- L Zhuang, C Li, X Hu, Q Yang, X Pei, G Jin. High expression of P4HA3 in obesity: a potential therapeutic target for type 2 diabetes.. Brazilian journal of medical and biological research = Revista brasileira de pesquisas medicas e biologicas., 2022. [DOI | PubMed]
- H Song, L Liu, Z Song, Y Ren, C Li, J Huo. P4HA3 is Epigenetically Activated by Slug in Gastric Cancer and its Deregulation is Associated With Enhanced Metastasis and Poor Survival.. Technology in cancer research & treatment., 2018. [DOI | PubMed]
- T Wang, W Wen, H Liu, J Zhang, X Zhang, Y Wang. Development and Validation of a Novel Prognosis Prediction Model for Patients With Stomach Adenocarcinoma.. Frontiers in medicine., 2021. [DOI | PubMed]
- A Atkinson, A Renziehausen, H Wang, C Lo Nigro, L Lattanzio, M Merlano. Collagen Prolyl Hydroxylases Are Bifunctional Growth Regulators in Melanoma.. The Journal of investigative dermatology., 2019. [DOI | PubMed]
- M Oliva, K Demanelis, Y Lu, M Chernoff, F Jasmine, H Ahsan. DNA methylation QTL mapping across diverse human tissues provides molecular links between genetic variation and complex traits.. Nature genetics., 2023. [DOI | PubMed]
- V Fedyuk, N Erez, N Furth, O Beresh, E Andreishcheva, A Shinde. Multiplexed, single-molecule, epigenetic analysis of plasma-isolated nucleosomes for cancer diagnostics.. Nature biotechnology., 2023. [DOI | PubMed]
- F. Lyko. The DNA methyltransferase family: a versatile toolkit for epigenetic regulation.. Nature reviews Genetics., 2018. [DOI | PubMed]
- J Yang, P Antin, G Berx, C Blanpain, T Brabletz, M Bronner. Guidelines and definitions for research on epithelial-mesenchymal transition.. Nature reviews Molecular cell biology., 2020. [DOI | PubMed]
- AW Lambert, RA Weinberg. Linking EMT programmes to normal and neoplastic epithelial stem cells.. Nature reviews Cancer., 2021. [DOI | PubMed]
- DC Singleton, A Macann, WR Wilson. Therapeutic targeting of the hypoxic tumour microenvironment.. Nature reviews Clinical oncology., 2021. [DOI | PubMed]
- I Kaymak, KS Williams, JR Cantor, RG Jones. Immunometabolic Interplay in the Tumor Microenvironment.. Cancer cell., 2021. [DOI | PubMed]
- K Yoshihara, M Shahmoradgoli, E Martinez, R Vegesna, H Kim, W Torres-Garcia. Inferring tumour purity and stromal and immune cell admixture from expression data.. Nature communications., 2013. [DOI | PubMed]
- Z Zhang, H Zhu, X Wang, S Lin, C Ruan, Q Wang. A novel basement membrane-related gene signature for prognosis of lung adenocarcinomas.. Computers in biology and medicine., 2023. [DOI | PubMed]
- B Ru, CN Wong, Y Tong, JY Zhong, SSW Zhong, WC Wu. TISIDB: an integrated repository portal for tumor-immune system interactions.. Bioinformatics., 2019. [DOI | PubMed]
- SS Onkar, NM Carleton, PC Lucas, TC Bruno, AV Lee, DAA Vignali. The Great Immune Escape: Understanding the Divergent Immune Response in Breast Cancer Subtypes.. Cancer discovery., 2023. [DOI | PubMed]
- HK Angell, D Bruni, JC Barrett, R Herbst, J Galon. The Immunoscore: Colon Cancer and Beyond.. Clinical cancer research: an official journal of the American Association for Cancer Research., 2020. [DOI | PubMed]
- SJ O’Keefe. Diet, microorganisms and their metabolites, and colon cancer.. Nature reviews Gastroenterology & hepatology., 2016. [DOI | PubMed]
- G Kroemer, C Galassi, L Zitvogel, L Galluzzi. Immunogenic cell stress and death.. Nature immunology., 2022. [DOI | PubMed]
- H Zhou, J Zou, C Shao, A Zhou, J Yu, S Chen. Prolyl 4-hydroxylase subunit alpha 3 facilitates human colon cancer growth and metastasis through the TGF-beta/Smad signaling pathway.. Pathology, research and practice., 2022. [DOI | PubMed]
- GR Steinberg, DG Hardie. New insights into activation and function of the AMPK.. Nature reviews Molecular cell biology., 2022. [DOI | PubMed]
- F Nakasuka, S Tabata, T Sakamoto, A Hirayama, H Ebi, T Yamada. TGF-beta-dependent reprogramming of amino acid metabolism induces epithelial-mesenchymal transition in non-small cell lung cancers.. Communications biology., 2021. [DOI | PubMed]
- TE Keenan, SM Tolaney. Role of Immunotherapy in Triple-Negative Breast Cancer.. Journal of the National Comprehensive Cancer Network: JNCCN., 2020. [DOI | PubMed]
- X Chen, E Song. The theory of tumor ecosystem.. Cancer communications (London, England)., 2022. [DOI | PubMed]
- J Zhang, D Huang, PE Saw, E Song. Turning cold tumors hot: from molecular mechanisms to clinical applications.. Trends in immunology., 2022. [DOI | PubMed]
- Y Kieffer, HR Hocine, G Gentric, F Pelon, C Bernard, B Bourachot. Single-Cell Analysis Reveals Fibroblast Clusters Linked to Immunotherapy Resistance in Cancer.. Cancer discovery., 2020. [DOI | PubMed]
- P Errarte, G Larrinaga, JI Lopez. The role of cancer-associated fibroblasts in renal cell carcinoma. An example of tumor modulation through tumor/non-tumor cell interactions.. Journal of advanced research., 2020. [DOI | PubMed]
- S Su, J Chen, H Yao, J Liu, S Yu, L Lao. CD10(+)GPR77(+) Cancer-Associated Fibroblasts Promote Cancer Formation and Chemoresistance by Sustaining Cancer Stemness.. Cell., 2018. [DOI | PubMed]
- W Zeng, L Xiong, W Wu, S Li, J Liu, L Yang. CCL18 signaling from tumor-associated macrophages activates fibroblasts to adopt a chemoresistance-inducing phenotype.. Oncogene., 2023. [DOI | PubMed]
- T Li, J Fu, Z Zeng, D Cohen, J Li, Q Chen. TIMER2.0 for analysis of tumor-infiltrating immune cells.. Nucleic acids research., 2020. [DOI | PubMed]
- GT Consortium. The Genotype-Tissue Expression (GTEx) project.. Nature genetics., 2013. [DOI | PubMed]
- Z Tang, B Kang, C Li, T Chen, Z Zhang. GEPIA2: an enhanced web server for large-scale expression profiling and interactive analysis.. Nucleic acids research., 2019. [DOI | PubMed]
- E Cerami, J Gao, U Dogrusoz, BE Gross, SO Sumer, BA Aksoy. The cBio cancer genomics portal: an open platform for exploring multidimensional cancer genomics data.. Cancer discovery., 2012. [DOI | PubMed]
- CJ Liu, FF Hu, MX Xia, L Han, Q Zhang, AY Guo. GSCALite: a web server for gene set cancer analysis.. Bioinformatics., 2018. [DOI | PubMed]
- M Ramos, L Geistlinger, S Oh, L Schiffer, R Azhar, H Kodali. Multiomic Integration of Public Oncology Databases in Bioconductor.. JCO clinical cancer informatics., 2020. [DOI | PubMed]
- A Subramanian, P Tamayo, VK Mootha, S Mukherjee, BL Ebert, MA Gillette. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles.. Proceedings of the National Academy of Sciences of the United States of America., 2005. [DOI | PubMed]